

Abstract
Early embryonic imaging of cardiovascular development in mammalian models requires a method that can penetrate through and distinguish the many tissue layers with high spatial and temporal resolution. In this paper we evaluate the capability of Optical Coherence Tomography (OCT) technique for structural 3D embryonic imaging in mouse embryos at different stages of the developmental process ranging from 7.5 dpc up to 10.5 dpc. Obtained results suggest that the collected data is suitable for quantitative and qualitative measurements to assess cardiovascular function in mouse models, which is likely to expand our knowledge of the complexity of the embryonic heart, and its development into an adult heart.
Keywords: Optical Coherence Tomography, non-invasive, mouse embryos, cardiovascular function
1. Introduction
During embryonic development, the establishment of the body plan and the formation of functional organs require dynamic changes in the cellular architecture. The ability to directly image these complex morphogenetic movements has aided greatly in our understanding of normal development as well as congenital abnormalities and mutant analysis in model systems. Vital imaging is very valuable but can also be very challenging. Vertebrate embryos are generally very small, requiring high spatial resolution (< 10 μm) to follow cell movements and distinguish tissue layers. Also, although some embryos, such as Zebrafish embryos, are transparent, others, such as mouse embryos, are quite turbid, which makes it difficult to image in depth through many tissue layers. Moreover, the speed at which processes such as heart contraction occur can be a challenge for the temporal resolution of many imaging systems. Although these challenges exist, recent improvements in imaging methodology have elevated the level of analysis that can be performed in several embryo species. Such advances have had a tremendous impact on understanding many aspects of development, but in particular, in the field of early cardiovascular development.
Many different methods have been used to image early cardiovascular development. Methods such as episcopic imaging and Optical Projection Tomography permit excellent analysis of the three-dimensional (3D) structure of embryos as they allow to label specific structures with fluorescent markers.1,2 Nevertheless, the tissue must be fixed for both methods and sectioned in the case of episcopic techniques. Other methods such as high-frequency ultrasound biomicroscopy and MRI are non-destructive and can be used to image complete volumes of whole embryos and specific regions therein, but suffer from low spatial resolution (∼ 30–50 μm), which makes the analysis of individual cells and thin tissue layers difficult. MRI is also limited by poor temporal resolution as it takes minutes to hours to collect full 3D volumes, particularly if high spatial resolution is required. Conversely, optical techniques such as confocal imaging offer excellent spatial (sub-micron lateral and axial) resolution and line scanning confocal systems enable very high temporal resolution while preserving the high axial resolution. This has enabled, for example, the capture of 3D volumes of the beating heart in Zebrafish embryos.3–5 Confocal and line-scanning confocal imaging have also been used to characterize the onset of circulation in early heart function in live, cultured mouse embryos;6–8 however, due to poor depth penetration it is not possible to acquire full 3D stacks of the entire heart of mouse embryo or to study flow patterns in vessels deep within the embryo. Selective plane illumination microscopy (SPIM) has also been used to image the beating heart in Zebrafish embryos. Although this technique extends the imaging depth, it requires that embryos be embedded in agarose for the tomography-like image collection from multiple angles, which is currently incompatible with murine embryo culture. Such fluorescence imaging methods have the advantage that live transgenic embryos can be used in which specific tissues express fluorescent proteins. For some species such as mouse, however, it can be time consuming and expensive to cross such markers into every mutant line that one wishes to image. Thus, for imaging early cardiovascular development and function in mouse embryos, there is a great need for imaging methods that can penetrate through and distinguish the many tissue layers with high spatial and temporal resolution for both the analysis of normal cardiovascular development and for the analysis of mutant animals. Recently our work and the work of others have shown that Optical Coherence Tomography (OCT) fits many of these requirements and is likely to offer the best compromise between spatial resolution and depth penetration of any available method for imaging chicken, frog, zebrafish, and mouse embryos.9–22
Frequently, techniques of gated embryonic cardiography are used for four-dimensional (4D) OCT imaging of heart dynamics in order to compensate for limited signal acquisition rate.13 Recent advances in the development of swept source OCT technology (SS-OCT) have enabled acquisition speeds at up to 115 kHz for conventional wavelength-swept lasers.23 Introduction of a new class of Fourier Domain Mode Locked (FDML) lasers24,25 allowed achieving sweeping speeds up to 370 kHz.26,27 Consequently, imaging of the embryonic avian heart has been demonstrated at 10 volumes/s (3D) without the aid of gating.15
For OCT imaging of early mouse physiology, embryos must be removed from the uterus or exposed for imaging due to significant light attenuation in the body wall. Thus, extensive expertise is required to maintain physiologically normal embryos to obtain meaningful measurements. In this paper we evaluate the capability of OCT for structural 3D embryonic imaging in mouse embryos at different stages of the developmental process ranging from 7.5 days post coitum (dpc) (0.5 is noon on the day of the vaginal plug) up to 10.5 dpc.
2. Materials and Methods
A schematic diagram of the Swept Source OCT (SS-OCT) system used in these studies is shown in Fig. 1. The current system employs a broadband swept-source laser (Thorlabs, SL1325-P16) with output power p = 12 mW at central wavelength λ0 = 1325 nm and spectral width Δλ = 110 nm. The scanning rate over the full operating wavelength range is 16 kHz. The light from the swept-source laser source is delivered to an interferometer where it is split into a reference arm and a sample arm in a 10/90 ratio. Light that returns from the sample and reference arms forms an interferogram. The latter is detected by a balanced-receiver configuration that reduces source intensity noise as well as auto-correlation noise from the sample (Thorlabs, PDB140C) and is digitized using a 14-bit analog-to-digital converter (ADC). A periodic optical filter, Mach-Zehnder interferometer (MZI)-based optical clock, which generates an equally spaced interferogram in frequency from a small portion of the source output, is used to re-map the signal from the optical time-delay domain in the frequency-domain before application of Fast Fourier Transform (FFT) algorithms. The FFT reconstructs an OCT intensity in-depth profile (A-scan) from a single scan over the operating wavelength range, resulting in an A-scan acquisition rate of 16 kHz. By scanning across the sample surface in the X and Y directions, 2D and 3D data sets are collected. The scanning head of the SS-OCT system is positioned inside a commercial 37°C CO2 incubator.
Fig. 1.
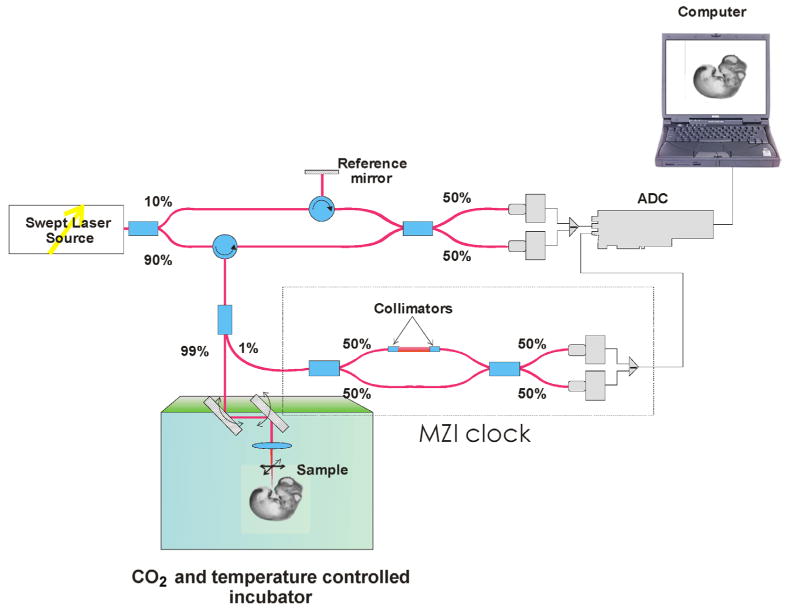
SS-OCT experimental setup for embryo imaging.
Wild type CD-1 embryos were dissected with the yolk sac intact at 7.5 to 10.5 dpc in the preheated dissecting medium (37°C) as previously reported for confocal microscopy.7 The imaging station was maintained at 37°C using a commercial 37°C CO2 incubator (VMD3001S9ABA, Symphony Inc.) and embryos were cultured in a rich culture media containing 50% home-made rat serum as described in Ref. 7. Live embryos can be maintained on the imaging stage with normal heart rates for at least 24 hours by maintaining temperature and proper gas environment.7
The basic reconstruction algorithm used in this study was previously successfully applied for 4D analysis of embryonic zebrafish heart, where imaging was performed by fast scanning confocal microscopy of vital fluorescent markers.4,5 For 4D reconstruction of the embryonic heartbeat, non-gated XZ time sequences were acquired for 10 cardiac cycles at a fixed Y position, repeating this process for multiple Y positions spaced 10 μm apart, for a total of 42 positions (420 μm). The acquisition rate for the XZ-sequences was 30 fps. Because of the repetitive nature of the heart movement, all cycles of the heartbeat were assumed identical within the total acquisition time. Each time lapse was de-noised by averaging between different cycles within the time lapse, resulting in a single de-noised heartbeat cycle, and then all the time lapses were synchronized using previously reported algorithms,4,5 that resulted in a 4D representation of the beating embryonic heart.
3. Results and Discussion
Figure 2 shows cross-sections of typical examples of 3D reconstructions of live embryos at different stages of development obtained by SS-OCT scanning. The whole embryo and specifically the heart undergoes dramatic changes in anatomy and function during each day of early development. Figure 2(a) shows an embryo at 7.5 dpc when embryonic heart just starts to form. At this stage cardiac progenitors migrate to the midline of the embryo to form a tube consisting of an endocardial layer surrounded by cardiomyocytes. Heart development coincides with the development of embryonic and extra-embryonic vessels and the formation of primitive erythroblasts in the proximal yolk sac. At the embryonic day 8.5 [Fig. 2(b)] the heart tube starts to contract and circulate plasma and in a few hours blood cells enter the circulation through the premature vascular system. Figure 2(c) shows an embryo just 24 hours later, at 9.5 dpc. During this time the embryo grows significantly and turns so that the heart moves away from the yolk sac, while the heart tube loops, and the vasculature remodels into more mature branching circulatory system, which is visible in the image. Further development continues as chambers and valves emerge. These transformations, which lead to the formation of a four-chambered heart, occur while the heart is functional and while circulation switches between nutrient sources, first the yolk sac, then the placenta and finally the lungs. As one can see from the images, many recognizable structures of the embryo are clearly outlined, as labeled, and the spatial resolution of the SS-OCT system (∼6μm in tissues) provides excellent details. For instance, the amnion labeled in (a) and (c) is only two-cell-layer thick and is clearly visible, even in very early 7.5 dpc embryos.
Fig. 2.
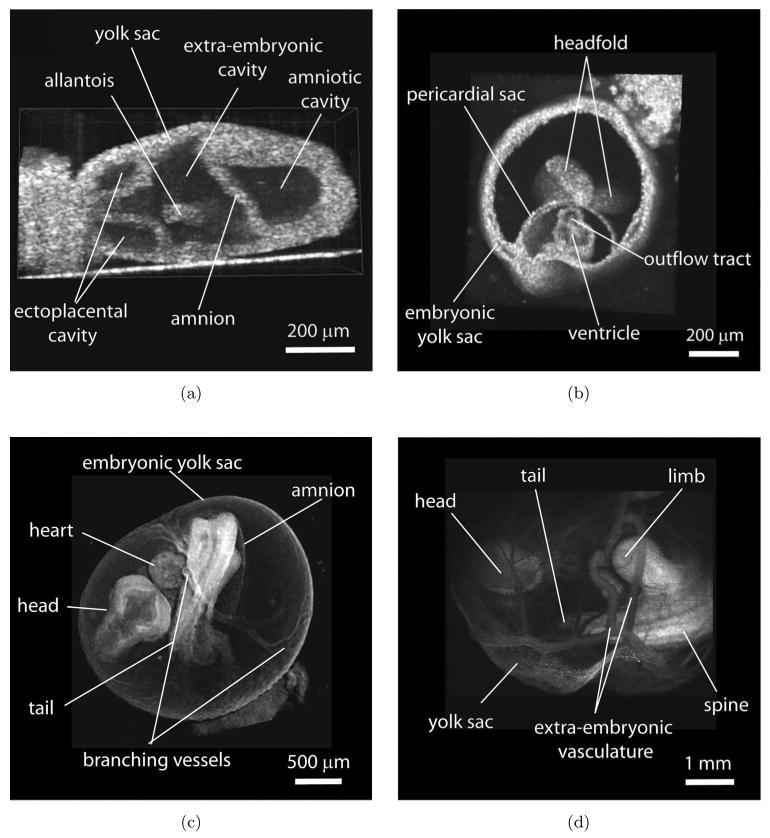
Structural imaging of early mouse embryos with SS-OCT. (a)–(d) 3D reconstructions of live embryos cultured with the yolk sac at 7.5 dpc, 8.5 dpc, 9.5 dpc, and 10.5 dpc, respectively.
As the embryo continues to grow, the development of mature extra-embryonic vasculature causes significant light attenuation and limits OCT imaging of the heart region, as seen in Fig. 2(d). Therefore, it might be challenging to image and characterize the dynamics of the heart at later stages of the development, i.e., above 10.5 dpc, and it requires careful positioning of the embryo on the imaging stage in order to access the region of interest. While removal of the yolk sac can provide much better structural information with excellent contrast and resolution as shown in Fig. 3, this procedure disrupts the blood circulatory network and, therefore, is not compatible with live imaging. To overcome this limitation, an incision in the yolk sac can be carefully made without disruption of major blood vessels to expose the organ of interest for the imaging, but this approach is technically challenging. Alternatively, optical clearing methods might potentially be utilized in order to enhance the OCT signal from the internal structures.28,29
Fig. 3.
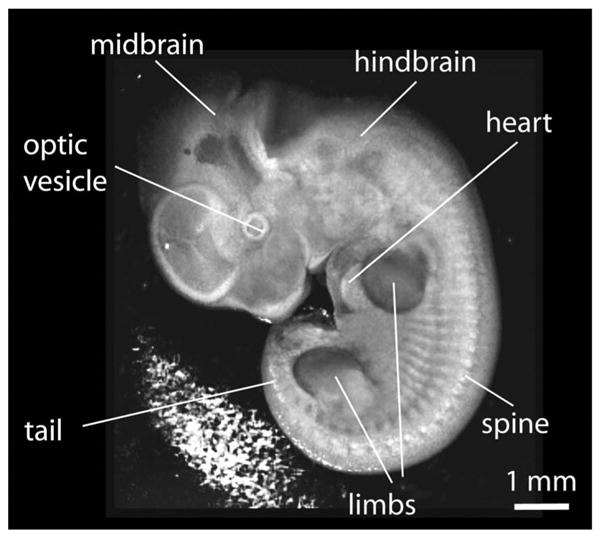
A snapshot from structural 3D image of mouse embryo at 10.5 dpc with the yolk sac removed.
We next focused on testing the possibility for reconstruction of live mouse embryonic heartbeat. The frame rate can be increased by decreasing the number of A-scans per frame and limiting the field of view precisely to the heart area, but an acquisition of a single volume of structural data consisting of 128 by 128 A-scans (X–Y) still takes about a second for 16-kHz SS-OCT system. Therefore, sequential acquisition of the volumes does not allow for a reasonable frame rate to analyze heart wall dynamics. In order to rapidly image live embryonic hearts in 4D structure, OCT can be gated to the stimulus of the heartbeat in order to sequence the temporal phases of the cardiac cycle.13,16 Non-gated reconstructions are advantageous because additional hardware for gating is not required and litters of embryos can be more rapidly analyzed if alignment with a gating signal is not a concern; however, non-gated reconstruction requires adequate sampling and can be prone to error if there is any arrhythmia.5 Here we used non-gated 4D reconstruction algorithm of the embryonic heartbeat as described above. Figure 4(a) shows representative frames from the reconstruction corresponding to different Y positions after noise reduction and synchronization, and Fig. 4(b) is a cross-sectional view of the same reconstruction. This result is the first 4D reconstruction of mammalian embryonic heartbeat in live embryo culture with OCT. The reconstruction provides sufficient details about tissue structure and might permit tracking of the heart wall movements during the cardiac cycle.
Fig. 4.
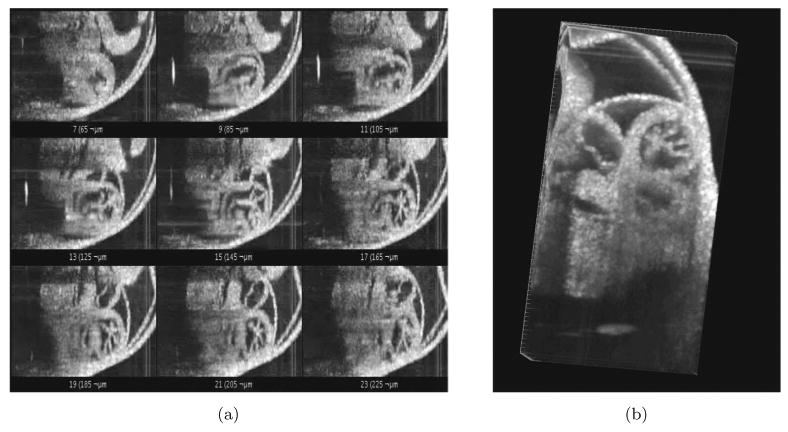
4D reconstruction of embryonic cardiodynamics. (a) A snapshot from a gallery of de-noised and synchronized time lapses acquired at different Y positions and used for 4D reconstruction of the heartbeat. (b) A snapshot from the corresponding 4D reconstruction.
4. Conclusions
In this paper, we presented structural 3D reconstructions of live mammalian embryos from 7.5 dpc stage, when the heart just starts to form, up to 10.5 dpc stage when the major organs are formed. The 4D reconstruction of the beating embryonic heart was performed by post-acquisition synchronization of non-gated time lapses sequentially acquired at different positions. The results from these studies demonstrate that SS-OCT combined with live static embryo culture is suitable for structural imaging of live mammalian embryos and functional characterization of beating embryonic heart, which potentially can expand our understanding of the dynamic complexity of the embryonic development.
Acknowledgments
The study was supported, in part, by Postdoctoral Fellowship from the AHA (IVL), grants from the NIH (HL077187 MED), and Office of Naval Research (KVL).
References
- 1.Ruffins SW, Jacobs RE, Fraser SE. Towards a Tralfamadorian view of the embryo: Multidimensional imaging of development. Curr Opin Neurobiol. 2002;12:580–586. doi: 10.1016/s0959-4388(02)00366-5. [DOI] [PubMed] [Google Scholar]
- 2.Sharpe J, Ahlgren U, Perry P, Hill B, Ross A, Hecksher-Sorensen J, Baldock R, Davidson D. Optical projection tomography as a tool for 3D microscopy and gene expression studies. Science. 2002;296:541–545. doi: 10.1126/science.1068206. [DOI] [PubMed] [Google Scholar]
- 3.Forouhar AS, Liebling M, Hickerson A, Nasiraei-Moghaddam A, Tsai HJ, Hove JR, Fraser SE, Dickinson ME, Gharib M. The embryonic vertebrate heart tube is a dynamic suction pump. Science. 2006;312:751–753. doi: 10.1126/science.1123775. [DOI] [PubMed] [Google Scholar]
- 4.Liebling M, Forouhar AS, Gharib M, Fraser SE, Dickinson ME. Four-dimensional cardiac imaging in living embryos via postacquisition synchronization of nongated slice sequences. J Biomed Opt. 2005;10:054001. doi: 10.1117/1.2061567. [DOI] [PubMed] [Google Scholar]
- 5.Liebling M, Forouhar AS, Wolleschensky R, Zimmermann B, Ankerhold R, Fraser SE, Gharib M, Dickinson ME. Rapid three-dimensional imaging and analysis of the beating embryonic heart reveals functional changes during development. Dev Dyn. 2006;235:2940–2948. doi: 10.1002/dvdy.20926. [DOI] [PubMed] [Google Scholar]
- 6.Jones EA, Baron MH, Fraser SE, Dickinson ME. Measuring hemodynamic changes during mammalian development. Am J Physiol Heart Circ Physiol. 2004;287:H1561–1569. doi: 10.1152/ajpheart.00081.2004. [DOI] [PubMed] [Google Scholar]
- 7.Jones EA, Crotty D, Kulesa PM, Waters CW, Baron MH, Fraser SE, Dickinson ME. Dynamic in vivo imaging of postimplantation mammalian embryos using whole embryo culture. Genesis. 2002;34:228–235. doi: 10.1002/gene.10162. [DOI] [PubMed] [Google Scholar]
- 8.Lucitti JL, Jones EA, Huang C, Chen J, Fraser SE, Dickinson ME. Vascular remodeling of the mouse yolk sac requires hemodynamic force. Development. 2007;134:3317–3326. doi: 10.1242/dev.02883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Boppart SA, Brezinski ME, Bouma B, Tearney G, Fujimoto JG. Investigation of developing embryonic morphology using optical coherence tomography. Developmental Biology. 1996;177:54–63. doi: 10.1006/dbio.1996.0144. [DOI] [PubMed] [Google Scholar]
- 10.Boppart SA, Tearney GJ, Bouma BE, Southern JF, Brezinski ME, Fujimoto JG. Noninvasive assessment of the developing Xenopus cardiovascular system using optical coherence tomography. Proc Natl Acad Sci USA. 1997;94:4256–4261. doi: 10.1073/pnas.94.9.4256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Yelbuz TM, Choma MA, Thrane L, Kirby ML, Izatt JA. Optical coherence tomography — A new high-resolution imaging technology to study cardiac development in chick embryos. Circulation. 2002;106:2771–2774. doi: 10.1161/01.cir.0000042672.51054.7b. [DOI] [PubMed] [Google Scholar]
- 12.Yang VXD, Gordon M, Seng-Yue E, Lo S, Qi B, Pekar J, Mok A, Wilson B, Vitkin I. High speed, wide velocity dynamic range Doppler optical coherence tomography (Part II): Imaging in vivo cardiac dynamics of Xenopus laevis. Opt Express. 2003;11:1650–1658. doi: 10.1364/oe.11.001650. [DOI] [PubMed] [Google Scholar]
- 13.Jenkins MW, Rothenberg F, Roy D, Nikolski VP, Hu Z, Watanabe M, Wilson DL, Efimov IR, Rollins AM. 4D embryonic cardiography using gated optical coherence tomography. Opt Express. 2006;14:736–748. doi: 10.1364/opex.14.000736. [DOI] [PubMed] [Google Scholar]
- 14.Luo W, Marks DL, Ralston TS, Boppart SA. Three-dimensional optical coherence tomography of the embryonic murine cardiovascular system. J Biomed Opt. 2006;11:021014. doi: 10.1117/1.2193465. [DOI] [PubMed] [Google Scholar]
- 15.Jenkins MW, Adler DC, Gargesha M, Huber R, Rothenberg F, Belding J, Watanabe M, Wilson DL, Fujimoto JG, Rollins AM. Ultra-high-speed optical coherence tomography imaging and visualization of the embryonic avian heart using a buffered Fourier domain mode locked laser. Opt Express. 2007;15:6251–6267. doi: 10.1364/oe.15.006251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mariampillai A, Standish BA, Munce NR, Randall C, Liu G, Jiang JY, Cable AE, Vitkin IA, Yang VXD. Doppler optical cardiogram gated 2D color flow imaging at 1000 fps and 4D in vivo visualization of embryonic heart at 45 fps on a swept source OCT system. Opt Express. 2007;15:1627–1638. doi: 10.1364/oe.15.001627. [DOI] [PubMed] [Google Scholar]
- 17.Davis AM, Rothenberg FG, Shepherd N, Izatt JA. In vivo spectral domain optical coherence tomography volumetric imaging and spectral Doppler velocimetry of early stage embryonic chicken heart development. J Opt Soc Am A. 2008;25:3134–3143. doi: 10.1364/josaa.25.003134. [DOI] [PubMed] [Google Scholar]
- 18.Kagemann L, Ishikawa H, Zou J, Charukamnoetkanok P, Wollstein G, Townsend KA, Gabriele ML, Bahary N, Wei X, Fujimoto JG, Schuman JS. Repeated, noninvasive, high resolution spectral domain optical coherence tomography imaging of zebrafish embryos. Mol Vis. 2008;14:2157–2170. [PMC free article] [PubMed] [Google Scholar]
- 19.Larina IV, Sudheendran N, Ghosn M, Jiang J, Cable A, Larin KV, Dickinson ME. Live imaging of blood flow in mammalian embryos using Doppler swept source optical coherence tomography. J Biomed Opt. 2008;13:0605063. doi: 10.1117/1.3046716. [DOI] [PubMed] [Google Scholar]
- 20.Bradu A, Ma L, Bloor JW, Podoleanu A. Dual optical coherence tomography/fluorescence microscopy for monitoring of Drosophila melanogaster larval heart. Journal of Biophotonics. 2009 doi: 10.1002/jbio.200910021. [DOI] [PubMed] [Google Scholar]
- 21.Davis A, Izatt J, Rothenberg F. Quantitative measurement of blood flow dynamics in embryonic vasculature using spectral Doppler velocimetry. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology. 2009;292:311–319. doi: 10.1002/ar.20808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Larina IV, Ivers SN, Syed SH, Dickinson ME, Larin KV. Hemodynamic measurements from individual blood cells in early mammalian embryos with Doppler swept source OCT. Opt Lett. 2009;34:986–988. doi: 10.1364/ol.34.000986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Oh WY, Yun SH, Tearney GJ, Bouma BE. 115 kHz tuning repetition rate ultrahigh-speed wavelength-swept semiconductor laser. Opt Lett. 2005;30:3159–3161. doi: 10.1364/ol.30.003159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Huber R, Adler DC, Srinivasan VJ, Fujimoto JG. Fourier domain mode locking at 1050 nm for ultra-high-speed optical coherence tomography of the human retina at 236,000 axial scans per second. Opt Lett. 2007;32:2049–2051. doi: 10.1364/ol.32.002049. [DOI] [PubMed] [Google Scholar]
- 25.Huber R, Wojtkowski M, Fujimoto JG. Fourier Domain Mode Locking (FDML): A new laser operating regime and applications for optical coherence tomography. Opt Express. 2006;14:3225–3237. doi: 10.1364/oe.14.003225. [DOI] [PubMed] [Google Scholar]
- 26.Potsaid B, Gorczynska I, Srinivasan VJ, Chen Y, Jiang J, Cable A, Fujimoto JG. Ultrahigh speed Spectral/Fourier domain OCT ophthalmic imaging at 70,000 to 312,500 axial scans per second. Opt Express. 2008;16:15149–15169. doi: 10.1364/oe.16.015149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Adler DC, Huber R, Fujimoto JG. Phase-sensitive optical coherence tomography at up to 370,000 lines per second using buffered Fourier domain mode-locked lasers. Opt Lett. 2007;32:626–628. doi: 10.1364/ol.32.000626. [DOI] [PubMed] [Google Scholar]
- 28.Tuchin VV. Optical Clearing of Tissues and Blood. PM 154. Bellingham, WA: SPIE Press; 2005. [Google Scholar]
- 29.Larina IV, Carbajal EF, Tuchin VV, Dickinson ME, Larin KV. Enhanced OCT imaging of embryonic tissue with optical clearing. Laser Phys Lett. 2008;5:476–480. [Google Scholar]