

Abstract
Loss of hepatic tissue triggers a regenerative response in the whole organ. Under typical normal conditions, all hepatic cells (epithelial: hepatocytes and biliary epithelial cells; non-epithelial: stellate cells, macrophages and endothelial cells) undergo one to three rounds of replication to establish the original number of cells and restore organ size. The review summarizes the literature of regenerative patterns in situations in which proliferation of either hepatocytes or biliary epithelial cells is inhibited. The evidence strongly suggests that under these circumstances, hepatocytes or biliary epithelial cells can function as facultative stem cells for each other and replenish the inhibited cellular compartment by a process of trans-differentiation, involving complex signaling pathways. These pathways are activated under experimental conditions in rodents and in fulminant hepatitis associated with liver failure in humans. Mechanistic analysis of these pathways has implications for liver biology and for potential therapeutic modalities in human liver disease.
Keywords: Liver regeneration, Hepatocytes, Oval Cells, Stem Cells, Transdifferentiation
Introduction
Liver regeneration is a very well studied response to loss of hepatic tissue. Such loss may occur as a result of toxic injury, exposure to viruses, trauma or surgical resection. Since hepatocytes are the major functional cells of the liver, typically a regenerative response is triggered when there is loss of hepatocytes at a large scale.
Experimental studies of liver regeneration have utilized animal models, in which loss of hepatic tissue is induced by exposure to a toxin such as CCl4, or by surgical resection. The most popular model for studies is liver regeneration after 2/3 partial hepatectomy in rodents (Higgins G.M., 1931). Rat and mouse liver are composed of five lobes. Three of these lobes can be removed by a simple surgical procedure. The remaining two lobes increase in size via cell proliferation and grow to an aggregate size equivalent to that of the total of the original five lobes. This experimental approach is much preferred than the one mediated by exposure to toxins. The primary reasons for the popularity of the 2/3 partial hepatectomy (PHx) rodent model are that there is no tissue necrosis with accompanying infiltration by macrophages and other extrahepatic cells whose presence distorts analysis of the biochemistry of the regenerative process; the surgical resection is well tolerated; the residual two lobes are histologically intact; and the process can be well timed from a specific starting point.
There have been several recent reviews of liver regeneration induced by PHx (Michalopoulos and DeFrances, 1997, Michalopoulos and DeFrances, 2005, Michalopoulos, 2007, Fausto, 2000, Fausto et al., 2006). These reviews have provided details on mechanisms associated with initiation and termination of liver regeneration, including signaling pathways, growth factors and cytokines, cell cycle associated proteins, contributions of extracellular matrix, etc. The reader is referred to these reviews for detail presentations of these topics. The purpose of the present review is to conduct a critical examination of the alternative cellular and signaling pathways that are mobilized in the liver when the standard regenerative process is thwarted, as, for example, when hepatocytes are prevented from proliferating. The review will also address the contributions of mature hepatic cell populations, i.e. hepatocytes and biliary epithelial cells, which may assume the role of facultative stem cells and provide progenitor cells for one another, in cases in which the regenerative capacity of hepatocytes or biliary cell fails. We will review the evidence that the biliary compartment is the source of progenitor cells that trans-differentiate to hepatocytes when hepatocyte proliferation is inhibited and liver needs to regenerate. In a reverse fashion, we will also review the evidence that when the biliary compartment needs to repair biliary damage and is unable to do so, populations of specific hepatocytes may also undergo transdifferentiation and provide progenitor cells that contribute to the repair of the biliary epithelium. Finally, we will also conduct a critical analysis of studies suggesting that extra-hepatic tissue sites may also contribute to progenitor cells for hepatocytes.
I. Role of progenitor cells in standard liver regeneration after PHx
Earlier studies critically inquired as to whether liver regeneration after PHx was mediated by a small number of stem cells which generated all hepatocytes needed to replenish the lost tissue. The alternative view was that most of the existing mature hepatocytes and the other hepatic cell populations were all undergoing few waves of proliferation to provide enough cells required for the restoration of the hepatic mass. A critical early experiment was performed by Stocker et al, in which tritiated thymidine was continually administered after PHx. In rats of up to 16 months of age, the approach resulted in labeling of almost 99% of hepatocytes (Stocker and Heine, 1971). The authors interpreted this as evidence that restoration of the numbers of hepatocytes was mediated by proliferation of most of the existing hepatocytes at the time of PHx. If the opposite was the case, i.e. if proliferation was primarily a property of a small number of stem cells which exclusively restored the hepatocyte numbers, than at least 1/3 of the hepatocytes (the ones residual to the liver after PHx) would not have nuclei labeled by tritiated thymidine. The same authors also demonstrated that liver could regenerate even after 12 sequential hepatectomies performed in the same animal (Stocker et al., 1973) The evidence for the hepatocytic origin of hepatocytes in liver regeneration after PHx was also critically summarized by Fausto in a recent review(Fausto, 2004).
II. Proliferative capacity of mature hepatocytes
Most of the studies on this matter up until 1994 considered hepatocytes as fully differentiated cells of limited proliferative capacity. This was reinforced by the fact that hepatocytes in culture could not undergo more than one or two rounds of replication. Subsequent studies however have demonstrated that this is not the case. Studies by Rhim et al.(Rhim et al., 1994) showed that mouse hepatocytes could repopulate the entire liver when transplanted in the failing livers of mice in which urokinase was expressed under the albumin promoter in hepatocytes. The high expression of urokinase was causing hepatocyte degeneration and liver failure. Transplantation of normal hepatocytes in the liver of these mice prevented liver failure and resulted in complete repopulation of the liver. It was estimated that the repopulation required 12 hepatocyte doublings (Rhim et al., 1995). The findings were further confirmed by a similar but more powerful approach by Grompe et al. utilizing mice suffering from congenital tyrosinemia due to deficiency in fumarylacetoacetate hydrolase (FAH) (Overturf et al., 1997). Treatment of the mice with the chemical NCTB prevents liver failure and the FAH deficient mice reproduce normally. Removal of NTCB form the drinking water induces liver failure. When this is accompanied by infusion of normal hepatocytes (from mice transgenic for expression of beta galactosidase) the result was complete repopulation of the liver with FAH+LacZ+ hepatocytes. When the FAH+LacZ+ hepatocytes were isolated from the liver of the first generation of rescued mice, they were equally successful in repopulating the liver of a second generation of mice. This was repeated seriatim for 10 times and it was estimated from the mathematics of the model that one mouse hepatocyte was capable of repopulating 30 mouse livers! Also of interest was the finding that diploid and polyploid hepatocytes were equally capable of contributing to liver repopulation in this model (Overturf et al., 1999). Repopulation models are also available for rat liver. Retrorsine, a pyrrolizidine alkaloid, can be metabolized by hepatocytic CYP enzymes to active intermediates causing cross-linking of hepatocyte DNA and inhibiting hepatocyte proliferation after PHx. When normal hepatocytes are injected following hepatic resection in the retrorsine-treated animals, the injected normal hepatocytes colonize most of the liver and restore normal liver weight. The colonization is demonstrable by using Fisher 344 rats of two strains, one positive and one negative for expression of DPP IV enzyme. The expression of the enzyme can be demonstrated by simple histochemistry. The colonization of the liver in all above models involves only the hepatocytes. Biliary epithelial cells remain those of the recipient liver (Laconi et al., 2001). The capacity of hepatocytes to generate clones in culture has also been demonstrated. In suitable media, hepatocytes expand as clones under the influence of HGF and EGF (Block et al., 1996). Other studies showed that EGF and HGF increase expression of telomerase in hepatocytes in primary culture (Nozawa et al., 1999). The extensive proliferation of hepatocytes in cellular transplantation models has been considered to be a unique property of rodent hepatocytes. Normal mouse and rat tissues, including liver, do express telomerase (Yamaguchi et al., 1998), whereas human tissues do not (Hytiroglou and Theise, 2006). On the other hand, it was also shown recently that human hepatocytes are also capable of colonizing mouse livers almost as effectively as mouse hepatocytes (Azuma et al., 2007). Altogether, these findings suggest that hepatocytes have a capacity to proliferate which far exceeds stereotypes for most known epithelial cells. This proliferation is mediated by hepatocytes themselves, and not through stem cell populations.
III. Hepatic progenitor cells: “Oval Cells”, “Ductular Hepatocytes”
“Oval Cells” is a name given by E. Farber (Tatematsu et al., 1984) to a population of cells in the liver which appear after PHx when hepatocyte proliferation is suppressed. (The name of the cells derives from the shape of their nucleus, which tends to be oval, as compared to normal hepatocytes, which have in their majority perfectly round nuclei). In the most studied model, suppression of hepatocyte proliferation in the rat is done by administration of the chemical Acetylaminofluorene (AAF) (Evarts et al., 1989). Oval cells appear within a few days after PHx, expand in the periportal regions of the lobule and reach a peak at 7–9 days after PHx. The details of the kinetics of the appearance of these cells vary with specific protocols (Shiojiri et al., 1991, Sirica and Cihla, 1984, Paku et al., 2001, Grozdanov et al., 2006, Pi et al., 2005, Tatematsu et al., 1984, Sell, 1994). In mice, there is no comparable model but oval cells can be induced by administering a variety of chemicals such as dipin (Factor et al., 1994). Oval cells also appear following administration of toxic diets which cause destruction of hepatocytes, thus triggering a regenerative response, in which hepatocytes cannot fully participate because of the toxicity associated with the diet (Sells et al., 1979, Shinozuka et al., 1978). Extensive analysis of the histochemical and biochemical properties of the oval cells shows that they have gene expression patterns intermediate between hepatocytes and biliary cells (Evarts et al., 1996, Evarts et al., 1989). They are arranged in sheets or in glandular/ductular structures. These ductular structures are often lined by cells having a diversity of size, some of them approaching morphology of hepatocytes whereas other cells in the same gland have a more typical biliary appearance. Specific antibodies are available, however, which are unique to oval cells and do not react with biliary epithelium (Hixson et al., 1997, Yang et al., 1993). Oval cells also intensely express alpha fetoprotein (AFP), a protein produced by fetal hepatocytes (Sell, 1980, Sell, 1978). As long as hepatocyte proliferation is blocked (e.g. by AAF), oval cells will appear whenever there is massive loss of hepatocytes. The oval cell response is elicited whether the cause of hepatocyte loss is surgical resection (as in PHx) or chemical injury causing centrilobular damage (e.g. carbon tetrachloride). When the damage causing loss of hepatocytes is periportal, however, as with allyl alcohol, then there is only proliferation of portal ductules but no expansion into a true oval cell response (Petersen et al., 1998).
A similar cell type and overall histologic pattern has also been indentified in humans during acute liver failure. The term “ductular reaction” is often used to describe it (Demetris et al., 1996). The cells are intermediary in phenotype and histology between biliary epithelial cells and hepatocytes. This reaction occurs regardless of the cause of liver failure, and it is seen in chemical toxicity or viral hepatitis cases. The term “ductular hepatocytes” has been used to describe these cells and they are similar in properties and overall behavior to the oval cells seen in rodents (Fiel et al., 1997, Haque et al., 1996)
Classic studies by Evarts and Thorgeirsson utilized pulse labeling with tritiated thymidine to unambiguously demonstrate that oval cells gradually transform themselves into small basophilic hepatocytes which then become fully mature hepatocytes and replace the lost liver mass (Evarts et al., 1996, Evarts et al., 1989, Evarts et al., 1987). This pathway was shown in essence to be that of an SOS type, rescuing liver regeneration if hepatocytes failed to respond (Alison et al., 1997).
The origin of the oval cells has been much debated. A central point of the debate has been whether oval cells are of biliary origin or whether they derive from another not readily identifiable, hepatic cell type, which would function as a stem cell or a progenitor cell and generate the oval cells. Since there are no cell types which are easily identifiable in large number by routine microscopy that can potentially be candidates for the origin of oval cells, another scenario involves the possibility that oval cells are derived from extrahepatic sources which migrate to the liver as needed and transform into the oval cells. None of the above three scenarios are mutually exclusive.
The overwhelming evidence from the accumulated literature suggests that oval cells derive from the biliary compartment. That is considered to include both portal biliary ductules and the canals of Hering. The latter are extensions of the portal biliary ductules and extend fairly deeply into the lobule, coming into direct contact with hepatocytes and forming the first tributaries of the flow system that delivers bile beyond the hepatocyte canaliculi and into the main biliary tree (Theise et al., 1999, Roskams et al., 2004). The evidence for the biliary origin of the oval cells is as follows:
The predominant patterns of gene expression at the earliest stages of the oval cell expansion are overwhelmingly biliary. The histologic arrangement of the cells also follows a tubular/ductular pattern (Hayner et al., 1984, Sirica et al., 1990).
Pretreatment with the biliary toxin DAPM prior to the initiation of the oval cell protocol causes massive loss of biliary epithelium and also eliminates the oval cell response (Petersen et al., 1997).
Soon after the initiation of the AAF/PHx protocol in rats, there intense proliferation of portal biliary ductules (Bisgaard et al., 1996). This is the only site in which proliferation is seen.
In the same study, portal biliary ductules begin expressing hepatocyte-associated transcription factors, including C/EBP alpha and beta and HNF4 (Bisgaard et al., 1996). This finding is very important because it indicates that biliary cells begin to undergo massive gene expression reprogramming towards the hepatocytic phenotype.
Similarly to the above findings in rats following AAF/PHx, biliary ductules in humans with massive hepatocyte loss and acute liver failure also express hepatocyte-associated transcription factors (HNF4, HNF6) and hepatocyte markers such as HEPPAR protein (Limaye et al., 2008a).
The issue of the relative contribution of portal ductules versus canals of Hering is not a settled one. The paucity of oval cells in the AAF/Allyl alcohol protocol suggests that the canals of Hering may be more important, as they are predominantly affected by the side-effect of the loss of the periportal hepatocytes and the associated toxicity. On the other hand the results of that study can also be interpreted to imply that damaging of the periportal extracellular matrix from the allyl alcohol is the reason for the failure of the portal biliary ductules to expand into an oval cell population (Petersen et al., 1998).
The origin of oval cells form hepatic cells other than biliary has been supported in several publications (Novikoff and Yam, 1998, Sell, 1983, Sell et al., 1981a, Sell et al., 1981b). The biliary origin of oval cells is not per se excluded in these studies. Data are presented however to suggest that other cell types, in small numbers, may be the cell types of origin. Such cell types include cells seen around the portal ductules (Novikoff and Yam, 1998). Though many studies suggest that liver should be seen in the prototype of small intestine, with a specific niche of tissue specific stem cells, it is also evident that the routine microscopic examination of normal liver sections does not contain a histologic feature set of clustered tissue stem cells comparable to the crypts of the small intestine. Several studies have isolated specific cell subtypes based on expression of specific markers and have demonstrated that these cells have properties of both hepatocytes and biliary cells. EpCAM, a cell adhesion molecule, was used in a recent study to isolate cells with “progenitor cell” properties (i.e. capable of evolving into hepatocytes and biliary cells) (Zhang et al., 2008). EpCAM positive cells are residing in canals of Hering. There was no evidence however that these cells possessed properties that set them out as special participants in the generation of the oval cell response, as compared to other biliary cells expressing hepatocyte specific transcription factors and proliferating early after induction of the protocol causing oval cell expansion.
The extrahepatic origin of oval cells has also been extensively investigated. Engraftment of mice or rats with bone marrow derived hematopoietic stem cells or more complex bone marrow preparations resulted in the appearance of oval cells carrying the specific marker of the transplanted bone marrow (Petersen et al., 1999). Mature hepatocytes carrying the marker of the transplanted bone marrow were also seen, depending on the experimental protocol (Stieger et al., 2007, Stieger et al., 2006). Bone marrow cells from FAH+/LacZ+ mice transplanted and stably engrafted into FAH−/LacZ− recipient mice resulted in colonization of the recipient mouse liver almost entirely by FAH+/LacZ+ hepatocytes (Lagasse et al., 2000). Further studies showed that myeloid precursors were the most likely source of the newly generated FAH+/LacZ+ hepatocytes (Willenbring et al., 2004, Willenbring and Grompe, 2004).
Subsequent studies demonstrated that hematopoietic stem cells have a capacity to fuse with other cell types (Terada et al., 2002). Several publications subsequently emerged to demonstrate that the appearance of new hepatocytes from bone marrow precursors in liver repopulation models was due not to transdifferentiation of the marrow stem cells to hepatocytes, but to fusion of the marrow cells with hepatocytes of the recipient (Wang et al., 2003). In the case of the FAH model, this fusion imparted to the hybrid cells the missing FAH gene and made the chimeric cell competent to proliferate. The fusion was demonstrated with extensive karyotypic analysis showing aneuploid or hyper-diploid karyotypes in the new hepatocytes. Subsequent studies with other tissues demonstrated comparable results. Other studies have claimed, however, that the fusion issue is peculiar to the FAH mouse tyrosinemia model and claims have been made of bona fide new hepatocyte emerging from transdifferentiation of bone marrow hematopoietic precursors. In the rat model of induction of oval cells (PHx plus AAF), a measurable percent of them appeared to derive from bone marrow precursors. When these oval cells were isolated and transplanted into a second generation of rats, they gave rise to karyotypically normal hepatocytes carrying the original DPP IV marker (Oh et al., 2007). In another study, bone marrow from GFP+ rats was engrafted into GFP- rats. With time after engraftment, GFP+ hepatocytes began to emerge in the liver of the recipient animals. It was estimated that the number of emerging hepatocytes was at the rate of 5,000 per day (Tomiyama et al., 2007). The authors characterized this as a relatively small contribution. The importance of the this study resides in the fact that there was no regenerative or proliferative pressure to the liver of the recipient rats to enforce the generation of the “new” hepatocytes. If the GFP+ hepatocytes are the result of fusion with bone marrow hematopoietic precursors, the rate of the fusion under spontaneous circumstances (5,000 cells per day in the rat) is not insubstantial regardless of the conservative characterization by the authors of the study. The mere occurrence of the event, regardless of its mechanism and “significance”, should be studied for its own merit. If on the other hand the new hepatocytes arise spontaneously by a transdifferentiation process, this phenomenon should be studied as well for the same reasons. There have been conflicting reports estimating the number of recipient-derived hepatocytes in liver allografts of humans. Some reports allege high percentages (Theise et al., 2000) whereas in other studies no such hepatocytes were found, and the only recipient derived cells in the liver allograft were endothelial cells of hepatic macrophages (Kupffer cells) (Wu et al., 2003). Bone marrow hematopoietic precursors in culture in the presence of HGF can transdifferentiate to gene expression patterns involving expression of albumin with some cells assuming hepatocytic morphology (Miyazaki et al., 2002, Stock et al., 2008). Trans-differentiation to hepatocytes was also observed in cultures of mesenchymal cells derived from bone marrow (Schwartz et al., 2002). While fusion with hepatocytes in whole animal experiments may be at play, it cannot explain the appearance of hepatocyte-like cells in cell cultures of bone marrow. These studies suggest that the transdifferentiation of bone marrow precursors into hepatocytes in animal models may be an event occurring at a low but measurable rates and the phenomenon definitely warrants further investigation.
IV. Signaling pathways controlling expansion and differentiation of hepatic progenitor cells
There has been significant work in the last two decades related to cell-cell interactions and signaling pathways controlling the expansion of hepatic progenitor cells. The PHx/AAF model is the most characterized. In summary, oval cell expansion emerges soon after there is evidence of enhanced proliferation and emergence of hepatocyte-associated transcription factors in biliary ductules following PHx in the AAF-suppressed rats (Bisgaard et al., 1996, Nagy et al., 1994). The expansion of the oval cell compartment is associated with proliferation of intermingled hepatic stellate cells (Paku et al., 2001). Enhanced expression of alpha fetoprotein is seen early in the response (Sell, 1980), with soon thereafter emergence of expression of albumin. Hepatocyte growth factor (HGF) is expressed by the accompanying stellate cells, and the oval cells do express MET, the receptor for HGF (Alison et al., 1993). Oval cells themselves express TGF alpha, FGF1 and FGF2 (Hu et al., 1996). They also express receptors for EGF, FGFs, VEGF as well as c-Kit, the receptor for Stem Cell Factor (Hu et al., 1996, Fujio et al., 1994). Stellate cells also produce TGFβ1 and the cognate receptors are expressed on the oval cells (Nakatsukasa et al., 1991, Evarts et al., 1990, Hu et al., 1995). TNF receptors are also expressed (Fausto, 2004, Fausto, 2005). The relative contributions or essentiality of any of these factors for oval cell expansion are not fully understood. The response of oval cells to HGF via MET and the potential autocrine loops with TGFα-EGFR and FGF growth factors and cognate receptors is similar to the patterns expressed by hepatocytes during liver regeneration. It is not clear in either case whether the potential autocrine loops are actually operative and control proliferation of the oval cells or the hepatocytes in liver regeneration (Michalopoulos, 2007).
More recent studies have emphasized the involvement of TWEAK, a member of the TNF family, in the proliferation of oval cells. It appears that TWEAK a selectively promotes proliferation of oval cells without having an effect on hepatocytes (Fausto, 2005, Jakubowski et al., 2005). The effect is mediated by the TWEAK receptor Fn14. Both TWEAK and its receptor increase in the standard liver regeneration but it appears to persist longer in the oval cell response. Other members of the same family such as lymphotoxin α and β (Akhurst et al., 2005), TNF itself may also be involved in this process (Brooling et al., 2005). Connective tissue growth factor, a protein of the pericellular matrix, also appears to affect expansion of oval cells. Inhibition of expression of CTGF limits the oval cell response. The mechanism of action of CTGF may involve pericellular matrix rearrangements by binding to fibronectin (Pi et al., 2008, Pi et al., 2005).
Much has been learned about the molecular pathways involved in triggering the expansion of oval cells as a salvage pathway by which the biliary cell compartment rescues the hepatocyte compartment when the latter is unable to proliferate. What is not clear however, is the initial trigger by which the oval cell pathway is activated. Earlier studies using alpha fetoprotein as a marker for the oval cells demonstrated that there is increased proliferation of cells producing alpha fetoprotein and associated with the biliary compartment as part of the regular liver regeneration (Bisgaard et al., 1994). The proliferation of the oval cells however does not continue beyond the termination of liver regeneration. One can speculate that the complement of growth factors and cytokines involved in liver regeneration is also the same with the one that triggers proliferation of the oval cells. When hepatocytes are unable to proliferate, the “regenerative stimuli” elevated in the plasma (Michalopoulos, 2007) continue to stay on. The reasons for the inhibition of hepatocytes may vary but, typically, conditions toxic for hepatocytes (viruses and chemicals) do not affect the capacity of proliferation of the biliary epithelium. In this scenario, absent the operation of pathways that cause a decrease in the levels of regenerative stimuli rising in the plasma (HGF, EGF, TNF, IL6 (itself a proven mitogen for biliary cells (Matsumoto et al., 1994))), their persistence continues to trigger proliferation of biliary cells. There are very little in terms of data however to offer any ground for speculation as to what would be the “signal” inducing expression of hepatocyte-associated genes and transcription factors in the proliferating biliary compartment and eventual complete transformation to hepatocytes. That part of the process is the least understood.
V. Hepatocytes as progenitor cells for biliary epithelium
Analysis of material from human pathology in cholestatic, biliary obstructive or biliary autoimmune disorders has shown that hepatocytes around the portal triads often begin to express genes characteristic of the biliary epithelium (Crosby et al., 1998b, Crosby et al., 1998a). This had raised the possibility that hepatocytes themselves may be precursor cells for the biliary epithelium if the capacity of the latter to proliferate and repair itself is for some reason compromised. The evidence was reinforced by demonstrating that in addition to specific markers, biliary-associated transcription factors were expressed in human hepatocytes in chronic biliary disease (Limaye et al., 2008a). Direct evidence for the transformation of hepatocytes to biliary cells was provided in cell cultures (Nishikawa et al., 2005). Using more complex in vitro systems, these studies were extended into organoid cultures, in which rat livers are completely dissociated into isolated cells and the fraction containing hepatocytes (plus a small contaminant of stellate cells) is placed in a roller bottle culture, in the presence of HGF, EGF, dexamethasone, insulin, transferin and a specially formulated medium HGM (hepatocyte growth medium (Block et al., 1996)). In fourteen days, the cellular elements form a reproducible histologic structure of multiple layers. The top cells facing the medium are biliary epithelial cells, whereas the cells in the layers underneath are hepatocytes, stellate cells and some endothelial cells (Michalopoulos et al., 2001). Using “tagged” hepatocytes (positive for DPP IV) from hybrid rat livers (made by use of the retrorsine/DPP IV protocol, it was demonstrated that the selective hepatocyte marker appeared in the biliary epithelial cells, in the same proportion as the percent of the DPP IV positive hepatocytes in the original cell isolate (Michalopoulos et al., 2002). These studies were extended to the same hybrid liver system in whole animals. Rats with DPP IV hybrid livers were exposed to the biliary toxin DAPM (it kills most of the biliary epithelial cells in 2–3 days) and then subjected to bile duct ligation, a procedure that typically triggers intense proliferation of the biliary epithelium (Michalopoulos et al., 2005). Under these conditions, 45% of the new biliary ductules expressed DPP IV, the marker of the hepatocytes which had not been exposed to retrorsine and were thus capable or re-colonizing the liver. The number of DPP IV positive biliary ductules was down to 2% if DAPM was not used. The latter is significant for two reasons. It shows the probability that periportal hepatocytes would contribute to the proliferation of biliary cells in the absence of any selective stimulus (such as toxic effects and killing of biliary cells). It also shows that the effect was not due to contamination of the hepatocyte preparation by lingering DPP IV positive biliary cells, because if the latter were the case, then DAPM would have decreased the percent of the new ductules positive for DPP IV. Organoid cultures were used to define the signaling pathways mediating the transformation of hepatocytes to biliary cells (Limaye et al., 2008b). It was shown that only HGF and EGF are capable of promoting this transdifferentiation in the organoid cultures. Other growth factors tested were not able to substitute, even though their receptors were expressed and activated in hepatocytes in these cultures. The signaling pathways involved PI3K but were independent of Akt.
VII: Alternative cellular regenerative pathways: Overall assessment
The convergence of evidence from several studies shows that biliary cells and hepatocytes can function as a source of replacement for each other, by a process that involves transdifferentiation, as evidenced by the appearance of transcription factors characteristic and controlling of the hepatocytic or biliary differentiation. The signals controlling these processes arrear very similar to liver regeneration, and HGF and EGFR ligands have been directly implicated in all three situations (standard liver regeneration, oval cell activation, hepatocytes becoming biliary cells). Obviously other signals are also involved and yet more probably remain to be discovered. Whereas oval cells appear as a morphologically distinct population of evanescent duration, the transdifferentiation of hepatocytes to biliary cells appears to be limited to the immediate periportal hepatocytes but there are no apparent distinct populations of intermediate cell forms (Michalopoulos et al., 2005). The restriction of participating hepatocytes to immediate periportal locations is of interest because this is the site from which biliary cells emerge during embryogenesis (Lee et al., 2005, Rubins et al., 2005, Lemaigre, 2003, Clotman et al., 2005). The transdifferentiation of biliary cells to hepatocytes (via oval cells) and vice versa is a very important phenomenon because it occurs in liver failure in humans and it may determine survival. All of these processes, and the comparable findings from studies of liver regeneration, demonstrate the high complexity by which liver as an organ regulates its tissue homeostasis.
Figure 1.
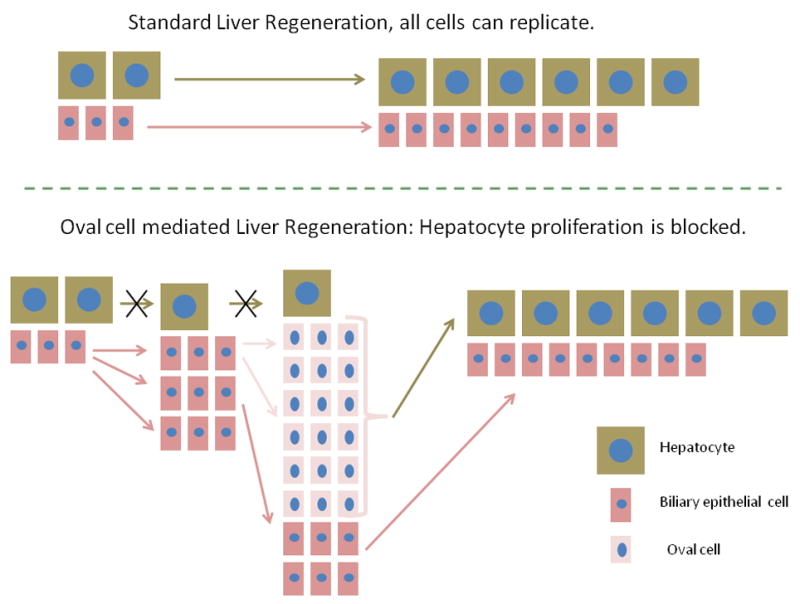
In standard liver regeneration, as after 2/3 partial hepatectomy, all cells are capable of proliferation. Hepatocytes give rise to hepatocytes and biliary cells generate biliary cells. When hepatocyte proliferation is blocked, biliary cells replicate themselves and they also expand into a large number of oval cells, with gene expression patterns of both biliary and hepatocyte specificity. Sources of oval cells are portal ductules and biliary cells of the canals of Hering. The oval cells transition into a hepatocyte phenotype and rescue the hepatocyte compartment.
Abbreviations
- PHx
2/3 partial hepatectomy
- FAH
fumarylacetoacetate hydrolase
- DPP IV
Dipeptidyl peptidase IV
- AAF
Acetylaminofluorene
- AFP
alpha fetoprotein
- C/EBP alpha
CAAT enhancer binding protein alpha
- HNF4
Hepatic Nuclear Factor 4
- HEPPAR
Hepatocyte paraffin antibody
- EpCAM
epithelial cell adhesion molecule
- GFP
green fluorescent protein
- HGF
Hepatocyte Growth Factor
- EGF
Epidermal growth factor
- EGFR
Epidermal growth factor receptor
- MET
HGF receptor protein
- FGF
Fibroblast growth factor
- TGF
transforming growth factor
- VEGF
vascular endothelial growth factor
- TWEAK
TNF related weak inducer of apoptosis
- CTGF
connective tissue growth factor
- DAPM
Methylene Dianiline
- TNF
Tumor necrosis factor
- IL6
Interleukin 6
- HGM
hepatocyte growth medium
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Akhurst B, Matthews V, Husk K, Smyth MJ, Abraham LJ, Yeoh GC. Differential lymphotoxin-beta and interferon gamma signaling during mouse liver regeneration induced by chronic and acute injury. Hepatology. 2005;41:327–335. doi: 10.1002/hep.20520. [DOI] [PubMed] [Google Scholar]
- Alison M, Golding M, Lalani EN, Nagy P, Thorgeirsson S, Sarraf C. Wholesale hepatocytic differentiation in the rat from ductular oval cells, the progeny of biliary stem cells. J Hepatol. 1997;26:343–352. doi: 10.1016/s0168-8278(97)80051-7. [DOI] [PubMed] [Google Scholar]
- Alison MR, Poulsom R, Jeffery R, Anilkumar TV, Jagoe R, Sarraf CE. Expression of hepatocyte growth factor mRNA during oval cell activation in the rat liver. J Pathol. 1993;171:291–299. doi: 10.1002/path.1711710410. [DOI] [PubMed] [Google Scholar]
- Azuma H, Paulk N, Ranade A, Dorrell C, Al-Dhalimy M, Ellis E, Strom S, Kay MA, Finegold M, Grompe M. Robust expansion of human hepatocytes in Fah−/−/Rag2−/−/Il2rg−/− mice. Nat Biotechnol. 2007;25:903–910. doi: 10.1038/nbt1326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bisgaard HC, Nagy P, Santoni-Rugiu E, Thorgeirsson SS. Proliferation, apoptosis, and induction of hepatic transcription factors are characteristics of the early response of biliary epithelial (oval) cells to chemical carcinogens. Hepatology. 1996;23:62–70. doi: 10.1002/hep.510230110. [DOI] [PubMed] [Google Scholar]
- Bisgaard HC, Nagy P, Ton PT, Hu Z, Thorgeirsson SS. Modulation of keratin 14 and alpha-fetoprotein expression during hepatic oval cell proliferation and liver regeneration. J Cell Physiol. 1994;159:475–484. doi: 10.1002/jcp.1041590312. [DOI] [PubMed] [Google Scholar]
- Block GD, Locker J, Bowen WC, Petersen BE, Katyal S, Strom SC, Riley T, Howard TA, Michalopoulos GK. Population expansion, clonal growth, and specific differentiation patterns in primary cultures of hepatocytes induced by HGF/SF, EGF and TGF alpha in a chemically defined (HGM) medium. J Cell Biol. 1996;132:1133–1149. doi: 10.1083/jcb.132.6.1133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brooling JT, Campbell JS, Mitchell C, Yeoh GC, Fausto N. Differential regulation of rodent hepatocyte and oval cell proliferation by interferon gamma. Hepatology. 2005;41:906–915. doi: 10.1002/hep.20645. [DOI] [PubMed] [Google Scholar]
- Clotman F, Jacquemin P, Plumb-Rudewiez N, Pierreux CE, Van der Smissen P, Dietz HC, Courtoy PJ, Rousseau GG, Lemaigre FP. Control of liver cell fate decision by a gradient of TGF beta signaling modulated by Onecut transcription factors. Genes Dev. 2005;19:1849–1854. doi: 10.1101/gad.340305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crosby HA, Hubscher S, Fabris L, Joplin R, Sell S, Kelly D, Strain AJ. Immunolocalization of putative human liver progenitor cells in livers from patients with end-stage primary biliary cirrhosis and sclerosing cholangitis using the monoclonal antibody OV-6. Am J Pathol. 1998a;152:771–779. [PMC free article] [PubMed] [Google Scholar]
- Crosby HA, Hubscher SG, Joplin RE, Kelly DA, Strain AJ. Immunolocalization of OV-6, a putative progenitor cell marker in human fetal and diseased pediatric liver. Hepatology. 1998b;28:980–985. doi: 10.1002/hep.510280412. [DOI] [PubMed] [Google Scholar]
- Demetris AJ, Seaberg EC, Wennerberg A, Ionellie J, Michalopoulos G. Ductular reaction after submassive necrosis in humans. Special emphasis on analysis of ductular hepatocytes. Am J Pathol. 1996;149:439–448. [PMC free article] [PubMed] [Google Scholar]
- Evarts RP, Hu Z, Omori N, Omori M, Marsden ER, Thorgeirsson SS. Precursor-product relationship between oval cells and hepatocytes: comparison between tritiated thymidine and bromodeoxyuridine as tracers. Carcinogenesis. 1996;17:2143–2151. doi: 10.1093/carcin/17.10.2143. [DOI] [PubMed] [Google Scholar]
- Evarts RP, Nagy P, Marsden E, Thorgeirsson SS. A precursor-product relationship exists between oval cells and hepatocytes in rat liver. Carcinogenesis. 1987;8:1737–1740. doi: 10.1093/carcin/8.11.1737. [DOI] [PubMed] [Google Scholar]
- Evarts RP, Nagy P, Nakatsukasa H, Marsden E, Thorgeirsson SS. In vivo differentiation of rat liver oval cells into hepatocytes. Cancer Res. 1989;49:1541–1547. [PubMed] [Google Scholar]
- Evarts RP, Nakatsukasa H, Marsden ER, Hsia CC, Dunsford HA, Thorgeirsson SS. Cellular and molecular changes in the early stages of chemical hepatocarcinogenesis in the rat. Cancer Res. 1990;50:3439–3444. [PubMed] [Google Scholar]
- Factor VM, Radaeva SA, Thorgeirsson SS. Origin and fate of oval cells in dipin-induced hepatocarcinogenesis in the mouse. Am J Pathol. 1994;145:409–422. [PMC free article] [PubMed] [Google Scholar]
- Fausto N. Liver regeneration. J Hepatol. 2000;32:19–31. doi: 10.1016/s0168-8278(00)80412-2. [DOI] [PubMed] [Google Scholar]
- Fausto N. Liver regeneration and repair: hepatocytes, progenitor cells, and stem cells. Hepatology. 2004;39:1477–1487. doi: 10.1002/hep.20214. [DOI] [PubMed] [Google Scholar]
- Fausto N. Tweaking liver progenitor cells. Nat Med. 2005;11:1053–1054. doi: 10.1038/nm1005-1053. [DOI] [PubMed] [Google Scholar]
- Fausto N, Campbell JS, Riehle KJ. Liver regeneration. Hepatology. 2006;43:S45–53. doi: 10.1002/hep.20969. [DOI] [PubMed] [Google Scholar]
- Fiel MI, Antonio LB, Nalesnik MA, Thung SN, Gerber MA. Characterization of ductular hepatocytes in primary liver allograft failure. Mod Pathol. 1997;10:348–353. [PubMed] [Google Scholar]
- Fujio K, Evarts RP, Hu Z, Marsden ER, Thorgeirsson SS. Expression of stem cell factor and its receptor, c-kit, during liver regeneration from putative stem cells in adult rat. Lab Invest. 1994;70:511–516. [PubMed] [Google Scholar]
- Grozdanov PN, Yovchev MI, Dabeva MD. The oncofetal protein glypican-3 is a novel marker of hepatic progenitor/oval cells. Lab Invest. 2006;86:1272–1284. doi: 10.1038/labinvest.3700479. [DOI] [PubMed] [Google Scholar]
- Haque S, Haruna Y, Saito K, Nalesnik MA, Atillasoy E, Thung SN, Gerber MA. Identification of bipotential progenitor cells in human liver regeneration. Lab Invest. 1996;75:699–705. [PubMed] [Google Scholar]
- Hayner NT, Braun L, Yaswen P, Brooks M, Fausto N. Isozyme profiles of oval cells, parenchymal cells, and biliary cells isolated by centrifugal elutriation from normal and preneoplastic livers. Cancer Res. 1984;44:332–338. [PubMed] [Google Scholar]
- Higgins GM, ARM Experimental pathology of the liver, 1: Restoration of the liver of the white rat following partial surgical removal. Arch Pathol. 1931;12:186–202. [Google Scholar]
- Hixson DC, Chapman L, McBride A, Faris R, Yang L. Antigenic phenotypes common to rat oval cells, primary hepatocellular carcinomas and developing bile ducts. Carcinogenesis. 1997;18:1169–1175. doi: 10.1093/carcin/18.6.1169. [DOI] [PubMed] [Google Scholar]
- Hu Z, Evarts RP, Fujio K, Marsden ER, Thorgeirsson SS. Expression of fibroblast growth factor receptors flg and bek during hepatic ontogenesis and regeneration in the rat. Cell Growth Differ. 1995;6:1019–1025. [PubMed] [Google Scholar]
- Hu Z, Evarts RP, Fujio K, Omori N, Omori M, Marsden ER, Thorgeirsson SS. Expression of transforming growth factor alpha/epidermal growth factor receptor, hepatocyte growth factor/c-met and acidic fibroblast growth factor/fibroblast growth factor receptors during hepatocarcinogenesis. Carcinogenesis. 1996;17:931–938. doi: 10.1093/carcin/17.5.931. [DOI] [PubMed] [Google Scholar]
- Hytiroglou P, Theise ND. Telomerase activation in human hepatocarcinogenesis. Am J Gastroenterol. 2006;101:839–841. doi: 10.1111/j.1572-0241.2006.00521.x. [DOI] [PubMed] [Google Scholar]
- Jakubowski A, Ambrose C, Parr M, Lincecum JM, Wang MZ, Zheng TS, Browning B, Michaelson JS, Baetscher M, Wang B, Bissell DM, Burkly LC. TWEAK induces liver progenitor cell proliferation. J Clin Invest. 2005;115:2330–2340. doi: 10.1172/JCI23486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laconi S, Pillai S, Porcu PP, Shafritz DA, Pani P, Laconi E. Massive liver replacement by transplanted hepatocytes in the absence of exogenous growth stimuli in rats treated with retrorsine. Am J Pathol. 2001;158:771–777. doi: 10.1016/s0002-9440(10)64019-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lagasse E, Connors H, Al-Dhalimy M, Reitsma M, Dohse M, Osborne L, Wang X, Finegold M, Weissman IL, Grompe M. Purified hematopoietic stem cells can differentiate into hepatocytes in vivo.[see comment] Nat Med. 2000;6:1229–1234. doi: 10.1038/81326. [DOI] [PubMed] [Google Scholar]
- Lee CS, Friedman JR, Fulmer JT, Kaestner KH. The initiation of liver development is dependent on Foxa transcription factors. Nature. 2005;435:944–947. doi: 10.1038/nature03649. [DOI] [PubMed] [Google Scholar]
- Lemaigre FP. Development of the biliary tract. Mech Dev. 2003;120:81–87. doi: 10.1016/s0925-4773(02)00334-9. [DOI] [PubMed] [Google Scholar]
- Limaye PB, Alarcon G, Walls AL, Nalesnik MA, Michalopoulos GK, Demetris AJ, Ochoa ER. Expression of specific hepatocyte and cholangiocyte transcription factors in human liver disease and embryonic development. Lab Invest. 2008a;88:865–872. doi: 10.1038/labinvest.2008.56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Limaye PB, Bowen WC, Orr AV, Luo J, Tseng GC, Michalopoulos GK. Mechanisms of hepatocyte growth factor-mediated and epidermal growth factor-mediated signaling in transdifferentiation of rat hepatocytes to biliary epithelium. Hepatology. 2008b;47:1702–1713. doi: 10.1002/hep.22221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsumoto K, Fujii H, Michalopoulos G, Fung JJ, Demetris AJ. Human biliary epithelial cells secrete and respond to cytokines and hepatocyte growth factors in vitro: interleukin-6, hepatocyte growth factor and epidermal growth factor promote DNA synthesis in vitro. Hepatology. 1994;20:376–382. [PubMed] [Google Scholar]
- Michalopoulos GK. Liver regeneration. J Cell Physiol. 2007 doi: 10.1002/jcp.21172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michalopoulos GK, Barua L, Bowen WC. Transdifferentiation of rat hepatocytes into biliary cells after bile duct ligation and toxic biliary injury. Hepatology. 2005;41:535–544. doi: 10.1002/hep.20600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michalopoulos GK, Bowen WC, Mule K, Lopez-Talavera JC, Mars W. Hepatocytes undergo phenotypic transformation to biliary epithelium in organoid cultures. Hepatology. 2002;36:278–283. doi: 10.1053/jhep.2002.34858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michalopoulos GK, Bowen WC, Mule K, Stolz DB. Histological organization in hepatocyte organoid cultures. Am J Pathol. 2001;159:1877–1887. doi: 10.1016/S0002-9440(10)63034-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michalopoulos GK, DeFrances M. Liver regeneration. Advances in Biochemical Engineering-Biotechnology. 2005;93:101–134. doi: 10.1007/b99968. [DOI] [PubMed] [Google Scholar]
- Michalopoulos GK, DeFrances MC. Liver regeneration. Science. 1997;276:60–66. doi: 10.1126/science.276.5309.60. [DOI] [PubMed] [Google Scholar]
- Miyazaki M, Akiyama I, Sakaguchi M, Nakashima E, Okada M, Kataoka K, Huh NH. Improved conditions to induce hepatocytes from rat bone marrow cells in culture. Biochem Biophys Res Commun. 2002;298:24–30. doi: 10.1016/s0006-291x(02)02340-9. [DOI] [PubMed] [Google Scholar]
- Nagy P, Bisgaard HC, Thorgeirsson SS. Expression of hepatic transcription factors during liver development and oval cell differentiation. J Cell Biol. 1994;126:223–233. doi: 10.1083/jcb.126.1.223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakatsukasa H, Evarts RP, Hsia CC, Marsden E, Thorgeirsson SS. Expression of transforming growth factor-beta 1 during chemical hepatocarcinogenesis in the rat. Lab Invest. 1991;65:511–517. [PubMed] [Google Scholar]
- Nishikawa Y, Doi Y, Watanabe H, Tokairin T, Omori Y, Su M, Yoshioka T, Enomoto K. Transdifferentiation of mature rat hepatocytes into bile duct-like cells in vitro. Am J Pathol. 2005;166:1077–1088. doi: 10.1016/S0002-9440(10)62328-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Novikoff PM, Yam A. Stem cells and rat liver carcinogenesis: contributions of confocal and electron microscopy. J Histochem Cytochem. 1998;46:613–626. doi: 10.1177/002215549804600507. [DOI] [PubMed] [Google Scholar]
- Nozawa K, Kurumiya Y, Yamamoto A, Isobe Y, Suzuki M, Yoshida S. Up-regulation of telomerase in primary cultured rat hepatocytes. J Biochem. 1999;126:361–367. doi: 10.1093/oxfordjournals.jbchem.a022458. [DOI] [PubMed] [Google Scholar]
- Oh SH, Witek RP, Bae SH, Zheng D, Jung Y, Piscaglia AC, Petersen BE. Bone marrow-derived hepatic oval cells differentiate into hepatocytes in 2-acetylaminofluorene/partial hepatectomy-induced liver regeneration. Gastroenterology. 2007;132:1077–1087. doi: 10.1053/j.gastro.2007.01.001. [DOI] [PubMed] [Google Scholar]
- Overturf K, Al-Dhalimy M, Finegold M, Grompe M. The repopulation potential of hepatocyte populations differing in size and prior mitotic expansion. Am J Pathol. 1999;155:2135–2143. doi: 10.1016/S0002-9440(10)65531-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Overturf K, al-Dhalimy M, Ou CN, Finegold M, Grompe M. Serial transplantation reveals the stem-cell-like regenerative potential of adult mouse hepatocytes. Am J Pathol. 1997;151:1273–1280. [PMC free article] [PubMed] [Google Scholar]
- Paku S, Schnur J, Nagy P, Thorgeirsson SS. Origin and structural evolution of the early proliferating oval cells in rat liver. Am J Pathol. 2001;158:1313–1323. doi: 10.1016/S0002-9440(10)64082-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petersen BE, Bowen WC, Patrene KD, Mars WM, Sullivan AK, Murase N, Boggs SS, Greenberger JS, Goff JP. Bone marrow as a potential source of hepatic oval cells. Science. 1999;284:1168–1170. doi: 10.1126/science.284.5417.1168. [DOI] [PubMed] [Google Scholar]
- Petersen BE, Zajac VF, Michalopoulos GK. Bile ductular damage induced by methylene dianiline inhibits oval cell activation. Am J Pathol. 1997;151:905–909. [PMC free article] [PubMed] [Google Scholar]
- Petersen BE, Zajac VF, Michalopoulos GK. Hepatic oval cell activation in response to injury following chemically induced periportal or pericentral damage in rats. Hepatology. 1998;27:1030–1038. doi: 10.1002/hep.510270419. [DOI] [PubMed] [Google Scholar]
- Pi L, Ding X, Jorgensen M, Pan JJ, Oh SH, Pintilie D, Brown A, Song WY, Petersen BE. Connective tissue growth factor with a novel fibronectin binding site promotes cell adhesion and migration during rat oval cell activation. Hepatology. 2008;47:996–1004. doi: 10.1002/hep.22079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pi L, Oh SH, Shupe T, Petersen BE. Role of connective tissue growth factor in oval cell response during liver regeneration after 2-AAF/PHx in rats. Gastroenterology. 2005;128:2077–2088. doi: 10.1053/j.gastro.2005.03.081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rhim JA, Sandgren EP, Degen JL, Palmiter RD, Brinster RL. Replacement of diseased mouse liver by hepatic cell transplantation. Science. 1994;263:1149–1152. doi: 10.1126/science.8108734. [DOI] [PubMed] [Google Scholar]
- Rhim JA, Sandgren EP, Palmiter RD, Brinster RL. Complete reconstitution of mouse liver with xenogeneic hepatocytes. Proc Natl Acad Sci U S A. 1995;92:4942–4946. doi: 10.1073/pnas.92.11.4942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roskams TA, Theise ND, Balabaud C, Bhagat G, Bhathal PS, Bioulac-Sage P, Brunt EM, Crawford JM, Crosby HA, Desmet V, Finegold MJ, Geller SA, Gouw AS, Hytiroglou P, Knisely AS, Kojiro M, Lefkowitch JH, Nakanuma Y, Olynyk JK, Park YN, Portmann B, Saxena R, Scheuer PJ, Strain AJ, Thung SN, Wanless IR, West AB. Nomenclature of the finer branches of the biliary tree: canals, ductules, and ductular reactions in human livers. Hepatology. 2004;39:1739–1745. doi: 10.1002/hep.20130. [DOI] [PubMed] [Google Scholar]
- Rubins NE, Friedman JR, Le PP, Zhang L, Brestelli J, Kaestner KH. Transcriptional networks in the liver: hepatocyte nuclear factor 6 function is largely independent of Foxa2. Mol Cell Biol. 2005;25:7069–7077. doi: 10.1128/MCB.25.16.7069-7077.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz RE, Reyes M, Koodie L, Jiang Y, Blackstad M, Lund T, Lenvik T, Johnson S, Hu WS, Verfaillie CM. Multipotent adult progenitor cells from bone marrow differentiate into functional hepatocyte-like cells. J Clin Invest. 2002;109:1291–1302. doi: 10.1172/JCI15182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sell S. AFP as a marker for liver cell injury: differentiation of tumor growth, hepatotoxicity, and carcinogenesis. UCLA Forum Med Sci. 1978;20:51–58. [PubMed] [Google Scholar]
- Sell S. Heterogeneity of alpha-fetoprotein(AFP) and albumin containing cells in normal and pathological permissive states for AFP production: AFP containing cells induced in adult rats recapitulate the appearance of AFP containing hepatocytes in fetal rats. Oncodev Biol Med. 1980;1:93–105. [PubMed] [Google Scholar]
- Sell S. Comparison of oval cells induced in rat liver by feeding N-2-fluorenylacetamide in a choline-devoid diet and bile duct cells induced by feeding 4,4′-diaminodiphenylmethane. Cancer Res. 1983;43:1761–1767. [PubMed] [Google Scholar]
- Sell S. Liver stem cells. Modern Pathology. 1994;7:105–112. [PubMed] [Google Scholar]
- Sell S, Leffert HL, Shinozuka H, Lombardi B, Gochman N. Rapid development of large numbers of alpha-fetoprotein-containing “oval” cells in the liver of rats fed N-2-fluorenylacetamide in a choline-devoid diet. Gann. 1981a;72:479–487. [PubMed] [Google Scholar]
- Sell S, Osborn K, Leffert HL. Autoradiography of “oval cells” appearing rapidly in the livers of rats fed N-2-fluorenylacetamide in a choline devoid diet. Carcinogenesis. 1981b;2:7–14. doi: 10.1093/carcin/2.1.7. [DOI] [PubMed] [Google Scholar]
- Sells MA, Katyal SL, Sell S, Shinozuka H, Lombardi B. Induction of foci of altered, gamma-glutamyltranspeptidase-positive hepatocytes in carcinogen-treated rats fed a choline-deficient diet. Br J Cancer. 1979;40:274–283. doi: 10.1038/bjc.1979.176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinozuka H, Lombardi B, Sell S, Iammarino RM. Early histological and functional alterations of ethionine liver carcinogenesis in rats fed a choline-deficient diet. Cancer Res. 1978;38:1092–1098. [PubMed] [Google Scholar]
- Shiojiri N, Lemire JM, Fausto N. Cell lineages and oval cell progenitors in rat liver development. Cancer Res. 1991;51:2611–2620. [PubMed] [Google Scholar]
- Sirica AE, Cihla HP. Isolation and partial characterizations of oval and hyperplastic bile ductular cell-enriched populations from the livers of carcinogen and noncarcinogen-treated rats. Cancer Res. 1984;44:3454–3466. [PubMed] [Google Scholar]
- Sirica AE, Mathis GA, Sano N, Elmore LW. Isolation, culture, and transplantation of intrahepatic biliary epithelial cells and oval cells. Pathobiology. 1990;58:44–64. doi: 10.1159/000163564. [DOI] [PubMed] [Google Scholar]
- Stieger B, Peters R, Sidler MA, Meier PJ. Hepatocyte transplantation: potential of hepatocyte progenitor cells and bone marrow derived stem cells. Swiss Med Wkly. 2006;136:552–556. doi: 10.4414/smw.2006.11419. [DOI] [PubMed] [Google Scholar]
- Stieger B, Peters R, Sidler MA, Meier PJ. Hepatocyte transplantation: potential of hepatocyte progenitor cells and bone marrow derived stem cells. Swiss Med Wkly. 2007;137(Suppl 155):55S–59S. [PubMed] [Google Scholar]
- Stock P, Staege MS, Muller LP, Sgodda M, Volker A, Volkmer I, Lutzkendorf J, Christ B. Hepatocytes derived from adult stem cells. Transplant Proc. 2008;40:620–623. doi: 10.1016/j.transproceed.2008.01.058. [DOI] [PubMed] [Google Scholar]
- Stocker E, Heine WD. Regeneration of liver parenchyma under normal and pathological conditions. Beitr Pathol. 1971;144:400–408. [PubMed] [Google Scholar]
- Stocker E, Wullstein HK, Brau G. [Capacity of regeneration in liver epithelia of juvenile, repeated partially hepatectomized rats. Autoradiographic studies after continous infusion of 3H-thymidine (author’s transl)] Virchows Arch B Cell Pathol. 1973;14:93–103. [PubMed] [Google Scholar]
- Tatematsu M, Ho RH, Kaku T, Ekem JK, Farber E. Studies on the proliferation and fate of oval cells in the liver of rats treated with 2-acetylaminofluorene and partial hepatectomy. Am J Pathol. 1984;114:418–430. [PMC free article] [PubMed] [Google Scholar]
- Terada N, Hamazaki T, Oka M, Hoki M, Mastalerz DM, Nakano Y, Meyer EM, Morel L, Petersen BE, Scott EW. Bone marrow cells adopt the phenotype of other cells by spontaneous cell fusion. Nature. 2002;416:542–545. doi: 10.1038/nature730. [DOI] [PubMed] [Google Scholar]
- Theise ND, Nimmakayalu M, Gardner R, Illei PB, Morgan G, Teperman L, Henegariu O, Krause DS. Liver from bone marrow in humans [In Process Citation] Hepatology. 2000;32:11–16. doi: 10.1053/jhep.2000.9124. [DOI] [PubMed] [Google Scholar]
- Theise ND, Saxena R, Portmann BC, Thung SN, Yee H, Chiriboga L, Kumar A, Crawford JM. The canals of Hering and hepatic stem cells in humans. Hepatology. 1999;30:1425–1433. doi: 10.1002/hep.510300614. [DOI] [PubMed] [Google Scholar]
- Tomiyama K, Miyazaki M, Nukui M, Takaishi M, Nakao A, Shimizu N, Huh NH. Limited contribution of cells of intact extrahepatic tissue origin to hepatocyte regeneration in transplanted rat liver. Transplantation. 2007;83:624–630. doi: 10.1097/01.tp.0000253942.16061.d9. [DOI] [PubMed] [Google Scholar]
- Wang X, Willenbring H, Akkari Y, Torimaru Y, Foster M, Al-Dhalimy M, Lagasse E, Finegold M, Olson S, Grompe M. Cell fusion is the principal source of bone-marrow-derived hepatocytes.[see comment] Nature. 2003;422:897–901. doi: 10.1038/nature01531. [DOI] [PubMed] [Google Scholar]
- Willenbring H, Bailey AS, Foster M, Akkari Y, Dorrell C, Olson S, Finegold M, Fleming WH, Grompe M. Myelomonocytic cells are sufficient for therapeutic cell fusion in liver. Nat Med. 2004;10:744–748. doi: 10.1038/nm1062. [DOI] [PubMed] [Google Scholar]
- Willenbring H, Grompe M. Delineating the hepatocyte’s hematopoietic fusion partner. Cell Cycle. 2004;3:1489–1491. doi: 10.4161/cc.3.12.1304. [DOI] [PubMed] [Google Scholar]
- Wu T, Cieply K, Nalesnik MA, Randhawa PS, Sonzogni A, Bellamy C, Abu-Elmagd K, Michalopolous GK, Jaffe R, Kormos RL, Gridelli B, Fung JJ, Demetris AJ. Minimal evidence of transdifferentiation from recipient bone marrow to parenchymal cells in regenerating and long-surviving human allografts. American Journal of Transplantation. 2003;3:1173–1181. doi: 10.1046/j.1600-6143.2003.00137.x. [DOI] [PubMed] [Google Scholar]
- Yamaguchi Y, Nozawa K, Savoysky E, Hayakawa N, Nimura Y, Yoshida S. Change in telomerase activity of rat organs during growth and aging. Exp Cell Res. 1998;242:120–127. doi: 10.1006/excr.1998.4102. [DOI] [PubMed] [Google Scholar]
- Yang L, Faris RA, Hixson DC. Characterization of a mature bile duct antigen expressed on a subpopulation of biliary ductular cells but absent from oval cells. Hepatology. 1993;18:357–366. [PubMed] [Google Scholar]
- Zhang L, Theise N, Chua M, Reid LM. The stem cell niche of human livers: symmetry between development and regeneration. Hepatology. 2008;48:1598–1607. doi: 10.1002/hep.22516. [DOI] [PubMed] [Google Scholar]