

Hearing - Survival & Society
Our normal activities require hearing, yet we take hearing for granted. This is largely because the ear does its job so well that we do not pay attention to it. Hearing is the only sensory system that allows us to know what is going on everywhere in our environment - we don’t have to be looking at the twig that is snapping to know there is something behind us in the dark. This ability imparts tremendous survival advantages for all animals. In addition, human social structures rely on speech communication which requires the sensitive, rapid processing of acoustic energy that the normal inner ear provides.
A Brief History of Hearing - From Early Anatomy to Active Hearing
Like other sensory organs that allow mankind to experience the environment, the ear has long held a special fascination for scientists. In mammals, the inner workings of the ear are encased in the hardest bone of the body. It contains the smallest bones, the smallest muscles, and the smallest, yet one of the most elegant organs of the body, the cochlea (part of the inner ear). Progress in understanding the structure and function of this tiny, relatively inaccessible organ has been slow and the milestones broadly spaced. By the 15th century, the presence of the ear drum and two of the three bones of the middle ear had been noted. Almost 300 years would pass before Domenico Cotugno would find that, in contrast to the air filled middle ear, the inner ear is fluid-filled. A century later Ernst Reissner described the presence of two distinct fluid compartments in the cochlea. Improvements in microscopic methods during the 19th century led to Alfonso Corti’s painstaking description of the cells comprising the sensory receptor organ of the inner ear.
Between 1877 and 1900 three machines were invented that greatly enhanced the role that hearing would play in our daily lives. Alexander Graham Bell gave us the telephone, Thomas Edison the phonograph, Nikola Tesla and Guglielmo Marconi, the radio. The manufacture and marketing of these inventions required many years but the public was fascinated with the capturing and transmitting of voices, music, or other sounds over great time and distances. This new sound technology had an instant and monumental impact upon our cultural imaginations that initiated the revolution in communication technologies so central to the history of the 20th century. With the added importance of hearing to our daily lives came investigations during the late 19th and first half of the 20th century by scientists such as Hermann von Helmholtz and Georg von Békésy. Their work led to the concept of the ear as an elegant, but essentially passive device for converting the mechanical energy of sound into electrical signals to the brain. These studies culminated in von Békésy winning the 1961 Nobel Prize in Medicine and Physiology. Significant progress was made during the next two decades particularly with regard to cochlear fine structure and the cellular mechanisms for converting mechanical signals to changes in electrical potentials. But, the prevailing view of cochlear function continued to be that of a passive mechanical receptor for sound-evoked pressure changes in the cochlear fluids. However, as early as 1948 there were suggestions that an active mechanism might be necessary to explain the exquisite frequency resolving powers of the cochlea. Our understanding of how the ear works entered an exciting phase about 20 years ago when it was discovered that the inner ear actually makes sounds. The remainder of this chapter will provide a contemporary overview of how hearing works and describe what is known about the inner ear cells that make sounds and contribute to what is now considered active hearing. The first step in this overview requires a description of the mechanical energy we call sound.
Sound - Mechanical Vibrations - Pressure Waves
The sensory organs of the eye, ear, tongue and skin are each sensitive to specific forms of energy. The nose and tongue detect chemical energy, the eye detects light energy, the skin detects heat and mechanical energy. Sound is a form of mechanical energy. Mechanical forces can be steady, like the weight of this journal in your hand, or they can vibrate, like your car when it goes over speed bumps. Sound is generated by mechanical vibrations (such as a vibrating piano string). This sets up small oscillations of air molecules that in turn cause adjacent molecules to oscillate as the sound propagates away from its source. Sound is called a pressure wave because when the molecules of air come closer together the pressure increases (compression) as they mover further apart the pressure decreases (rarefaction). Since a pressure wave consists of a molecular disturbance, sound waves cannot travel through a vacuum. The velocity of sound in air is around 1,100 ft/sec which is why dividing the seconds between seeing lightning and hearing thunder by 5 gives a rough measure in miles of how far the lightning is from you. Sound waves travel fastest in solids, slower in liquids and slowest in air. Sound vibrations extend from a few cycles per second to millions of cycles per second. Human hearing is limited to a range of between 20 to 20,000 cycles per second. Sound at a vibration rate of greater than 20,000 cycles per second is called ultrasound. Other mammals can hear ultrasound, some such as whales approach 100,000 cycles per second. Physicians now use imaging techniques based on ultrasound (mechanical vibrations at millions of cycles per second) to examine the unborn child.
A sound is characterized by its frequency and intensity. The frequency of a sound contributes to its pitch and is measured by counting the number of cycles per second in the vibration. Intensity is a measure of loudness. If you have ever played a piano, you know where middle C is on the keyboard (see Figure 1). If the piano is properly tuned, middle C has a frequency of 256 cycles per second, high C (7 white keys to the right) has a frequency of 512 cycles per second. People with normal hearing can tell the difference between two sounds that differ by less 0.5%. In order to appreciate how small a difference in frequency this is you need only realize that middle C differs from C sharp (the black piano key immediately to its right of C) by more than 5%.
Figure 1.

The pressure waves of sound are represented by the advancing concentric lines radiating away from the vibrating source. Middle C has a frequency of 256 cycles per second, while upper C has a frequency of 512 cycles per second.
The intensity of a sound is a measure of its loudness and reflects how tightly packed the molecules of air become during the compression phase of a sound wave. The ear can detect sounds where the vibration of the air at the ear drum is less than the diameter of a hydrogen molecule. The ear has the ability to discriminate intensities over a 100,000 fold difference in energy. Still louder sounds can cause pain and damage the structures of the inner ear.
The task of all hearing organs is to analyze environmental sounds and transmit the results of that analysis to the brain. The brain interprets the ear’s analysis. All sensory organs have specialized sensory cells that convert an environmental signal into electrical energy. The change in electrical energy is then converted to a type a digital code that is transmitted to the brain. The human auditory system performs an analysis of sound entering the ear prior to the conversion to the neural code. The inner ear first determines how much energy is contained at the different frequencies that make up a specific sound. The cochlea is designed so that it is most sensitive to a specific frequency (say middle C) at one location and most sensitive to another frequency (say high C) at another. These different locations then transmit information to the brain. If the brain receives an increase in activity from the middle C location it then knows that the original sound contained energy at that frequency. This “mapping” of frequency information is just one of several strategies that the ear uses to code incoming information. The frequency analysis of environmental sounds begins in the external ear.
Getting Sound to the Inner Ear, the Analysis of Sound Begins
Millions of years ago marine animals had hearing organs that allowed them to detect sounds in the water. When land dwelling animals evolved they now had to detect environmental sounds that traveled through the air. This created a special challenge because their inner ears continued to be fluid filled. When sound passes from one media to another (as, in this example, from air to water) some energy is reflected by the surface and does not pass to the new media. In order to reduce these reflections and maximize the transfer of sound energy from the air filled environment to the fluid filled inner ear, land animals evolved external ears as sound collectors and middle ears as mechanical force amplifiers. It is fascinating that the tiny bones in the middle ear appear to have evolved from gills that were no longer needed.
The outer portion of the external ear reflects sound towards the ear canal (see Figure 2). Once in the ear canal, the pressure waves are aligned so they strike the ear drum at right angles. The reflection of sounds of different frequency is not the same and as a result the relative amplitude of some frequencies is greater than others. The result is that the relative amplitude of different frequencies at the ear drum differs, even if sound begins at the same intensity for all frequencies. Modification of the original sound by the external ear is a type of analysis that your brain learns to interpret. The frequency composition of familiar sounds aids your auditory system in determining where a sound is coming from. You can perform a simple experiment to appreciate the “frequency analysis” performed by the external ear. Cupping your hands over your ears and bending the top of your ears down changes the energy of different frequencies at your ear drum. Ask a friend to snap their fingers or clap their hands behind you before and after bending your ears. The sound will appear to come from different locations, particularly if the sound is coming from above or below your head, because your brain attempts to analyze the ear’s input to the brain based on the normal frequency pattern.
Figure 2.

Pressure waves from the speaker pass through the air to the external ear which collects the sound and passes it to the ear drum. The middle ear consists of the ear drum, the middle ear bones, and the membrane over the oval window at the entrance to the inner ear. The cochlea of the inner ear is named with the Greek word for “snail” because of its spiral shape. Did you know that the left cochlea spirals to the left and the right cochlea spirals to the right?
The middle ear bones conduct sound from the ear drum to the fluids of the inner ear. The ear drum is bigger than the oval window. The decrease in the area of these two membranes leads to an increase in pressure (pressure is equal to force divided by area and as the area gets smaller the pressure increases). The middle ear bones act as mechanical levers and further increase the pressure of the sound at the entrance to the cochlea. All of this is necessary to maximize the sound energy that gets to the fluids of the inner ear. There is a tube (called the eustachian tube) that connects the middle ear to the nose. It’s purpose is to allow the air pressure in middle ear to be equal to the air pressure in the environment. The pressure balance allows the ear drum to vibrate freely. Sometimes when you have a cold the tube is blocked and the middle ear pressure can not be balanced. You may have experienced the discomfort and even pain that can result if you are rapidly changing altitude (as when an airplane is landing). The freedom of movement of the middle ear bones can be reduced by certain diseases which leads to hearing loss. Any problem in the outer or middle ear that leads to a reduction of the sound energy entering the inner ear leads to what is called a conductive hearing loss. Many of these problems can be corrected either though medicine or surgery and contrast with the long term hearing problems that arise from damage to the structures in the inner ear.
The Inner Ear - A Closed Shop with a Division of Labor
The inner ear contains the sensory systems of balance and hearing. Its location is close to the center of the skull and it is encased in the hardest bones in the body which make it one of the best protected sensory systems. This protection reflects the importance of the hearing and balance for survival. The organs of balance are much older than those of hearing and evolved with the earliest multicellular organisms. All vertebrate balance organs are similar in structure and function. The organ of hearing evolved from one of the balance organs and this heritage is retained when it buds off from the balance organ early in fetal development. The basic structure of the human inner ear is present in the fetus at 6 months and hearing begins before birth. The auditory portion of the inner ear of mammals differs structurally from that of birds, reptiles and fish but its function in all animals is the same - to tell the brain how much energy is contained in an environmental sound and at what frequencies that energy is located.
The Inner Ear Battery - One Group of Cells Powers Another Group
The inner ear is divided into two fluid filled chambers - one inside the other. Figure 3 illustrates the basic organization of both the organs of hearing and balance. The fluid in the two chambers differs on the basis of the kind of salt that each contains. The fluid in the outer or bony chamber is filled with a sodium salt solution (called perilymph) that resembles the salt composition in the blood or the fluids found in the brain. The inner or membranous chamber is filled with a potassium salt solution (endolymph) that resembles the fluid that is normally found inside the cells of the body. Specialized cells that line parts of the membranous chamber and “pump” potassium into the membranous chamber maintain the difference in concentration between the two chambers. The difference in the chemical composition of these two fluids provides chemical energy (like a battery) that powers the activities of the sensory cells.
Figure 3.
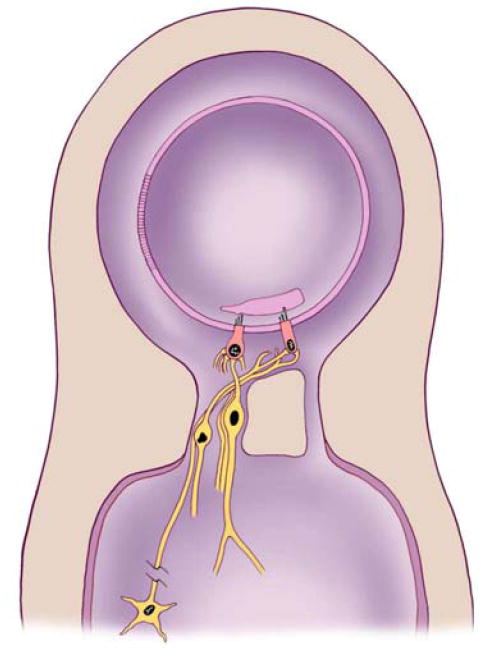
Simplified diagram showing the organization of inner ear organs of hearing and balance. The inner ear contains two fluid chambers, a membranous and a bony chamber. The membranous chamber is filled with endolymph while the bony chamber is filled with perilymph. The wall of the membranous chamber is made up of many cells that are so tightly joined together that they prevent the two fluids from mixing. The sensory epithelium makes up only a small portion of the wall of the membranous chamber and contains sensory receptor cells and surrounding supporting cells (supporting cells are not shown in the drawing).
This division of labor is unique to the inner ear because the function of the principal cells relies on chemical energy provided by other cells. In virtually all other systems, whether it is heart muscles, the brain, or the retina of the eye, the principal cells must combine nutrients and oxygen to produce the energy they use to perform their functions. In the inner ear, metabolic processes are performed by an organ called the stria vascularis located a half a millimeter from the hearing organ. The stria vascularis is essentially a battery whose electrical current powers hearing. It is powerful enough that if its power could be harnessed it could used instead of batteries for hearing aids.
Small Size Means Greater Mechanical Sensitivity
A sensory epithelium makes up a small portion of the wall of the membranous chamber. It is made up of the sensory cells together with “supporting” cells that help to support the function of the sensory cells both structurally and functionally. Some of the supporting cells may pump potassium into the endolymph and others will contribute to making substances that are connected to the ends of the sensory cells that are exposed to the endolymph. The inner ear sensory epithelium are among the smallest organs in the body, containing less than 20,000 sensory cells. Compare this with the million photoreceptors in the eye. The inner ear organs must be small because any increase in their size would increase their mass. An increase in mass would increase the mechanical force that would be required to make them vibrate. Any increase in the driving force would represent a decrease in the sensitivity of the system (a hearing loss). The small number of cells in the hearing organ means that the loss of even a small number affects hearing. While millions of photoreceptors in the retina improve visual resolution, millions of hair cells would decrease the sensitivity of the hair cell organs. Loss of a 1000 photoreceptors is hardly noticed - loss of a 1000 sensory cells in the ear leads to hearing loss. The mass of the cochlear sensory epithelium is further reduced because it has only a small number of blood vessels. The number of blood vessels required by the organ of Corti is reduced by a unique system for converting the metabolic energy from sugar and oxygen in the blood into an electrical potential. The indirect method for supplying the metabolic demands of the organ of Corti is adaptive because the auditory system is sensitive enough to “hear” the vibrations associated with blood moving through blood vessels. It is fortunate they are located away from the organ of Corti.
Hair Cells Convert Mechanical Energy to Electrical Energy
Hair cells are the sensory receptor cells of hearing and balance. They are as important for the senses of the inner ear as chemoreceptors are for taste and smell; as photoreceptors are for vision; and as the mechanoreceptors of the skin, muscles and joints are for touch and body sense. Hair cells are specialized mechanoreceptors that convert the mechanical stimuli associated with hearing and balance into neural information for transmission to the brain. The conversion of one type of energy to another is called transduction and hair cells are mechanotransducers.
Sensory hair cells are not in any way related to body hair, they are called hair cells because of their appearance under the microscope. They have a collection of some 100 thin cellular projections at one end of the cell. These projections (stereocilia) are similar to smaller projections (microvilli) associated with the surface of cells that line the gut or upper respiratory pathways. The stereocilia are arranged in rows that increase in length towards one side of the cell. Individual stereocilia are long and packed with a stiff filament-like molecule called “actin”. The actin serves as a type of skeleton (called a cytoskeleton because it is inside the cell). A stereocilia bundle is located at the “apical” end of the cell so that the cells appear to have hair in photomicrographs, hence the name hair cell. The hair style varies between the different hearing and balance organs. Hearing hair cells have something like a 50s crewcut while some balance hair cells possess a style that borders on punk rock. The different hair styles reflect differences in function between the hair cell organs.
Synapses - Communication to and from the Brain
Hair cells have synapses located at the end of the cell opposite the stereocilia bundle. Synapses are structures that permit communication between neuronal cells. One side of the synapse is presynaptic and the other postsynaptic. A chemical known as a “neurotransmitter” is secreted from the presynaptic cell and changes the membrane potential of the postsynaptic cell. There are two types of synapses associated with hair cells. “Afferent” synapses convey information into the central nervous system by exciting “action potentials” in the afferent nerve fibers that enter the brain. Action potentials are essentially pulses that travel down the fiber and carry information in a type of digital code. “Efferent” synapses modulate the membrane potential of the hair cell in response to neurotransmitter release from their presynaptic element which is the terminal of a nerve fiber that originates deep in the brainstem. The neural signals from the brain conveyed by these efferent fibers may be viewed as having the ability to change the “gain” (amplification, similar to changing the volume control on a radio) of the hair cells they innervate. The brain cannot change the gain of chemoreceptor or photoreceptor sensory cells directly because, in contrast to hair cells, they are not innervated by nerve fibers from the central nervous system.
Stereocilia Bundle Organization and Mechanotransduction
The sensory role of hair cells is accomplished by activating the auditory or vestibular nerve fibers that carry information into the brain. Neurotransmitter release at an afferent synapse is regulated by changes in the membrane potential of the hair cell in response to bending its stereocilia bundle. The bending of the bundle is a mechanical event which is why hair cells are mechanoreceptors (photoreceptors and chemoreceptors change their membrane potential in response to light and chemicals respectively, the change in the membrane potential of a sensory receptor cell in response to an appropriate stimulus is called a receptor potential). The stereocilia of each hair cell are arranged in a precise geometry. This arrangement is asymmetrical and polarized because the stereocilia are arranged in rows of short, intermediate and tall stereocilia. There is a stepwise progression from the shortest row to the tallest row that gives the bundle the appearance of a pipe organ. The bundle’s organization from short to tall rows is related to the functional consequences of bending the bundle on the cell’s membrane potential. Bending the bundle in the direction of the tallest row leads to an increase in neural activity to the brain. Bending the bundle in the opposite direction leads to a decrease in activity. Each hair cell therefore codes the direction and degree of stereocilia bundle bending by either increasing or decreasing the firing rate of the postsynaptic afferent fiber in proportion to the magnitude of the bend.
The Organs of Hearing and Balance
Inner ear sensory organs differ in the way their hair cell stereocilia bundles are mechanically bent. The hair cells in each organ are grouped in one of three types of sensory epithelium. The maculae and the cristae are the sensory epithelium of the vestibular system (balance) and the organ of Corti is the sensory epithelium of the cochlea. There are two maculae (the saccule and the utricle), three cristae and one organ of Corti on each side of the head. The hair cell stereocilia of each macula are linked to an overlying structure whose movement causes them to bend. Tilting your head or slamming on your car’s brakes cause the overlying mass in the maculae to move relative to the hair cells, bending the stereocilia bundle and activating the afferent nerve fibers connected to the hair cells. Your brain interprets the incoming information and tells the muscles of the body what to do without your awareness. The stereocilia in a cristae are bent in response to movement of the fluid in a semicircular canal (see Figure 4). The fluids are set in motion by rotation about the axis of the canal. For instance, when watching a tennis match you rotate your head to follow the ball. This causes the fluid in the “horizontal canal” to move relative to the crista and the nerve fibers are stimulated. The end result is that the muscles of your eyes move (again, without your conscious attention) so as to stabilize the visual field. While the hair cells in the vestibular end organs remain fixed relative to the bony cavity in which they are located, the hair cells in the organ of Corti move (or vibrate) in response to sound. Organ of Corti hair cell stereocilia insert into an overlying structure (the tectorial membrane) which vibrates less than the hair cells, and the differential movements bend the stereocilia bundle. This arrangement can vibrate at frequencies of between 20 cycles/sec to 20,000 cycles/sec for humans.
Figure 4.
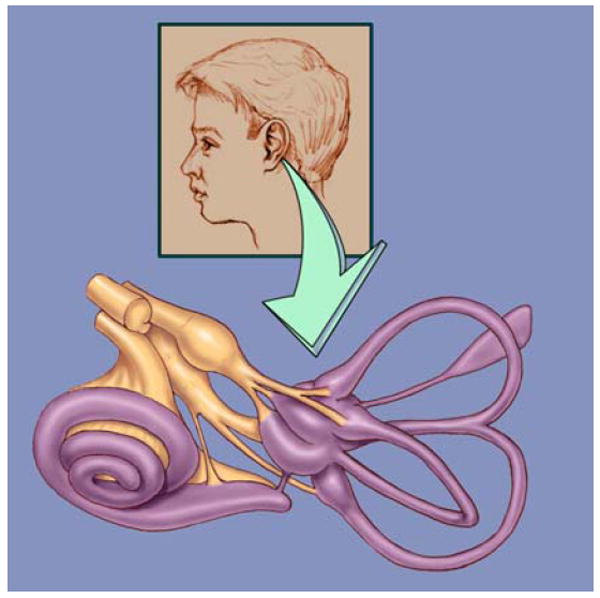
A drawing of the bony chambers of the inner ear. The bony chambers (purple) of the inner ear form precise geometric shapes. There are five sensory epithelia in the balance portion of the inner ear (one for each of the three semicircular canals and two in a chamber where the three canals come together). The nerve fibers (brown) can be seen on their way to the hair cells in the sensory epithelia. The nerve is made up of the neuronal projections that connect the hair cells with the brain and is called the eighth nerve because it is one of 12 nerves that come off the brain in the skull. The spiral shaped cochlea originates from one of the balance organs and contains the sensory epithelium for hearing.
The organ of Corti -The Temple of Hearing
The hearing organ in mammals is a spiraling structure called the “cochlea” from the Greek word for snail. It spirals out from the saccule (one of the balance organs). There are 2½ turns in the human cochlea and if you were to unwind the cochlea it would stretch to nearly an inch in length. By winding itself in a spiral, the organ takes up less space.
The sensory epithelium of the inner ear is called the organ of Corti after the Italian scientist who first described it. Its orderly rows of outer hair cells is unique among the organs of the body. Figure 5 shows a short section of the organ of Corti as it spirals in the cochlea. The organ of Corti is larger and the basilar membrane on which it sits is longer as it gets further away from the base of the cochlea. This difference in size is consistent with the fact that different frequencies of sound result in greater vibrations of the organ of Corti depending on where along the length of the cochlea you are measuring. The shorter, smaller structures near the base of the cochlea respond best to high frequencies, while the longer, larger structures near the top of cochlea respond best to low frequencies. This is similar to the organization of a piano where the longer, larger strings produce the lower frequency sounds.
Figure 5.

A drawing of the organ of Corti. The wide base of the cochlea from which this segment comes is towards the bottom of the page. The central axis of the spiraling cochlea is to the left of the drawing. Eighth nerve fibers pass through a bony shelf on their way to the hair cells (orange). The organ of Corti is made up of hair cells and supporting cells (purple and blue, respectively) that sit on a flexible basilar membrane which is anchored to the bony shelf on the left and a ligament (not shown) on the right. A single flask shaped inner hair cell is shown on the left and three rows of cylindrically shaped outer hair cells are seen on the right. The tips of the outer hair cell stereocilia are imbedded in a gelatinous mass called the tectorial membrane which lies on top of the organ of Corti and is secreted from cells (not shown) on the left.
When sound is transmitted to the inner ear the organ of Corti begins to vibrate up and down. Since the basilar membrane is attached to bone and ligament at its two ends, the area of maximal vibration is near the third (furthest right) row of outer hair cells. The overlying tectorial membrane is not as flexible so the stereocilia are bent as the organ of Corti moves up and down against it. The electrical potential inside the hair cells changes as the stereocilia are bent.
In no other organ in the body is it as easy to see the precise organization of the principal cells. The supporting cells of the organ of Corti are not found immediately adjacent to the outer hair cells so that for most of the length of these cylindrically shaped cells are surrounded by a relatively large fluid filled space (Figure 6 provides a view of a row of outer hair cells). The popular press has long appreciated the photogenic appearance of the organ of Corti. Pictures obtained with electron microscopes are routinely published showing the organ of Corti’s colonnade appearance. The three rows of columns are the outer hair cells and the organ is beautiful for the same reason that ancient Greek temples are beautiful. The space around the cells allows you to see their organization and appreciate their role in supporting the rest of the structure. In no other organ in the body do you find large fluid filled spaces around the principal cells. Neurons in the brain are surrounded by supporting cells. The cells in the muscles of the heart are close to one another. We now know that the spaces around the outer hair cell allow the cells to change their length during hearing.
Figure 6.

View of portion of the third row of outer hair cells. The view is what would be seen if you were looking towards the central axis of the cochlea and the most lateral set of supporting cells were removed. The organ of Corti is spiraling towards the top of the cochlea to the right. Stereocilia are on the top and radial fibers of the basilar membrane are seen on the bottom. The outer hair cells sit in a cup formed by a supporting cell. The supporting cells send out a narrow filament that angles towards the base of the cochlea. This unique structural organization means that the supporting cells touch the outer hair cells only at their top and bottom.
Inner and Outer Hair Cells
The presence of two types of hair cells, the inner and outer hair cells, in the organ of Corti was appreciated nearly a 100 years ago but the function of the two has only become clear in the last 20 years. The first clue that they might play different roles in hearing came about 40 years ago as the result of a painstaking anatomical investigation which revealed that most of the nerve fibers that carry information to the brain contact only the inner hair cells. This meant that most the information about the acoustic world reached the brain via the inner hair cells. What then was the role of outer hair cells which are over three times more numerous? The mystery was compounded by the discovery that neural fibers originating from neurons deep in the brain, which send information back to the hair cells, only touch outer hair cells. It was later determined that outer hair cell stereocilia are firmly embedded in the overlying tectorial membrane while inner hair cell sterocilia make only a tenuous connection. The outer hair cells are located near the center of the basilar membrane where vibrations will be greatest while the basilar membrane is anchored under the inner hair cells (see Figure 5). These observations suggest that the movement of stereocilia and the resulting modulation of their ionic currents is likely to be greater for outer hair cells than for inner hair cells. Several studies that had examined the inner ears of deaf people shortly after they died demonstrated that outer hair cells were required for hearing. It was clear that the inner hair cells served to transmit information to the brain but the role of the outer hair cells remained a mystery.
The mechanical vibrations of the organ of Corti had been analyzed by engineers since the 1940s. Their analysis was able to explain the frequency selectivity originally measured by von Bekesy in cochlea obtained from cadaver ears. It was known at the time that the measured frequency selectivity and the frequency selectivity computed from the engineering analysis did not approach the frequency selectivity of the human hearing or the frequency selectivity that could be measured from individual nerve fibers. Shortly after WW2 an American astrophysicist who had worked on radar during the war suggested that the frequency selectivity of the cochlea could be enhanced if a source of mechanical energy were present in the cochlea. His suggestion was largely ignored until several engineering groups in the late 70s rediscovered the potential benefits of this hypothetical energy source. They were forced to consider the possible existence of this “cochlear amplifier” when improved measures from living (as opposed to dead) ears revealed that the mechanical frequency selectivity in the living ear began to approach that of human hearing. The concept that a source of mechanical energy exists in the cochlea appeared validated when in the late 70s it was discovered that sound is produced by the inner ear. These sounds can be measured by placing a sensitive microphone in the ear canal. They were called otoacoustic emissions and they are now routinely measured in the clinic to assess hearing. Their discovery was amazing for sensory physiology because it was equivalent to finding that light comes out of the eye (which has never been observed). Within five years it was discovered that the outer hair cell could be made to elongate and shorten by electrical stimulation. The function of the outer hair cell in hearing is now perceived as that of a cochlear amplifier that refines the sensitivity and frequency selectivity of the mechanical vibrations of the cochlea.
Outer Hair Cell Electromotility
Outer hair cells have a cylindrical shape. Their diameter is about 9 micrometers which is slightly larger than the diameter of a red blood cell. Their apical end is capped with a rigid flat plate into which the stereocilia are embedded and their synaptic end is a hemisphere (see Figure 7). Each of these three regions (flat apex, middle cylinder and hemispheric base) has a specific function. The stereocilia at the top of the cell are responsible for converting the mechanical energy of sound into electrical energy. Synaptic structures are found at the base of the hair cell and they are responsible for converting electrical energy into chemical energy by modulating the release of neurotransmitters that activate the 8th nerve fibers contacting the cell. The top and the bottom of the outer hair cell perform functions that are common to all hair cells. The elongated cylindrical portion of the outer hair cell is where electrical energy is converted into mechanical energy. This function is unique to the outer hair cell. No other hair cell (nor any other kind of cell) is able to change its length at acoustic frequencies in response to electrical stimulation. These length changes can be greater than 1% of the cell’s original length if the electrical stimulation is large. The physical mechanism responsible for this electromotility is not known but there is recent evidence that cellular membranes can move in response to high frequency electrical stimuli. It is possible that all membranes generate mechanical force but that the outer hair cell is specialized to generate length changes. It should come as no surprise that there are structural features that are unique to the outer hair cell which facilitate its ability to undergo rapid length changes.
Figure 7.

Drawing of outer hair cells showing the effect of different internal pressures. The outer hair cell is divided in three parts. The top part is capped with a flat plate into which the stereocilia are inserted. The base of the cell is hemispheric. It contains the cell nucleus (round ball) and synaptic structures (not shown) for communicating with the central nervous system. The middle part of the cell is cylindrical in shape. The shape of the outer hair cell is maintained by a pressurized fluid core that pushes against an elastic wall. The wall is reinforced by additional layers of cytoskeletal material and membranes (shown in drawing by concentric cylinders). If the cell is at a normal pressure as on the left it will shorten and become fatter (middle) when there is an increase in intracellular fluid. When fluid is lost the cell elongates and the sides of the cell collapse (right) from the loss of pressure.
The Outer Hair Cell is Pressurized
Most of the cells in your body have an internal “skeleton” that maintains the shape of the cell. Several types of relatively stiff large molecules normally make up the so called cytoskeleton. These large molecules join together to form a scaffold which spans the inside of a cell. The stereocilia of all hair cells are stiff because of an abundance of one of these cytoskeletal proteins. All hair cells except the outer hair cell also have the cell spanning cytoskeleton throughout their cell body. Because such an internal skeleton would reduce electromotility, nature appears to have removed the central cytoskeleton from the cylindrical portion of the outer hair cell in order to improve the cell’s flexibility. The outer hair cell must be more than flexible, it must also be strong enough to transmit force to the rest of the organ of Corti. As a result the outer hair cell is pressurized. Pressurized cells are common in the plant kingdom but rarely found in cells of the animal kingdom. Plant cells, such as those found at the base of a tree, are highly pressurized. This allows the plant cells to hold the weight of the tree and still be flexible enough to bend and not shatter in a wind. You are familiar with inflatable tires which are also pressurized. You may remember the solid rubber tires that were on your little red wagon and how the wagon vibrated as you hit stones in the road. In contrast, your first bicycle didn’t vibrate as much because the air filled tires absorbed the bumps in the road. The pressurized fluid that fills the outer hair cell is only part of the story. Most of the cells in our body will not tolerate internal pressure because the membrane that encloses them has the strength of a soap bubble. A conventional pressurized cell will expand till it bursts. The outer hair cell has reinforced the membrane along the cylindrical part of the cell to prevent it from bursting and to maintain the cylindrical shape.
Outer hair cells are isolated from the organ of Corti and kept alive to record their electromotility while recording their movements on video. During these experiments the outer hair cells will collapse (like the cell on the right of Figure 7) when exposed to large amounts of aspirin (equivalent to what you would have in your blood after taking 8 aspirin tablets). Aspirin has long been known to cause a reversible hearing loss and more recently it has been shown to block otoacoustic emissions. The experiments show that aspirin causes both effects by a direct action on the outer hair cell.
Most of the reinforcement is provided by two cytoskeletal proteins that are located immediately below the cell’s membrane. The stiffer of these is oriented circumferentially. These molecules serve the same function as the steel threads in steel belted radial tires. It is interesting that they are not directly attached to the cell membrane but instead appear directly bonded to another membranous structure that is located immediately under them. The membranes of this stucture form what is equivalent to the inner tube of older tires. This “inner tube” lines the cylindrical portion of the cell and it is a structure that is found no where else in biology. It is called the subsurface cisterna and it is known to be essential for the full expression of electromotility. Manipulations that alter the subsurface cisterna reduce electromotility. The three layers that make up the side of the cylindrical portion of the outer hair cell are incredibly close together. The distance between the outside of the cell to the inside of the subsurface cisterna is much less than the wavelength of light. Up until recently the only way to see the layers was to use an electron microscope.
Active Hearing Improves Frequency Selectivity
The end result of having outer hair cell electromotility is that we are able to discriminate between sounds that are very close in frequency. The reason for this may be easily understood by considering a simple children’s swing. A playground swing is a mechanically tuned system that we are all familiar with. This fact is known to any parent who has pushed a child on a swing. The frequency of the system is determined by the length of the rope and the mass of the child. The tuning of the system is best appreciated when you attempt to push at a frequency other than that determined by the natural frequency of the swing. If you attempt to push at a different frequency you know that the system fights against you and you have to exert considerably more energy if you really want it to move at a different frequency.
A child sitting passively on a swing is an example of a passive in contrast to an active system. In order to understand what is meant by an active system you need only recall your own early experiences on a swing. After a short time you learned that you could make the swing to go higher and higher by pumping energy into the system through a combination of kicking your legs and tilting your upper body. Not only could you make the swing go very high but you probably experimented with kicking at a frequency other than the natural (passive) best frequency. When you did you found that you rapidly suppressed the magnitude of the swing. The new “active” system with you pumping had become effectively more narrowly tuned than the original system in which you just sat on the swing.
One way to imagine the cochlea is to envision a row of 3000 swings that have progressively longer ropes and heavier children as you move to the right. Each swing has a preferred frequency that is lower than the swing to the left. The parents behind each child are equivalent to sound vibrations and if the children do not pump the entire set of swings is equivalent to a dead cochlea with relatively poor frequency selectivity. If the children pump the set of swings is now equivalent to a living or active cochlea with greatly improved frequency selectivity.
The role of the outer hair cells in hearing is both sensory and mechanical. When the organ of Corti begins to vibrate in response to the incoming sound, each hair cell will sense the vibration through the bending of its stereocilia. The bending results in a change in the outer hair cell’s internal electrical potential which drives electromotility. If the resulting mechanical force is at the natural frequency of that portion of the cochlea then the magnitude of the vibration will increase. If the electomotile force is at a different frequency, the vibrations will decrease. The system now has greater sensitivity and frequency selectivity than when the outer hair cells are missing or damaged. The refined mechanical vibrations of the organ of Corti are transmitted to the inner hair cells which excite the 8th nerve fibers at their base and tell the central nervous system that there is energy at a specific frequency in the sound entering the ear. One consequence of having an active system is that oscillations can occur even when no energy is coming into the system from the outside. This happens in the cochlea and the resulting sound vibrations can be measured in the ear canal. These are called spontaneous otoacoustic emissions and are only observed in living ears. Measuring otoacoustic emissions has become an important diagnostic tool for determining if outer hair cells are working.
Regeneration - The Problems of Reorganizing and Rewiring
Hair cells in the organ of Corti are “born” during the first three months of fetal development. They resemble neurons because they must last a lifetime. Unlike the cells that make up our liver, heart and kidney, cochlear hair cells and neurons do not replace themselves. One reason that both hair cells and neurons do not replace themselves may lie in the number of contacts they have with other neurons. During development, neurons and hair cells make contact with many other neurons with which they communicate. The process that controls the formation of contacts appears to be both genetic and competitive. Many contacts are made, correct connections that benefit the brain survive and inappropriate connections disappear. If a new hair cell were to come into existence in an adult ear it would have to be rewired to the myriad of fibers that connect it with the brain. The precise organization of the organ of Corti poses yet another obstacle for a regenerating hair cell. The cells that might change into hair cells are located near the central axis of the cochlea, if one of these could be made to change into an outer hair cell it would have to migrate outwards past the inner hair cells, through all the supporting cells to find its correct place in the precise rows of outer hair cells. Even though outer hair cells change length when electrically stimulated, they cannot move in a manner that would allow them to make their way through the intervening supporting cells.
Several years ago it was reported that bird ears recover function after hair cell damage and this was followed by a report that new hair cells can appear in human balance organs. These reports triggered considerable optimism that regeneration might be possible in the adult organ of Corti, but so far there is no evidence for cochlear hair cell regeneration. The problem of hair cell regeneration is comparable to that of neural regeneration. The severe spinal cord injuries of former Governor Wallace and the actor Christopher Reeve have focused a great deal of attention on research into neural regeneration. One benefit of the research was that when Mr. Reeve sustained his injuries he was quickly treated with drugs that reduced the amount of neural damage. He is now regaining some sensation that might not have been possible 10 years ago. A similar strategy would be to investigate the use of similar drugs to protect hair cells when they are subjected to potentially damaging situations. Just as research continues on neural regeneration, research should continue on hair cell regeneration and perhaps someday we will be able to coax new hair cells to grow. Even if we are successful in achieving regrowth, considerable work will have to take place in learning how to promote correct rewiring of the hair cells with the central nervous system. The process could be more challenging than learning a foreign language as an adult.
The Central Hearing System - Where Sound Is Reconstructed
In all sensory systems an important part of the neural code is determined by what location of the sensory organ is stimulated. In the case of the eye, a spot of light falls on a few photoreceptors and they excite nerves that map onto a representation of the visual world in the brain. In the ear, the acoustic world is coded by a 1 dimensional representation of frequency. This frequency map then projects to the brain which performs the almost unbelievable task of reconstructing the original 3 dimensional acoustic world. There are actually parts of the brain that contain a true 3 dimensional representation of the outer world so that the sound of a twig snapping behind you excites nerve cells in one location while a twig snapping on your right excites nerve cells in another spatially precise location. The analysis of speech appears to take place in parts of the brain that are highly developed only in man. The amazing machinery that accomplishes the reconstruction of the acoustic world relies on the delicate structures of the inner ear that deconstruct the original sounds.
Figure 8.

A playground swing is a simple example of mechanical frequency selectivity with a passive system. The swing moves back and forth at a frequency that is determined by the length of the rope and the mass of the child. The parent can make the swing move at a different frequency by exerting considerably more effort than pushing at the natural frequency.
Figure 9.

Active tuning is achieved when the child pumps energy into the system. When the pumping is done at the natural frequency and at the correct time in the cycle the swing goes higher and tuning is improved. The improved tuning is best appreciated by attempting to pump at a frequency that is different than the natural frequency. In contrast with the parent pushing, no amount of pumping will make the swing move at the new frequency, in fact the magnitude of the swinging rapidly decreases. This ability to narrow the range of frequencies at which the swing will oscillate is equivalent to what the outer hair cell does in hearing.
Recommended Resources: Books, Articles and the Internet
Books
Pickles, J.O. (1989). An introduction to the physiology of hearing (2nd ed.). Academic Press. And Geisler, C.D. (1998). From Sound to Synapse : Physiology of the Mammalian Ear. Oxford University Press. These single-authored books provide excellent overviews of hearing mechanisms from the external ear to the brain. Both are easily read by someone with a good high school or early college background in biology.
Berlin C.I. (Ed.). (1996). Hair Cells and Hearing Aids. Singular Publishing Group, Inc. San Diego. This book contains contributions to a one day symposium with the same title. The level of the chapters range from intermediate to advanced and the book includes an instructive CD that demonstrates the role of outer hair cells in hearing.
Dallos, P., Popper, A.N., Fay, R.R. (Eds). (1996). The Springer Handbook of Auditory Research. The Cochlea. Springer, New York. This book provides up to date research reviews on the inner ear with contributions by many scientists. Information on the cochlea is more complete and more technical than the Pickles and Geisler books.
Articles
Liberman, C. (1997). Audiological Processing in Sensorineural Hearing Loss: What the Ear Tells the Brain. Hearing Loss, March/April, 9-13. This easy to read paper provides an excellent description of how auditory information is “coded” in the nerve fibers that leave the cochlea for the brain. Hearing Loss is published by Self Help for Hard of Hearing People, Inc. (SHHH), a national organization dedicated to improving hard of hearing people’s lives though education, advocacy and self help.
The Internet
The internet provides a great deal of information on hearing. There are a number of websites on hearing research and several support groups for the hearing impaired. The net, with its multimedia capabilities, is an exciting educational tool and a tremendous amount of information is available. As with any media which thrives on free expression, you must exercise careful judgment as to the accuracy of specific websites. Four URLs that will get you started are:
http://www.bcm.tmc.edu/oto/research/cochlea/index.html - Cochlear biophysics lab at the Department of Otolaryngology and Communicative Sciences at the Baylor College of Medicine.
http://www.bcm.tmc.edu/oto/othersd.html - The Department of Otolaryngology and Communicative Sciences at the Baylor College of Medicine. Otolaryngology Resources on the Internet
http://www.aro.org/ - The Association for Research in Otolaryngology. See the Virtual Library for resources around the world.
http://www.agbell.org/index.html/ Alexander Graham Bell Association for the Deaf established in 1890 to empower persons who are hearing impaired to function independently by promoting universal rights and optimal opportunities to learn to use, maintain, and improve all aspects of their verbal communications.
http://www.shhh.org/ Self Help for Hard of Hearing People, Inc.