

Abstract
Oysters (Crassostrea virginica) were a central component of the Chesapeake Bay ecosystem in 1607 when European settlers established Jamestown, VA, the first permanent English settlement in North America. These estuarine bivalves were an important food resource during the early years of the James Fort (Jamestown) settlement while the colonists were struggling to survive in the face of inadequate supplies and a severe regional drought. Although oyster shells were discarded as trash after the oysters were eaten, the environmental and ecological data recorded in the bivalve geochemistry during shell deposition remain intact over centuries, thereby providing a unique window into conditions during the earliest Jamestown years. We compare oxygen isotope data from these 17th century oyster shells with modern shells to quantify and contrast estuarine salinity, season of oyster collection, and shell provenance during Jamestown colonization (1609–1616) and the 21st century. Data show that oysters were collected during an extended drought between fall 1611 and summer 1612. The drought shifted the 14 psu isohaline above Jamestown Island, facilitating individual oyster growth and extension of oyster habitat upriver toward the colony, thereby enhancing local oyster food resources. Data from distinct well layers suggest that the colonists also obtained oysters from reefs near Chesapeake Bay to augment oyster resources near Jamestown Island. The oyster shell season of harvest reconstructions suggest that these data come from either a 1611 well with a very short useful period or an undocumented older well abandoned by late 1611.
Keywords: Chesapeake Bay, Crassostrea virginica, environmental reconstruction, oxygen isotope, scleroarchaeology
During the latter phases of sea level rise in the early Holocene, the Susquehanna River valley was inundated by the Atlantic Ocean, forming the modern Chesapeake Bay (1, 2). As sea level rose, eastern oysters (Crassostrea virginica) invaded the Bay, forming large populations and biogenic structures that expanded up the southern tributaries (1). By the early 17th century, self-perpetuating oyster populations dominated the system ecologically and spatially (1). Oysters provided habitat for numerous other species (3–5), created reef structures that delineated channels and funneled tidal flow (1, 6), enhanced bank stability between the estuary and tidal marshes (7), and provided a food resource for Native Americans (8). Dense oyster populations also provided ecological services that were central to estuarine trophic structure (9–11).
As the first settlers sailed up the James River in May 1607 and began construction of the James Fort on Jamestown Island (Fig. 1), they were unaware that the survival of the first permanent English settlement in North America would be intimately linked to local oyster populations. Although Jamestown was well established by the 1620s as the seat of colonial government (12, 13), shortages of food (14) and fresh drinking water (14, 15), combined with poor leadership (14, 15), nearly destroyed the colony during its first decade. The colonists had no way of knowing that their arrival in Virginia coincided with the beginning of a severe regional drought that included the driest 7 y (1606–1612) in nearly 8 centuries (16). The drought caused crop failures for the native Algonquin people (8, 14), reducing the likelihood that the colonists could successfully barter for large quantities of food. In May 1609, John Smith sent men downriver to live “on the oyster banks” for 9 wk to reduce pressure on meager food resources at the Fort (14, 17). During and immediately after the winter of 1609–1610 (“The Starving Time”) (14), the settlers relied on oysters for food as they recovered from a winter in which ~44% of the colony died from causes including food and fresh water shortages (15).
Fig. 1.

Map of Chesapeake Bay (A) showing the James River, VA (B) including collection locations for historic and modern oyster shells, modern water samples for isotope analyses, and modern water temperature (Fig. 3A) and salinity (Fig. 3B) monitoring stations. Fig. S1 is a larger version of this map.
Initially, a fresh water source was absent from Jamestown, and the colonists drank brackish James River water (14). The lack of fresh water likely contributed to the high mortality rates at Jamestown from 1607 to 1609 (15). Three Jamestown wells were documented from 1609 to 1624, although more may have been dug. In 1609 John Smith recorded the first well (14), and in May 1611 Governor Thomas Dale ordered construction of another well (14). The next well was documented in 1617 when Governor Samuel Argall either renovated the existing well or dug a new well to establish a reliable supply of fresh water (13).
Abandoned James Fort wells were filled with the colony's trash including oyster shells (18). The infill is relevant archaeologically and the artifacts within a well provide insight into the time period and time course of well fill. Thus, archaeologists have concluded that a well [Association for the Preservation of Virginia Antiquities (APVA) Structure ID 170] discovered in 2003 just outside the James Fort containing artifacts dating to 1619–1625 is likely the Argall well of 1617 (13, 18).
During spring 2006, archaeologists discovered another well (APVA Structure ID 177; Fig. 2) within the James Fort (19) which is not clearly discernible as either the original Smith well or the Dale well on the basis of descriptive examination alone. Although this well cannot be from before 1609, foundations found directly over it date to 1617 (19), indicating that it was filled and capped by this date. Thus the period of use is limited to 1609–1616. Large numbers of oyster shells were part of the backfill in this well and form the foundation of this study.
Fig. 2.

Profile of Jamestown well (Structure 177) that was the source of oyster shells used in this study. Shells from P, U, and Z layers (shown in black) were used. Profile is modified from Fig. 114 in ref. 19 and is used with permission.
The growth, life cycle, and ecology of C. virginica have been studied extensively from mid-Atlantic estuaries (20, 21). Within the Chesapeake Bay system, oyster shell growth occurs between ~8 and 25 °C (22). Each oyster shell contains a geochemical (18O/16O) record of growth, season of collection, estuarine temperatures and salinities (23, 24) and information that can identify locations of source oyster reefs. The James River and watershed have changed dramatically between 1607 and the present in terms of terrestrial vegetation (25), land use (25, 26), sedimentation rates (27, 28), nutrient sources (29, 30), and trophic structure (1, 9, 27, 30, 31). Because the James Fort well excavated in 2006 was abandoned and filled within a short time window, geochemical analyses of the oysters in this well provide an unparalleled opportunity to document estuarine environmental conditions during the earliest period of colonization. Thus, the Jamestown oysters provide a valuable contrast with modern James River oysters. Here, we use oyster shell oxygen isotope data to quantitatively compare modern and early 17th century James River estuarine conditions and to describe the season of early Jamestown oyster collection, to reconstruct oyster provenance and the timing and pattern of well fill, and to provide evidence to place this well within the Jamestown timeline.
Results and Discussion
Modern Oyster Ecology and Distribution in the James River.
The modern James River estuary displays salinity zones along its length. End member salinities are controlled by freshwater influx and tidal exchange. James River salinities are lower between March and May due to elevated rainfall runoff (32–34) than between June and January (33, 34). In the modern James River, the 2-psu isohaline migrates seasonally from Mulberry Point (MP) during wet conditions, to just above Jamestown Island (JI) during dry conditions (Fig. 1). Wreck Shoal (WS) typically experiences salinities between 10 and 23 psu, although salinities as low as 3 psu have been recorded during heavy rainfall events (Fig. 3B). In the lower estuary, Old Point Comfort (OPC), and Middle Ground (MG) experience annual salinity ranges of 15–28 psu (Fig. 3B).
Fig. 3.

Measured bottom water temperatures (A, °C) and salinities (B, psu) for WS and OPC, James River, VA, from 2002 to 2006 (SI Text). Salinity data, converted to δ18Owater data using relationships in Fig. S3, were combined with temperature to predict δ18Ocalcite values for WS oysters that were alive during this period (C).
Salinity sets the upriver boundary for modern James River oyster populations at approximately Deep Water Shoal, ~24 km downriver of JI (34–36) (Fig. 1). Although oysters can tolerate salinities as low as 2 psu for short periods (36, 37), salinities of 5–7 psu typically describe the low salinity threshold for James River oyster populations (22, 34, 36). Modern oyster distributions are also set by factors such as availability of oyster shell habitat, predators, diseases, and fishing.
The modern James River oyster distribution is in stark contrast to the distribution of early 17th century oyster reefs described by John Smith and his contemporaries from JI downriver to OPC (Tyndall, 1608, in ref. 14; Vingaboons, 1617, in ref. 13). These records, combined with colonial descriptions of brackish water at JI (15) and descriptions of foraminifera (Elphidium sp.) in marsh sediment cores from Kennon Marsh (38) (Fig. 1) indicate the 14-psu isohaline was upriver of JI from 1606 to 1612.
Modern Oyster δ18O Geochemistry.
Shell δ18O values from James River oysters are controlled by temperature (T) and salinity (S, as δ18Owater) (Eqs. 1 and 2). Warming T (late spring, early summer) usually occurs during periods of reduced S, whereas cooling T (fall, early winter) is associated with elevated S (Fig. 3). Because oyster δ18O varies with δ18Owater and T (Eq. 3), the inverse James River T vs. S distribution means that a 1-psu S decrease produces a 0.21–0.25‰ reduction in δ18Ocalcite (slope in Eqs. 1 and 2), whereas a 1 °C increase in T decreases δ18Ocalcite by 0.20–0.24‰, and vice versa. The modern seasonal James River T and S pairing that produces wetter conditions in spring and early summer results in an amplification of the oyster δ18Ocalcite signal as both environmental parameters serve to decrease or increase (during drier, cold winters) δ18Ocalcite.
Four WS oysters (Fig. 4 E–H) have shell δ18O values that display clear seasonal oscillations indicating onset of growth (settlement) by WS 8 and WS 16 during the summer of 2002, WS 2 in late fall 2002, and WS 9 in early summer 2003. Assuming one cold (δ18O maxima) and one warm period (δ18O minima) each year, these δ18O data document the age at terminal length relationship for modern WS oysters (Table 1 and Fig. S2). The isotope profiles in Fig. 4 record oyster growth from the outer (oldest) edge of the shell cross-section just above the resilium to the inner (most recent, 0 value) edge.
Fig. 4.

Measured δ18O data for historic (A–D) and modern (E–H) James River oysters.
Table 1.
Morphological and geochemical description of modern Wreck Shoal (WS) and historic Jamestown Structure 177 (JT) oyster shells
| Source | Shell ID | SL | SW | SL/SW ratio | Age (yr) | Avg δ18O | SD | δ18O min | δ18O max |
| WS | 2 | 110 | 86 | 1.28 | 4.33 | −4.74 | 1.12 | −5.63 | −2.84 |
| 8 | 111 | 75 | 1.48 | 4.33 | −4.26 | 1.37 | −5.59 | −2.64 | |
| 9 | 103 | 62 | 1.66 | 3.33 | −4.8 | 0.9 | −5.58 | −2.73 | |
| 16 | 113 | 80 | 1.41 | 4.33 | −4.55 | 1.29 | −5.64 | −2.58 | |
| JT | Z 1.2 | 148 | 94 | 1.57 | 3 | −3.5 | 1.09 | −4.4 | −1.35 |
| Z 2.1 | 129 | 74 | 1.72 | 3.5 | −3.62 | 1.58 | −4.73 | −0.97 | |
| P 5.4 | 119 | 90 | 1.32 | 3.83 | −3.38 | 1.46 | −4.64 | −1.18 | |
| P 14.1 | 101 | 68 | 1.49 | 2.83 | −3.5 | 1.19 | −4.43 | −1.16 | |
| U 9.2 | 121 | 86 | 1.41 | 7.75 | −3.52 | 1.1 | −4.52 | −1.42 | |
| U 13.5 | 118 | 76 | 1.55 | 8.75 | −3.4 | 0.8 | −4.42 | −1.97 |
Oxygen isotope data (Avg, SD, min, max) are presented graphically in Fig. S4. SL, shell length, maximum dimension from the hinge to the growth edge, mm; SW, shell width, maximum dimension perpendicular to shell length, mm; Age, age estimate based on oxygen isotope data corrected for collection month in years per Harding et al. (41); Avg, average; Max, maxima; Min, minima.
All modern oysters record increasing shell δ18O after the summer 2006 δ18O minimum, indicating that the shell geochemistry captured the initial seasonal T reduction during fall 2006 just before collection. If we (i) assign a S of ~18 psu to this time on the basis of measured salinities (Fig. 3B), (ii) use the average of the four final δ18Ocalcite values among the WS oysters, −4.32‰, for the collection geochemistry, (iii) convert S to δ18Owater with Eq. 1 and (iv) insert this average WS δ18Ocalcite and fall δ18Owater into Eq. 3 we obtain an ambient T prediction of 23.5 °C in excellent agreement with measured T at this time (Fig. 3A). Of importance here is our ability to accurately reconstruct the season of collection from these final data points as well as to demonstrate the robustness of the oxygen isotope thermometer.
These four WS shells record a combined δ18Ocalcite range from −1.3 to −6.7 ‰ (Fig. 4 E–H). Winter shell δ18O data capture all but the coldest estuarine temperatures (Figs. 3 and 4). Using the most positive predicted δ18Ocalcite value from winter 2004/2005 (Fig. 3C; −0.3‰) as being representative of winter conditions when water temperatures are 4 °C (Fig. 3A), we see that the shells did not record these coldest conditions; rather, the most positive shell δ18Ocalcite values are ~1‰ lower than predicted, suggesting the oysters stopped calcifying at a slightly higher T of ~8 °C. This calculation agrees with previous determinations of winter growth cessation in James River oysters (22, 34).
Minimum WS shell δ18O data are not as low as expected for oysters calcifying throughout the summer; instead, the lowest δ18Ocalcite value of ~ −6.7‰ is seen in three of the WS oysters during summer 2003 rather than the predicted δ18Ocalcite value of ~−8‰ (Fig. 4 E, F, and H). These data suggest Virginia oysters stop calcifying at an upper threshold T of 24–25°C (Figs. 3 and 4), which is close to the temperature of growth cessation determined for Florida oysters (~26–30 °C) (24). Most of the seasonal differences in minima (and maxima) δ18Ocalcite values in the WS oysters likely result from interannual shifts in S due to its control on δ18Owater (Fig. 3) because the seasonal T cycle at WS is similar from year to year (Fig. 3A). In summary, the δ18Ocalcite data from modern oysters demonstrate that we can (i) determine oyster age by counting seasonal δ18O cycles, (ii) identify the season of collection via the δ18O of the last calcite precipitated along the hinge margin, and (iii) determine that winter shell calcification ceases at ~8 °C and summer shell calcification stops at ~25 °C.
Historical Oyster δ18O and Archaeological Insights.
Seasonal cycles in δ18O are also observed in “historical” oysters from Structure 177 Z and P layers (Fig. 4 A–D). The final δ18O values in the P layer shells (Figs. 2 and 4 A and B) were incorporated following the decrease in shell δ18O that corresponds to spring warming. Because these final values are similar to the earlier summer δ18O minima, these data indicate the oysters were collected in early summer (~May or June) as water T rose above 24–25 °C (T of growth cessation). The final δ18O data from the U layer shells (Fig. 5) also record decreasing values after the winter δ18O maximum (March/April), suggesting the oysters were collected during spring and early summer. In contrast, the final Z layer δ18O data are considerably more positive and lack data from a spring warming, indicating a late fall/early winter collection near the seasonal δ18O maximum (December/January; Fig. 5). δ18O data obtained from the inner hinge edge of 10 additional Jamestown oysters from these three layers support these interpretations (Table S1). Paleobotanical data from the AA layer (19) (Fig. 2, directly underneath the Z layer) also suggest that this layer was deposited during late summer/early fall temporally preceding the Z layer and maintaining stratigraphic continuity.
Fig. 5.

Measured δ18O data from historic oyster shells from high-salinity (A and B) and medium-salinity (C and D) environments.
Given these δ18O data and historical accounts, we conclude that the oysters in the Z, U and P layers were collected during winter (December 1611/January 1612) and spring through early summer (April/June 1612), respectively. Geochemically, the isotope profiles for Z and P layer shells display similar amplitudes, suggesting exposure to similar growth conditions. If these oysters had been alive after 1612, we would expect to see a change in the δ18O signal reflecting changes in estuarine salinity with the end of the drought established by tree ring chronologies (16). The U layer oysters, which were alive before and throughout the drought (Fig. 5), display a unique δ18O profile that is discussed below.
Together, these data provide a coherent record of oyster collection between late 1611 and summer 1612. Given the location of the historic water table at the AA layer (19) (Fig. 2), the shells were put in the well after it had been abandoned. The lower position of Z layer, harvested in early winter 1611–1612, in the well shaft (Fig. 2) indicates that if Structure 177 is the Dale well dug in May 1611, it was open only for a very short time (~6 months or less). Structure 177 had a wooden frame made of rough hewn oak boards and a large amount of silt was found in the AB and AC layers (Fig. 2) during excavation (19). With the constant water pumping required to excavate the well, large quantities of silt probably entered the well through the boards with water (19). Because the Dale well would have gone into service in June 1611 during the drought, it is likely that large volumes of water were drawn from the well during summer 1611 to meet the needs of colonists and their livestock, particularly if a pump was placed on the large wooden wellhead frame. It is possible that the accompanying volume of sediment might have ruined the well in a very short time.
The possibility also exists that Structure 177 is an earlier well dating to 1609 or 1610 and not the Dale well. If this well was dug in 1609 or 1610, it would have been used for at least a year before abandonment, infill, and the deposition of Z layer. The presence of a ceremonial halberd from Lord De La Warre's guard in AB layer (19) is noteworthy in that the halberd was not in Virginia before June 1610 and would not have been discarded until after De La Warre's departure in April 1611 (14). Other artifacts indicate that Structure 177 was in use when the halberd was discarded (19). An older well, in use during May 1611, is referred to by Dale as “…a new well for the amending of the most unwholesome waters which the old afforded…(14, p. 523). The temporal sequence of fill for Structure 177 with late summer botanical material in AA and AB layers (19) combined with the oyster collections from Z (early winter 1611–1612), U and P layers (spring/early summer 1612) has parallels to the gradual conversion of an abandoned well to a trash pit beginning in mid-summer 1611. The season of harvest information from these shells combined with their stratigraphic sequence in the well describes either a 1611 well with a very short useful period or an undocumented older well abandoned in late 1611.
Historical Oyster Paleoenvironmental Implications.
The complete δ18O profiles obtained from Z and P layer oysters (Fig. 4 A–D) are similar to each other in terms of general seasonal trends and range of observed values (~0.3 to −5.7 ‰), suggesting that these oysters were collected from reef(s) with similar seasonal T and S conditions. This conclusion is also supported by a comparison of average minimum and maximum shell δ18O values between these shells, which indicates that the P and Z layer δ18O data are indistinguishable (Table 1 and Fig. S4).
Unlike the Z and P layer shells, the δ18O profiles from U layer oysters (Fig. 5 A and B) display seasonal cycles that reflect lower annual growth rates and, in general, record a narrower range of δ18O values. Although summer (minimum) U layer δ18O values are similar to Z and P layer δ18O minima (Figs. 4 A–D and 5 C and D), U layer winter maxima values are lower than either Z or P layer shells. If we assume that U layer oysters stopped calcifying below 8 °C and above 25 °C (similar to modern WS oysters), then the geochemical differences indicate that U layer oysters were collected from a reef that was exposed to less saline winter conditions than oysters from the Z and P layers. Thus, we will discuss the U layer shells separately.
Z and P layer oysters display an annual seasonal δ18O cycle that is similar in duration to modern WS oysters (Fig. 4). However, the average “historical” summer (−4.55 ± 0.16‰) and winter (−1.16 ± 0.16‰) values are 1.06‰ and 1.54‰ more positive, respectively, than corresponding average modern WS summer (−5.61 ± 0.03‰) and winter (−2.70 ± 0.16‰) values (Table 1 and Fig. S4). If we assume that growth was constrained by temperature limits of 25 °C and 8 °C, then we can compute summer and winter salinities by first computing δ18Owater from Eq. 3 and converting these values to salinity with Eq. 1. Here, we are assuming the δ18Owater vs. S relationship of the James River during the early 1600s was similar to that of modern times and that the most likely source(s) of the Z and P layer oysters were reefs near JI above MP (Fig. 1). The resulting average summer and winter salinities are 18.4 psu (δ18Owater = −2.48‰) and 16 psu (δ18Owater = −3.08‰), respectively. Whereas the calculated summer S is similar to that for the modern WS oysters during the drier summer season, the historic Jamestown winter S is considerably higher than modern. These data confirm a significant reduction in winter precipitation over the James River between 1609 and 1612 compared with precipitation between 2002 and 2006, in agreement with documentation of drought conditions at that time (16).
Drought of 1606–1612 and Implications for Historical Oyster Ecology.
Severe regional drought between 1606 and 1612 has been described independently using tree rings (16), benthic foraminifera (39, 40), and pollen data (25). The suggestion that regional salinities in the Chesapeake Bay increased by 10–15 psu during 1606–1612 relative to modern salinities (40) is in agreement with our winter salinity estimates from Z and P layer oysters. Thus, James River salinities between JI and MP (Fig. 1) would have been mesohaline (10–23 psu) year round during the 1606–1612 drought.
Maps by Tyndall (1608; 14) and Vingaboons (1617; 13) record oysters in the James River as far upriver as the mouth of the Chickahominy River above JI (Fig. 1). It is likely that the severity and duration of the 1606–1612 drought would have facilitated the upriver expansion of oyster habitat from MP toward JI. The drought would have shifted the James River salinity gradient upriver through a combination of increased tidal incursion and a 40–50% reduction in freshwater discharge (40). Beginning in 1606, drought induced salinities of 14 psu (38) between JI and MP would have encouraged oyster recruitment. By 1608, reefs could have formed near Jamestown containing oysters suitably sized for food (~2-yr-old oysters, SI Text) (41) within easy access for the colonists.
Environmental and Historical Implications of U Layer Oyster Geochemistry.
Based on the stratigraphic relationship of the U layer between Z and P layers and our reconstruction of the collection timing of the Z and P layer oysters, we infer that U layer shells were harvested during spring/early summer 1612, given the range of hinge edge values (Fig. 5 A and B and Table S1). Age estimates using winter δ18O maxima indicate that U layer oysters were older at collection than the other oysters examined (Table 1) and appear to have settled before the drought began (1604 or 1605; Fig. 5).
Of particular interest is the uniqueness of the U layer δ18O records relative to WS, Z and P layer oysters. First, the U layer δ18O records contain 5–7 yr of seasonal signal in contrast to the 3–4 yr of growth documented in the other shells (Figs. 4 and 5). Second, although the average δ18O minima (summer, −4.47 ± 0.07‰) in oysters U 9.2 and 13.5 are indistinguishable from the Z and P layer shells, the average δ18O maxima (winter, −1.70 ± 0.39‰) are more negative than Z and P layer maxima (>0.5‰; Table 1 and Fig. S4). This suggests fresher winter conditions or minimum temperatures that did not drop below 8 °C (Table 1 and Figs. 4 and 5). We prefer the former explanation, as it is unlikely that winter/summer extreme temperatures varied much across the James River oyster habitat. Oxygen isotope data from Z and P layer shells are in agreement with modern water temperature and oxygen isotope data which show that James River temperatures are below 8 °C for at least a short period each year (Fig. 3 and Fig. S5). An intertidal harvest site in a small watershed close to the James River confluence with the Chesapeake Bay could explain the U layer δ18O signal.
Shell U9.2 (Fig. 5A) displays δ18O values and growth cycles of the same timing and magnitude (0 to −5‰) predicted by the annual salinity cycle at OPC (Figs. 1 and S5C), although the δ18O pattern recorded in shell U13.5 displays a smaller range than that in U9.2 (comparing Fig. 5 A vs. B). The maxima of −0.5‰ displayed by shell U9.2 in the winters of 1609–1610 and 1607–1608 are absent in U13.5. If U13.5 grew closer to the air–water interface on the source intertidal oyster reef than U9.2, this shell would have been exposed at low tide more often and for longer periods than U9.2. Exposure to air and relatively shallow water would be commensurate with longer and more intense exposure to temperatures above or below the 8–25 °C window required for calcification, thereby resulting in a truncated or compressed growth trajectory of U13.5 relative to U9.2 and the Z and P layer shells. Furthermore, shallow exposure would have exposed the oysters to the less saline near surface estuarine layer, thereby explaining the reduced U layer winter δ18O maxima values.
Given the temporal relationship of the Z, U and P layers combined with the unique U layer stable isotope records, we hypothesize that the U layer oysters were transported to Jamestown from a downriver tributary source closer to the James River confluence with the Chesapeake Bay. By 1612, the colonists had established settlements between the Hampton River and Mill Creek (Ft. Algernoune; Fig. 1) and on the Nansemond River (17) (Fig. 1) and had contact with Native American settlements in the Elizabeth River drainage (Fig. 1). All four of these tributaries historically supported oysters (14, 42). Upon their arrival in the Chesapeake Bay, ships from England typically stopped at Ft. Algernoune (Fig. 1) before proceeding upriver to Jamestown. Both Nansemond and Hampton River oysters would have been accessible to supply ships moving upriver from Ft. Algernoune. The region around Ft. Algernoune was also identified by colonists as a reliable source of oysters, crabs, and fish because of its proximity to the Bay and the Atlantic Ocean (14). The colony had a shallow draft “barge” (8) that could have been used to harvest and transport shellfish upriver independently of larger supply ships. By spring 1612, the number of colonists at Jamestown had increased, whereas their mortality rate had decreased (15). This net population increase combined with years of harvest pressure was probably a strain on the oysters near JI and transport of seafood upriver by ships was likely necessary to keep Jamestown adequately supplied.
Conclusion
The James River was a different habitat in the early 17th century than it is today. Oyster shells from James Fort Structure 177 document estuarine conditions during the 1606–1612 drought, thereby providing an estuarine analog to the terrestrial bald cypress chronologies (16) as well as a unique view of the James River and Jamestown during the earliest years when the colony was fighting to survive. Although the drought presented numerous challenges for the colonists, conditions in the estuary may have facilitated oyster growth and extension of habitat upriver as more saline conditions migrated above JI. Despite reefs near JI, the colonists appear to have augmented local resources with oysters from reefs near the Chesapeake Bay. The oyster shell season of harvest information combined with their stratigraphic relationships in the well describe either a 1611 well with a very short useful period or an undocumented older well abandoned by late 1611. Because oysters are finely tuned stationary barometers in historic and modern estuarine environments, they are a critical source of paleoecological, paleoenvironmental, and archaeological information for studies of early colonization of the New World.
Materials and Methods
Oyster Shell Sources.
In spring 2006, APVA archaeologists excavating a Jamestown well (APVA Structure ID 177, Fig. 1) categorized material that was removed into 29 layers (Fig. 2) on the basis of its apparent coherence within geologic features and archaeological context (19). Layers AC (deepest, bottom of well) through D (shallow) correspond to a coherent time period within the colony between 1609 and 1616 (19). There was no mixing of artifacts from within the well with those from later time periods beginning with layer N (19) (Fig. 2). Intact left oyster shell valves were selected from three well layers: P (n = 6, relatively shallow), U (n = 6, intermediate), and Z (n = 4, deep) for stable isotope analyses. These layers were selected because they span the available depth profile and each had a relatively large number of intact left valves (483, 726, and 166 intact valves, respectively for the P, U, and Z layers).
For comparative purposes, live oysters were collected from WS (n = 4) during November 2006 and killed to obtain modern left oyster valves with a known collection date. Only left valves with a straight resilium were used in this study (41). Oyster shells were embedded and sectioned along a straight axis extending from the hinge to the growth edge through the resilium parallel to the axis of maximum growth (23, 44).
Geochemical Sample Collection and Analyses.
Oyster shell sections were mounted on a computer-aided triaxial sampler (45). For two shells from each layer, samples were collected every 0.3 mm along a 0.8-mm-wide path drilled near the resilium from the outer (oldest) to the inner (youngest) layer. Transects were drilled perpendicular to growth bands using a 0.5-mm carbide dental burr. In the remaining shells from each layer, samples were collected from the most recent 10 mm of the internal shell margin to encompass the last ~6 months of life. Powdered shell calcite was collected and roasted in vacuo for 30 min at 375 °C before analysis on a Fisons Optima isotope ratio mass spectrometer using an Isocarb common acid bath device. Acid reaction temperature was 90 °C. Water δ18O was determined via CO2 equilibration using an automated equilibrator attached to a Finnigan MAT 251 isotope ratio mass spectrometer.
All oxygen isotope data are presented relative to the Vienna Pee Dee Belemnite (V-PDB) standard, whereas water oxygen isotope data are presented relative to Vienna Standard Mean Ocean Water (V-SMOW) standard. Data are presented in standard per mil (‰) notation where:
Analytical precision (±1 σ) of the carbonate δ18O data was ±0.06‰ based on repeat analyses of an in-house calcite standard. The precision of water replicates was ±0.03‰ (±1 σ).
Data Analyses: Relationship Between Salinity and δ18Owater.
Water samples from WS and MG in the James River (Fig. 1) were analyzed for salinity and stored in sealed vials for δ18O analyses (SI Text). Relationships between salinity and δ18Owater (=δ18Ow) were calculated for WS and MG waters (Fig. S3) using linear regression analyses, yielding:
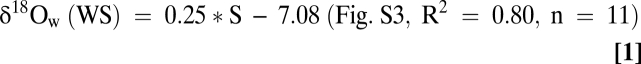 |
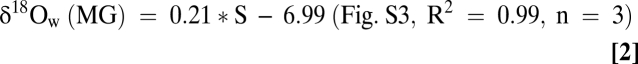 |
We compute a predicted δ18Ocalcite (=δ18Oc) time series for oysters growing between 2002–2006 by combining water temperatures (T) from WS (SI Text) and δ18Ow values computed from Eq. 1 by inverting the T vs. δ18Ocalcite relationship of Epstein et al. (46).
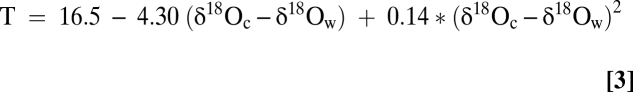 |
to yield
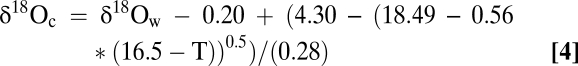 |
Here, 0.20‰ is subtracted from δ18Ow to correct for the V-PDB and V-SMOW differences in this relationship (47). Similar predictions are made for Old Point Comfort (Fig. 1) using Eq. 2.
Supplementary Material
Acknowledgments
We are grateful to the Association for the Preservation of Virginia Antiquities staff including W. Kelso, B. Straube, D. Schmidt, D. Givens, M. Gavin, D. Gamble, J. May, and C. Taylor for providing access to the oysters as well as discussions on archaeology and Jamestown with J.M.H. We appreciate the insight and historical perspective offered by M. Nichols (Virginia Institute of Marine Science) on the James River estuary as well as access to some of his original foraminifera core figures. We thank D. Winter and J. Oster (University of California–Davis) for assistance with stable isotope analyses and shell sampling, and M. Southworth and M. Harris (Virginia Institute of Marine Science) for collecting modern oyster and water samples. During a part of this project, H.J.S. was supported by the National Science Foundation (NSF) while he worked at the Foundation. We thank three anonymous reviewers and W. Kelso and B. Straube for comments that significantly improved the final version of this manuscript. This is Contribution 3093 from the Virginia Institute of Marine Science.
Footnotes
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1001052107/-/DCSupplemental.
References
- 1.Hargis WJ., Jr. In: Oyster Reef Habitat Restoration: A Synopsis of Approaches. Luckenbach MW, Mann R, Wesson JA, editors. Gloucester Point, VA: Virginia Institute of Marine Science; 1999. pp. 5–24. [Google Scholar]
- 2.Hobbs C. Geological history of Chesapeake Bay, USA. Quat Sci Rev. 2004;23:641–661. [Google Scholar]
- 3.Wells H. The fauna of oyster beds, with special reference to the salinity factor. Ecol Monogr. 1961;31:239–266. [Google Scholar]
- 4.Coen L, Luckenbach M, Breitburg D. In: Fish Habitat: Essential Fish Habitat and Rehabilitation. Benaka LR., editor. Bethesda, MD: Am Fish Soc Symp 22; 1999. pp. 438–454. [Google Scholar]
- 5.Harding JM, Mann R. Fish species richness in relation to restored oyster reefs, Piankatank River, Virginia. Bull Mar Sci. 1999;65:289–300. [Google Scholar]
- 6.DeAlteris J. The geomorphic development of Wreck Shoal, a subtidal oyster reef of the James River, Virginia. Estuaries. 1988;11:240–249. [Google Scholar]
- 7.Bahr L, Lanier W. The Ecology of Intertidal Oyster Reefs of the South Atlantic Coast: A Community Profile. US FWS, Washington, DC: US Fish and Wildlife Series; 1981. FWS/OBS-81/15. [Google Scholar]
- 8.Rountree HC, Clark W, Mountford K. John Smith's Chesapeake Voyages: 1607-1609. Charlottesville: Univ of VA Press; 2007. pp. 12–15.pp. 48pp. 66–67.pp. 131 [Google Scholar]
- 9.Newell RIE. In: Understanding the Estuary: Advances in Chesapeake Bay Research. Lynch MP, Krome EC, editors. Gloucester Point, VA: Chesapeake Research Consortium; 1988. pp. 536–546. [Google Scholar]
- 10.Ulanowicz RE, Tuttle JH. The trophic consequences of oyster stock rehabilitation in Chesapeake Bay. Estuaries. 1992;15:298–306. [Google Scholar]
- 11.Newell RIE, Fisher TR, Holyoke RR, Cornwell JC. In: The Comparative Roles of Suspension Feeders in Ecosystems. Dame R, Olenin S, editors. Vol 47. The Netherlands: Springer; 2005. pp. 93–120. NATO Series: IV—Earth and Environmental Sciences. [Google Scholar]
- 12.Hatch CE. The First Seventeen Years: Virginia, 1607-1624. Charlottesville: Univ of VA Press; 1957. pp. 4–34. [Google Scholar]
- 13.Kelso W. Jamestown: The Buried Truth. Charlottesville: Univ of VA Press; 2006. pp. 11–27.pp. 89–93.pp. 115–124. [Google Scholar]
- 14.Haile EW. Jamestown Narratives: Eyewitness Accounts of the Virginia Colony. Champlain, VA: Roundhouse Publishers; 1998. pp. 96–100.pp. 118–120.pp. 128–129.pp. 319–343.pp. 416–431.pp. 497–519. [Google Scholar]
- 15.Earle CV. In: The Chesapeake in the Seventeenth Century: Essays on Anglo-American society. Tate T, Ammerman D, editors. Chapel Hill: Univ of NC Press; 1979. pp. 97–125. [Google Scholar]
- 16.Stahle DW, Cleaveland MK, Blanton DB, Therrell MD, Gay DA. The lost colony and Jamestown droughts. Science. 1998;280:564–567. doi: 10.1126/science.280.5363.564. [DOI] [PubMed] [Google Scholar]
- 17.Brown A. The First Republic in America, An Account of the Origin of this Nation Written from Records then (1624) Concealed by the Council rather than from the Histories then Licensed by the Crown. New York: Houghton Mifflin; 1898. pp. 68–125. [Google Scholar]
- 18.Kelso W, Straube B. Jamestown Rediscovery 1994-2004. Richmond, VA: Association for the Preservation of Virginia Antiquities; 2004. pp. 131–135. [Google Scholar]
- 19.Kelso W, Straube B, editors. 2000-2006 Interim Report on the APVA excavations at Jamestown, Virginia. Richmond, VA: Association for the Preservation of Virginia's Antiquities; 2008. pp. 55–69. [Google Scholar]
- 20.Brooks WK. The Oyster. Baltimore: Johns Hopkins Univ Press; 1891. p. 230. [Google Scholar]
- 21.Galtsoff P. US Fish Wildlife Service Fish Bull 64. 1964. The American Oyster Crassostrea virginica Gmelin; pp. 397–456. [Google Scholar]
- 22.Mann R, Evans D. Site selection for oyster habitat rehabilitation in the Virginia portion of the Chesapeake Bay. J Shellfish Res. 2004;23:41–49. [Google Scholar]
- 23.Kirby MX, Soniat T, Spero HJ. Stable isotope sclerochronology of Pleistocene and recent oyster shells (Crassostrea virginica) Palaios. 1998;13:560–569. [Google Scholar]
- 24.Surge D, Lohmann K, Dettman D. Controls on isotopic chemistry of the American oyster, Crassostrea virginica: Implications for growth patterns. Palaeogeogr Palaeoclimatol Palaeoecol. 2001;172:283–296. [Google Scholar]
- 25.Brush G. Natural and anthropogenic changes in Chesapeake Bay during the last 1000 years. Hum Ecol Risk Assess. 2001;7:1283–1296. [Google Scholar]
- 26.Brush G. Geology and Paleoecology of Chesapeake Bay: A long-term monitoring tool for management. J Wash Acad Sci. 1986;76:146–160. [Google Scholar]
- 27.Cooper S, Brush G. A 2,500 year history of anoxia and eutrophication in Chesapeake Bay. Estuaries. 1993;16(3B):617–626. [Google Scholar]
- 28.Colman S, Bratton J. Anthropogenically induced changes in sediment and biogenic silica fluxes in Chesapeake Bay. Geology. 2003;31:71–74. [Google Scholar]
- 29.Cornwell J, Conley D, Owens M, Stevenson J. A sediment chronology of the eutrophication of the Chesapeake Bay. Estuaries. 1996;19:488–499. [Google Scholar]
- 30.Zimmerman A, Canuel E. Sediment geochemical records of eutrophication in the mesohaline Chesapeake Bay. Limnol Oceanogr. 2002;47:1084–1093. [Google Scholar]
- 31.Kennedy V. The ecological role of the eastern oyster, Crassostrea virginica, with remarks on disease. J Shellfish Res. 1996;15:177–183. [Google Scholar]
- 32.Nichols M, editor. Sediments of the James River Estuary, Virginia. New York, NY: Geological Society of America; 1972. pp. 169–212. The Geological Society of America. The GSA Memoir 133. [Google Scholar]
- 33.Brooks T, Fang C. Gloucester Point, VA: Virginia Institute of Marine Science; 1983. James River slack water data report: Temperature, salinity, and dissolved oxygen 1971-1980. VIMS Data Report No. 12. [Google Scholar]
- 34.Mann R, Southworth MJ, Harding JM, Wesson JA. Population studies of the native oyster, Crassostrea virginica (Gmelin, 1791), in the James River, Virginia, USA. J Shellfish Res. 2009;28:193–220. [Google Scholar]
- 35.Haven DS, Whitcomb JP. The origin and extent of oyster reefs in the James River, Virginia. J Shellfish Res. 1983;3:141–151. [Google Scholar]
- 36.Mann R, Evans D. Estimation of oyster, Crassostrea virginica, standing stock, larval production and advective loss in relation to observed recruitment in the James River, Virginia. J Shellfish Res. 1998;17:239–254. [Google Scholar]
- 37.Gunter G. Seasonal population changes and distributions as related to salinity, of certain invertebrates of the Texas coast, including the commercial shrimp. Publ Inst Mar Sci Univ Texas. 1950;1:7–51. [Google Scholar]
- 38.Ellison RL, Nichols M, editors. Halifax, Nova Scotia: Dalhousie University; 1976. Modern and holocene foraminifera in the Chesapeake Bay region. Maritime Sediments Special Publication 1. 1st International Symposia on Bethonic Foraminifera of Continental Margins; pp. 131–151. [Google Scholar]
- 39.Karlsen A, et al. Historical trends in Chesapeake Bay dissolved oxygen based on benthic foraminifera from sediment cores. Estuaries. 2000;23:488–508. [Google Scholar]
- 40.Cronin T, et al. Climactic variability in the eastern United States over the past millenium from Chesapeake Bay sediments. Geology. 2000;28:3–6. [Google Scholar]
- 41.Harding JM, Mann R, Southworth MJ. Shell length-at-age relationships in James River oysters (Crassostrea virginica) collected four centuries apart. J Shellfish Res. 2008;27:1109–1115. [Google Scholar]
- 42.Baylor JB. Method of Defining and Locating Natural Oyster Beds, Rocks and Shoals. Oyster Records (pamphlets, one for each Tidewater, Virginia county) Richmond, VA: Board of Fisheries of VA; 1896. [Google Scholar]
- 43.Surge D, Lohmann K, Goodfriend G. Reconstructing estuarine conditions: Oyster shells as recorders of environmental change, Southwest Florida. Estuar Coast Shelf Sci. 2003;57:737–756. [Google Scholar]
- 44.Harding JM, Mann R. Age and growth of wild suminoe (Crassostrea ariakensis, Fugita 1913) and Pacific (C. gigas, Thunberg 1793) oysters from Laizhou Bay, China. J Shellfish Res. 2006;25:73–82. [Google Scholar]
- 45.Quinn T, Taylor F, Crowley T, Link S. Evaluation of sampling resolution in coral stable isotope records: A case study using records from New Caledonia and Tarawa. Paleoceanography. 1996;11:529–542. [Google Scholar]
- 46.Epstein S, Buchsbaum R, Lowenstam H, Urey H. Revised carbonate water isotopic temperature scale. Geol Soc Am Bull. 1953;2:417–425. [Google Scholar]
- 47.Bemis BE, Spero HJ, Bijma J, Lea DW. Reevaluation of the oxygen isotope composition of planktonic foraminifera: Experimental results and revised paleotemperature equations. Paleoceanography. 1998;13:150–160. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.