

Abstract
The current study used event-related potentials (ERPs) and masked repetition priming to examine the time-course of picture processing. We manipulated the stimulus-onset asynchrony (110 ms, 230 ms, 350 ms, 470 ms) between repeated and unrepeated prime-target pairs while holding the prime duration constant (50 ms) (Experiment 1) as well as the prime duration (30 ms, 50 ms, 70 ms, 90 ms) (Experiment 2) with a constant SOA of 110 ms in a masked repetition priming paradigm with pictures. The aim of this study was to further elucidate the mechanisms underlying previously observed ERP components in masked priming with pictures. We found both the N/P190 and N400 are modulated by changes in prime duration and SOA, however, it appears that longer prime exposure rather than a longer SOA leads to more in-depth processing as indexed by larger N400 effects.
Keywords: event-related potentials (ERPs), masked priming, prime duration, object processing, stimulus-onset asynchrony (SOA)
1. Introduction
As we move through our environment, we rapidly recognize objects without any apparent effort. How we achieve this feat of object recognition with such ease has eluded researchers for decades, as the processes underlying recognition are not well understood. One way the mechanisms underlying object recognition have been studied is through priming. Priming consists of a visible presentation of a prime (word, picture, etc.) that is followed by a target that is either related to/a repetition of the prime or unrelated to the prime. The basic assumption is that presentation of the prime activates certain representations in memory and that the subsequent processing of the related or identical target item, which occurs shortly thereafter, benefits from the overlap of representations between the two items. In the case of unrelated primes and targets, there is little or no overlap in representations and therefore processing of target items is not facilitated. The typical behavioral effect observed in unmasked repetition priming is facilitation in reaction time or accuracy to target objects/word preceded by the same or related prime in comparison to targets preceded by unrelated or unrepeated primes.
Forster and Davis (1984) modified the standard priming paradigm and reduced the visibility of the prime by using a forward mask. They successfully used masked priming with words, where the visibility of the prime is limited, to show the same behavioral repetition effects observed with unmasked primes. The idea behind masked repetition priming is that the briefly presented prime followed by a pattern masked interrupts the processing of the prime prior to conscious identification; thus by examining subtle differences in target stimulus processing that immediately follows the masked prime that is either the same or different stimulus, it should be possible to examine the residual processing influences of the prime on the system. Because participants are not aware of the prime, effects of supraliminal processes are reduced in these masked priming behavioral studies (Forster, Mohan, & Hector, 2003) and masking has been shown to reduce recurrent processing between V1 and higher level cortical areas (Lamme, Zipsser, & Spekreijse, 2002).
Masked priming has been successfully combined with event-related potential (ERP) recordings to provide precise temporal information about the mechanisms involved in recognition (see Eddy, Schmid, & Holcomb, 2006; Holcomb & Grainger, 2006; Eddy & Holcomb, 2009). ERPs are ideal for examining the processes involved in masked priming because of their precise temporal resolution. ERPs provide an advantage over behavioral measures, as they give a measure of brain activity within milliseconds (ms) of stimulus presentation, whereas behavioral responses occur several hundred milliseconds after the presentation of the stimulus. Previous ERP experiments aimed at elucidating these neural mechanisms have examined recognition using masked and unmasked repetition priming with pictures.
There are fewer studies combining these two methodologies with pictures (see Eddy Schmid, & Holcomb, 2006; Eddy & Holcomb, 2009) than with words; however a similar pattern of results has emerged. ERP studies investigating both masked and unmasked picture priming reveal a series of components that occur between 100-500 ms (Eddy & Holcomb, 2009; Eddy, Schmid, & Holcomb, 2006; McPherson & Holcomb, 1999; Holcomb & McPherson, 1994). Even using brief masked prime presentation, robust effects have been observed (Eddy, Schmid & Holcomb, 2006). Masked priming has been widely used in the study of visual word recognition (see Forster et al., 2003; Grainger and Jacobs, 1999). These studies using a short prime duration and a short stimulus onset asynchrony (SOA) have found a series of effects (N150/P150, N250, and N400) (e.g. see Holcomb & Grainger, 2006) reflecting a cascade of processes involved in word recognition. The earliest of these ERP components, the N150/P150 is though to reflect the activation of a perceptual feature representation (N150/P150). Once initial processing of features occurs, a more form specific representation is activated, reflected by the N250 effect. The representation then becomes more abstract, activating semantic, meaning based representation (N400). Some studies only report N400 effects and not earlier ERP components (Misra and Holcomb, 2003; Holcomb et al. 2005). The main difference between studies that describe earlier effects (N150/P150; N250) and those in which only later effects are noted (N400) is the SOA between the prime and target: the experiments yielding only N400 effects were designed with longer SOAs, between 500 to 1000 ms, while those showing earlier effects had SOAs shorter than 500ms (e.g., 70 ms).
To further explore the relationship between SOA and the series of word ERP components reported in the masked and unmasked priming literature, Holcomb and Grainger (2007) systematically examined the effects of SOA and prime duration on masked repetition priming ERP components. Overall, they found no effect of repetition priming at shorter prime durations (10 and 20ms), but the typical cascade of visual word priming effects were observed at longer prime durations (30 and 40ms). They suggest that only a minimal amount of prime exposure is necessary to start the cascade of processing that leads to earlier and later ERP components. Additionally, by manipulating the SOA (60, 180, 300, 420 ms), they were able to examine how masked priming effects with words are affected by increasing the time interval between the prime and target presentation. They found N400 effects for all SOA manipulations; while, the N250 effect was only observed for the 180 and 300 ms SOAs. These observations suggest that the effect of early perceptual processing of word features fades quickly after the presentation of the prime.
There exists a similar disparity in the picture priming literature. Unmasked repetition priming studies that use longer SOAs (e.g., 500-1000 ms) and longer prime durations (e.g., 400 ms) (McPherson and Holcomb, 1999; Holcomb and McPherson, 1994), have found N300 and N400 effects. However, with masked repetition priming, effects begin as early as 100 ms after target stimulus presentation (N/P190) (Eddy, Schmid, & Holcomb, 2006; Eddy & Holcomb, 2009) and are followed by the previously observed N300 and N400 effects with a short SOA (110 ms) and prime duration (50 ms). The early effect (N/P190), not previously observed in unmasked priming, presents itself as an early anterior negativity that inverses in polarity in posterior cortices, occurring between 100-250 ms (N/P190; Eddy, Schmid, & Holcomb, 2006). This effect has also been observed when using masked priming with faces (Henson, Mouchlianitis, Matthews, & Kouider, 2008). Specifically, Henson et al. (2008) found a repetition effect between 100 and 150 ms with greyscale faces. This effect likely parallels the process observed in the N190/P190 effect with objects. In addition, the observation of this early effect with faces and objects is similar to the effect reported with masked word priming studies where word were either partial or full repetitions (e.g., Holcomb & Grainger, 2006). The next component typically observed with objects is the N300 effect, a middle level, object representation specific component, observed with both masked and unmasked repetition priming and may parallel the form specific N250 found with words. With pictures, the N300 is a greater negativity for unrepeated/unrelated pictures compared to repeated/related pictures and is found less reliably with masked priming (Eddy, Schmid, & Holcomb, 2006; McPherson & Holcomb, 1999; Holcomb & McPherson, 1994). We are describing this component as a “mid level” effect because it seems to parallel mapping of perceptual information to a more form specific percept in memory that matches the primed object best, however, it is still reliant upon the physical form of the object and not higher level abstract conceptual and semantic features of the object. This idea is consistent with models of object recognition such as Schendan and Kutas’ (2007) where this second level representation allows for higher level cognitive processes to occur. The third component observed in these studies, the N400 is thought to reflect a general process mediated by semantics. This component is larger for unrepeated/unrelated pictures compared to repeated/related pictures in masked and unmasked conditions and reportedly occurs with both longer and shorter SOAs and prime durations (Eddy & Holcomb, 2009; Eddy, Schmid, & Holcomb, 2006; McPherson & Holcomb, 1999; Holcomb & McPherson, 1994).
The N/P190 effects are not observed with unmasked picture priming paradigms, leaving a question about the potential role of prime duration or SOA unanswered. In addition, reports that middle (N300) and later (N400) effects are observed with both masked and unmasked priming leaves unresolved the issue of whether the longer prime exposure or longer SOA is responsible for the discrepancy between masked and unmasked picture priming. Typically the N300 and N400 effects observed with unmasked primes are typically larger in magnitude than those observed with masked primes. These findings leave open the possibility that the prime duration or SOA modulates ERP effects observed in masked and unmasked repetition priming. In this study, we aim to examine how the duration of the prime and the SOA modulates these middle and late effects, as well as the early effect (N/P190) observed in rapid masked repetition priming.
We predict if longer exposure to the prime in the unmasked studies leads to larger amplitude N300 and N400 effects, then increasing the prime duration in a masked priming paradigm would result in larger amplitude N300 and N400 effects. Alternatively, if larger effects are observed in the unmasked studies due to more in-depth prime processing because of the longer prime-target SOA, then increasing the SOA between the prime and target in a masked priming paradigm, while keeping the prime duration constant at a short duration, should produce larger N300 and N400 effects. Experiments 1 and 2 are aimed at addressing these issues by systematically manipulating the SOA (Experiment 1) and prime duration (Experiment 2) while holding the other parameter constant.
1.1. Experiment 1
The first experiment examined how manipulating the interval (110, 230, 350, 470 ms) between the prime and target (SOA) affects ERP components while holding the prime duration constant (50 ms). Participants were presented with brief color picture primes (50 ms) that were forward and backward pattern masked and followed by a visible target picture for 300 ms (see Figure 1). The target picture was either a repetition of the prime or completely unrelated to the prime. The SOA was manipulated by varying the duration of the backward mask (60, 180, 300, 420 ms). Participants performed a semantic categorization task where they were asked to press a button to non-critical, occasional food pictures. The critical conditions for this experiment contained non-probe pictures.
FIGURE 1.
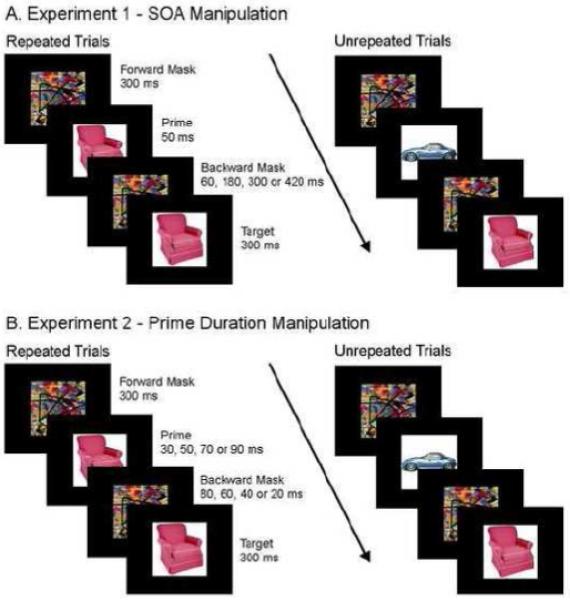
Typical trials and timing for (A) Experiment 1 and (B) Experiment 2.
By manipulating the SOA between the prime and target we were able to determine whether object processing reaches a more in-depth level with increased time between prime and target presentation. In particular, we hypothesized the N300 and N400 effects should be larger in amplitude for longer SOAs if more in-depth processing of the prime occurs with an increased duration between the prime and target. It is also predicted that the N/P190 effect may be limited to only shorter prime-target SOA durations based on the results Holcomb and Grainger (2007) reported where the N250 effect dissipated with longer SOAs (sometime between 180 and 300ms).
1.2. Experiment 2
The second experiment manipulated the duration of the prime (30, 50, 70, 90 ms) while keeping the SOA constant (110 ms). Participants were presented with brief color picture primes for either 30, 50, 70 or 90 ms that were forward and backward pattern masked and followed by a visible target picture for 300 ms. The target picture was either a repetition of the prime or completely unrelated to the prime. The SOA was held constant by varying the backward masked duration (80, 60, 40, 20 ms). Like in Experiment 1, participants performed a semantic categorization task where they were asked to press a button to non-critical, occasional food pictures. The critical conditions for this experiment contained non-probe pictures.
For this experiment, it was predicted that if increasing prime duration leads to more in-depth processing, then larger N300 and N400 effects would be expected. However, if more time is necessary for elaborate processing to occur before presentation of the target, then larger effects are not predicted with increased prime duration when the SOA (110 ms) remains constant across conditions. Additionally, the N/P190 effect should remain intact across all prime durations, but may be larger for longer prime durations since more perceptual information processing may be occurring with longer exposures. However, because the SOA is sufficiently short enough between the prime and target, the N/P190 not be disrupted at the shorter prime durations.
2. Results
2.1. Experiment 1 results
Experiment 1 manipulated the SOA between the prime and target while holding the prime duration constant. Participants performed a go no-go semantic categorization task where they pressed a button anytime a food item appeared. Since the prime duration was held constant at 50 ms for the four SOA manipulations the d’ values for probe items were collapsed across SOA. The average d’ for probe items in the prime position was 1.86 and the average d’ for probe food items appearing in the target position was 4.16. These d’ values demonstrate that the masking worked effectively and that participants were still processing targets for meaning (t(23) = −10.54, p < 0.00001).
Plots from 29 electrode channels for each SOA, time-locked to the target onset, can be seen in Figure 2 A-D. A subset of electrode sites (FC2, CP2, and O2) from these plots is shown at a larger scale and the distribution of these effects in Figure 3. The effects at the shorter SOA (110ms) were more likely to be due to overlap in residual processing of the prime while effects at the longer SOAs were most likely due to target processing and some residual processing of the backward mask. Therefore, only effects involving interactions of REPETITION and SOA were of interest in this first experiment.
FIGURE 2.
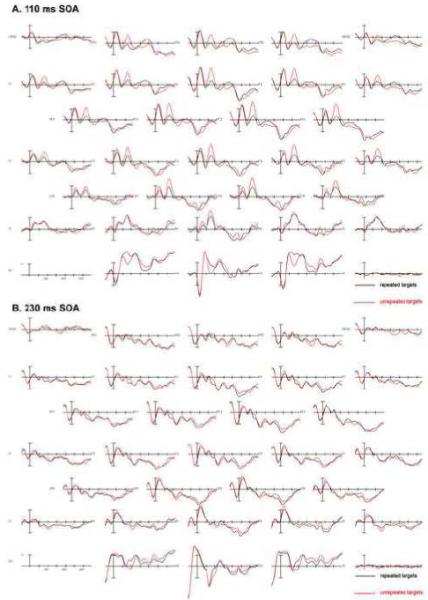
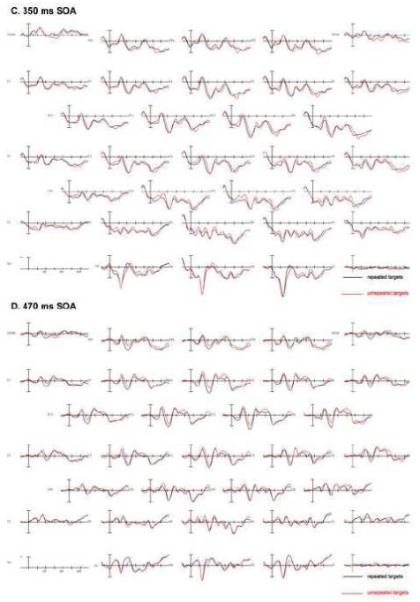
Experiment 1: SOA manipulation grand average ERPs (N=24) for each SOA. Repeated targets are in black, unrepeated in red. A: Grand average for 110 ms SOA; B: Grand average for the 230 ms SOA; C: Grand average from 350 ms SOA; D: Grand average from 470 ms SOA.
FIGURE 3.
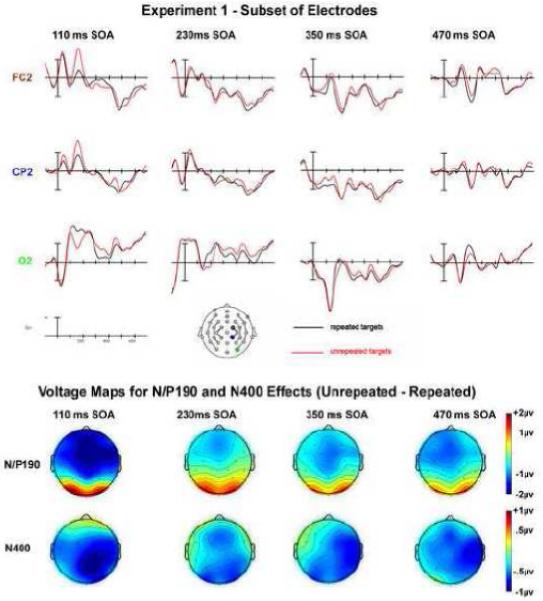
Experiment 1: SOA manipulation subset of electrodes and voltage maps. The voltage maps for the N/P190 are plotted at the peak of the effects for each SOA: 110 ms SOA at 155 ms post-target onset; 230 and 350 ms SOAs at 180 ms; 470 ms SOA at 175 ms. All of the voltage maps for the N400 effect are plotted at 450 ms post-target onset, except for the 470 ms SOA condition where the N400 can slightly earlier (400 ms).
2.1.1. N/P190 Epoch (100-225ms)
Repetition effects were observed during this epoch in the two shorter SOAs in the form of an anterior negativity for unrepeated target objects compared to repeated targets objects and in posterior sites a greater positivity for the unrepeated targets (Figs. 2-3). This effect of anterior negativity and a posterior positivity when comparing unrepeated to repeated targets was widely significant in the omnibus comparisons including all SOAs (REPETITION x SOA x electrode site x hemisphere interactions: C1: F(6,128) = 3.26, p = 0.017, ε = 0.629; C2: F(9,207) = 2.33, p = 0.046, ε = 0.559 and a REPETITION x SOA x electrode site interaction at Column 3 (F(12,276) = 3.465, p = 0.013, ε = 0.313). The interactions of REPETITION with SOA were mainly driven by the large anterior negativity and posterior positivity for the 110 ms condition, whereas the other conditions did not show as robust effects of perceptual processing on target objects from previous exposure to a repeated prime object (see voltage maps in Fig. 3). Additionally, the anterior negativity observed at the shorter SOA became more central in its distribution and the posterior positivity also became smaller with an increase in SOA (Fig. 3). A more detailed analysis of the anterior/posterior dissociation comparing two electrodes sites (FC2/O2) demonstrated that the size of this effect varied across SOA and also the distribution of this effect changed across SOA where, for example in the 230 ms SOA, the effect was larger in posterior electrodes, whereas in anterior electrodes the effect was slightly bigger for the 350 and 470 ms SOAs (SOA x REPETITION x electrode position F(3,69) = 6.354, p = 0.002, ε = 0.790).
Pairwise comparisons between the four different SOAs confirmed that the 110 ms SOA condition is mainly driving the above described interactions and also that it differs in distribution from the other SOAs. When examining the repetition priming effect in an anterior electrode (FC2) and a posterior electrode (O2), differences between each SOA and the 110 ms SOA were found that varied by anterior-posterior distribution of this effect (SOA x REPETITION x electrode site interactions: 110 vs. 230: F(1,23) = 10.178, p = 0.004; 110 vs. 350: F(1,23) = 17.093, p = 0.0004; 110 vs. 470: F(1,23) = 9.039, p = 0.006). However, no significant differences were found between the 3 later SOAs (230, 350, 470 ms) in pairwise comparisons (all F’s < 1.5, all p’s > 0.3).
When examining repetition priming effects in individual SOAs, the effect of REPETITION was particularly clear for the 110 ms SOA where there was an overall difference between unrepeated and repeated trials (as indicated by a main effect of REPETITION midline F(1,23) = 8.841, p = 0.007; C1 F(1,23) = 16.811, p = 0.0004; C2 F(1,23) = 12.702, p = 0.002; C3 F(1,23) = 4.434, p = 0.046). In addition, this difference between conditions varied in its anterior-posterior distribution, reflected by an interaction of REPETITION x electrode site (midline F(4,92) = 24.336, p = 0.00000002, ε = 0.551; C1 F(2,46) =10.767, p = 0.0003, ε = 0.909; C2 F(3,69) = 15.573, p = 0.00004, ε = 0.531; C3 F(4,92) = 33.003, p = 0.0000002, ε = 0.360). Moreover, in one electrode column the difference between repeated and unrepeated trials not only varied between anterior and posterior electrode sites but the difference between repeated and unrepeated trials was slightly larger over the right hemisphere compared to the left (see voltage maps, Fig. 3; REPETITION x electrode site x hemisphere interaction (C1 F(2,46) = 9.646, p = 0.001, ε = 0.737). Examining a small subset of anterior (FC2)/posterior (O2) electrodes confirmed this effect was driven by a difference in polarity between the anterior electrode site and the posterior electrode site with unrepeated trials being more negative-going than repeated trials in the anterior electrode and more positive-going in the posterior electrode (REPETITION x position: F(1,23) = 48.291, p = 0.000001).
The priming effect for the 230 ms SOA was primarily driven by a difference between the unrepeated and repeated conditions in the posterior electrodes (REPETITION by electrode site interaction at Column 3 F(4,92) = 6.246, p = 0.013, ε = 0.312). This interaction reflects the larger posterior positivity in comparison to a small negativity in anterior electrodes when comparing unrepeated to repeated targets. This was confirmed by a comparison of one anterior (FC2) and one posterior (O2) electrode site (REPETITION x position interaction F(1,23) = 5.953, p = 0.023).
A less robust effect was observed for the 350 ms SOA where both anterior and posterior electrode sites showed only small differences between the unrepeated and repeated targets. The effect observed at this SOA was slightly different than the previously observed effects since there was a small negativity in centro-parietal and posterior electrodes (REPETITION x electrode site interaction (midline: F(4,92) = 6.336, p = 0.002, ε = 0.578; C3: F(4,92) = 5.648, p = 0.008, ε = 0.456) and larger in the right hemisphere posterior electrodes (see Figure 2C electrodes C3 and C4: REPETITION x electrode site x hemisphere C1: F(2,46) = 5.986, p = 0.011,, ε = 0.728). When isolating a subset of electrodes (FC2 compared to O2) to examine if the same anterior-posterior change in polarity occurred for this effect, it was evident the same flip in polarity observed prior is not observed here (see Figure 3). However, as can be seen in Figure 2C, when comparing FC2 and O2, there was a larger difference between unrepeated and repeated trials in the more posterior electrode (O2) compared to the anterior electrode (FC2) driving a REPETITION x position effect (F(1,23) = 17.483, p = 0.0004).
For all electrode columns in the 470 ms SOA the difference between unrepeated and repeated trials interacted with the anterior-posterior distribution of this effect (Fig. 2D). In anterior electrodes the effect was more negative for unrepeated trials than repeated and more positive in posterior electrodes for this comparison (REPETITION x electrode site interactions midline: F(4,92) = 6.315, p = 0.005, ε = 0.445; C1: F(2,46) = 4.332, p = 0.034, ε = 0.696; C2: F(3,69) = 5.472, p = 0.016, ε = 0.476; C3: F(4,92) = 8.775, p = 0.002, ε = 0.362). Again, this effect showed an effect of distribution when comparing an anterior electrode (FC2) and a posterior electrode (O2), where the difference between unrepeated and repeated trials was larger in the posterior electrode (O2; indicated by a REPETITION by electrode interaction F(1,23) = 16.262, p = 0.001; see Figure 3).
2.1.1.1. Peak Latency Analysis: N/P190 Epoch (100-225ms)
An additional analysis of peak latency for this N/P190 effect revealed differences in the peak of this effect between the 110 ms SOA condition and others conditions. Peak latencies were examined for the difference waves (unrepeated-repeated trials) at one anterior (FP2) and one posterior (O2) electrode site for each of the SOAs (a comparison with analogous left hemisphere electrodes did not show an interaction of this effect with hemisphere: SOA x anterior-posterior x hemisphere interaction: F(3,69) = 0.920, p = 0.418, ε = 0.779). Mean peak latency differences can be seen in Figure 4 which shows difference waves for repetition effects of each SOA and the mean peak latencies. It can be clearly seen when examining the effect in FP2/O2 that the N/P190 effect peaks much earlier for the 110 ms SOA compared to the rest of the SOA manipulations. The overall ANOVA including all four SOAs revealed a SOA x electrode site interaction (F(3,69) = 3.098, p = 0.040, ε = 0.862) warranting further comparison.
FIGURE 4.
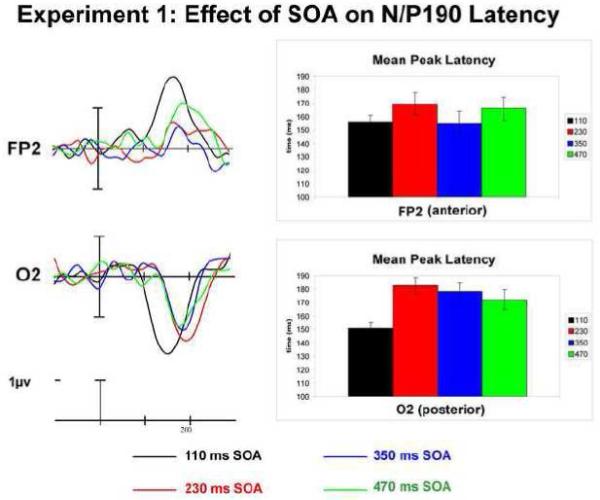
Experiment 1: The Effect of SOA on N/P190 Latency. Mean peak latency of N/P190 effect at anterior (FP2) and posterior (O2) electrode sites.
When comparing the 110 ms and 230 ms SOA conditions, the N/P190 effect peaked later for the 230 ms SOA than the 110 ms SOA (significant main effect of SOA for 230 ms SOA: F(1,23) = 19.214, p = 0.0002; see Figure 4). The anterior and posterior portions of this effect for the 230 ms SOA appear to peak around the same time. While for the longer SOAs, there appears to be not only a difference between SOAs when this effect peaks, but also a difference in when the anterior and posterior portions of this effect peak within each SOA. For example, the N190 portion of the effect peaked around a similar time for the 110 ms SOA compared to the 350 ms SOA at the anterior electrodes sites, but at a later time for the posterior portion of this effect for the 350 ms SOA (SOA x electrode site interaction for 110 ms vs. 350 ms SOA: F(1,23) = 7.906, p = 0.010) as does the 470 ms SOA (SOA x electrode site interaction 110 ms vs. 470 ms SOA: F(1,23) = 8.187, p = 0.009). The 230 ms SOA condition also differed in peak latency from the 350 ms (main effect of SOA: F(1,23) = 7.909, p = 0.010) and 470 ms condition (main effect of SOA: F(1,23) = 10.037, p = 0.004). The two longer SOAs did not differ in peak latency (350 ms vs. 470 ms; F<1, p’s > 0.35).
2.1.2. N300 Epoch (225-325ms)
The overall comparison including all SOAs indicated no effect of SOA on the N300 effect (all F’s < 2, p < 0.1). Examination of the grand averages (Figures 2 A-D) confirms this lack of interaction. However, because we predicted this effect may be modulated by SOA, priming effects for each individual SOA were examined. In contrast to the widespread effects observed for each SOA in the N/P190 time window, N300 effects were mainly observed in the 110ms SOA (see Figure 2A) although not in the typical direction and in a small subset of electrodes in the 350ms SOA (see Figure 2C). The N300 effect is typically characterized by a greater negativity for unrepeated target pictures compared to repeated pictures. In the 110ms SOA (Fig. 2A) the effect was observed in a small set of posterior electrodes, whereas in the anterior electrode sites the repeated trials are more negative going than unrepeated trials, indicating the direction of the effect is not the typical N300 effect (REPETITION by electrode interactions observed at midline: F(4,92) = 5.532, p = 0.014, ε = 0.371; C2: F(3,69) = 5.309, p = 0.018, ε = 0.471; C3: F(4,92) = 4.449, p = 0.027 and REPETITION x electrode x hemisphere interactions at C3: F(4,92) = 3.408, p = 0.045). In the 350ms SOA (Fig. 2C) the N300 effect is limited to the peripheral column (C3) and is posterior in distribution as the REPETITION x electrode interaction reveals (F(4,92) = 3.879, p = 0.035).
2.1.3. N400 Epoch (325-500ms)
The N400 effect did not interact with SOA, however, there were robust interactions of REPETITION and electrode site in the omnibus ANOVA (midline: F(4,92) = 6.216, p = 0.004, ε = 0.519; C1: F(2,46) = 4.357, p = 0.024, ε = 0.860; C2: F(3,69) = 6.669, p = 0.008, ε = 0.483; C3: F(4,92) = 9.613, p = 0.002, ε = 0.341). There was a linear trend for SOA interacting with these variables (SOA x REPETITION x electrode site - midline: F(1,23) = 5.072, p = 0.034; C3: F(1,23) = 6.381, p = 0.019). Visual inspection of the data confirms this interaction and linear trend where the effects of repetition are more posterior at the shorter SOA (110 ms) and become more anterior at the longer SOA (470 ms).
2.2. Discussion of Experiment 1
The early priming effects (N/P190) previously observed by Eddy, Schmid, and Holcomb (2006) with a short SOA of 110 ms, were found in this experiment using the same SOA. Additionally, this effect was found with longer SOAs, although, it was much less robust. These findings are consistent with Holcomb and Grainger’s (2007) findings with words where they reported that increasing the SOA leads to a reduction in early ERP components as well as with Henson, Rylands, Ross, Vuilleumeir, and Rugg’s (2004) findings of increased repetition lag leading to reduced repetition priming between 150 and 300 ms with supraliminal pictures. The results of Experiment 1 are also support the idea that masked priming reflects a short-lived process, especially the earlier components. Since this early priming effect is most likely reflecting early perceptual processing of visual features of the objects, it is not surprising that when a greater amount of time comes between the prime and target, this effect dissipates. One interesting observation is the change in onset and peak of this early component as the SOA becomes longer. In addition, with the increase in SOA, there is a divergence in timing of this peak between anterior and posterior electrodes. Along with a later peak, the amplitude of this effect becomes smaller as well. One explanation for the delay in peak of this effect could be reflecting less benefit from the perceptual information available from the prime leading to an increased amount of time to see a perceptual effect on the target. As the time increases between the onset of the prime and target, this effect is less likely to reflect overlap in perceptual processing of the prime and target and more likely to be refractory.
The fact that we did not observe a robust N300 is not surprising, given our previous finding where the N300 effect was either small or non-existent in masked priming (Eddy, Schmid, & Holcomb, 2006; Eddy & Holcomb, 2009). There are a variety of possibilities for why the N300 is not robustly modulated with a short prime duration of 50 ms. One possibly is in the shorter SOA, because the prime and target come close together the process indicated by the N300 is either bypassed or gets pushed back in time and becomes combined with the N400. In the 230 and 350 ms SOA conditions, there is a hint of an N300, suggesting with a longer SOA the process reflected by the N300 has more time to happen and have an effect target processing on the form specific representation. From visual inspection of the data, it appears we have an N400 effect for each SOA, although with an increasing anterior distribution with an increase in SOA. This is consistent with the findings of Holcomb and Grainger (2007) who found that longer latency priming effects remained intact even with a longer duration between the prime and target.
Unlike Holcomb and Grainger’s (2007) findings, the N400 effect did show trends toward interacting with SOA. This most likely reflects a difference in distribution of the effect than size of the repetition priming effect, although the N400 was not as large in magnitude as the N400 found by Holcomb and Grainger (2007). A point to consider, when comparing our results to Holcomb and Grainger’s (2007), is that the N400 effect is much smaller in magnitude than the N400 effect they observed possibly reflecting an additional burden in processing complex pictures compared to words.
From the results of this first experiment, it does not appear that increasing the duration between the prime and target allows for more in-depth, higher level processing effects to occur as the N400 effect was significant in omnibus comparisons and did not interact with SOA except in its distribution. It appears the process underlying the N400 effect may change with an increase in SOA as the distribution of the effect becomes more anterior. With regard to prior unmasked picture priming studies (Holcomb & McPherson, 1994; McPherson & Holcomb, 1999), it does not appear the longer SOA in these experiments accounts for the more robust N400 effects observed. Experiment 2 aimed to address the other possibility: increased prime duration.
2.3. Experiment 2 results
The second experiment tested whether increased prime exposure leads to increased processing of object-related information. Prime durations were varied (30, 50, 70, 90 ms) while holding the SOA at a constant 110 ms. As in experiment 1, participants performed a go no-go semantic categorization task where they only pressed a button to pictures of food items.
The average d’ was calculated for probe items (food pictures) in the prime position for each of the prime durations. For the 30ms duration the d’ was 0.37, for the 50ms duration it was 1.90, the 70ms duration 3.00 and for the 90ms duration it was 2.88, suggesting that participants were able to identify food objects in the prime position at the 70 and 90ms prime durations, but not at the two shorter durations (main effect of duration: F(3,69) = 64.14, p < 0.0000001). Pairwise t-tests confirmed the shorter durations drove this difference in d’ score, where the 30 and 50 ms values differed significantly from not only each other, but the longer duration (all t’s < −4, all p’s < 0.0001). The d’ for the 70 and 90 ms durations did not differ significantly (t(23) = 1.021, p = 0.318). The average d’ for food items appearing in the target position was 4.09. These d’ values demonstrate that the masking worked effectively at the shorter durations and that participants were still processing targets for meaning. However, as has been argued elsewhere (Holcomb & Grainger, 2007), even though participants detected a significant number of probe items at the longer prime durations, this does not necessarily mean that there was enough time between prime and target onset for significant high level processing of the prime to influence early target processing.
For the duration manipulation, the timing of repetition priming effects were slightly different than those observed in Experiment 1, as reflected in the grand averages (see Figures 5 A-D). Therefore, the time windows for the data analysis were adjusted accordingly to best describe the effects. The N/P190 was measured between 100 and 250 ms; the N300 between 250 and 350 ms; the N400 between 350 and 500 ms.
FIGURE 5.


Experiment 2: Prime duration manipulation grand average ERPs (N=24) for each prime duration. Repeated targets are in black, unrepeated in red. The grand averages are time-locked to the onset of the target, base-lined to the prime period. A: Grand average for 30 ms prime; B: Grand average for the 50 ms prime; C: Grand average from 70 ms prime; D: Grand average from 90 ms prime.
2.3.1. N/P190 Epoch (100-250 ms)
Widespread effects at all prime durations are evident in Figure 5 A-D in the earliest time window analyzed. An omnibus ANOVA confirmed the visual inspection, where REPETITION, prime duration and electrode site interacted at all electrode columns (Midline: F(12,276) = 4.46, p = 0.002, ε = 0.373; C1: F(6, 138) = 2.953, p = 0.034, ε = 0.548; C2: F(9, 207) = 2.949, p = 0.033, ε = 0.374, C3: F(12, 276) = 5.385, p = 0.001, ε = 0.285) while REPETITION x prime duration x electrode site x hemisphere (F(12,276) = 2.340, p = 0.030, ε = 0.550) interacted in Column 3 and REPETITION, prime duration and hemisphere interacted significantly in Column 1 (F(3, 69) = 4.359, p = 0.010, ε = 0.900). Overall repetition effects can be seen in Figure 5 and Figure 6 where there was a greater negativity for unrepeated targets in the anterior sites (e.g., FC2) and a greater positivity in posterior electrode sites (e.g., O2).
FIGURE 6.
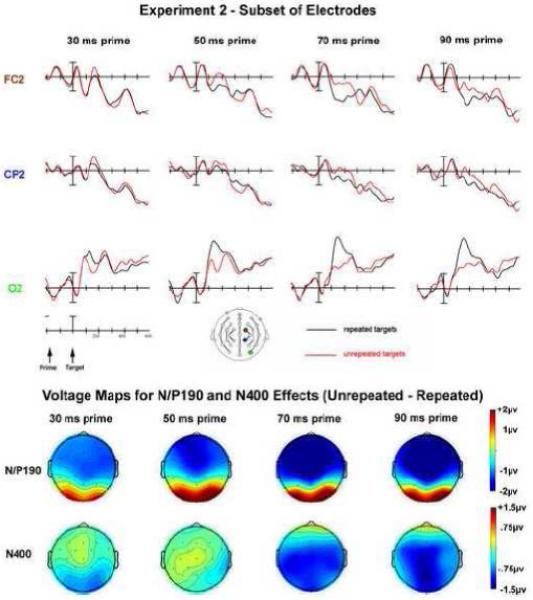
Experiment 2: Prime duration manipulation - subset of electrodes and voltage maps. The voltage maps for the N/P190 are plotted at 165 ms post-target onset; the voltage maps for the N400 effect are plotted at 420 ms post-target onset.
To further investigate the overall effects observed in the omnibus comparison, repetition effects were examined for each of the prime durations. The anterior/posterior dissociation was observed for all prime durations as can be seen in Figure 6, however, it was largest at the 70 ms prime duration confirmed by examining the interaction between REPETITION by electrode position (FC2 vs O2) in each prime duration (30ms: F(1,23) = 6.708, p = 0.016, ηp2 = 0.226; 50ms: F(1,23) = 40.283, p = 0.0000002, ηp2 = 0.637; 70ms: F(1,23) = 118.358, p = 0.0000000002, ηp2 = 0.837; 90ms: F(1,23) = 53.863, p = 0.0000002, ηp2 = 0.701). This increase in the N/P190 effect with prime duration was also confirmed by significant linear trends for the overall ANOVA (interaction of REPETITION x prime duration x electrode site (all F’s > 4.8, p’s < 0.04).
2.3.2 N300 Epoch (250-350 ms)
Examination of the ERP waveforms (see Figure 5A-D) shows a more positive-going deflection for the unrepeated targets than the repeated targets in all the conditions, which is inconsistent with the typical N300 effect (where the unrepeated should have a more negative going deflection than the repeated trials). This positivity, in the overall ANOVA produces only an interaction of REPETITION, prime duration, electrode site, and hemisphere in Column 2 (F(9, 207) = 2.682, p = 0.027, ε = 0.527). While this interaction was observed, we did not perform further analyses since this effect was not in the predicted direction of a typical N300.
2.3.3. N400 Epoch (350-500 ms)
In the overall ANOVA main effects of REPETITION were observed in Column 2 (F(1,23) = 9.563, p = 0.005) and Column 3 (F(1,23) =16.147, p = 0.001). In addition, an interaction of REPETITION, prime duration and electrode site was observed in Column 1 (F(6,138) = 2.902, p = 0.030, ε = 0.612). These overall interactions were driven by the unrepeated condition being more negative than the repeated condition (e.g., larger repetition effect) in the 70ms and 90ms conditions more so than the 30 and 50ms conditions (Fig. 5 and Fig. 6). In addition, for the prime durations where visual inspection of the waves suggests a significant N400 effect, the N400 has a central-posterior distribution (which explains the interaction with electrode sites).
To further confirm this visual inspection, repetition effects were examined for each of the prime durations. The 30 ms prime duration condition did not show any significant repetition priming effect, confirming that the interaction of prime duration and REPETITION in the overall comparison was not being driven by this prime duration. The 50 ms prime condition only showed an interaction of REPETITION x electrode x hemisphere in one electrode column (Column 2 F(3,69) = 5.991, p = 0.006, ε = 0.625). This interaction is reflecting how the effect in the right hemisphere (Column 2 electrodes) show a greater negativity for unrepeated compared to repeated trials, whereas in the left hemisphere electrodes, the repeated trials are showing a greater negativity (Fig. 5B). More robust N400 effects are clearly seen in visual inspection of the waveforms for the 70 ms prime duration (Figure 5C). The N400 effect has a central-posterior distribution as indicated by a REPETITION x electrode interaction (Midline: F(4,92) = 5.109, p = 0.013, ε = 0.448; C2: F(3,69) = 4.376, p = 0.026, ε = 0.550; C3: F(4,92) = 5.562, p = 0.010, ε = 0.434). In the 90 ms prime duration, an N400 effect was also observed, confirmed by a main effect of REPETITION (C2: F(1,23) = 7.058, p = 0.014; C3: F(1,23) = 8.565, p = 0.008
In addition, there was no significant relationship between the mean amplitude of the N400 effect observed at midline electrode sites and d’ scores for each of the prime durations (all r’s < .2, all p’s > 0.381).
2.4 Discussion of Experiment 2
Experiment 2 tested the hypothesis that additional processing of the prime produces larger high level (semantic) effects in masked repetition priming, rather than a longer time between the prime and target (tested in Experiment 1). In agreement with Experiment 1, the 50 ms prime duration produced an N/P190 and a small N400 effect, although the N400 effect in Experiment 1 was more widespread than in Experiment 2. While a similar pattern of effects was observed for the 30 ms prime duration, these effects were not as robust and did not prove to be statistically significant for the N400 epoch. These finding suggest there is a minimal amount of exposure necessary to produce benefits in target processing that occurs somewhere around 30 ms. More fine-grained analysis of different prime durations is necessary to determine this precisely.
The main finding of this study was a change in the size of the N400 effect with prime exposure. While earlier perceptual effects did not differ, the N400 effect increased in amplitude linearly with an increase in prime duration. And as suggested above, while the prime becomes more visible at the longer prime exposures (70 ms and 90 ms), indicated by d’ scores, the SOA is short enough to be primarily reflecting rapid object recognition processes. the N400 effect was not attributable to prime awareness or ability to see the prime, as no correlation was found between the size of the N400 effect and d’ scores for each of the durations. Other studies such as Holcomb et al. (2005) have found a relationship between prime awareness and N400 priming effects, suggesting that awareness of the prime identity may influence the N400 effect in addition to the increased processing of the prime when the prime duration is lengthened.
These findings are comparable to those in the Holcomb and Grainger’s (2007) manipulation of prime duration. They found with the shortest prime durations (10 ms and 20 ms) that there was an absence of all priming effects, but as the prime duration increased (30 ms and 40 ms) the N250 effect (reflecting orthographic processing) emerged and the N400 effect became more robust. This is not unlike the pattern of findings, where the shortest duration only produced perceptual priming effects (N/P190) whereas the longer prime durations showed gradually larger priming effects on the N400. Given that our prime durations were significantly longer than the ones used by Holcomb and Grainger (2007) we cannot draw exact parallels between our findings and their findings, however, it is reasonable to speculate that picture processing takes considerably more time than single word processing, therefore our durations may be more similar to their prime durations in terms of the amount of processing that is taking place.
3. General Discussion
The results of Experiment 1 and 2 suggest that longer prime exposure leads to more in-depth processing as indexed by larger N400 effects rather than a longer time window between the prime and target to allow for more extensive processing. Effects of prime duration on later processing components, observed with a relatively short SOA (110ms), further support the idea that more time between the prime and target does not lead to increased processing of the prime. However, since we limited the prime duration to 50 ms while manipulating SOA, we do not know the effects of longer durations in combination with longer SOAs.
The manipulations of SOA and duration with a 50 ms prime duration and 110 ms SOA replicate findings from our previous masked picture repetition study (Eddy, Schmid, & Holcomb, 2006). Specifically, N/P190 and N400 effects previously reported for those prime durations and SOAs were observed in the current experiments as well. Overall, increasing the prime duration led to a linear increase in the size of the N400 repetition priming effect, while increasing the SOA did not change the size of the N400 effect. However, our prime duration results do diverge from the findings of Holcomb and Grainger (2007) since we did not find an N400 effect at the 30 ms prime duration. One explanation for these differential results could be that pictures require more initial processing than words. Words are less complex than real pictures of objects and contain far less perceptual information. In order for a later semantic effect to be observed, sufficient processing of this perceptual information must occur to map onto a higher level semantic representation. This would account for our overall smaller N400 effects in both the prime duration and SOA manipulations in comparison to these manipulations with words.
In both experiments, we did not observe robust N300 effects when manipulating the SOA or prime duration. The short prime duration in Experiment 1 could be one possible explanation for the lack of this effect, leading to rapid processing where the mechanism underlying the N300 happens rapidly and becomes combined with the N400 effect. But even when we extend the prime duration to 90 ms in Experiment 2 we do not observe an N300 effect, possibly suggesting both prime duration and SOA play a role in eliciting this effect. In addition there is a hint of an N300 effect at the 230 and 350 ms SOAs and also at the longer prime duration. There could be two different mechanisms that could explain this pattern. In the SOA experiment, the N300-like effect comes later, closer to the N400, possibly suggesting the time between the prime and target allows for activating a higher level form specific representation that then affects target processing, whereas, with the longer prime duration, the same could be true, but because the SOA is short, the N300 gets pushed earlier in time and overlaps with the N/P190 effect. These two explanations suggest when the SOA is lengthened, the N300 gets pushed later in time because the longer SOA allows, even with a short prime duration, for the form specific object representation to get activated, whereas with a short prime duration and short SOA this does not happen, leading to no effect whatsoever. In the longer prime duration, because the SOA is still short, but the prime exposure longer, a higher level representation gets activated by the prime, but the target comes so quickly afterwards that the perceptual and form specific processing happen in one quick sweep.
The N/P190 effect did show sensitivity when the SOA was manipulated, similar to the findings of Holcomb and Grainger (2007) where an N250 effect was only observed at shorter SOAs, suggesting ERP repetition priming components sensitive to stimulus form show less benefit when the time between the prime and target exposure is lengthened. And with our pictures, the N/P190 effect, reflecting perceptual processing, became smaller and shifted in latency as the SOA increased, whereas, when the prime duration was manipulated this priming effect did not change. This suggests a mechanism of perceptual decay, where longer intervals between the prime and target lead to dissipating perceptual priming effects, while longer prime exposure does not change the magnitude of this effect.
As mentioned, the latency shift of the N/P190 in the SOA manipulation could be indicative of the decay of perceptual information and that the benefit in perceptual processing takes slightly longer to occur when the prime and target are separated by more time or requires some recursive processing. It is possible that the prime duration does play a role in this – if the backward mask limits recursive processing then it is just decay of the representation activated by the prime. To further examine the combined effects of prime duration and SOA, future studies could extend this paradigm to a longer prime duration while still manipulating the SOA to see if this changes the pattern of priming effects.
4. Experimental Procedure
4.1. Experiment 1
4.1.1. Participants
Twenty-four volunteers (10 female, mean age = 21, range 20-27, SD = 1.65), all undergraduate students at Tufts University, were paid $20 to participate in this experiment. All were right-handed with normal or corrected to normal visual acuity.
4.1.2. Stimuli and Procedure
Color photographs of 620 common objects taken from conventional views were displayed on a white background (each 256 × 256 pixels) on a 19-in. display (visual angle 2°) time-locked to the vertical refresh signal of the video card (100 Hz resolution). Each subject viewed 390 pseudorandomly arranged trials composed of a unique prime–target pairs of objects. In the 320 critical trials (those used to form the ERPs of interest), four sets of 40 pairs of stimuli were presented where the same picture occurred in the prime and target positions (repeated) for each of the four SOAs. Another four sets of 40 pairs of pictures were presented where different unrelated objects were shown in the prime and target positions (unrepeated; see Figure 1a). The remaining 70 trials contained a ‘probe” object (food item) paired with a nonfood object (40 in the prime position and 30 in the target position divided among the four SOAs) matched with non-food fillers. Twelve counterbalanced lists resulted in each repeated and unrelated object being presented an equal number of times across participants, but each object being presented in only one trial for each participant.
Stimuli were presented with a forward and backward mask, with a prime presented for 50ms in between. The forward mask was presented for 300 ms and the backward mask was presented for 60, 180, 300, or 420ms to manipulate the SOA between the prime and target to be 110, 230, 350 or 470ms. The target was then presented for 300ms. Figure 1a depicts examples of the two types of critical trials and their timing. Participants were instructed to attend to the screen and rapidly press a button whenever they detected an object depicting a food item. Participants were given clear instructions on what constituted a food item (e.g., live animals are not food items; food related items are not food items). All other items, including the critical items, were to be viewed passively.
4.1.3. EEG Recording
A 32-channel electrode cap (Electro-cap International) was used to collect the electroencephalogram (EEG; see Figure 7 for electrode locations). All electrodes, including one over the right mastoid, were referenced to an electrode over the left mastoid (the right mastoid was used to monitor differential left mastoid activity; none was found, when examining SOA x repetition interactions – all F’s < 1.8, all p’s > 0.17). Horizontal and vertical eye movements blinks were detected from electrodes placed below and to the side of the eyes (scalp impedances of <2 kΩ). The EEG (200 Hz sampling rate, bandpass 0.01 and 40 Hz) was recorded continuously and ERPs averages were formed by time-locking to the onset of targets. Trials with blinks, eye movements, and muscle artifact were rejected prior to averaging (approximately 10% of trials).
FIGURE 7.
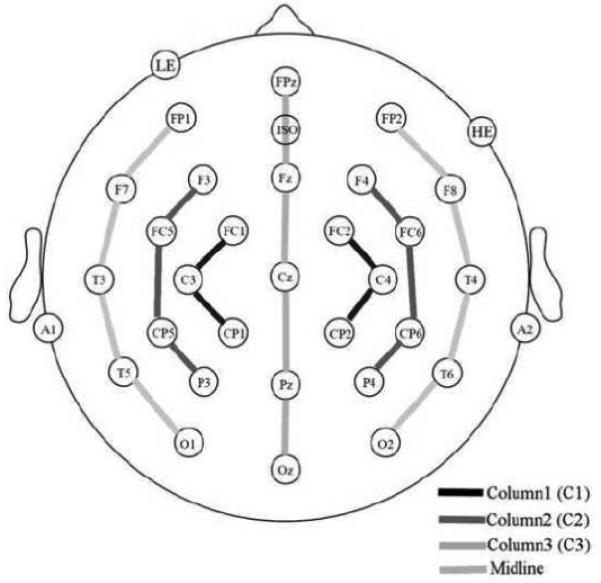
Electrode montage and the four columns used for analysis.
4.1.4. Data Analyses
Based on our previous research using masked picture priming and examination of the grand average ERPs we selected three time windows for analyses. Mean amplitude measurements were made in an early region 100–225 ms; the N300 region 225–325ms, and the N400 region 325–500 ms. ANOVAs were performed with SOA, Repetition, Electrode, and Hemisphere (except midline) as within-subject factors in four separate columnar analyses (e.g., Holcomb, Reder, Misra, & Grainger, 2005; see Figure 7 for four columns). The Geisser and Greenhouse (1959) correction was applied to all repeated measures having more than one degree of freedom and the corresponding ε values are reported. An additional analysis examining the peak latency differences between anterior and posterior sites in the early effect time window (100-225 ms) were performed comparing electrodes FP2 and O2, the electrodes where the response for this effect was maximal.
4.2. Experiment 2
4.2.1. Participants
Twenty-four volunteers (14 female, mean age = 20, range 19-26, SD = 1.71), all undergraduate students at Tufts University, were paid $20 to participate in this experiment. All were right-handed with normal or corrected to normal visual acuity.
4.2.2. Stimuli and procedure
The critical and non-critical stimuli for this experiment were the same color pictures used in Experiment 1. All procedures were also identical with the exception of the SOA and prime durations. In Experiment 2 the SOA was held constant at 110 ms and the duration of primes was varied across four levels: 30 ms, 50 ms, 70 ms and 90 ms. A typical trial is illustrated in Figure 1b.
4.2.3. EEG Recording
The same recording parameters from Experiment 1 were used for this Experiment (see section 4.1.3). Again no differential mastoid activity was observed, as assessed by the prime duration x repetition interaction (all F’s < 0.4, p > 0.8).
4.2.4. Data Analyses
The same data analyses were performed for this manipulation of duration using slightly different time windows that characterized the effects the best: early (100-250 ms), middle (250-350 ms), and later (350-500 ms). ANOVAs were performed with Prime Duration, Repetition, Electrode, and Hemisphere (except midline) as within-subject factors in four separate columnar analyses (e.g., Holcomb, Reder, Misra, & Grainger, 2005; see Figure 7 for four columns). The Geisser and Greenhouse (1959) correction was applied to all repeated measures having more than one degree of freedom and the corresponding ε values are reported.An additional analysis examining the differences between anterior and posterior direction of the early effect were performed comparing electrodes FC2 and O2 where the response for this effect was maximal. In addition, to examine the relationship between the N400 effect and prime visibility, we examined the correlation between the d’ score for each prime duration and the mean amplitude of the N400 effect at midline electrodes.
Acknowledgements
This research was supported by grant number HD25889. The authors would like to thank Alexandra Cheetham, Tarun Sridharan and the students in Psychology 49 at Tufts University for their help in collecting data.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Eddy M, Schmid A, Holcomb PJ. Masked repetition priming and event-related brain potentials: A new approach for tracking the time-course of object perception. Psychophysiology. 2006;43(6):564–568. doi: 10.1111/j.1469-8986.2006.00455.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eddy MD, Holcomb PJ. Electrophysiological evidence for size invariance in masked picture repetition priming. Brain and Cognition. 2009;71(3):397. doi: 10.1016/j.bandc.2009.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eddy MD, Schnyer D, Schmid A, Holcomb PJ. Spatial dynamics of masked picture repetition effects. NeuroImage. 2007;34(4):1723–1732. doi: 10.1016/j.neuroimage.2006.10.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forster K, Mohan K, Hector J. The mechanics of masked priming. In: K S, L SJ, editors. Masked priming: State of the art. Psychology Press; Hove, UK: 2003. pp. 3–37. [Google Scholar]
- Forster KI, Davis C. Repetition priming and frequency attenuation in lexical access. Journal of Experimental Psychology: Learning, Memory, & Cognition. 1984;10(4):680–698. [Google Scholar]
- Geisser S, Greenhouse S. On methods in the analysis of profile data. Psychometrica. 1959;24:95–112. [Google Scholar]
- Grainger J, Jacobs A. Temporal integration of information in orthographic priming. Visual Cognition. 1999;6:461–492. [Google Scholar]
- Henson RN, Rylands A, Ross E, Vuilleumeir P, Rugg MD. The effect of repetition lag on electrophysiological and haemodynamic correlates of visual object priming. NeuroImage. 2004;21(4):1674. doi: 10.1016/j.neuroimage.2003.12.020. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ, Grainger J. On the time course of visual word recognition: An event-related potential investigation using masked repetition priming. Journal of Cognitive Neuroscience. 2006;18(10):1631–1643. doi: 10.1162/jocn.2006.18.10.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holcomb PJ, Grainger J. Exploring the temporal dynamics of visual word recognition in the masked repetition priming paradigm using event-related potentials. Brain Research. 2007;1180:39–58. doi: 10.1016/j.brainres.2007.06.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holcomb PJ, McPherson WB. Event-related brain potentials reflect semantic priming in an object decision task. Brain & Cognition. 1994;24(2):259–276. doi: 10.1006/brcg.1994.1014. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ, Reder L, Misra M, Grainger J. The effects of prime visibility on erp measures of masked priming. Cognitive Brain Research. 2005;24(1):155–173. doi: 10.1016/j.cogbrainres.2005.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamme VAF, Zipser K, Spekreijse H. Masking interrupts figure-ground signals in v1. Journal of Cognitive Neuroscience. 2002;14(7):1044–1053. doi: 10.1162/089892902320474490. [DOI] [PubMed] [Google Scholar]
- McPherson W, Holcomb PJ. An electrophysiological investigation of semantic priming with pictures of real objects. Psychophysiology. 1999;36(1):53–65. doi: 10.1017/s0048577299971196. [DOI] [PubMed] [Google Scholar]
- Misra M, Holcomb PJ. Event-related potential indices of masked repetition priming. Psychophysiology. 2003;40:115–130. doi: 10.1111/1469-8986.00012. [DOI] [PubMed] [Google Scholar]
- Schendan H, Kutas M. Neurophysiological evidence for the time course of activation of global shape, part, and local contour representations during visual object categorization and memory. Journal of Cognitive Neuroscience. 2007;19(5):734–749. doi: 10.1162/jocn.2007.19.5.734. [DOI] [PubMed] [Google Scholar]