

Abstract
Leishmania parasites have been widely used in experimental models to understand generation, maintenance and failure of immune responses underlying resistance and susceptibility to infection. The clinical outcomes of Leishmania infection depend on the infecting species and the immune status of the host. Noticeably most people exposed Leishmania never develop overt disease. Understanding the immunological events that result in failure or successful control of the parasites is fundamental to both design and evaluation of vaccines and therapies against the leishmaniases. Recent studies visualizing immune response to Leishmania major in the skin have given new insights into the different immune cells acting as hosts the parasite during different stage of infection. Control of Leishmania infection and disease progression has been associated with generation of T-helper (Th) 1 and Th2 responses respectively. Though still valid in several aspects, the Th1/Th2 paradigm is an oversimplification in need of revision. Th2 polarization has never explained severity of human leishmanial disease and a number of other T-cell subsets, including regulatory T- and Th17- cells, have important roles in susceptibility and resistance of both experimental and human leishmanial disease. This review gives an updated overview of immunological response considered to be of importance in protection, susceptibility, disease progression and cure of leishmaniasis, with a special emphasis on human diseases.
Keywords: Leishmania, Immunology, T cells, Innate immunity, Human, Apoptosis, Vaccine
INTRODUCTION
All natural Leishmania infections start when Leishmania promastigotes are injected into the skin dermis of humans and other warm-blooded animals. To survive, the parasite must resist exposure to host serum component and destruction by innate immune cells present in or rapidly recruited to the skin. The skin is a complex immunological organ in which multiple innate immune cells [Table 1] function to protect the host from infectious pathogens. Normal skin of adult humans also contains a substantial number of T cells, nearly twice that present in the circulation,[1] which may play an important role in the local response.
Table 1.
Innate immune cells in the skin and their role in leishmanial disease (Leish)
| Cell type | General function | Observations in murine Leish | Observations in Leish in vitro |
|---|---|---|---|
| Keratinocytes | Sensors of injury & infection | - | Source of IL-10, associated with PKDL[111] |
| Location: epidermis | |||
| Langerhans cells | Antigen presentation in certain infections | Uncertain, not necessary for induction of Th1 responses[33] | Correlation between high LC density and acute cutaneous L. tropica disease[112] |
| Location: epidermis | Induction of peripheral tolerance | ||
| Th2 induction | |||
| Cross priming of naïve CD8+ T cells | |||
| Dermal DC | Immune surveillance | Sensors of infection[33] | - |
| Location: dermis | Antigen presentation | ||
| Cross presentation to CD8+ T cells | |||
| Dermal macrophages | Antimicrobial activity and production of pro- and anti-inflammatory mediators | Can act as host cells [8,32] | Host cells (non-human primate skin)[6] |
| Location: dermis | |||
| Plasmacytoid DC | IFNα production | Leishmania loaded pDC can induce protective immunity[113] | - |
| Location: dermis | Activation of NK cells, B cells T cells and myeloid DC cells | ||
| Mast cells | Regulating later Inflammatory response by Neutrophils | Sentinels, contribute to DC recruitment[114] Tissue pathology[115] | Elevated numbers in MCL lesion[117] |
| Location:Dermis | Susceptibility[116] | Possibly an association with wound healing[118] | |
| Monocyte derived | Inflammatory cells | Induction of protective immunity[7] | Species dependent production of IL-12, co-stimulation[36] |
| Inflammatory DC | T cell stimulation Production of IL-12, iNOS and TNFα | Primary cells harboring parasites in later stages of disease development (healing mice)[28] | |
| Location: Inflamed dermis | |||
| Neutrophils polymorph nucleated cells (PMN) | Uptake and destruction of pathogens | Temporary early major host cells facilitating L. major infection[8] | *Human PMN can kill promasitigotes and amastigotes.[14] |
| Silent Transfer of parasites into macrophages[13] | |||
| Location: dermis | Protective[16] | Found in lesions[18,20] | |
| Tissue pathology – in later stages of disease[22,23] | Can harbor parasites in VL[9] | ||
| NK cells | Early source of IFNγ | Contribute to early resistance against the parasite[37,119] | Associated with protection and cure[43,45] |
| Location: Inflamed dermis |
The general functions of cells have been adapted from Nestle et al.[110]
The interaction with the complement system depends on the developmental stage and the species of the parasite. In short, exponentially multiplying log-phase promastigotes are sensitive to complement mediated lysis and perform poorly in experimental infections while metacyclic promastigotes and amastigotes are more resistant and more infective.[2,3] However, no parasite is completely resistant to physiological plasma complement levels.[4] Prompt infection of susceptible host cell may be essential for survival. While surface deposition of complement can cause destruction, Leishmania can use deposited C3b, which is rapidly converted into iC3b, to facilitate parasite entry into macrophages and neutrophils via complement receptor (CR)3.[3,5]
NEUTROPHILS – TROJAN HORSES, EXPLOITED INTERMEDIATE HOST CELLS OR PARASITE KILLERS?
Leishmania infection has been assumed to be initiated by direct parasitization of skin resident macrophages[6] whereas uptake by skin DCs has been linked to priming and shaping the T cell response.[7] Recent studies have, however, slightly changed this view; in vivo imaging of sandfly transmitted L. major infection revealed neutrophils as the first cells to be infected by the parasites.[8]
Neutrophils are primary antimicrobial effector cells, with the main function to phagocytose and destroy invading pathogens. Neutrophils are rapidly recruited to sites of the body where tissue damaged has occurred, such as the site of a sandfly bite. Most microorganisms are rapidly killed when taken up by neutrophils, but a few mostly intracellular pathogens, can survive the destructive milieu of these cells.
Following sandfly transmission or needle inoculation with L. major, invading neutrophils were observed to rapidly and efficiently capture parasites.[8] Infiltrating neutrophils did not destroy the parasites, instead they facilitated infection as depletion of neutrophils prior to infection reduced the parasites load and delayed onset of disease.[8] Infection of neutrophils is transient and within a week post infection macrophages/monocytes take over as the primary host cell.[8] However, in human VL neutrophils have been reported to harbor parasites during active disease.[9,10] It may be noted that human blood is much more neutrophil rich than that of mice.[11]
Laskay and colleagues have supported by in vitro studies showing that Leishmania infected apoptotic neutrophils can be taken up by macrophages that allow parasites to thrive, proposed a model in which neutrophils, act as “Trojan horses” for Leishmania.[12,13] Apoptotic neutrophils are normally cleared without triggering activation of macrophages. Thus, uptake of infected apoptotic neutrophils could facilitate silent entry and infection of macrophages. Leishmania may also extend the life span of a neutrophil by delaying apoptosis, suggested to give monocytes time to infiltrate the site of infection and become infected by apoptotic neutrophils.[13] However, the in vivo images by Peters et al. were not able to capture neutrophils uptake by monocytes/macrophages.[8] Instead parasites were observed to egress dying neutrophils, to invade macrophages. Regardless if neutrophils act as Trojan horses or not, compelling evidence indicates that the Leishmania parasite, in the early infection phase, can both evade and exploit neutrophils to ensure its survival.
Nevertheless, when appropriately activated, neutrophils can kill intracellular pathogens such as Leishmania[14] and there are several reports suggesting that neutrophils play a role in early protection against Leishmanial disease.[15–17] Leishmania amazonensis promastigotes can induce and be killed by neutrophils extracellular traps (NETs). The same would appear to be the case for amastigotes, albeit not to the same extent as promastigotes. More interestingly meshes of DNA and elastate suggestive of NET were found in skin biopsies of patients.[18] Furthermore, in a mouse model of Leishmania donovani infection, using amastigotes for infection, neutrophils were found to have a protective function.[16]
The capacity of neutrophils to function as immune evasion targets probably depend on the genetic background of the host, the parasite strain and the developmental stage of the parasite used.[19] While metacyclic promastigotes may survive in neutrophils, non-metacyclic ones can rapidly be killed. Neutrophils may act in disease-stage specific way, being permissive hosts for metacyclic promastigotes while contributing to the over-all inflammatory and parasitocidal response in active lesions. In both human and murine leishmaniasis neutrophils are prominent infiltrates in lesions;[20,21] their presence at the site of infection can cause immune mediated tissue pathology.[22,23]
MANIPULATING ANTIGEN PRESENTING CELLS A KEY STRATEGY FOR SURVIVAL
For a productive infection, the Leishmania parasites need to establish in macrophages. Macrophages possess potent antimicrobial functions and activated macrophages can kill Leishmania. To survive the parasite need to avoid macrophage activation and recognition by T cells. The ability to survive in macrophages is partly stage specific with metacyclic promastigotes having better capacity to survive compare to pro-cyclic promastigotes.[3] The parasites have several strategies by which macrophage activation can be prevented including: silent entry utilizing engagement of non-triggering receptors such as CR1 and the phosphatidyl serine (PS) receptor;[24,25] more direct inhibition of macrophage function by interfering with NFB transcription and IL-12 production, down regulation of MHC class II; promoting production of regulatory cytokines like IL-10 and TGFβ.[26,27]
Dermal macrophages are readily infected with Leishmania and permit differentiation and growth and may initiate the infection.[6] However, most targets for Leishmania would appear to be infiltrating monocytes/macrophages, which enter the site of infection one to two days post infection.[8,28] Interestingly, recent studies show that macrophages may not be the main host cells for the parasites in chronic stages of healing disease. In chronic self-healing L. major infection (in C57Bl/6) mice) TNF and iNOS producing CD11b+CD11c+Ly6C+MHC-II+DC (TIP-DC), which most likely are derived from monocytes, host the majority parasites in the skin.[28,29] If this is related to cure and generation of a protective Th1 response and/or preparing the parasite for transmission to blood feeding sandflies, is not known.
APOPTOSIS – A WAY TO AVOID IMMUNE ACTIVATION AND PROMOTE SURVIVAL
Exposure of PS is fundamental for the non-inflammatory phagocytosis of apoptotic cells. Recognition of this phospholipid by macrophages induces TGFβ secretion, IL-10 synthesis and inhibits NO production. Utilizing apoptotic cells as vectors or mimicking mammalian apoptotic cells is a strategy to escape host protective inflammatory response. Leishmania parasites have evolved several strategies to use apoptosis to it advantage [Figure 1].
Figure 1.
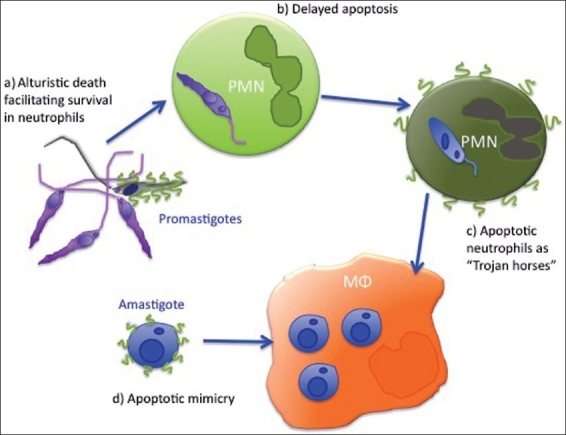
Apoptosis - a way to avoid immune activation and promote survival; a) Sandflies inject apoptotic parasites together with viable. Apoptotic promastigotes facilitate infection and prevent activation of neutrophils (PMN).[108,109]; b) Parasites delay neutrophil apoptosis, giving monocytes/macrophages (Mφ) time to enter the site of infection; c) Silent entry of parasite into macrophages via apoptotic neutrophils.[12]; d) Viable amastigotes expose PS and mimic apoptotic cells. This facilitates internalization and increases macrophage susceptibility to leishmanial growth[25]
DENDRITIC CELLS ORCHESTRATING THE IMMUNE RESPONSE
Leishmania parasites-DC, interactions are complex, inconsistent and may lead to the control of infection or progression of disease. Activation of DC varies in quantity and quality depending upon developmental stage and the species/strain of Leishmania as well as DC cell subset and exogenous stimuli involved in different studies.[30]
Several different types of DC with different location and probably different function exist in the skin. The first studies of murine skin DC indicated epidermal Langerhans cells (LC) as the important cells for sensing, uptake and transport of Leishmania to the lymph node.[31] More recent studies have shown that it is dermal DCs that are involved in the early recognition of the parasite. Dermal DC can efficiently take up and incorporate parasites in vacuoles[32] and have been suggested by some to act as principal antigen presenting cells in leishmaniasis,[33] while other suggest lymph node resident DC as the initiators of the immune response.[34]
Leishmania have evolved several strategies to avoid or dampen DC, while some appear general to Leishmania other tend to be species related.[30] This may be, in part, the explanation for the severity of disease caused by the different species. In general, more papers report inhibitory effects of L. donovani and the South American species (L. amazonensis, Leishmania braziliensis) compared to L. major. In human cells L. donovani has been suggested to block maturation of human DC.[35] Production of IL-12 by DC, which is essential for the initiation of a protective immune response in mice (and probably also in humans) is differently affected by L. donovani and L. major: while uptake of L. major by human monocyte derived DC efficiently prime DC for IL-12 production uptake of L. donovani does not.[36]
A RECOVERED ROLE FOR NATURAL KILLER (NK) CELLS IN INDUCTION OF IMMUNITY
Together with phagocytes, NK cells represent the first line of defense against pathogens by two principal mechanisms, cytolytic destruction of infected cells and secretion of pro-inflammatory cytokines (e.g. IFNγ, TNFα).
Early studies of experimental leishmaniasis in C3H/HeN mice indicated that IFNγ production by NK cells was important for generation of protective immune responses and control of infection. Subsequent studies, which of note, were mainly done in mice on C57Bl/6 background, demonstrated, however, that NK cells are not required for generation of adequate T helper type-1 response and protective immunity. Their presence however, may delay onset of disease as Balb/c mice lacking NK cells develop lesions faster and harbor more parasite.[37] Thus, NK cells can serve a function in control of Leishmania burden during early phases of infection through their ability to rapidly respond with IFNγ production. In vitro, human NK cells have been shown to have the ability to be directly activated to IFNγ production by Leishmania promastigotes or their LPG.[38,39]
A new interest in NK cells and Leishmania infection has evolved from the interaction between these cells and DC. Activated NK cells promote DC maturation, while they may kill autologous immature DC.[40] DC can on the other hand efficiently prime resting NK cells. In vitro resting NK cells have been shown to promote activation of DCs pre-infected with L. amazonensis promastigotes and these activated DCs can, in turn, mostly via cell contact-dependent mechanisms stimulate NK cells.[41] L. major infection induces NK cells to secretion of IFNγ and in vivo imaging has shown that NK cells are recruited to the paracortex, a strategic area in the lymph node, where they can interact with DC and regulate co-localized CD4 T cells responses.[42]
In patients, NK cell number and activity has mainly been associated with protection against or healing of disease. Patients with active leishmaniasis (cutaneous and visceral) have been reported to have a reduction in the frequency of peripheral NK cells[43,44] and recently an increased frequency NK cells, following immunotherapy, in a L. amazonensis diffuse cutaneous leishmaniasis (DCL) patients was associated with cure.[45]
EFFECTOR T CELLS IN CONTROL AND FAILURE OF LEISHMANIAL INFECTION
Protective immune responses against Leishmania, in self-healing strains of mice (C57BL/6, C3H, CBA), are intimately linked to development of a Th1 response and IFNγ production. In experimental self-healing disease DC are stimulated to produce IL-12, which drives the generation of Th1 cells (effector and memory). T cell dependent IFNγ in turn activates macrophages to upregulation of iNOS and NO production, which results in killing of intracellular parasites and control of disease.[46,47] Disease progression has to a large extent been viewed as development of Th2 responses and IL-4 based on studies of L. major infection in Balb/c mice. The polarized Th1/Th2 responses in mice have been more thoroughly reviewed by others[48] and is not the scope of this review.
Most data point to the fact that same or similar Th1 dependent mechanisms are involved in control of human disease. Self-healing forms of leishmaniasis and cure of VL is typically accompanied by parasite specific proliferation and IFNγ production. Human macrophages are activated to kill intracellular parasites by IFNγ and exogenous IFNγ can promote cure of human CL.[49] Though Th2 responses can act in favor of the parasite, polarized Th2 response has never been able to explain non-curative or visceralizing human disease. Th2 independent disease progression is also supported by studies on non-healing disease in the Th1 phenotypic B6 mice.[50] In this context it can also be noted that in patients with VL the effect of IFNγ administration was limited[51] and in human CL, IFNγ production by CD4+ cells, alone, in response to Leishmania antigens is not predictive of protection or disease development.[52] This indicates that other mechanisms acting in synergy with IFNγ or counteracting the effects of IFNγ are as important.
ADVANCING THE TH1/TH2 PARADIGM: TH17 AND REGULATORY T CELLS AND THEIR ROLE IN PATHOLOGY OF LEISHMANIAL DISEASES
Th17 and Treg are today widely accepted subsets with important functions in induction and control of the inflammatory response. Both Th17 and Treg have a greater degree of plasticity in their differentiation decision, as compared to conventional Th1 and Th2 cells, enabling response to signals provided by the environment in which they reside.[53]
Th17 cells are pro-inflammatory T helper cells, hallmarked by their ability to secrete IL-17. IL-17 is involved in recruitment, migration and activation of neutrophils and Th17 cells have an important function in protecting surfaces against certain extracellular bacteria and fungal pathogens, but can also mediate severe immune pathologies.[54]
In experimental leishmaniasis Th17 cells have been associated with tissue destruction: IL-17 deficient Balb/c mice develop smaller lesions, have decreased CXCL2 accumulation and fewer neutrophils in lesions as compared to wild type, while elevated IL-17 conferred no reduction in parasite load.[22]
Interestingly, a recent study of human VL linked IL-17 and IL-22 (a pro-inflammatory cytokine, produced by Th17 and NK cells), to protection against human kala-azar caused by L. donovani. L. donovani was furthermore shown to stimulate generation of cells producing IL-17, IL-22 as well as IFNγ by human T cells.[55] IL-27 is an IL-12 related cytokine, produced mainly by macrophages and DC, thought to be important in regulation of Th17 cells. In C57Bl/6 mice IL-27 is important in early Th1 development, mediating suppression of the early IL-4 burst that occur in B6 mice.[56] IL-27 also has anti-inflammatory properties, mediated through the ability of IL-27 to suppress Th17 cells[57] and induction of naïve human CD4 cells to IL-10 production.[58]
Interleukin-27 receptor deficient mice display enhanced resistance to L. donovani infection[59] and findings of elevated levels of IL-27 together with low RoRγT/IL-17 in VL patients before treatment, implicate IL-27 in VL pathogenesis.[60] However mice deficient in IL-27R develop severe liver immunopathology when infected with L. donovani[59] and more severe cutaneous lesions in infection with non-healing L. major.[57] Both studies showed that CD4 T cells were linked to pathology, in the latter study IL-27 was found to regulate both IL-10 and IL-17, and tissue pathology was associated with IL-17 producing T cells. Thus, the elevated levels of IL-27 in human VL may serve an important function suppression of IL-17 producing CD4 T cells and subsequent tissue damage by neutrophils.
REGULATORY T CELLS AND PARASITE PERSISTENCE
Regulatory CD4 T cells can broadly be divided into two categories - natural Foxp3+ (CD4+CD25high) regulatory T cells (nTreg) that arise in the thymus and inducible regulatory T cells generated in the periphery, the latter can be both, Foxp3+ (iTreg) and adaptive FoxP3-, type-1 regulatory T cells (Tr1). All regulatory T cells act to counteract inflammatory immune response to limit tissue damage and absence of regulatory T cells is linked to a number of autoimmune conditions.
The skin (and other epithelial surfaces) has a high frequency of steady state, nTreg that function to suppress the generation of harmful immune response to infectious and non-infectious antigens to, which the skin is regularly exposed. The presence of these cells may, however, precondition the skin for survival of Leishmania parasites and favor long-term parasite survival.[61] In humans CD4+CD25+ Treg cells are found in cutaneous lesions[62] and elevated intra lesional FoxP3 and IL-10 have been associated with unresponsiveness to treatment during L. amazonensis infection.[63]
IL-10 is a cytokine intimately linked with disease progression of both murine and human Leishmania infection.[64] Experimental models have clearly demonstrated the central role played by IL-10 in pathology and parasite persistence.[65–67] In human VL, elevated levels of IL-10/IL-10 mRNA are found systemically as well as in spleen, bone marrow and lymph nodes. A role for IL-10 in human VL pathology is supported by studies indicating that IL-10 blockade can enhance VL PBMC IFNγ responses and inhibit VL serum promoted parasite replication in macrophages.[44,68,69] However, if the IL-10, as assumed, is a major suppressor of effector T cell in VL patients, remains to be proved. In human CL, elevated IL-10 has been demonstrated in lesions.[70–72] A recent genetic analysis of IL10-819C/T polymorphism, in the IL10 promoter, showed that the C allele, which is linked to higher levels of IL-10 production, is associated with increased risk of developing cutaneous lesions in populations exposed to L. braziliensis.[72]
All regulatory T cells can be sources of IL-10. In leishmanial infection most data point to antigen-induced Foxp3- T cells, producing IL-10, as being responsible for delayed healing associated with disease progression.[50] In line with this T cells other than those expressing FoxP3 would appear to be the main source of IL-10 in human VL.[43,44] The B6 mouse model of non-healing disease indicates that Th1, which secrete IFNγ cells, can be the main source IL-10.[50] IL-10 secretion by Th1 cells is a self-regulating mechanism evolved to minimize T cell mediated immune-pathology.
Central cytokines in healing and progression of leishmanial disease are summarized in Table 2.
Table 2.
Expanding the Th1/ Th2 paradigm: A brief summary of central cytokines in healing and progression of Leishmania infection
| Cytokine | Producer/s | Function in murine Leish | Human Leish (correlation) |
|---|---|---|---|
| IL-4 | Th2 cells | Inhibition of Th1 responses | Some association with non-healing CL and VL[120,121] |
| Mast cells basophils | Responsible for progression in Balb/c mice[48] | ||
| IL-10 | Many, including monocytes/macrophages, T cells and epithelial cells | Promote parasite persistence | Associated with visceral and non-healing disease.[64,111,122] |
| Down regulation of macrophage function. Counter act Th1 cells[65,66,99] | |||
| IL-12 | Dendritic cells | Required for induction of protective Th1 response[123] | Addition to VL PBMC induce IFNγ and cytotoxic Monocytes response[124,125] |
| Neutrophils | |||
| B cells | |||
| IL-17 | Th17 cells, neutrophils | Disease progression in susceptible Balb/C mice[22] | Associated with protection from disease[55] |
| IL-22 | Th17 cells | - | Associated with protection from disease[55] |
| NK-22 cells | |||
| IL-27 | Dendritic cells Monocytes | Fewer parasites, but more tissue pathology due to impaired regulatory response[57,59] | Associated with active VL[60] |
| Macrophages | |||
| IFNγ | Many, most importantly Th1 cells and NK cells | Required for protective responses, KO mice cannot control infection[48] | Antigen specific INFγ response by PBMC are associated with cure and protection[126] |
| May promote cure of CL[49] | |||
| TNFα | Many, mainly macrophages | Required for control of most, leishmania strains.[127] | Associated with protection and cure[126] |
| Cause tissue destruction and loss of splenic architecture in experimental VL[128] | Case reports of VL in TNF antagonist treated patients[129] | ||
| High levels associated with tissue pathology[130] | |||
| TGFβ | Monocytes /macrophages | Regulatory function associated with disease progression.[131] | Associated with non-healing phenotype (MCL, PKDL)[131,134] |
| T (reg) cells Chondrocytes | Suppression of IFNγ by NK cells[132] | ||
| Act in synergy with IL-10[133] |
Important producers of the respective cytokines in leishmanial disease
CD8 T CELLS ARE PROTECTIVE AND DESTRUCTIVE
The role of CD8+ T cells is still not completely defined in Leishmania infection. Though a number of early reports suggested a role for CD8+ cells in immunity against L. major infection,[73,74] CD8+ T cells, were for a long time thought to play a secondary role as CD8 cells alone could not induce protective immunity and CD8 defective mice, were able to control infection.[75] However, Belkaid et al. later demonstrated that CD8 cells actually were required for healing when C57BL/6 mice were infected with a low, and more physiological relevant, dose of parasites and in experimental infection with L. donovani both CD8 and CD4 can on their own cells prevent reactivation of disease.[76,77]
CD8+ T cells participate in protection against pathogens by two major mechanisms: production of cytokines (IFNγ and TNF-α) and by direct killing of infected cells. In Leishmania infection the main contribution of CD8 T cells in immunity is considered to be through IFNγ production. Cytotoxic T-lymphocyte (CTL)-mediated mechanisms in the regulation and control of Leishmania infection remain largely unexplored.[78] Perforin (together with IFNγ) has, however, been suggested as an important effector molecule in vaccination induced immunity against L. amazonensis.[79] Both murine and human Leishmania infection can prime CD8 T cells for killing of antigen pulsed macrophages.[80,81] In vivo studies have moreover indicated that the Fas-Fas ligand (L) pathway contributes to healing of lesions induced by L. major,[82] as Fas- and FasL-deficient mice cannot control infection despite upregulation of IL-12 and NO production. Moreover, CD95 is required for the early control of parasite burden in the liver of L. donovani -infected mice.[83] In contrast, elevated levels of Fas in human CL lesions have been suggested to contribute to ulcer formation.[84]
CD8+ T-cells have been associated with both cure and pathology in human leishmaniasis: An expanded CD8+ cell population was observed in the draining lymph node prior to ulcer development, implicating CD8 mediated immunity in the early containment of Leishmania infection.[85] An increase in responding CD8+ cells has been associated with cure of L. braziliensis CL.[86,87] Exacerbated CD8+ activity, in addition to a poor regulatory response, could however, underlie an unfavorable fate with regard to MCL. Recruitment of CD8+ T cells expressing granzyme associated with lesion progression of CL caused by L. braziliensis and more CD8 cells were found in relapse cases.[88,89] Accumulation of CD8 cells have also been linked to PKDL.[90]
CD8 T cells can just like CD4 T cells have natural or acquired regulatory properties.[91] IL-10 producing CD8 cells of memory phenotype have been identified in humans infected with Leishmania guanyensis.[92] The function of these IL-10 producing CD8 T cells in leishmanial disease is unknown.
B CELLS AND ANTIBODY RESPONSES: HARMFUL OR PROTECTIVE
B cells and antibodies are generally not considered to be of major importance in protective immunity against Leishmania. Antibodies are not effective at killing the parasite as it hides inside the parasitophorus vacuole and antibody responses in self-healing cutaneous disease are very modest.
High levels of Leishmania specific antibodies are observed in patients with VL and other severe forms of leishmanial disease and there are accumulating evidence that B cells and antibodies correlate with pathology.
A model where immunoglobulin (IgG) promotes infection by inducing IL-10 was proposed by Kane and Mosser, who showed that IgG coated amastigotes (L. major) could ligate Fc-receptors on murine macrophages and induce IL-10 production.[67] In support of this models in vivo studies found that Fc-deficient mice infected with L. amazonensis produce less IL-10 and are less susceptible to infection.[93] Moreover, a regulatory role for B cells has been suggested in a VL model demonstrating that B cell depleted animal exhibit extensive neutrophil mediated pathology.[23]
There is still much to learn about how antibodies function in leishmaniasis and it should not be ruled out that certain antibodies might contribute to protection. Immunization with, the for dogs licensed vaccine, Leishmune, which confers some protection against leishmaniasis, result in seroconversion and an increase in the proportion of B cells.[94]
IMMUNE RESPONSES ASSOCIATED WITH VISCERAL DISEASE
Infection with L. donovani and Leishmania infantum results in the establishment of the parasite in the liver, spleen and bone marrow in mice. The liver is, in most mouse strains, the site of an acute resolving infection associated with the development of inflammatory granulomas around infected Kupffer cells, and resistance to re-infection. In contrast to the liver, parasites persist in the spleen. Persistent parasites are characterized by lack of granuloma formation, splenomegaly, enhanced hematopoietic activity and disruption of lymphoid tissue micro-architecture, the latter postulated to contribute to the immuno-compromised status of the host. Splenic pathology is linked to high levels of both TNFα and IL-10. TNFα mediates destruction of marginal zone macrophages and gp38+stromal cells, while IL-10 is responsible for impaired DC migration into T cell areas and defective T cell priming. Furthermore, the altered stromal cells function can promote development of IL-10 producing DC, with immuno-regulatory properties (reviewed in[95]).
The mechanisms underlying the failure to control growth and spread of parasites in human VL are less well understood. Most humans infected with visceralizing Leishmania species never develop disease. How control is mediated in these people is largely unknown. Absence of antigen specific Th1 responses, in peripheral blood mononuclear cells of VL patients is, thought to be, causally related to disease progression. However, there appears to be no inherent defect in antigen induced Th1 response as patient cell respond after cure and the finding that patients express elevated levels of IFNγ mRNA in T cells, in lesional tissue and the multiple pro-inflammatory cytokines, found systemically and at the site of infection indicates that neither can their immunological defect simply be explained by immune tolerance or Th2 polarization. Most clinical evidence point to that the host regulatory response, generated to limit collateral tissue damage, promotes spread and growth of microorganism in chronic infections, like VL. Human VL is clearly associated with elevated levels of the regulatory cytokines IL-10.[64] The inhibitory effect of IL-10 on macrophage may be detrimental, as IL-10 renders macrophages unresponsive to activating cytokines. In addition IL-10 may contribute to loss of important effector T cells in VL. It is still unclear if patients with VL have developed antigen specific T cell responses that are suppressed by regulatory responses or if antigen specific T cell responses are never appropriately generated.
THE QUEST FOR A VACCINE AGAINST LEISHMANIASIS
After clinical cure from leishmanial disease people are considered to have acquired lifelong immunity to infection with the same parasite, making vaccination a feasible measure. However, developing a human vaccine against Leishmania has proven difficult despite the many vaccine candidates reported to be protective against murine leishmaniasis. In humans, vaccination with autoclaved L. major (considered protective in mice) adjuvanted with BCG has had limited success.[96] The only reliable preventive measure against clinical disease remains leishmanzation (LZ), an ancient method where viable parasites are inoculated at a covered part of the body to protect against subsequent disfiguring disease. While this method may be used to protect against mild cutaneous disease, it cannot be used against VL. Moreover, LZ has been questioned due to fear of non-healing and disseminating ulcers.
Vaccination in mice may, however, not be as straight forward as initially thought. It was recently shown that inoculation of killed Leishmania in mice that resolved their primary L. major infection resulted in rapid and relatively sustained loss of infection-induced immunity.[97] Moreover, while vaccination with killed parasites plus CpG adjuvant confer protection against needle challenge, vector transmitted infection can abrogate this protective immunity. Only live infection was able to protect against subsequent vector mediated transmission.[98] Indeed, studies indicate that parasite persistence may be required for maintenance of protective T cell immunity and “sterile” cure would again make the animal permissive for disease.[61,99]
Vaccination with attenuated parasites may be the solution for preventing leishmanial disease. Several methods have been used to develop live attenuated Leishmania parasites including long term in vitro cultures, selection for temperature sensitivity, chemical mutagenesis and irradiation.[100] Attenuated lines have been shown to confer substantial protection in animal models, but undefined mutations and concerns regarding conversion back to virulence make them unsuitable for human use. Targeted elimination of virulence or essential genes could, if carefully done, solve this problem. Recently, a L. donovani strain completely deficient in the centrin gene, (required for growth in the amastigote stage) was found to be both safe and protective against both homologous and heterologous parasites in rodent models.[101] While more studies are required this may be an attractive vaccine candidate against VL.
COINFECTION – A REALITY RARELY CONSIDERED IN EXPERIMENTAL MODELS
The poor populations, which are mainly affected by leishmaniasis, are also plagued with many other chronic infections such as helminths, other protozoa, tuberculosis and HIV/AIDS.
The visceralizing species of Leishmania can in many aspects be considered as opportunistic infections, and patients with HIV are at much greater risk of developing VL.[102] From an immunological perspective the two agents ability to escape and manipulate the immune response seem to a large extent work in synergy, resulting in a dangerous liaison where the immune system rapidly can be exhausted and control of pathogens lost.[103–105] HIV/AIDS can also slow down diagnosis of VL as antibody based tests may not be indicative of disease in AIDS patients. Interestingly, while HIV may have a profound impact on VL the evolution of purely cutaneous disease would only seem to be only moderately affected and L. major have not been reported to visceralize in HIV patients.[102]
Helminths are another group of pathogens, which may favor survival of Leishmania parasites by the Th2 and regulatory immune responses they induce. In this respect, it has been shown that mice have better capacity to deal with L. donovani and L. major infection in the absence of Schistosoma mansoni infection.[106,107]
Coinfection are a reality that need to be considered when developing and evaluating vaccines against human leishmaniais in endemic populations. Treating co-infections, in particular worms may be a measure to enhance vaccine and therapeutic efficacy.
Acknowledgments
We would like to thank Hannah Akuffo for critical reading of the manuscript.
Footnotes
Source of Support: Financial support for Susanne Nylén and Shalini Gautam has been provided by SIDA/Sarec and the Indian Research Council, respectively.
Conflict of Interest: None declared.
REFERENCES
- 1.Clark RA, Chong B, Mirchandani N, Brinster NK, Yamanaka K, Dowgiert RK, et al. The vast majority of CLA+ T cells are resident in normal skin. J Immunol. 2006;176:4431–9. doi: 10.4049/jimmunol.176.7.4431. [DOI] [PubMed] [Google Scholar]
- 2.Sacks DL, Perkins PV. Identification of an infective stage of Leishmania promastigotes. Science. 1984;223:1417–9. doi: 10.1126/science.6701528. [DOI] [PubMed] [Google Scholar]
- 3.Ueno N, Bratt CL, Rodriguez NE, Wilson ME. Differences in human macrophage receptor usage, lysosomal fusion kinetics and survival between logarithmic and metacyclic Leishmania infantum chagasi promastigotes. Cell Microbiol. 2009;11:1827–41. doi: 10.1111/j.1462-5822.2009.01374.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Moreno I, Molina R, Torano A, Laurin E, Garcia E, Dominguez M. Comparative real-time kinetic analysis of human complement killing of Leishmania infantum promastigotes derived from axenic culture or from Phlebotomus perniciosus. Microbes Infect. 2007;9:1574–80. doi: 10.1016/j.micinf.2007.09.009. [DOI] [PubMed] [Google Scholar]
- 5.Wozencraft AO, Blackwell JM. Increased infectivity of stationary-phase promastigotes of Leishmania donovani: correlation with enhanced C3 binding capacity and CR3-mediated attachment to host macrophages. Immunology. 1987;60:559–63. [PMC free article] [PubMed] [Google Scholar]
- 6.Locksley RM, Heinzel FP, Fankhauser JE, Nelson CS, Sadick MD. Cutaneous host defense in leishmaniasis: interaction of isolated dermal macrophages and epidermal Langerhans cells with the insect-stage promastigote. Infect Immun. 1988;56:336–42. doi: 10.1128/iai.56.2.336-342.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Leon B, Lopez-Bravo M, Ardavin C. Monocyte-derived dendritic cells formed at the infection site control the induction of protective T helper 1 responses against Leishmania. Immunity. 2007;26:519–31. doi: 10.1016/j.immuni.2007.01.017. [DOI] [PubMed] [Google Scholar]
- 8.Peters NC, Egen JG, Secundino N, Debrabant A, Kimblin N, Kamhawi S, et al. In vivo imaging reveals an essential role for neutrophils in leishmaniasis transmitted by sand flies. Science. 2008;321:970–4. doi: 10.1126/science.1159194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Pahwa R, Gupta SK, Singh T, Nigam S. Acute fulminant visceral leishmaniasis in children--a report of two cases. Indian J Pathol Microbiol. 2004;47:428–30. [PubMed] [Google Scholar]
- 10.Wickramasinghe SN, Abdalla SH, Kasili EG. Ultrastructure of bone marrow in patients with visceral leishmaniasis. J Clin Pathol. 1987;40:267–75. doi: 10.1136/jcp.40.3.267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ganz T. Defensins: antimicrobial peptides of innate immunity. Nat Rev Immunol. 2003;3:710–20. doi: 10.1038/nri1180. [DOI] [PubMed] [Google Scholar]
- 12.Laskay T, van Zandbergen G, Solbach W. Neutrophil granulocytes as host cells and transport vehicles for intracellular pathogens: apoptosis as infection-promoting factor. Immunobiology. 2008;213:183–91. doi: 10.1016/j.imbio.2007.11.010. [DOI] [PubMed] [Google Scholar]
- 13.van Zandbergen G, Klinger M, Mueller A, Dannenberg S, Gebert A, Solbach W, et al. Cutting edge: neutrophil granulocyte serves as a vector for Leishmania entry into macrophages. J Immunol. 2004;173:6521–5. doi: 10.4049/jimmunol.173.11.6521. [DOI] [PubMed] [Google Scholar]
- 14.Pearson RD, Steigbigel RT. Phagocytosis and killing of the protozoan Leishmania donovani by human polymorphonuclear leukocytes. J Immunol. 1981;127:1438–43. [PubMed] [Google Scholar]
- 15.Lima GM, Vallochi AL, Silva UR, Bevilacqua EM, Kiffer MM, Abrahamsohn IA. The role of polymorphonuclear leukocytes in the resistance to cutaneous Leishmaniasis. Immunol Lett. 1998;64:145–51. doi: 10.1016/s0165-2478(98)00099-6. [DOI] [PubMed] [Google Scholar]
- 16.McFarlane E, Perez C, Charmoy M, Allenbach C, Carter KC, Alexander J, et al. Neutrophils contribute to development of a protective immune response during onset of infection with Leishmania donovani. Infect Immun. 2008;76:532–41. doi: 10.1128/IAI.01388-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rousseau D, Demartino S, Ferrua B, Michiels JF, Anjuere F, Fragaki K, et al. In vivo involvement of polymorphonuclear neutrophils in Leishmania infantum infection. BMC Microbiol. 2001;1:17. doi: 10.1186/1471-2180-1-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Guimaraes-Costa AB, Nascimento MT, Froment GS, Soares RP, Morgado FN, Conceicao-Silva F, et al. Leishmania amazonensis promastigotes induce and are killed by neutrophil extracellular traps. Proc Natl Acad Sci U S A. 2009;106:6748–53. doi: 10.1073/pnas.0900226106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ritter U, Frischknecht F, van Zandbergen G. Are neutrophils important host cells for Leishmania parasites? Trends Parasitol. 2009;25:505–10. doi: 10.1016/j.pt.2009.08.003. [DOI] [PubMed] [Google Scholar]
- 20.Donnelly KB, Lima HC, Titus RG. Histologic characterization of experimental cutaneous leishmaniasis in mice infected with Leishmania braziliensis in the presence or absence of sand fly vector salivary gland lysate. J Parasitol. 1998;84:97–103. [PubMed] [Google Scholar]
- 21.Palma GI, Saravia NG. In situ characterization of the human host response to Leishmania panamensis. Am J Dermatopathol. 1997;19:585–90. doi: 10.1097/00000372-199712000-00006. [DOI] [PubMed] [Google Scholar]
- 22.Lopez Kostka S, Dinges S, Griewank K, Iwakura Y, Udey MC, von Stebut E. IL-17 promotes progression of cutaneous leishmaniasis in susceptible mice. J Immunol. 2009;182:3039–46. doi: 10.4049/jimmunol.0713598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Smelt SC, Cotterell SE, Engwerda CR, Kaye PM. B cell-deficient mice are highly resistant to Leishmania donovani infection, but develop neutrophil-mediated tissue pathology. J Immunol. 2000;164:3681–8. doi: 10.4049/jimmunol.164.7.3681. [DOI] [PubMed] [Google Scholar]
- 24.Rosenthal LA, Sutterwala FS, Kehrli ME, Mosser DM. Leishmania major-human macrophage interactions: cooperation between Mac-1 (CD11b/CD18) and complement receptor type 1 (CD35) in promastigote adhesion. Infect Immun. 1996;64:2206–15. doi: 10.1128/iai.64.6.2206-2215.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wanderley JL, Moreira ME, Benjamin A, Bonomo AC, Barcinski MA. Mimicry of apoptotic cells by exposing phosphatidylserine participates in the establishment of amastigotes of Leishmania (L) amazonensis in mammalian hosts. J Immunol. 2006;176:1834–9. doi: 10.4049/jimmunol.176.3.1834. [DOI] [PubMed] [Google Scholar]
- 26.McDowell MA, Sacks DL. Inhibition of host cell signal transduction by Leishmania: observations relevant to the selective impairment of IL-12 responses. Curr Opin Microbiol. 1999;2:438–43. doi: 10.1016/S1369-5274(99)80077-0. [DOI] [PubMed] [Google Scholar]
- 27.Kima PE. The amastigote forms of Leishmania are experts at exploiting host cell processes to establish infection and persist. Int J Parasitol. 2007;37:1087–96. doi: 10.1016/j.ijpara.2007.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.De Trez C, Magez S, Akira S, Ryffel B, Carlier Y, Muraille E. iNOS-producing inflammatory dendritic cells constitute the major infected cell type during the chronic Leishmania major infection phase of C57BL/6 resistant mice. PLoS Pathog. 2009;5:e1000494. doi: 10.1371/journal.ppat.1000494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Filipe-Santos O, Pescher P, Breart B, Lippuner C, Aebischer T, Glaichenhaus N, et al. A dynamic map of antigen recognition by CD4 T cells at the site of Leishmania major infection. Cell Host Microbe. 2009;6:23–33. doi: 10.1016/j.chom.2009.04.014. [DOI] [PubMed] [Google Scholar]
- 30.Soong L. Modulation of dendritic cell function by Leishmania parasites. J Immunol. 2008;180:4355–60. doi: 10.4049/jimmunol.180.7.4355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Moll H, Fuchs H, Blank C, Rollinghoff M. Langerhans cells transport Leishmania major from the infected skin to the draining lymph node for presentation to antigen-specific T cells. Eur J Immunol. 1993;23:1595–601. doi: 10.1002/eji.1830230730. [DOI] [PubMed] [Google Scholar]
- 32.Ng LG, Hsu A, Mandell MA, Roediger B, Hoeller C, Mrass P, et al. Migratory dermal dendritic cells act as rapid sensors of protozoan parasites. PLoS Pathog. 2008;4:e1000222. doi: 10.1371/journal.ppat.1000222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ritter U, Meissner A, Scheidig C, Korner H. CD8 alpha- and Langerin-negative dendritic cells, but not Langerhans cells, act as principal antigen-presenting cells in leishmaniasis. Eur J Immunol. 2004;34:1542–50. doi: 10.1002/eji.200324586. [DOI] [PubMed] [Google Scholar]
- 34.Iezzi G, Frohlich A, Ernst B, Ampenberger F, Saeland S, Glaichenhaus N, et al. Lymph node resident rather than skin-derived dendritic cells initiate specific T cell responses after Leishmania major infection. J Immunol. 2006;177:1250–6. doi: 10.4049/jimmunol.177.2.1250. [DOI] [PubMed] [Google Scholar]
- 35.Tejle K, Lindroth M, Magnusson KE, Rasmusson B. Wild-type Leishmania donovani promastigotes block maturation, increase integrin expression and inhibit detachment of human monocyte-derived dendritic cells--the influence of phosphoglycans. FEMS Microbiol Lett. 2008;279:92–102. doi: 10.1111/j.1574-6968.2007.01013.x. [DOI] [PubMed] [Google Scholar]
- 36.McDowell MA, Marovich M, Lira R, Braun M, Sacks D. Leishmania priming of human dendritic cells for CD40 ligand-induced interleukin-12p70 secretion is strain and species dependent. Infect Immun. 2002;70:3994–4001. doi: 10.1128/IAI.70.8.3994-4001.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Laurenti MD, Gidlund M, Ura DM, Sinhorini IL, Corbett CE, Goto H. The role of natural killer cells in the early period of infection in murine cutaneous leishmaniasis. Braz J Med Biol Res. 1999;32:323–5. doi: 10.1590/s0100-879x1999000300012. [DOI] [PubMed] [Google Scholar]
- 38.Nylen S, Maasho K, Soderstrom K, Ilg T, Akuffo H. Live Leishmania promastigotes can directly activate primary human natural killer cells to produce interferon-gamma. Clin Exp Immunol. 2003;131:457–67. doi: 10.1046/j.1365-2249.2003.02096.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Becker I, Salaiza N, Aguirre M, Delgado J, Carrillo-Carrasco N, Kobeh LG, et al. Leishmania lipophosphoglycan (LPG) activates NK cells through toll-like receptor-2. Mol Biochem Parasitol. 2003;130:65–74. doi: 10.1016/s0166-6851(03)00160-9. [DOI] [PubMed] [Google Scholar]
- 40.Piccioli D, Sbrana S, Melandri E, Valiante NM. Contact-dependent stimulation and inhibition of dendritic cells by natural killer cells. J Exp Med. 2002;195:335–41. doi: 10.1084/jem.20010934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sanabria MX, Vargas-Inchaustegui DA, Xin L, Soong L. Role of natural killer cells in modulating dendritic cell responses to Leishmania amazonensis infection. Infect Immun. 2008;76:5100–9. doi: 10.1128/IAI.00438-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bajenoff M, Breart B, Huang AY, Qi H, Cazareth J, Braud VM, et al. Natural killer cell behavior in lymph nodes revealed by static and real-time imaging. J Exp Med. 2006;203:619–31. doi: 10.1084/jem.20051474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Maasho K, Sanchez F, Schurr E, Hailu A, Akuffo H. Indications of the protective role of natural killer cells in human cutaneous leishmaniasis in an area of endemicity. Infect Immun. 1998;66:2698–704. doi: 10.1128/iai.66.6.2698-2704.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Nylen S, Maurya R, Eidsmo L, Manandhar KD, Sundar S, Sacks D. Splenic accumulation of IL-10 mRNA in T cells distinct from CD4+CD25+ (Foxp3) regulatory T cells in human visceral leishmaniasis. J Exp Med. 2007;204:805–17. doi: 10.1084/jem.20061141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Pereira LI, Dorta ML, Pereira AJ, Bastos RP, Oliveira MA, Pinto SA, et al. Increase of NK cells and proinflammatory monocytes are associated with the clinical improvement of diffuse cutaneous leishmaniasis after immunochemotherapy with BCG/Leishmania antigens. Am J Trop Med Hyg. 2009;81:378–83. [PubMed] [Google Scholar]
- 46.Murray HW, Nathan CF. Macrophage microbicidal mechanisms in vivo: reactive nitrogen versus oxygen intermediates in the killing of intracellular visceral Leishmania donovani. J Exp Med. 1999;189:741–6. doi: 10.1084/jem.189.4.741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wei XQ, Charles IG, Smith A, Ure J, Feng GJ, Huang FP, et al. Altered immune responses in mice lacking inducible nitric oxide synthase. Nature. 1995;375:408–11. doi: 10.1038/375408a0. [DOI] [PubMed] [Google Scholar]
- 48.Sacks D, Noben-Trauth N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat Rev Immunol. 2002;2:845–58. doi: 10.1038/nri933. [DOI] [PubMed] [Google Scholar]
- 49.Harms G, Zwingenberger K, Chehade AK, Talhari S, Racz P, Mouakeh A, et al. Effects of intradermal gamma-interferon in cutaneous leishmaniasis. Lancet. 1989;1:1287–92. doi: 10.1016/s0140-6736(89)92686-x. [DOI] [PubMed] [Google Scholar]
- 50.Anderson CF, Mendez S, Sacks DL. Nonhealing infection despite Th1 polarization produced by a strain of Leishmania major in C57BL/6 mice. J Immunol. 2005;174:2934–41. doi: 10.4049/jimmunol.174.5.2934. [DOI] [PubMed] [Google Scholar]
- 51.Sundar S, Rosenkaimer F, Lesser ML, Murray HW. Immunochemotherapy for a systemic intracellular infection: accelerated response using interferon-gamma in visceral leishmaniasis. J Infect Dis. 1995;171:992–6. doi: 10.1093/infdis/171.4.992. [DOI] [PubMed] [Google Scholar]
- 52.Nylen S, Khamesipour A, Mohammadi A, Jafari-Shakib R, Eidsmo L, Noazin S, et al. Surrogate markers of immunity to Leishmania major in leishmanin skin test negative individuals from an endemic area re-visited. Vaccine. 2006;24:6944–54. doi: 10.1016/j.vaccine.2006.05.016. [DOI] [PubMed] [Google Scholar]
- 53.Lee YK, Mukasa R, Hatton RD, Weaver CT. Developmental plasticity of Th17 and Treg cells. Curr Opin Immunol. 2009;21:274–80. doi: 10.1016/j.coi.2009.05.021. [DOI] [PubMed] [Google Scholar]
- 54.Korn T, Bettelli E, Oukka M, Kuchroo VK. IL-17 and Th17 Cells. Annu Rev Immunol. 2009;27:485–517. doi: 10.1146/annurev.immunol.021908.132710. [DOI] [PubMed] [Google Scholar]
- 55.Pitta MG, Romano A, Cabantous S, Henri S, Hammad A, Kouriba B, et al. IL-17 and IL-22 are associated with protection against human kala azar caused by Leishmania donovani. J Clin Invest. 2009;119:2379–87. doi: 10.1172/JCI38813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Artis D, Johnson LM, Joyce K, Saris C, Villarino A, Hunter CA, et al. Cutting edge: early IL-4 production governs the requirement for IL-27-WSX-1 signaling in the development of protective Th1 cytokine responses following Leishmania major infection. J Immunol. 2004;172:4672–5. doi: 10.4049/jimmunol.172.8.4672. [DOI] [PubMed] [Google Scholar]
- 57.Anderson CF, Stumhofer JS, Hunter CA, Sacks D. IL-27 regulates IL-10 and IL-17 from CD4+ cells in nonhealing Leishmania major infection. J Immunol. 2009;183:4619–27. doi: 10.4049/jimmunol.0804024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Murugaiyan G, Mittal A, Lopez-Diego R, Maier LM, Anderson DE, Weiner HL. IL-27 is a key regulator of IL-10 and IL-17 production by human CD4+ T cells. J Immunol. 2009;183:2435–43. doi: 10.4049/jimmunol.0900568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Rosas LE, Satoskar AA, Roth KM, Keiser TL, Barbi J, Hunter C, et al. Interleukin-27R (WSX-1/T-cell cytokine receptor) gene-deficient mice display enhanced resistance to leishmania donovani infection but develop severe liver immunopathology. Am J Pathol. 2006;168:158–69. doi: 10.2353/ajpath.2006.050013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Ansari NA, Kumar R, Gautam S. In: Evidence and implications of IL-27/IL-17 pathway in human visceral leishmaniasis. Woods H, editor. WHIP; 2009. [Google Scholar]
- 61.Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, Sacks DL. CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature. 2002;420:502–7. doi: 10.1038/nature01152. [DOI] [PubMed] [Google Scholar]
- 62.Campanelli AP, Roselino AM, Cavassani KA, Pereira MS, Mortara RA, Brodskyn CI, et al. CD4+CD25+ T cells in skin lesions of patients with cutaneous leishmaniasis exhibit phenotypic and functional characteristics of natural regulatory T cells. J Infect Dis. 2006;193:1313–22. doi: 10.1086/502980. [DOI] [PubMed] [Google Scholar]
- 63.Bourreau E, Ronet C, Darcissac E, Lise MC, Sainte Marie D, Clity E, et al. Intralesional regulatory T-cell suppressive function during human acute and chronic cutaneous leishmaniasis due to Leishmania guyanensis. Infect Immun. 2009;77:1465–74. doi: 10.1128/IAI.01398-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Nylen S, Sacks D. Interleukin-10 and the pathogenesis of human visceral leishmaniasis. Trends Immunol. 2007;28:378–84. doi: 10.1016/j.it.2007.07.004. [DOI] [PubMed] [Google Scholar]
- 65.Anderson CF, Oukka M, Kuchroo VJ, Sacks D. CD4(+)CD25(−)Foxp3(−) Th1 cells are the source of IL-10-mediated immune suppression in chronic cutaneous leishmaniasis. J Exp Med. 2007;204:285–97. doi: 10.1084/jem.20061886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Murphy ML, Wille U, Villegas EN, Hunter CA, Farrell JP. IL-10 mediates susceptibility to Leishmania donovani infection. Eur J Immunol. 2001;31:2848–56. doi: 10.1002/1521-4141(2001010)31:10<2848::aid-immu2848>3.0.co;2-t. [DOI] [PubMed] [Google Scholar]
- 67.Kane MM, Mosser DM. The role of IL-10 in promoting disease progression in leishmaniasis. J Immunol. 2001;166:1141–7. doi: 10.4049/jimmunol.166.2.1141. [DOI] [PubMed] [Google Scholar]
- 68.Carvalho EM, Bacellar O, Brownell C, Regis T, Coffman RL, Reed SG. Restoration of IFN-gamma production and lymphocyte proliferation in visceral leishmaniasis. J Immunol. 1994;152:5949–56. [PubMed] [Google Scholar]
- 69.Ghalib HW, Piuvezam MR, Skeiky YA, Siddig M, Hashim FA, el-Hassan AM, et al. Interleukin 10 production correlates with pathology in human Leishmania donovani infections. J Clin Invest. 1993;92:324–9. doi: 10.1172/JCI116570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Bourreau E, Prevot G, Gardon J, Pradinaud R, Launois P. High intralesional interleukin-10 messenger RNA expression in localized cutaneous leishmaniasis is associated with unresponsiveness to treatment. J Infect Dis. 2001;184:1628–30. doi: 10.1086/324665. [DOI] [PubMed] [Google Scholar]
- 71.Bourreau E, Ronet C, Darsissac E, Lise MC, Marie DS, Clity E, et al. In leishmaniasis due to Leishmania guyanensis infection, distinct intralesional interleukin-10 and Foxp3 mRNA expression are associated with unresponsiveness to treatment. J Infect Dis. 2009;199:576–9. doi: 10.1086/596508. [DOI] [PubMed] [Google Scholar]
- 72.Salhi A, Rodrigues V, Jr, Santoro F, Dessein H, Romano A, Castellano LR, et al. Immunological and genetic evidence for a crucial role of IL-10 in cutaneous lesions in humans infected with Leishmania braziliensis. J Immunol. 2008;180:6139–48. doi: 10.4049/jimmunol.180.9.6139. [DOI] [PubMed] [Google Scholar]
- 73.Farrell JP, Muller I, Louis JA. A role for Lyt-2+ T cells in resistance to cutaneous leishmaniasis in immunized mice. J Immunol. 1989;142:2052–6. [PubMed] [Google Scholar]
- 74.Holaday BJ, Sadick MD, Wang ZE, Reiner SL, Heinzel FP, Parslow TG, et al. Reconstitution of Leishmania immunity in severe combined immunodeficient mice using Th1- and Th2-like cell lines. J Immunol. 1991;147:1653–8. [PubMed] [Google Scholar]
- 75.Wang ZE, Reiner SL, Hatam F, Heinzel FP, Bouvier J, Turck CW, et al. Targeted activation of CD8 cells and infection of beta 2-microglobulin-deficient mice fail to confirm a primary protective role for CD8 cells in experimental leishmaniasis. J Immunol. 1993;151:2077–86. [PubMed] [Google Scholar]
- 76.Belkaid Y, Von Stebut E, Mendez S, Lira R, Caler E, Bertholet S, et al. CD8+ T cells are required for primary immunity in C57BL/6 mice following low-dose, intradermal challenge with Leishmania major. J Immunol. 2002;168:3992–4000. doi: 10.4049/jimmunol.168.8.3992. [DOI] [PubMed] [Google Scholar]
- 77.Murray HW. Prevention of relapse after chemotherapy in a chronic intracellular infection: mechanisms in experimental visceral leishmaniasis. J Immunol. 2005;174:4916–23. doi: 10.4049/jimmunol.174.8.4916. [DOI] [PubMed] [Google Scholar]
- 78.Ruiz JH, Becker I. CD8 cytotoxic T cells in cutaneous leishmaniasis. Parasite Immunol. 2007;29:671–8. doi: 10.1111/j.1365-3024.2007.00991.x. [DOI] [PubMed] [Google Scholar]
- 79.Colmenares M, Kima PE, Samoff E, Soong L, McMahon-Pratt D. Perforin and gamma interferon are critical CD8+ T-cell-mediated responses in vaccine-induced immunity against Leishmania amazonensis infection. Infect Immun. 2003;71:3172–82. doi: 10.1128/IAI.71.6.3172-3182.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Brodskyn CI, Barral A, Boaventura V, Carvalho E, Barral-Netto M. Parasite-driven in vitro human lymphocyte cytotoxicity against autologous infected macrophages from mucosal leishmaniasis. J Immunol. 1997;159:4467–73. [PubMed] [Google Scholar]
- 81.Kima PE, Ruddle NH, McMahon-Pratt D. Presentation via the class I pathway by Leishmania amazonensis-infected macrophages of an endogenous leishmanial antigen to CD8+ T cells. J Immunol. 1997;159:1828–34. [PubMed] [Google Scholar]
- 82.Huang FP, Xu D, Esfandiari EO, Sands W, Wei XQ, Liew FY. Mice defective in Fas are highly susceptible to Leishmania major infection despite elevated IL-12 synthesis, strong Th1 responses, and enhanced nitric oxide production. J Immunol. 1998;160:4143–7. [PubMed] [Google Scholar]
- 83.Alexander CE, Kaye PM, Engwerda CR. CD95 is required for the early control of parasite burden in the liver of Leishmania donovani-infected mice. Eur J Immunol. 2001;31:1199–210. doi: 10.1002/1521-4141(200104)31:4<1199::aid-immu1199>3.0.co;2-6. [DOI] [PubMed] [Google Scholar]
- 84.Eidsmo L, Nylen S, Khamesipour A, Hedblad MA, Chiodi F, Akuffo H. The contribution of the Fas/FasL apoptotic pathway in ulcer formation during Leishmania major-induced cutaneous Leishmaniasis. Am J Pathol. 2005;166:1099–108. doi: 10.1016/S0002-9440(10)62330-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Bomfim G, Andrade BB, Santos S, Clarencio J, Barral-Netto M, Barral A. Cellular analysis of cutaneous leishmaniasis lymphadenopathy: insights into the early phases of human disease. Am J Trop Med Hyg. 2007;77:854–9. [PubMed] [Google Scholar]
- 86.Coutinho SG, Da-Cruz AM, Bertho AL, Santiago MA, De-Luca P. Immunologic patterns associated with cure in human American cutaneous leishmaniasis. Braz J Med Biol Res. 1998;31:139–42. doi: 10.1590/s0100-879x1998000100019. [DOI] [PubMed] [Google Scholar]
- 87.Da-Cruz AM, Conceicao-Silva F, Bertho AL, Coutinho SG. Leishmania-reactive CD4+ and CD8+ T cells associated with cure of human cutaneous leishmaniasis. Infect Immun. 1994;62:2614–8. doi: 10.1128/iai.62.6.2614-2618.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Faria DR, Souza PE, Duraes FV, Carvalho EM, Gollob KJ, Machado PR, et al. Recruitment of CD8(+) T cells expressing granzyme A is associated with lesion progression in human cutaneous leishmaniasis. Parasite Immunol. 2009;31:432–9. doi: 10.1111/j.1365-3024.2009.01125.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Tuon FF, Gomes-Silva A, Da-Cruz AM, Duarte MI, Neto VA, Amato VS. Local immunological factors associated with recurrence of mucosal leishmaniasis. Clin Immunol. 2008;128:442–6. doi: 10.1016/j.clim.2008.05.007. [DOI] [PubMed] [Google Scholar]
- 90.Ghosh MK, Nandy A, Addy M, Maitra TK, Ghose AC. Subpopulations of T lymphocytes in the peripheral blood, dermal lesions and lymph nodes of post kala-azar dermal leishmaniasis patients. Scand J Immunol. 1995;41:11–7. doi: 10.1111/j.1365-3083.1995.tb03527.x. [DOI] [PubMed] [Google Scholar]
- 91.Smith TR, Kumar V. Revival of CD8+ Treg-mediated suppression. Trends Immunol. 2008;29:337–42. doi: 10.1016/j.it.2008.04.002. [DOI] [PubMed] [Google Scholar]
- 92.Bourreau E, Ronet C, Couppie P, Sainte-Marie D, Tacchini-Cottier F, Launois P. IL-10 producing CD8+ T cells in human infection with Leishmania guyanensis. Microbes Infect. 2007;9:1034–41. doi: 10.1016/j.micinf.2007.04.009. [DOI] [PubMed] [Google Scholar]
- 93.Buxbaum LU, Scott P. Interleukin 10- and Fcgamma receptor-deficient mice resolve Leishmania mexicana lesions. Infect Immun. 2005;73:2101–8. doi: 10.1128/IAI.73.4.2101-2108.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Borja-Cabrera GP, Santos FN, Bauer FS, Parra LE, Menz I, Morgado AA, et al. Immunogenicity assay of the Leishmune vaccine against canine visceral leishmaniasis in Brazil. Vaccine. 2008;26:4991–7. doi: 10.1016/j.vaccine.2008.07.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Kaye PM, Svensson M, Ato M, Maroof A, Polley R, Stager S, et al. The immunopathology of experimental visceral leishmaniasis. Immunol Rev. 2004;201:239–53. doi: 10.1111/j.0105-2896.2004.00188.x. [DOI] [PubMed] [Google Scholar]
- 96.Noazin S, Modabber F, Khamesipour A, Smith PG, Moulton LH, Nasseri K, Sharifi I, Khalil EA, Bernal ID, Antunes CM, Kieny MP, Tanner M. First generation leishmaniasis vaccines: A review of field efficacy trials. Vaccine. 2008;26:6759–67. doi: 10.1016/j.vaccine.2008.09.085. [DOI] [PubMed] [Google Scholar]
- 97.Okwor I, Liu D, Beverley SM, Uzonna JE. Inoculation of killed Leishmania major into immune mice rapidly disrupts immunity to a secondary challenge via IL-10-mediated process. Proc Natl Acad Sci U S A. 2009;106:13951–6. doi: 10.1073/pnas.0905184106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Peters NC, Kimblin N, Secundino N, Kamhawi S, Lawyer P, Sacks DL. Vector transmission of leishmania abrogates vaccine-induced protective immunity. PLoS Pathog. 2009;5:e1000484. doi: 10.1371/journal.ppat.1000484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Belkaid Y, Hoffmann KF, Mendez S, Kamhawi S, Udey MC, Wynn TA, et al. The role of interleukin (IL)-10 in the persistence of Leishmania major in the skin after healing and the therapeutic potential of anti-IL-10 receptor antibody for sterile cure. J Exp Med. 2001;194:1497–506. doi: 10.1084/jem.194.10.1497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Selvapandiyan A, Duncan R, Debrabant A, Lee N, Sreenivas G, Salotra P, et al. Genetically modified live attenuated parasites as vaccines for leishmaniasis. Indian J Med Res. 2006;123:455–66. [PubMed] [Google Scholar]
- 101.Selvapandiyan A, Dey R, Nylen S, Duncan R, Sacks D, Nakhasi HL. Intracellular replication-deficient Leishmania donovani induces long lasting protective immunity against visceral leishmaniasis. J Immunol. 2009;183:1813–20. doi: 10.4049/jimmunol.0900276. [DOI] [PubMed] [Google Scholar]
- 102.Alvar J, Aparicio P, Aseffa A, Den Boer M, Canavate C, Dedet JP, et al. The relationship between leishmaniasis and AIDS: the second 10 years. Clin Microbiol Rev. 2008;21:334–59. doi: 10.1128/CMR.00061-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Garg R, Barat C, Ouellet M, Lodge R, Tremblay MJ. Leishmania infantum Amastigotes Enhance HIV-1 Production in Cocultures of Human Dendritic Cells and CD4 T Cells by Inducing Secretion of IL-6 and TNF-alpha. PLoS Negl Trop Dis. 2009;3:e441. doi: 10.1371/journal.pntd.0000441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Garg R, Trudel N, Tremblay MJ. Consequences of the natural propensity of Leishmania and HIV-1 to target dendritic cells. Trends Parasitol. 2007;23:317–24. doi: 10.1016/j.pt.2007.05.006. [DOI] [PubMed] [Google Scholar]
- 105.Wolday D, Berhe N, Akuffo H, Britton S. Leishmania-HIV interaction: immunopathogenic mechanisms. Parasitol Today. 1999;15:182–7. doi: 10.1016/s0169-4758(99)01431-3. [DOI] [PubMed] [Google Scholar]
- 106.Hassan MF, Zhang Y, Engwerda CR, Kaye PM, Sharp H, Bickle QD. The Schistosoma mansoni hepatic egg granuloma provides a favorable microenvironment for sustained growth of Leishmania donovani. Am J Pathol. 2006;169:943–53. doi: 10.2353/ajpath.2006.051319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.La Flamme AC, Scott P, Pearce EJ. Schistosomiasis delays lesion resolution during Leishmania major infection by impairing parasite killing by macrophages. Parasite Immunol. 2002;24:339–45. doi: 10.1046/j.1365-3024.2002.00473.x. [DOI] [PubMed] [Google Scholar]
- 108.van Zandbergen G, Bollinger A, Wenzel A, Kamhawi S, Voll R, Klinger M, et al. Leishmania disease development depends on the presence of apoptotic promastigotes in the virulent inoculum. Proc Natl Acad Sci U S A. 2006;103:13837–42. doi: 10.1073/pnas.0600843103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Wanderley JL, Pinto da Silva LH, Deolindo P, Soong L, Borges VM, Prates DB, et al. Cooperation between apoptotic and viable metacyclics enhances the pathogenesis of Leishmaniasis. PLoS One. 2009;4:e5733. doi: 10.1371/journal.pone.0005733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Nestle FO, Di Meglio P, Qin JZ, Nickoloff BJ. Skin immune sentinels in health and disease. Nat Rev Immunol. 2009;9:679–91. doi: 10.1038/nri2622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Gasim S, Elhassan AM, Khalil EA, Ismail A, Kadaru AM, Kharazmi A, et al. High levels of plasma IL-10 and expression of IL-10 by keratinocytes during visceral leishmaniasis predict subsequent development of post-kala-azar dermal leishmaniasis. Clin Exp Immunol. 1998;111:64–9. doi: 10.1046/j.1365-2249.1998.00468.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Meymandi S, Dabiri S, Dabiri D, Crawford RI, Kharazmi A. A quantitative study of epidermal Langerhans cells in cutaneous leishmaniasis caused by Leishmania tropica. Int J Dermatol. 2004;43:819–23. doi: 10.1111/j.1365-4632.2004.02359.x. [DOI] [PubMed] [Google Scholar]
- 113.Remer KA, Apetrei C, Schwarz T, Linden C, Moll H. Vaccination with plasmacytoid dendritic cells induces protection against infection with Leishmania major in mice. Eur J Immunol. 2007;37:2463–73. doi: 10.1002/eji.200636780. [DOI] [PubMed] [Google Scholar]
- 114.Maurer M, Lopez Kostka S, Siebenhaar F, Moelle K, Metz M, Knop J, et al. Skin mast cells control T cell-dependent host defense in Leishmania major infections. FASEB J. 2006;20:2460–7. doi: 10.1096/fj.06-5860com. [DOI] [PubMed] [Google Scholar]
- 115.Wershil BK, Theodos CM, Galli SJ, Titus RG. Mast cells augment lesion size and persistence during experimental Leishmania major infection in the mouse. J Immunol. 1994;152:4563–71. [PubMed] [Google Scholar]
- 116.Romao PR, Da Costa Santiago H, Ramos CD, De Oliveira CF, Monteiro MC, De Queiroz Cunha F, et al. Mast cell degranulation contributes to susceptibility to Leishmania major. Parasite Immunol. 2009;31:140–6. doi: 10.1111/j.1365-3024.2008.01084.x. [DOI] [PubMed] [Google Scholar]
- 117.Amato VS, Tuon FF, Nicodemo AC, Duarte MI. Histopathology of mast cells and cytokines during healing of human mucosal leishmaniasis. Parasite Immunol. 2008;30:637–40. doi: 10.1111/j.1365-3024.2008.01056.x. [DOI] [PubMed] [Google Scholar]
- 118.Tuon FF, Duarte MI, Amato VS. A quantitative and morphometric study of mast cells in cutaneous leishmaniasis. Parasite Immunol. 2008;30:641–5. doi: 10.1111/j.1365-3024.2008.01057.x. [DOI] [PubMed] [Google Scholar]
- 119.Laskay T, Rollinghoff M, Solbach W. Natural killer cells participate in the early defense against Leishmania major infection in mice. Eur J Immunol. 1993;23:2237–41. doi: 10.1002/eji.1830230928. [DOI] [PubMed] [Google Scholar]
- 120.Ajdary S, Alimohammadian MH, Eslami MB, Kemp K, Kharazmi A. Comparison of the immune profile of nonhealing cutaneous Leishmaniasis patients with those with active lesions and those who have recovered from infection. Infect Immun. 2000;68:1760–4. doi: 10.1128/iai.68.4.1760-1764.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Sundar S, Reed SG, Sharma S, Mehrotra A, Murray HW. Circulating T helper 1 (Th1) cell- and Th2 cell-associated cytokines in Indian patients with visceral leishmaniasis. Am J Trop Med Hyg. 1997;56:522–5. doi: 10.4269/ajtmh.1997.56.522. [DOI] [PubMed] [Google Scholar]
- 122.Akuffo H, Maasho K, Blostedt M, Hojeberg B, Britton S, Bakhiet M. Leishmania aethiopica derived from diffuse leishmaniasis patients preferentially induce mRNA for interleukin-10 while those from localized leishmaniasis patients induce interferon-gamma. J Infect Dis. 1997;175:737–41. doi: 10.1093/infdis/175.3.737. [DOI] [PubMed] [Google Scholar]
- 123.Mattner F, Magram J, Ferrante J, Launois P, Di Padova K, Behin R, et al. Genetically resistant mice lacking interleukin-12 are susceptible to infection with Leishmania major and mount a polarized Th2 cell response. Eur J Immunol. 1996;26:1553–9. doi: 10.1002/eji.1830260722. [DOI] [PubMed] [Google Scholar]
- 124.Bacellar O, Brodskyn C, Guerreiro J, Barral-Netto M, Costa CH, Coffman RL, et al. Interleukin-12 restores interferon-gamma production and cytotoxic responses in visceral leishmaniasis. J Infect Dis. 1996;173:1515–8. doi: 10.1093/infdis/173.6.1515. [DOI] [PubMed] [Google Scholar]
- 125.Ghalib HW, Whittle JA, Kubin M, Hashim FA, el-Hassan AM, Grabstein KH, et al. IL-12 enhances Th1-type responses in human Leishmania donovani infections. J Immunol. 1995;154:4623–9. [PubMed] [Google Scholar]
- 126.Kemp K, Theander TG, Hviid L, Garfar A, Kharazmi A, Kemp M. Interferon-gamma- and tumour necrosis factor-alpha-producing cells in humans who are immune to cutaneous leishmaniasis. Scand J Immunol. 1999;49:655–9. doi: 10.1046/j.1365-3083.1999.00554.x. [DOI] [PubMed] [Google Scholar]
- 127.Wilhelm P, Ritter U, Labbow S, Donhauser N, Rollinghoff M, Bogdan C, et al. Rapidly fatal leishmaniasis in resistant C57BL/6 mice lacking TNF. J Immunol. 2001;166:4012–9. doi: 10.4049/jimmunol.166.6.4012. [DOI] [PubMed] [Google Scholar]
- 128.Engwerda CR, Ato M, Cotterell SE, Mynott TL, Tschannerl A, Gorak-Stolinska PM, et al. A role for tumor necrosis factor-alpha in remodeling the splenic marginal zone during Leishmania donovani infection. Am J Pathol. 2002;161:429–37. doi: 10.1016/s0002-9440(10)64199-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Xynos ID, Tektonidou MG, Pikazis D, Sipsas NV. Leishmaniasis, autoimmune rheumatic disease, and anti-tumor necrosis factor therapy, Europe. Emerg Infect Dis. 2009;15:956–9. doi: 10.3201/eid1506.090101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Gaze ST, Dutra WO, Lessa M, Lessa H, Guimaraes LH, Jesus AR, et al. Mucosal leishmaniasis patients display an activated inflammatory T-cell phenotype associated with a nonbalanced monocyte population. Scand J Immunol. 2006;63:70–8. doi: 10.1111/j.1365-3083.2005.01707.x. [DOI] [PubMed] [Google Scholar]
- 131.Reed SG. TGF-beta in infections and infectious diseases. Microbes Infect. 1999;1:1313–25. doi: 10.1016/s1286-4579(99)00252-x. [DOI] [PubMed] [Google Scholar]
- 132.Laouar Y, Sutterwala FS, Gorelik L, Flavell RA. Transforming growth factor-beta controls T helper type 1 cell development through regulation of natural killer cell interferon-gamma. Nat Immunol. 2005;6:600–7. doi: 10.1038/ni1197. [DOI] [PubMed] [Google Scholar]
- 133.Anderson CF, Lira R, Kamhawi S, Belkaid Y, Wynn TA, Sacks D. IL-10 and TGF-beta control the establishment of persistent and transmissible infections produced by Leishmania tropica in C57BL/6 mice. J Immunol. 2008;180:4090–7. doi: 10.4049/jimmunol.180.6.4090. [DOI] [PubMed] [Google Scholar]
- 134.Saha S, Mondal S, Ravindran R, Bhowmick S, Modak D, Mallick S, et al. IL-10- and TGF-beta-mediated susceptibility in kala-azar and post-kala-azar dermal leishmaniasis: the significance of amphotericin B in the control of Leishmania donovani infection in India. J Immunol. 2007;179:5592–603. doi: 10.4049/jimmunol.179.8.5592. [DOI] [PubMed] [Google Scholar]