

Abstract
While anecdotally there appear to be differences in the way native speakers use and comprehend their native language, most empirical investigations of language processing study university students and none have studied differences in language proficiency which may be independent of resource limitations such as working memory span. We examined differences in language proficiency in adult monolingual native speakers of English using an event-related potential (ERP) paradigm. ERPs were recorded to insertion phrase structure violations in naturally spoken English sentences. Participants recruited from a wide spectrum of society were given standardized measures of English language proficiency, and two complementary ERP analyses were performed. In between-groups analyses, participants were divided, based on standardized proficiency scores, into Lower Proficiency (LP) and Higher Proficiency (HP) groups. Compared to LP participants, HP participants showed an early anterior negativity that was more focal, both spatially and temporally, and a larger and more widely distributed positivity (P600) to violations. In correlational analyses, we utilized a wide spectrum of proficiency scores to examine the degree to which individual proficiency scores correlated with individual neural responses to syntactic violations in regions and time windows identified in the between-group analyses. This approach also employed partial correlation analyses to control for possible confounding variables. These analyses provided evidence for the effects of proficiency that converged with the between-groups analyses. These results suggest that adult monolingual native speakers of English who vary in language proficiency differ in the recruitment of syntactic processes that are hypothesized to be at least in part automatic as well as of those thought to be more controlled. These results also suggest that in order to fully characterize neural organization for language in native speakers it is necessary to include participants of varying proficiency.
Anecdotally, it appears that there exist differences in the way native speakers use and comprehend their native language. Beginning with Chomsky's (1965) claim that linguistic theory should be concerned with an ideal speaker-listener with perfect linguistic knowledge, differences in linguistic proficiency among native speakers have often been assumed to be the result of resource limitations or performance errors considered to be independent of and irrelevant to grammatical knowledge. Numerous studies of bilinguals, young children, children with specific language impairment, and deaf adults have found that event-related potentials (ERPs) are sensitive to differences in language proficiency (Mills, Coffey-Corina, & Neville, 1993; Neville, Coffey, Holcomb, & Tallal, 1993; Neville, Mills, & Lawson, 1992; Weber-Fox & Neville, 1996).
Additionally, several ERP studies have examined differences in language processing in native speakers. However, most of these studies have not separately assessed confounds between language processing and other cognitive resource limitations because they specifically studied complex syntactic structures to examine individuals with differences in working memory capacity (Friederici, Steinhauer, Mecklinger, & Meyer, 1998; King & Kutas, 1995; Mecklinger, Schriefers, Steinhauer, & Friederici, 1995; Vos & Friederici, 2003; Vos, Gunter, Kolk, & Mulder, 2001; Vos, Gunter, Schriefers, & Friederici, 2001). Also, most of these latter studies looked at university students processing sentences in the visual modality. Because the majority of language use in everyday life occurs in the auditory modality and between speakers with a wide variety of educational backgrounds, there remains the question of the extent to which proficiency differences in syntactic processing may be apparent under conditions which more closely approximate real-world language use. In the current study we tested the hypothesis that differences in grammatical proficiency in native speakers are indexed by differences in ERP components related to the processing of phrase structure violations in auditorily presented sentences. To this end, we examined differences in grammatical processing proficiency in participants recruited from a wide spectrum of society using a natural speech paradigm which does not place high demands on working memory resources. In order to more fully characterize these differences, we conducted two complementary analyses: first, we characterized differences in the ERP response to syntactic violations between two groups which differed in standardized proficiency scores; second, we conducted correlational analyses with a larger group of participants in order to assess the degree to which the relationship between proficiency and the neural response to syntactic violations held across a wide spectrum of proficiency scores and while controlling for other possible mediating factors.
ERP Studies of Language Processing
ERPs provide an on-line, multidimensional index of cognitive processes with a temporal resolution of milliseconds and thus have emerged as one of the more widely used methodologies to examine on-line language processing. Along with other methods, ERP studies have demonstrated that separate linguistic subsystems are mediated by non-identical neural mechanisms. Numerous studies in both the visual and auditory modalities have found that semantically unexpected words elicit a negative-going potential peaking around 400 ms (N400) compared to contextually appropriate words (e.g., Friederici, Pfeifer, & Hahne, 1993; Holcomb & Neville, 1991; Kutas & Hillyard, 1980), leading to the hypothesis that the N400 component indexes semantic processes of lexical integration.
While the N400 has consistently been related to aspects of semantic processing, at least two components have been identified which index syntactic processing. One of these is a negative-going wave, typically larger over left anterior electrode sites between 100-500 ms, known as the left anterior negativity (LAN). The LAN has been elicited by a variety of syntactic violation types, such as phrase structure violations (Friederici, et al., 1993; Gunter, Friederici, & Hahne, 1999; Hahne & Friederici, 1999; Neville, Nicol, Barss, Forster, & Garrett, 1991; Yamada & Neville, 2007) and morphosyntactic violations (Coulson, King, & Kutas, 1998a; Friederici, et al., 1993; Münte, Heinze, & Mangun, 1993). The LAN typically occurs in one or both of two time windows (100-300 ms and 300-500 ms), which has lead some researchers to propose the existence of two distinct, separate components, with the first, termed the early left anterior negativity (ELAN), indexing processes different from those indexed by the second, LAN (Friederici, 1995; Friederici & Mecklinger, 1996; Hahne & Jescheniak, 2001). Two recently proposed theories of online sentence processing account for these components in different ways. Friederici (2002) proposes that the ELAN reflects early and automatic processing of word category violations, a process hypothesized to be autonomous and independent of contextual or semantic influences. In contrast, Hagoort and colleagues (Hagoort, 2003; Hagoort, 2005; van den Brink & Hagoort, 2004) propose that semantic and syntactic information are processed in parallel as soon as they are available and that the timing differences reported between LAN and ELAN effects are a result of differences in the online availability of morphosyntactic and word category information, not as the result of a fundamental functional distinction between them.
The second component which has been observed in ERP studies of syntactic processing is a large positive-going wave usually largest over bilateral posterior regions and peaking between 500-1000 ms., known as the P600 (Osterhout & Holcomb, 1993). The P600 is consistently elicited by syntactic violations (Hagoort, Brown, & Groothusen, 1993; Hagoort & Brown, 2000; Hahne & Friederici, 1999; Osterhout & Holcomb, 1992; Osterhout & Mobley, 1995) as well as by violations of preferred syntactic structure (Osterhout & Holcomb, 1992; Osterhout, Holcomb, & Swinney, 1995) or in well-formed sentences of higher syntactic complexity (Kaan, Harris, Gibson, & Holcomb, 2000; Kaan & Swaab, 2003a, 2003b). While the distribution of the P600 is usually posterior, several studies have reported a late positivity with a more frontal distribution to grammatically correct but non-preferred structures (Friederici, Hahne, & Saddy, 2002; Kaan & Swaab, 2003a, 2003b; Osterhout & Holcomb, 1992). This has led to the proposal that the frontally distributed P600 reflects processing difficulties related to revision in the face of non-preferred structures, while the posteriorally distributed P600 reflects processes related to the failure of a parse and related processes of repair and meaning rescue (Friederici, et al., 2002; Hagoort & Brown, 2000) or to syntactic integration difficulty (Kaan, et al., 2000).
While overall there is a good deal of consistency in the elicitation of this biphasic response across studies using different violation types in different languages and modalities, there still exists a great deal of variability, in particular with regard to the distribution of the anterior negativity effect. Specifically, several studies have reported an anterior negativity effect in monolingual native speakers which is more extended temporally and/or more bilateral in distribution (e.g., Friederici & Frisch, 2000; Friederici & Mecklinger, 1996; Friederici, et al., 1993; Hagoort, Wassenaar, & Brown, 2003; Hahne, 2001; Hahne & Friederici, 2002; Hahne & Jescheniak, 2001; Isel, Hahne, Maess, & Friederici, 2007; Münte, Matzke, & Johannes, 1997; Rossi, Gugler, Friederici, & Hahne, 2006). While possible sources of this variability are potentially informative for theories of online sentence processing, little is known about this issue. One possible source of this variability is within-group differences in participant characteristics, such as language proficiency. While it is difficult to assess the possibility that such differences may have contributed to the variability observed in previous studies, here we specifically address the possibility that differences in participant characteristics may be reflected in differences in the distribution and timing of ERPs elicited by syntactic violations.
Individual Differences and Effects of Experience
Previous studies of individual differences in adult native speakers have primarily focused on differences in working memory (WM) using paradigms involving garden-path sentences or manipulations of syntactic complexity. Early behavioral studies found that individual differences in WM capacity are predictive of comprehension and speed of processing, as subjects with lower WM spans were slower and less accurate in comprehending complex syntactic constructions compared to subjects with higher WM spans (Just & Carpenter, 1992; King & Just, 1991). Subsequent ERP studies have found that these differences are reflected electrophysiologically as well. The most common finding is that the disambiguating element in object-first relative clauses compared to subject-first relative clauses elicits a P600, but only in subjects who performed faster on sentence comprehension tasks (Mecklinger, et al., 1995) or who had high WM spans (Friederici, et al., 1998; Vos & Friederici, 2003). While these studies provide evidence that ERPs are sensitive to individual differences in WM and syntactic processing, the focus on differences in WM raises the question of the degree to which individual differences in proficiency may occur independently of other resource limitations such as WM span.
Several studies of both monolingual native speakers and of bilinguals suggest that language experience affects both linguistic proficiency and related neural systems indexed in ERP paradigms. ERP studies of monolingual children suggest that brain organization is predicted by language proficiency: children with larger vocabularies (Mills, et al., 1993) and children who score higher on tests of language comprehension (Adamson-Harris, Mills, & Neville, 2000) show more mature patterns of brain organization for language, including greater focalization, as compared with children with smaller vocabularies or those who score lower on comprehension tests. ERP studies of deaf adults suggest that the effects of early language experience can endure into adulthood, as individuals exposed to American Sign Language (ASL) from an early age recruit right hemisphere areas in addition to left hemisphere language areas when processing ASL. However, those not exposed to ASL at an early age do not show this bilateral response to ASL and score lower on tests of ASL grammar (Neville, Coffey, Lawson, Fischer, & et al., 1997; Newman, Bavelier, Corina, Jezzard, & Neville, 2002; Newport, 1990). More evidence from ERP studies of bilinguals suggest that linguistic subsystems are differentially sensitive to the effects of language experience, with the syntactic subsystem displaying less focal neural organization with delays in second language exposure as short as 4-6 years, while the semantic subsystem appears to be affected by delays in second language exposure only after 11-13 years of age (Weber-Fox & Neville, 1996). Similar results have been observed for native and late learners of ASL (Capek, et al., 2002; Capek, et al., 2009).
This evidence of the effects of language experience raises the question of the extent of these effects. Do significant differences in proficiency exist within normal monolingual adults, and would such differences be indexed by ERP components related to syntactic processing? Or do effects of early experience fade with time such that neural systems underlying language processing in adult monolinguals are relatively homogeneous?
Data from one ERP experiment suggests that significant differences in proficiency do exist in monolingual adults and are linked to altered neural organization as indexed by ERPs. Weber-Fox, Davis, and Cuadrado (2003) found that individuals who scored higher on a spoken grammar test had an earlier N280 to closed-class words over left anterior regions, suggesting greater efficiency related to syntactic processing in higher proficiency individuals. If ERP components associated with syntactic processing are sensitive to differences in linguistic proficiency, then the evidence discussed above raises the hypothesis that components elicited by phrase structure violations in low proficiency participants should be less focal than those elicited in higher proficiency individuals.
The Present Study
The present study further explored the relationship between proficiency and neural organization for language in monolingual native speakers as measured by ERPs during on-line syntactic processing. We attempted to maximize proficiency differences by recruiting participants from a wide spectrum of society. We attempted to minimize the effects of other potential resource limitations such as WM by using a paradigm which examined the brain response to phrase structure violations in simple, single-clause sentences, which were presented auditorily in an attempt to increase ecological validity while minimizing confounds related to literacy. In an effort to more fully characterize differences in neural organization related to proficiency, we conducted two complementary analyses. In the first, we examined two groups of participants with scores at or near the upper and lower quartiles on standardized tests of English proficiency. In the second, we examined the relationship between proficiency and the neural response to syntactic violations in a group of 72 participants with a wide range of proficiency scores, employing a correlational approach which allowed us to control for other possible mediating factors including education level, working memory span, and socioeconomic status of origin.
Method
Participants
Right-handed, normal hearing, native monolingual speakers of English, recruited from both the university and non-university populations, participated in the study. Participants were paid for their time. A total of 116 participants were run in the behavioral testing paradigm described below, and of these 80 were also run in the event-related potential paradigm described below. From this group of 80, eight participants were removed from the final analysis either after being identified as outliers on the behavioral measures or due to excessive ERP artifact. This left a group of 72 participants with good behavioral and ERP data, and from this group 34 were selected for the between-group analyses based on behavioral performance to form Lower Proficiency (LP; N = 17) and Higher Proficiency (HP; N = 17) groups. An average standardized score for the three subtests of the Test of Adolescent and Adult Language-3 (TOAL-3; (Hammil, Brown, Larsen, & Wiederholt, 1994) was calculated for all participants, and the LP and HP groups were formed by selecting the participants with the lowest and highest average standardized scores. In order to reduce group differences in WM capacity, an initial selection of the participants with the lowest and highest average standardized scores was modified by removing the two participants from each group with the lowest and highest Carpenter Reading Span scores, respectively.
Behavioral Language Inventories
Three subtests of the TOAL-3 were administered to assess proficiency. The TOAL-3 Listening/Vocabulary subtest is a test which requires participants to match a vocabulary word with two pictures relating to that word (out of four pictures shown). The TOAL-3 Listening/Grammar subtest requires participants to determine, out of three sentences presented auditorily, which two sentences have similar meaning. The TOAL-3 Speaking/Grammar subtest requires participants to repeat exactly sentences said by the examiner; the sentences gradually increase in syntactic difficulty. Participants were also given the Saffran and Schwartz Grammaticality Judgment Test (Linebarger, Schwartz, & Saffran, 1983), a 78-item assessment in which participants are asked to judge the grammaticality of sentences containing a variety of syntactic violations, adapted for purposes of this study into the auditory modality. In order to assess working memory capacity, participants were also given the Carpenter Reading Span Test (Daneman & Carpenter, 1980), a widely-used assessment in which participants must recall the final word of two or more sentences after reading them consecutively. Participants also completed a questionnaire which gathered data about educational attainment, as well as about language usage, television, and video game habits both as children and as adults. This questionnaire also gathered data used to calculate the socioeconomic status of the family in which participants were raised until 18 years of age or independence. This was measured by the Hollingshead Four Factor Index of Social Status (Hollingshead, 1975), which takes into consideration parental education and occupation.
Stimuli
In the ERP paradigm, participants heard both English sentences and Jabberwocky sentences, in which open-class words were replaced with pronounceable nonwords to greatly reduce the semantic context; only the results for the English stimuli are presented here. The English stimuli were sentences which were canonical (50%) or which contained an insertion phrase structure violation (50%) in which an additional closed-class word was inserted in a sentence-final prepositional phrase. In all cases, the phrase structure violation clearly occurred at the onset of either a demonstrative (50%) or possessive (50%) pronoun directly following the inserted pronoun. The ERPs to the onset of the target word (underlined below) in the canonical and violation (*) sentences were compared:
| English: | Timmy can ride the horse at his farm. |
| *Timmy can ride the horse at my his farm. |
A number of measures were undertaken in order to provide prosodic variability as well as to ensure that subjects listened fully to the sentences and did not focus only on the location of the critical violation. In 5% of the experimental sentences an additional prepositional phrase was added to the beginning of the sentences, and in 20% of the experimental sentences an adjective was placed directly after the target word so that the target word was not invariably in the penultimate position in the sentence. In addition, filler sentences and probe questions were constructed. Filler sentences contained a permutation phrase structure violation in which a main verb and the determiner of the object noun phrase were reversed. Probe questions took the form “Did you hear the word (blank)?” Most participants heard 62 sentences of each condition; a subset (N = 10) heard 40 sentences per condition. Filler sentences (10% of total) were pseudo-randomly interspersed between the experimental sentences, as were probe questions (5% of total), such that filler sentences and probe questions occurred equally across quarter stimulus blocks and were always separated by at least two experimental sentences.
All sentences were recorded using SoundEdit 16 Version 2 with 16-bit resolution and a 16 Khz sampling rate then transferred to a PC for presentation. The sentences were spoken by a female with natural tempo and prosody and critical word onsets were identified and coded by three trained coders using both auditory cues and visual inspection of sound spectrographs for increased accuracy. Any sentences in which codes differed by more than 20 milliseconds between coders were re-coded by all three coders together until a consensus was reached by all three to ensure reliability.
Procedure
Most participants were tested in one three-hour session, with the standardized tests of language administered right before ERP testing. A subset of participants (N = 24) were given the behavioral measures and ERP testing in separate sessions. In each ERP session a 32-channel electrode cap was applied while the participant completed an information sheet which included questions about their education, socioeconomic status, handedness, neurological history, and language habits. Participants were then seated in a comfortable chair in an electrically shielded, sound-attenuating booth. Sentences were presented via a speaker placed centrally on a monitor 70 in. from the participant. Participants were given auditory instructions including examples of both sentence types and emphasizing the need to judge the sentences based on grammatical, and not semantic, correctness. On each trial, participants pushed one of two response buttons to play a sentence. While the sentences were playing, participants were asked to refrain from blinking or moving their eyes as a box with a central fixation cue (‘*’) was displayed. After each sentence, participants were cued to make a judgment with a display of “Yes or No?” on the screen. The judgment was made with a button press with either the left or right hand, counterbalanced across participants. Participants proceeded at their own pace and were given two regularly scheduled breaks and additional breaks as requested.
EEG Equipment and Analysis
The EEG was recorded using tin electrodes mounted in an appropriately sized elastic cap (Electro-Cap International) over 29 scalp sites based on Standard International 10-20 electrode locations: F7/F8, F3/F4, FT7/FT8, FC5/FC6, T3/T4, C5/C6, CT5/CT6, C3/C4, T5/T6, P3/P4, TO1/TO2, O1/O2, FP1/FP2, Fz, Cz, and Pz. Scalp electrode impedances were kept below 3 Kohms. Data from all scalp electrodes were referenced on-line to the EEG from an electrode placed over the right mastoid and later referenced off-line to the mathematical average of the left and right mastoids. Horizontal eye movements were monitored using electrodes placed at the outer canthus of each eye and referenced to each other, while vertical eye movements were monitored using an electrode placed beneath the right eye and referenced to the right mastoid. The raw EEG signal was collected at a sampling rate of 250 Hz and was amplified using Grass Amplifiers with high- and low-pass filter settings of 0.01 Hz and 100 Hz, respectively.
Only trials on which subjects responded correctly were included in the ERP analyses. The EEG data for each participant were examined for eye movements, muscle artifact, and amplifier saturation and drift, and any trials contaminated by these artifacts were excluded from final data analyses. ERPs were computed for 1200 ms after the onset of the target word relative to a 100 ms prestimulus baseline. ERP waveforms were measured within specific time windows determined by visual inspection of individual and group averages; specific time windows are described in the Results section. Based on a priori hypotheses from previous studies and on visual inspection of the effects, the anterior negativity effect was characterized by analyzing the 12 anterior electrode sites. Additional analyses conducted on different time windows and electrode sites are noted in the Results section.
For the between-group analyses, mean voltage amplitude was measured within each time window and analyzed using ANOVAs with repeated measures, including two levels of condition (C: canonical, violation), two levels of hemisphere (H: left, right), three levels of anterior-posterior (A: frontal, fronto-temporal, temporal (anterior sites); central, parietal, and occipital (posterior sites)), and two levels of lateral-medial (L: lateral, medial), as well as a between-participants factor, proficiency, with two levels (P: Lower Proficiency, Higher Proficiency). Following omnibus ANOVAs, additional analyses were performed in step-down fashion in order to isolate any significant interactions, collapsing across factors with which an interaction was not found. When significant between-group interactions were found, separate ANOVAs were performed for each group to better characterize group differences. For all distributional comparisons, analyses were performed on both the raw data and on data normalized following the procedure recommended by McCarthy and Wood (1985); as no differences between the two analyses were found, only analyses performed on the raw data are reported. Greenhouse-Geisser corrections were applied to all ANOVAs with greater than one degree of freedom.
For correlational analyses, for each of the 72 participants the average difference amplitude (violation – canonical) was calculated for each electrode site. Based on the results from the between-group analyses, three time windows were analyzed to capture the anterior negativity: 100-300 ms, 300-700 ms, and 700-1200 ms. Within these time windows, the average difference amplitude was calculated across left anterior lateral sites (F7, FT7, T3), left anterior medial sites (F3, FC5, C5), right anterior medial sites (F4, FC6, C6), and right lateral sites (F8, FT8, T4). A laterality index was also calculated for anterior lateral sites (left anterior lateral – right anterior lateral), anterior medial sites (left anterior medial – right anterior medial), and anterior sites (left anterior – right anterior). The 300-1000 ms time window was analyzed to capture the P600 effect; within this time window the average difference amplitude was calculated across all 12 posterior sites. Zero-order correlations were then calculated between individual average difference amplitudes and individual proficiency scores, quantified as a composite individual average standardized score for the three subtests of the TOAL-3. In order to control for the possible influence of other variables which significantly correlated with proficiency, partial correlations controlling for these variables were also conducted. Any correlations driven by extreme scores, as defined by lying more than two standard deviations from the mean, are not reported.
Results: Between-group Analyses
Behavioral Results
Lower Proficiency (LP) and Higher Proficiency (HP) groups were determined by standardized scores on the two TOAL-3 subtests used as described in the methods section. The mean average standardized scores for the resulting LP (M = 8.09, SD = 1.65) and HP (M = 13.06, SD = 1.20) groups were statistically independent (t(32) = -10.04, p < .0001). The mean scores for each behavioral measure are displayed in Table 1. Means for the groups were statistically independent for each language measure. While the groups did not differ significantly on the measure of WM capacity, there was a trend toward a group difference. While all participants were within normal limits for native speakers, the groups were distinct in terms of TOAL-3 standardized scores: the mean scores for the LP group were at or below the 34th percentile on each subtest (below the 25th percentile for the grammar subtests), while the mean scores for the HP group were at or above the 75th percentile for each subtest. In the ERP grammaticality judgment task, there was a trend for a higher percentage of correct responses by the HP group (M = 98.64, SD = 1.45) compared to the LP group (M = 94.96, SD = 9.94) which did not reach significance (t(32) = 1.512, p = .14). Scores on measures used to ensure that participants were listening to the entire sentence indicated that this was indeed the case. The HP group answered 100% of the probe questions correctly and the LP group, while performing significantly worse, still answered 94.18% of the questions correctly (t(32) = 2.411, p < .05).
Table 1. Mean scores by proficiency.
| Group | TOAL-3 L-V** |
TOAL-3 L-G** |
TOAL-3 S-G** |
Saffran and Schwartz* |
Carpenter Span+ |
|---|---|---|---|---|---|
| LP | |||||
| (n = 17, 7 F) | |||||
| M | 23.06 | 19.00 | 17.06 | 74.29 | 2.79 |
| (SD) | (5.69) | (7.98) | (3.36) | (3.08) | (.53) |
| Range | 14-33 | 7-32 | 9-22 | 67-78 | 2-4 |
| Percentile | 37 | 25 | 16 | N/A | N/A |
| HP | |||||
| (n = 17, 10 F) | |||||
| M | 31.18 | 30.71 | 22.59 | 76.12 | 3.15 |
| (SD) | (2.55) | (4.79) | (1.62) | (.992) | (.66) |
| Range | 27-35 | 17-35 | 20-25 | 75-78 | 2-5 |
| Percentile | 84 | 75 | 91 | N/A | N/A |
p < .05
p < .0001
p = .095
The mean average standardized scores for the groups correlated significantly with several personal history factors. Highest education level achieved correlated significantly with proficiency scores (r = .632, p < .0001), and the HP group had significantly more education than the LP group (t(32) = 3.875, p < .01). In addition, the socioeconomic status of the family in which participants were raised until 18 years of age or independence was calculated. This was measured by the Hollingshead Four Factor Index of Social Status (Hollingshead, 1975), which takes into consideration parental education and occupation. Proficiency scores also correlated significantly with family SES (r = .320, p < .05); though the HP group had a higher family SES, this difference did not reach significance.
ERP Results
The ERP data to the critical word in English sentences for both groups are shown in Figure 1. Visual inspection of the waveforms revealed clear patterns and clear differences between proficiency groups. Both groups displayed a biphasic response to phrase structure violations: an anterior negativity with onset around 100 ms and a posterior positivity peaking around 600 ms. In the HP group, the anterior negativity effect was short in duration and larger over left hemisphere sites. In contrast, the anterior negativity in the LP group was more extended temporally and spatially, with the effect extending beyond 400 ms and more prominent over right hemisphere sites than in the HP group. In addition, the anterior negativity effect extended to 1200 ms over lateral sites, and a late negativity began at 700 ms over medial sites for the LP group. Group differences were also evident in the P600 effect, which was larger and more broadly distributed in the HP group compared to the LP group.
Figure 1.

ERPs to English phrase structure violations for HP and LP groups showing representative electrode rows illustrating the anterior negativity (frontal and fronto-temporal) and P600 (parietal) effects.
Anterior Negativity
Analyses across anterior electrode sites in the 100-300 ms time window revealed a group interaction with the factors hemisphere and lateral-medial (C × H × L × P: F(1, 32) = 4.22, p < .05), reflecting significant differences in the distribution of the anterior negativity effect across proficiency groups. These distributional differences are illustrated by voltage maps in Fig. 2. The effect in the HP group was more lateralized to left-hemisphere sites, in particular across medial sites, while the effect in the LP group was more widespread and bilateral in distribution. Further analyses of this interaction confirmed this interpretation. In the HP group the effect was significantly larger over left-hemisphere sites overall (C × H: F(1, 16) = 8.71, p < .01). In the LP group, the effect was bilateral overall (C × H: F(1, 16) = 1.81, NS). While there was a greater degree of left lateralization over lateral sites (C × H × L: F(1, 16) = 4.65, p < .05) in the LP group, a direct comparison of lateral sites by hemisphere did not reach significance (t(16) = 1.762, p = .097). These further analyses also confirmed distribution differences across lateral and medial sites, as there was a trend for the effect to be larger over lateral sites in the HP group (C × L: F(1, 16) = 4.25, p = .056), while it was evenly distributed across lateral and medial sites in the LP group (C × L: F(1, 16) = 0.53, NS).
Figure 2.
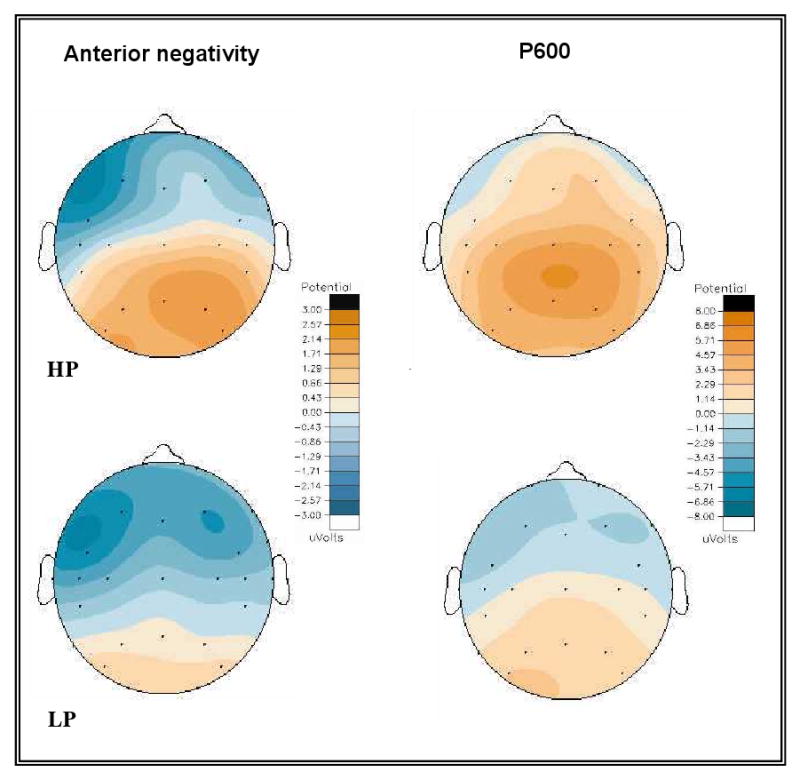
Voltage maps for HP and LP groups illustrating the distribution of the anterior negativity effect for English in over the 100-300 ms time window and the posterior positivity (P600) effect over the 300-1000 ms time window.
Visual inspection also suggested group differences in the distribution of the negativity to the central row of electrodes (CT5-6, C3-4), and an interaction in the 100-300 ms time window across all electrode sites which neared significance confirmed that this effect extended to the central row, maximally over medial sites, in the LP group but not the HP group (C × A × L × P: F(5, 160) = 2.72, p = .055).
Late Negativity
As visual inspection suggested that the anterior negativity was longer in duration in the LP group, additional analyses across anterior sites were conducted in later time windows. A group interaction between 300-700 ms confirmed that the negativity extended in this time window in the LP group only, maximally over anteriormost lateral sites; in contrast, violations elicited a positivity maximal over medial sites in this time window in the HP group (C × A × L × P: F(2, 64) = 6.27, p < .01). As visual inspection revealed an anterior negativity in the 700-1200 time window in the LP group, additional analyses in this time window were conducted. These analyses confirmed that while the negativity extended across this epoch in the LP group, this was not the case in the HP group (C × P: F(1, 32) = 10.41, p < .005). This negativity was bilaterally distributed in the LP group in both the 300-700 ms time window (C × H: F(1, 16) = .281, NS) and the 700-1200 ms time window (C × H: F(1, 16) = .434, NS).
Posterior Positivity (P600)
Because visual inspection suggested group differences in amplitude and distribution in the posterior positivity effect across all electrode sites, initial analyses were conducted across all sites. An analysis in the 300-1200 ms time window revealed that this effect was larger overall in the HP group than in the LP group (C × H: F(1, 32) = 11.65, p < .005) and that it extended to anterior medial sites in the HP group but not in the LP group (C × A × L × P: F(5, 160) = 6.20, p < .005).
As individual group analyses confirmed that the posterior positivity was confined to the three posterior rows in the LP group (C × A: F(5, 80) = 27.66, p < .0001), additional group comparisons limited to these rows were conducted. These analyses revealed that the effect was larger in the HP group compared to the LP group over sites where the effect was present in both groups (C × P: F(1, 32) = 8.24, p < .01).
Results: Correlational Analyses
Behavioral Results
In order to limit the number of variables considered in the correlational analyses with average ERP difference amplitudes, the relationship between proficiency and several potentially confounding variables was explored. Proficiency correlated significantly with socioeconomic status of origin (henceforth SES) (r = .460, p < .0001), working memory span (r = .561, p < .0001), and education level (r = .368, p < .005). Proficiency did not correlate significantly with participant age, and analysis by gender revealed no significant differences in proficiency between males (N = 35) and females (N = 37); therefore age and gender were not included in the ERP correlational analyses. As results from the between-group analyses provided specific hypotheses concerning the direction of the effects, all significance levels reported are one-tailed unless otherwise specified.
Correlational analyses also revealed possible environmental factors contributing to proficiency differences in adult monolingual native speakers. Proficiency correlated with the amount participants reported reading as children (r = .234, p < .05) and there was a nearly significant correlation with the frequency participants reported being read to when children (r = .190, p = .055), while there was a negative correlation with the amount of time participants reported playing video games as children (r = -.334, p < .005). The amount of time participants reported reading as children also correlated positively with the amount of time participants reported reading as adults (r = .450, p < .0001), as did the frequency participants reported being read to as children (r = .298, p < .01). All four self-report measures of childhood environment correlated significantly with SES: amount participants reported reading (r = .378, p < .005), frequency participants reported being read to (r = .538, p < .0001), watching television (r = -.265, p < .05), and playing video games as children (r = -.319, p < .005).
While proficiency did not correlate with the amount of time participants reported reading as adults, there was a significant negative correlation between proficiency and the amount of time participants reported watching television as adults (r = -.429, p < .0001). In addition, a self-rating measure in which participants were asked to rate their language skills compared to other adult native speakers on a five-point scale correlated significantly with both SES (r = .317, p < .005) and proficiency (r = .437, p < .0001).
ERP Results
Overall, results from the correlational analyses supported the findings from the between-group analyses and provided converging evidence from a complementary analytical approach for effects of proficiency on neural organization for syntactic processing. These analyses revealed the strongest effects of proficiency in later time windows over both anterior and posterior electrode sites. Importantly, these effects of proficiency were found to be independent of other confounding variables, as partial correlations were used to control for the effects of SES, working memory span, and education level. In addition to proficiency effects, these analyses also revealed an effect of SES on the amplitude of the early left anterior negativity.
Anterior Negativity
100-300 ms TW
While no zero-order correlations reached significance in this time window, there was a near-significant partial correlation with proficiency and average difference amplitude over right anterior medial sites (r = .194, p = .052). Consistent with the results from the between-groups analyses, this analysis revealed that lower proficiency participants showed an increased negative response over these sites.
300-700 ms TW
Significant zero-order correlations were found between proficiency and average difference amplitude over both left anterior (r = .365, p < .005) and right anterior (r = .334, p < .005) sites (Figure 3). Partial correlations across these sites were also significant (left anterior: r = .406, p < .0001; right anterior: r = .276, p < .05), as were partial correlations across left anterior lateral (r = .337, p < .005), left anterior medial (r = .430, p < .0001), and right anterior medial (r = .325, p < .005) sites. The relationship was weakest across right anterior lateral sites, where the partial correlation tended toward significance (r = .184, p = .065). These correlations revealed that lower proficiency participants showed a more negative response while higher proficiency participants showed a more positive response across this time window, consistent with the results from the between-groups analyses.
Figure 3.

Correlation between average difference amplitude (violation – canonical, in μV) over left anterior and right anterior sites and proficiency in the 300-700 ms time window.
700-1200 ms TW
Significant zero-order correlations were found between proficiency and average difference amplitude over both left anterior (r = .203, p < .05) and right anterior (r = .230, p < .05) sites, revealing that lower proficiency participants showed a more negative response across this time window. Partial correlations across these sites only approached significance (left anterior: r = .193, p = .057; right anterior: r = .177, p = .076).
Posterior Positivity (P600)
As shown in Figure 4, a significant zero-order correlation was found in the 300-1000 ms TW across posterior sites (r = .279, p < .01). A significant partial correlation was also observed across these sites (r = .274, p < .05). Consistent with the results from the between-groups analyses, these correlations revealed that higher proficiency participants showed a larger P600 response. Additional analyses revealed a relationship between the size of the P600 across posterior sites and the presence of a positivity across anterior sites in the 300-700 ms time window: there were significant positive zero-order correlations between the posterior P600 and average difference amplitude between 300-700 ms across both left (r = .448, p < .0001) and right (r = .420, p < .0001) anterior sites. This relationship was stronger over medial sites in both left (medial: r = .505, p < .0001; lateral: r = .334, p < .001) and right (medial: r = .506, p < .0001; lateral: r = .263, p < .05) hemispheres.
Figure 4.

Correlation between average difference amplitude (violation – canonical, in μV) over posterior sites and proficiency in the 300-1000 ms time window (P600).
Socioeconomic Status
In addition to effects of proficiency, exploratory analyses using the correlational approach revealed significant relationships between SES and neural organization for syntactic processing, specific to left anterior sites. While a zero-order correlation across left anterior sites in the 100-300 ms time window only approached significance (r = -.176, p = .069), a partial correlation across these sites controlling for proficiency, WM, and education level was significant (r = -.218, p < .05), revealing that participants from higher SES backgrounds showed a larger amplitude negativity to syntactic violations over left anterior sites. This relationship was slightly stronger over left anterior lateral sites (partial correlation: r = -.223, p < .05) than over left anterior medial sites (partial correlation: r = -.197, p = .052).
In order to investigate the contributions of factors related to self-reported childhood experience, the relationship between left anterior average difference amplitude and SES was examined while controlling for the amount participants reported reading, being read to, watching television, and playing video games as children. These factors were found to moderate the relationship between the neural response to syntactic violations and SES, as the partial correlation between average difference amplitude over left anterior sites and SES was reduced over both the 100-300 ms (r = -.081, NS) and 300-700 ms (r = -.021, NS) time windows. None of these factors were found to moderate any of the correlations with proficiency described above.
Discussion
In this study event-related potentials elicited by phrase structure violations were examined as native speakers of English listened to simple sentences in English. Participants were recruited from a wide spectrum of society and given standardized measures of English language proficiency, and two types of analysis employing complementary approaches were performed. In the between groups analyses, participants were divided, based on standardized proficiency scores, into Lower Proficiency (LP) and Higher Proficiency (HP) groups. Analyses revealed differences in brain organization between the two proficiency groups, with HP participants showing an early anterior negativity that was more restricted both spatially and temporally, and a larger and more widely distributed positivity to violations in English. In the correlational analyses, we explored the relationship between proficiency and the neural organization for syntactic processing across a wide spectrum of proficiency scores by examining the degree to which individual proficiency scores correlated with individual neural responses to syntactic violations in regions and time windows identified in the between-group analyses. This approach also employed partial correlation analyses to control for possible confounding variables. These correlational analyses provided converging evidence for the effects of proficiency discovered in the between-groups analyses, confirming that differences in proficiency predict neural indices of syntactic processing indexed by both anterior negativity and posterior positivity components. Additionally, these analyses revealed links with socioeconomic status that were specific to left anterior sites. Below we discuss possible interpretations of these results and their implications for theories of sentence processing and future directions for research, and comment on the potential of research into proficiency differences for enriching our knowledge about language processing.
Anterior Negativity
While the component referred to as the left anterior negativity, or LAN, is so named because it most often has been reported to display a distribution which is maximal over left anterior electrode sites (e.g., Coulson, King, & Kutas, 1998b; Gunter, et al., 1999; Hahne & Friederici, 1999; Münte, et al., 1993; Neville, et al., 1991) this is not always the case. Several studies have reported an anterior negativity which is more bilateral in distribution (e.g., Friederici & Frisch, 2000; Friederici & Meyer, 2004; Friederici, et al., 1993; Frisch, Hahne, & Friederici, 2004; Hagoort, et al., 2003; Hahne, 2001; Hahne & Friederici, 1999, 2002; Hahne & Jescheniak, 2001; Münte, et al., 1997; Rossi, et al., 2006; Yamada & Neville, 2007; Ye, Luo, Friederici, & Zhou, 2006). To date no theory of language processing has sought to address the functional significance of these differences in distribution, although the question has been identified as an important one for future research (Hahne & Friederici, 2002).
While the differences in distribution of the anterior negativity effect have sometimes been attributed to subtle differences in the stimuli used (Hagoort, et al., 2003), in other cases a bilateral negativity has been found using the same stimuli and experimental methods as in other studies which did find a left lateralized anterior negativity (Hahne & Friederici, 2002). This raises the hypothesis that proficiency differences in the participants tested may have at least contributed to the observed differences in distribution. The results of the current study support this hypothesis. In the between-groups analyses, participants who scored higher on standardized tests of English proficiency showed a more focal, left-lateralized early anterior negativity to straightforward phrase structure violations in English, while participants who scored lower showed an affect that was more widespread both spatially and temporally. This between-groups spatial difference was reflected in an increased negativity over right anterior medial sites in the 100-300 ms time window in lower proficiency participants, a finding which was supported by a correlation between proficiency and average difference amplitude over right anterior medial sites in this time window.
The anterior negativity in this study began early, around 100 ms, and was elicited by insertion phrase structure violations. Such early components elicited by word category violations have been interpreted by some researchers to reflect early and automatic processes in which a word is integrated into the phrase structure of the preceding sentence fragment (Friederici, 2002). While other theories dispute the automaticity of these processes with regard to the influence of contextual information (van den Brink & Hagoort, 2004), both theories are consistent in their view that early anterior negative components reflect processes which make immediate use of incoming input to guide online parsing. Given these interpretations, our results suggest that such early and immediate sentence parsing processes may operate differently, or be used differently, in adult monolingual native speakers who differ in their linguistic proficiency when processing their native language.
A preliminary, testable functional explanation for these differences is that LP participants recruited additional resources related to early processing in order to parse straightforward phrase structure violations in simple sentences in their native language. Given the overall similarity of the effect reported here to those previously reported for monolinguals, and the spatial and temporal differences between groups, it seems likely that these differences reflect the recruitment of additional resources in similar systems. While the early effect is larger over right anterior medial sites in the LP group, there is still a small negative deflection in the same location in the HP group. Thus it appears that these differences may reflect the less efficient use in LP participants of neural systems similar to those used by HP participants. While in both groups these systems make immediate use of incoming input to guide syntactic parsing, in HP participants these systems are highly focal and short in duration. This interpretation is consistent with some ERP evidence from bilinguals. Some studies have failed to find an anterior negativity effect to phrase structure violations in low proficiency late bilinguals (Hahne, 2001; Hahne & Friederici, 2001), suggesting that a certain level of proficiency is necessary to recruit the neural mechanisms indexed by the anterior negativity for online syntactic processing. Other evidence suggests that when these resources are recruited in late bilinguals, the distribution is related to proficiency, as the anterior negativity to phrase structure violations has been reported to be more bilateral and widespread in distribution with increases in age of immersion and decreases in English proficiency (e.g., Rossi, et al., 2006; Weber-Fox & Neville, 1996). Evidence from neuroimaging studies of sentence processing also supports this interpretation. Activation in perisylvian language areas of the left hemisphere (e.g., Caplan, Alpert, & Waters, 1999; Caplan, et al., 2002; Stromswold, Caplan, Alpert, & Rauch, 1996) and bilaterally (Just, Carpenter, Keller, Eddy, & Thulborn, 1996) has been found to vary as a function of syntactic complexity and, presumably, the amount of resources necessary to process more complex syntactic structures.
The between-group analyses also revealed that the anterior negativity effect was more focal temporally in HP participants, while the effect was temporally more extended in LP participants. This was supported by the correlational analyses, which revealed that lower proficiency correlated with increased negativity in both the 300-700 ms and 700-1200 ms time windows. ERP studies of language development have reported a pattern in which language-related ERP effects become more focal spatially and temporally with increases in age and/or language ability (e.g., Adamson-Harris, et al., 2000; Hahne, Eckstein, & Friederici, 2004; Mills, et al., 1993; Neville, et al., 1993). While caution is necessary when comparing studies of children and adults, this raises the hypothesis that individual differences in the development of neural systems important for some aspects of language processing may endure into adulthood. This hypothesis will be discussed more below with relation to the correlational results with socioeconomic status.
The finding that the anterior negativity effect extended to the 700-1200 ms time window in the LP group was somewhat unexpected. Previous studies have reported an N400-like “wrap-up” effect which has been observed to the final word in sentences judged to be unacceptable due to either a semantic or syntactic violation (e.g., Hagoort, et al., 1993; Hagoort & Brown, 1999; Osterhout & Holcomb, 1992, 1993). The late negativity observed in this study could reflect similar processes engaged when participants encountered the final word in the sentence. While the distribution of this “wrap-up” effect is usually more widespread and central than the distribution of the late negativity observed here, the relatively late position of the violation creates a temporal and spatial overlap with the posterior positivity which makes a definitive interpretation of this effect difficult based solely on this evidence. However, additional evidence from a study using ERP and fMRI data gathered from the same participants in this paradigm allows for a degree of clarification regarding the interpretation of this component (Pakulak, 2008; Pakulak, Dow, & Neville, 2009). That analysis provides evidence that one of the neural generators of this component is anatomically close to the neural generator of the anterior negativity in the 100-300 ms time window, suggesting that the negativity over anterior sites in the later time window indexes processes similar to the negativity in the earlier time window.
Posterior Positivity (P600)
We found a typical biphasic response in which the anterior negativity was followed by a later, posterior positivity which is consistent in latency and distribution with the component typically referred to as the P600. The P600 has been interpreted as an index of more controlled processes related to syntactic repair and revision in the face of a violation (Friederici, 2002) or to the cost of syntactic integration (Kaan, et al., 2000), and more recently as an index of the amount of competition between alternative options as the parser attempts to unify linguistic elements in the comprehension of an utterance (Hagoort, 2003). As with the anterior negativity, the between-group analyses revealed differences in the P600 effect between groups: the P600 was larger in amplitude in the HP group compared to the LP group and was more widespread in distribution, extending to anterior medial sites in the HP group. These findings were confirmed by the correlational analyses, in which proficiency positively correlated with average difference amplitude over anterior sites in the 300-700 ms time window and over posterior sites in the 300-1000 ms time window.
A more frontally distributed P600 has been hypothesized to reflect processing difficulties associated with revision when the parser encounters grammatical but non-preferred continuations in syntactically complex and/or ambiguous sentences, while a P600 with a more posterior distribution has been hypothesized to reflect a parsing failure and/or resulting repair processes when the parser encounters a grammatical violation (Friederici, et al., 2002; Hagoort & Brown, 2000). Given these past findings concerning the distribution of the P600, it is unclear why straightforward, unambiguous violations would elicit a more frontally distributed P600 in the HP group. However, the correlational analyses revealed strong correlations between the positivity across anterior sites and the positivity across posterior sites across all 72 participants, suggesting that the frontally distributed P600 may reflect part of a more widespread effect indexing similar as opposed to separate processes. While both groups showed a posterior P600 to phrase structure violations, this effect was larger in HP participants and positively correlated with proficiency across all participants. The size of the P600 has been shown to be reduced when participants do not perform a grammaticality judgment task (Hahne & Friederici, 2002), which suggests that this difference in amplitude may reflect greater engagement of processes related to revision and repair in the context of the grammaticality judgment task on the part of the HP participants. However, as there was no task manipulation which explicitly required repair processes this interpretation is necessarily tentative and calls for more research.
Overall, the strength of the correlations between average difference amplitude and proficiency reveal that the strongest effects of proficiency were found in the 300-700 ms time window over anterior sites and in the 300-1000 ms time window over posterior sites. While lower proficiency is associated with an extended negativity or weak positivity over anterior sites and a smaller P600 across posterior sites, higher proficiency is associated with a robust and widespread P600 extending to anterior sites. While the functional interpretation of the P600 is still being clarified, it is clear that the process or processes indexed by this component are more engaged in participants with higher proficiency. Be they related to repair and/or reanalysis in the face of a violation, syntactic integration in general, or competition between alternatives in the unification of linguistic elements for comprehension, activation of processes which become engaged when the parser encounters difficulty is likely to be associated with better performance across a variety of tasks involving syntactic processing, and the results from this study show that this is indeed the case.
Optimal Neural Organization for Syntactic Processing
Overall, the results presented here present a profile of neural organization for syntactic processing which is predicted by higher proficiency. Reflected in this organization is an interaction between processes considered to be more automatic and those hypothesized to be more controlled. It is reasonable to propose that the response associated with higher proficiency represents the most efficient allocation of these processes: a more focal early anterior negativity, which indexes more efficient detection of word category violations, “frees up” more controlled resources involved with repair and reanalysis reflected in the widespread P600, and it is this allocation which represents an optimal neural organization for syntactic processing in monolinguals. While it is not possible to draw a definitive causal link between these processes, the relative timing makes it more likely that more efficient earlier processes would free up resources reflected in later processes and not the other way around. A different picture emerges with lower proficiency: less efficient automatic processes involved in the detection of word category violations, reflected in more widespread distribution spatially and temporally, are followed by reduced recruitment of more controlled processes. Here, again given the relative timing it is likely that the less efficient nature of these early processes does not allow for the recruitment of later processes in lower proficiency individuals to the same degree as in higher proficiency individuals.
Other Possible Contributing Factors
The group differences in syntactic processing found in this study have been discussed in terms of English language proficiency, and the correlational analyses allowed for the control for other possible mediating factors. Still, it is possible that other factors contributed to the results.
While in theory one possibility is that the pattern of results is due to differences in dialect, this is unlikely to explain the differences found. The experimental materials used were chosen to minimize any effects of dialect differences: the sentences used in the ERP paradigm were all simple, single-clause sentences created for a paradigm that was also used to investigate syntactic processing in 36 month-olds and the violations used were insertion phrase structure violations which are syntactic violations in any known dialect of English. All of the participants were native speakers of American English who came from the Eugene, Oregon area.
Another possibility is that individual resource limitations not explicitly controlled for are contributing to results found. One possibility is general intelligence. As our goal was to more completely characterize language proficiency while gathering all data in a single session, we were unable to include a direct assessment of intelligence. However, there is considerable evidence that WM correlates with intelligence (e.g., Conway, Cowan, Bunting, Therriault, & Minkoff, 2002), and the correlational analyses revealed that the results with regard to proficiency were not significantly affected by WM. Still, it is possible that differences in general intelligence contributed to our findings, and the relationship between general intelligence and brain organization for language is an important consideration in future research. Another factor which may have affected our results is attention. While we were unable to control for individual differences in the several types of attention, such as executive function and sustained endogenous attention, which may influence performance in experimental paradigms such as ours, several facts suggest that attention had little or no effect on our results. The behavioral measures were given in a small, enclosed room one-on-one with the lead author, a setting specifically designed to minimize differences in attention, and all participants performed the tasks in a similar time frame. In the between-groups analyses, all participants attended to the stimuli well enough to answer more than 84% of the probe questions accurately and score above 84% on the grammaticality judgment task, and only correct responses were used in the ERP analyses. As discussed above, the pattern of ERP results and modulation of ERP components reported are consistent with results from other populations (e.g., bilinguals). This pattern of results suggests that domain-general differences in attention are unlikely to have produced the specific pattern of results observed. More research is certainly needed into the interplay between WM, attention, intelligence, and language proficiency. Still, our results suggest that there exist proficiency differences in on-line syntactic processing which are independent of certain resource limitations, a finding which calls for further study.
Socioeconomic Status
This study also examined the relationship between linguistic proficiency and neural organization for syntactic processing in adults and the socioeconomic status environment in which they were raised. In both cases, significant relationships were found which raise important hypotheses for future research.
In the correlational analyses of 72 participants, self-reported childhood SES correlated significantly with adult linguistic proficiency as assessed by standardized measures. This is agreement with a large literature documenting profound effects of SES on language in children (e.g., Dollaghan, et al., 1999; Hart & Risley, 1995; Hoff, 2003; Hoff, Laursen, & Tardif, 2002; Huttenlocher, Vasilyeva, Cymerman, & Levine, 2002; Noble, McCandliss, & Farah, 2007; Noble, Norman, & Farah, 2005) and raises the hypothesis that such differences endure into adulthood. While these analyses are correlational and therefore can not make any causal inferences or rule out genetic or epigenetic factors, additional evidence supports this hypothesis. Several self-reported measures of childhood environment - amount of time spent reading, being read to, watching television, and playing video games - correlated significantly with childhood SES, and there were relationships between proficiency and all of these measures but one. Of particular relevance is that the two measures of childhood reading showed the strongest correlations with SES, and both showed a positive relationship with adult proficiency. This is consistent with evidence that time spent reading or reading with an adult can have a positive effect on language development (Payne & et al., 1994; Scarborough & Dobrich, 1994). Other evidence suggests that there are differences in the speech mothers use when reading with their children, such as using more utterances per unit of time, utterances with greater structural complexity, and a larger vocabulary, and that these speech differences can affect language development (Hoff-Ginsberg, 1991; Hoff, 2006). Additionally, there is evidence that interventions which specifically seek to promote parent-child reading can positively affect vocabulary development (Mendelsohn, et al., 2001; Sharif, Reiber, & Ozuah, 2002). While caution is necessary, evidence for some specificity in the relationship between childhood environmental factors and adult proficiency raises the hypothesis that some effects of childhood environment related to SES may have enduring effects on language proficiency. Future research which specifically addresses causal relationships with the use of targeted interventions, and which specifically addresses the role of other factors in the relationship between childhood SES and language development, will provide important evidence bearing on this hypothesis.
We also observed a correlation between childhood SES and the neural response to syntactic violations, such that participants from higher SES backgrounds showed a larger negative response to violations than participants from lower SES backgrounds. This effect was specific to left anterior sites in both the 100-300 ms and 300-700 ms time windows, and the results of the partial correlation analysis revealed that this effect was independent of proficiency, as well as working memory and education level. However, given the results discussed above with regard to SES and proficiency, the interpretation that the relationship between SES and left anterior negativity amplitude is independent of proficiency should be treated with caution. It is possible that this relationship is moderated to some degree by aspects of linguistic proficiency which were not reflected by the proficiency measures used, though further clarification is beyond the scope of this study. Still, these results raise the additional hypothesis that differences related to childhood SES environment which can affect language development may also have effects on neural organization for language which endure into adulthood. An alternate hypothesis is that these differences in the neural response of left anterior sites are the result of genetic differences which covary with our measurement of childhood SES. However, this relationship disappeared when childhood environmental factors - amount of time spent reading, being read to, watching television, and playing video games – were controlled for. This provides evidence that our measure of childhood SES is a mediating variable for specific aspects of childhood environment and strengthens the hypothesis that experiential factors may have effects on neural organization for syntactic processing which endure into adulthood. While necessarily preliminary and cautious, the hypothesis that environmental effects of SES which impact language development may also have a lasting impact on neural organization for syntactic processing provides an intriguing direction for future research.
The results relating to SES discussed above were the result of correlational analyses utilizing data collected from 72 adults from a wide spectrum of society and a wide range of childhood SES backgrounds. Reviews of results from studies of the effects of SES on language development in children suggest that the magnitude of differences related to SES depends on the range of SES in the sample studied (e.g., Hoff, 2006). Overall, this suggests that in order to obtain a more complete picture of neural organization for syntactic processing, or of neural organization more generally, it will be important to study samples from a wider and more diverse spectrum of society.
References
- Adamson-Harris AM, Mills DL, Neville HJ. Children's processing of grammatical and semantic information within sentences: Evidence from event-related potentials. Cognitive Neuroscience Society 2000;7:58. [Google Scholar]
- Capek C, Corina D, Grossi B, McBurney SL, Neville HJ, Newman AJ, et al. Electrophysiological evidence for the effects of age of acquisition on brain systems mediating semantic and syntactic processing in American Sign Language. Cognitive Neuroscience Society 2002;9:135. [Google Scholar]
- Capek CM, Grossi G, Newman AJ, McBurney SL, Corina D, Roeder B, et al. Brain systems mediating semantic and syntactic processing in deaf native signers: biological invariance and modality specificity. Proc Natl Acad Sci U S A. 2009;106(21):8784–8789. doi: 10.1073/pnas.0809609106. Journal Article; Research Support, N.I.H., Extramural; Research Support, Non-U.S. Gov't. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caplan D, Alpert N, Waters G. PET studies of syntactic processing with auditory sentence presentation. Neuroimage. 1999;9(3):343–351. doi: 10.1006/nimg.1998.0412. [DOI] [PubMed] [Google Scholar]
- Caplan D, Vijayan S, Kuperberg G, West C, Waters G, Greve D, et al. Vascular responses to syntactic processing: event-related fMRI study of relative clauses. Hum Brain Mapp. 2002;15(1):26–38. doi: 10.1002/hbm.1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conway AR, Cowan N, Bunting MF, Therriault DJ, Minkoff SR. A latent variable analysis of working memory capacity, short-term memory capacity, processing speed, and general fluid intelligence. Intelligence. 2002;30(2):163–184. [Google Scholar]
- Coulson S, King JW, Kutas M. ERPs and domain specificity: Beating a straw horse. Language and Cognitive Processes. 1998a Dec;13(6):653–672. 1998. [Google Scholar]
- Coulson S, King JW, Kutas M. Expect the Unexpected: Event-Related Brain Response to Morphosyntactic Violations. Language and Cognitive Processes. 1998b;13(1):21–58. [Google Scholar]
- Daneman M, Carpenter PA. Individual differences in working memory and reading. Journal of Verbal Learning & Verbal Behavior. 1980 Aug;19(4):450–466. [Google Scholar]
- Dollaghan CA, Campbell TF, Paradise JL, Feldman HM, Janosky JE, Pitcairn DN, et al. Maternal education and measures of early speech and language. Journal of Speech, Language, and Hearing Research. 1999 Dec;42(6):1432–1443. doi: 10.1044/jslhr.4206.1432. [DOI] [PubMed] [Google Scholar]
- Friederici AD. The time course of syntactic activation during language processing: a model based on neuropsychological and neurophysiological data. Brain Lang. 1995;50(3):259–281. doi: 10.1006/brln.1995.1048. [DOI] [PubMed] [Google Scholar]
- Friederici AD. Towards a neural basis of auditory sentence processing. Trends in Cognitive Sciences. 2002;6(2):78–84. doi: 10.1016/s1364-6613(00)01839-8. [DOI] [PubMed] [Google Scholar]
- Friederici AD, Frisch S. Verb argument structure processing: The role of verb-specific and argument-specific information. Journal of Memory and Language. 2000;43(3):476–507. [Google Scholar]
- Friederici AD, Hahne A, Saddy D. Distinct neurophysiological patterns reflecting aspects of syntactic complexity and syntactic repair. Journal of Psycholinguistic Research. 2002;31(1):45–63. doi: 10.1023/a:1014376204525. [DOI] [PubMed] [Google Scholar]
- Friederici AD, Mecklinger A. Syntactic parsing as revealed by brain responses: first-pass and second-pass parsing processes. J Psycholinguist Res. 1996;25(1):157–176. doi: 10.1007/BF01708424. [DOI] [PubMed] [Google Scholar]
- Friederici AD, Meyer M. The brain knows the difference: two types of grammatical violations. Brain Res. 2004;1000(1-2):72–77. doi: 10.1016/j.brainres.2003.10.057. [DOI] [PubMed] [Google Scholar]
- Friederici AD, Pfeifer E, Hahne A. Event-related brain potentials during natural speech processing: Effects of semantic, morphological and syntactic violations. Cognitive Brain Research. 1993;1(3):183–192. doi: 10.1016/0926-6410(93)90026-2. [DOI] [PubMed] [Google Scholar]
- Friederici AD, Steinhauer K, Mecklinger A, Meyer M. Working memory constraints on syntactic ambiguity resolution as revealed by electrical brain responses. Biol Psychol. 1998;47(3):193–221. doi: 10.1016/s0301-0511(97)00033-1. [DOI] [PubMed] [Google Scholar]
- Frisch S, Hahne A, Friederici AD. Word category and verb-argument structure information in the dynamics of parsing. Cognition. 2004;91(3):191–219. doi: 10.1016/j.cognition.2003.09.009. [DOI] [PubMed] [Google Scholar]
- Gunter TC, Friederici AD, Hahne A. Brain responses during sentence reading: Visual input affects central processes. Neuroreport: For Rapid Communication of Neuroscience Research. 1999;10(15):3175–3178. doi: 10.1097/00001756-199910190-00009. [DOI] [PubMed] [Google Scholar]
- Hagoort P. How the brain solves the binding problem for language: a neurocomputational model of syntactic processing. NeuroImage. 2003;20:S18–29. doi: 10.1016/j.neuroimage.2003.09.013. [DOI] [PubMed] [Google Scholar]
- Hagoort P. On broca, brain, and binding: A new framework. Trends in Cognitive Sciences. 2005 Sep;9(9):416–423. doi: 10.1016/j.tics.2005.07.004. [DOI] [PubMed] [Google Scholar]
- Hagoort P, Brown C, Groothusen J. The Syntactic Positive Shift (SPS) as an ERP Measure of Syntactic Processing. Language and Cognitive Processes. 1993;8(4):439–483. [Google Scholar]
- Hagoort P, Brown CM. Gender electrified: ERP evidence on the syntactic nature of gender processing. J Psycholinguist Res. 1999;28(6):715–728. doi: 10.1023/a:1023277213129. [DOI] [PubMed] [Google Scholar]
- Hagoort P, Brown CM. ERP effects of listening to speech compared to reading: the P600/SPS to syntactic violations in spoken sentences and rapid serial visual presentation. Neuropsychologia. 2000;38(11):1531–1549. doi: 10.1016/s0028-3932(00)00053-1. [DOI] [PubMed] [Google Scholar]
- Hagoort P, Wassenaar M, Brown CM. Syntax-related ERP-effects in Dutch. Cognitive Brain Research. 2003;16(1):38–50. doi: 10.1016/s0926-6410(02)00208-2. [DOI] [PubMed] [Google Scholar]
- Hahne A. What's different in second-language processing? Evidence from event-related brain potentials. Journal of Psycholinguistic Research. 2001;30(3):251–266. doi: 10.1023/a:1010490917575. [DOI] [PubMed] [Google Scholar]
- Hahne A, Eckstein K, Friederici AD. Brain signatures of syntactic and semantic processes during children's language development. Journal of Cognitive Neuroscience. 2004 Sep;16(7):1302–1318. doi: 10.1162/0898929041920504. [DOI] [PubMed] [Google Scholar]
- Hahne A, Friederici AD. Electrophysiological evidence for two steps in syntactic analysis: Early automatic and late controlled processes. Journal of Cognitive Neuroscience. 1999;11(2):194–205. doi: 10.1162/089892999563328. [DOI] [PubMed] [Google Scholar]
- Hahne A, Friederici AD. Processing a second language: Late learners' comprehension mechanisms as revealed by event-related brain potential. Bilingualism: Language & Cognition. 2001;4(2):123–141. [Google Scholar]
- Hahne A, Friederici AD. Differential task effects on semantic and syntactic processes as revealed by ERPs. Cognitive Brain Research. 2002;13(3):339–356. doi: 10.1016/s0926-6410(01)00127-6. [DOI] [PubMed] [Google Scholar]
- Hahne A, Jescheniak JD. What's left if the Jabberwock gets the semantics? An ERP investigation into semantic and syntactic processes during auditory sentence comprehension. Cognitive Brain Research. 2001;11(2):199–212. doi: 10.1016/s0926-6410(00)00071-9. [DOI] [PubMed] [Google Scholar]
- Hammil DD, Brown VL, Larsen SC, Wiederholt JL. Test of adolescent and adult language. 3rd. Austin, Tx.: 1994. [Google Scholar]
- Hart B, Risley TR. Meaningful differences in the everyday experience of young American children. Baltimore: P.H. Brookes; 1995. [Google Scholar]
- Hoff-Ginsberg E. Mother-child conversation in different social classes and communicative settings. Child Development. 1991 Aug;62(4):782–796. doi: 10.1111/j.1467-8624.1991.tb01569.x. [DOI] [PubMed] [Google Scholar]
- Hoff E. The specificity of environmental influence: socioeconomic status affects early vocabulary development via maternal speech. Child Dev. 2003;74(5):1368–1378. doi: 10.1111/1467-8624.00612. [DOI] [PubMed] [Google Scholar]
- Hoff E. How social contexts support and shape language development. Developmental Review. 2006 Mar;26(1):55–88. [Google Scholar]
- Hoff E, Laursen B, Tardif T. Socioeconomic status and parenting. In: Bornstein Marc H., editor. Handbook of parenting: Vol 2: Biology and ecology of parenting. 2nd 2002. [Google Scholar]
- Holcomb PJ, Neville HJ. Natural speech processing: An analysis using event-related brain potentials. Psychobiology. 1991;19(4):286–300. [Google Scholar]
- Hollingshead A. Four factor index of social status. New Haven: Yale University Department of Sociology; 1975. Unpublished manuscript. [Google Scholar]
- Huttenlocher J, Vasilyeva M, Cymerman E, Levine S. Language input and child syntax. Cognitive Psychology. 2002;45(3):337–374. doi: 10.1016/s0010-0285(02)00500-5. [DOI] [PubMed] [Google Scholar]
- Isel F, Hahne A, Maess B, Friederici AD. Neurodynamics of sentence interpretation: ERP evidence from French. Biological Psychology. 2007;74(3):337–346. doi: 10.1016/j.biopsycho.2006.09.003. [DOI] [PubMed] [Google Scholar]
- Just MA, Carpenter PA. A capacity theory of comprehension: Individual differences in working memory. Psychological Review. 1992;99(1):122–149. doi: 10.1037/0033-295x.99.1.122. [DOI] [PubMed] [Google Scholar]
- Just MA, Carpenter PA, Keller TA, Eddy WF, Thulborn KR. Brain activation modulated by sentence comprehension. Science. 1996;274(5284):114–116. doi: 10.1126/science.274.5284.114. [DOI] [PubMed] [Google Scholar]
- Kaan E, Harris A, Gibson E, Holcomb P. The P600 as an index of syntactic integration difficulty. Language & Cognitive Processes. 2000;15(2):159–201. [Google Scholar]
- Kaan E, Swaab TY. Electrophysiological evidence for serial sentence processing: A comparison between non-preferred and ungrammatical continuations. Cognitive Brain Research. 2003a;17(3):621–635. doi: 10.1016/s0926-6410(03)00175-7. [DOI] [PubMed] [Google Scholar]
- Kaan E, Swaab TY. Repair, revision, and complexity in syntactic analysis: An electrophysiological differentiation. Journal of Cognitive Neuroscience. 2003b;15(1):98–110. doi: 10.1162/089892903321107855. [DOI] [PubMed] [Google Scholar]
- King J, Just MA. Individual differences in syntactic processing: The role of working memory. Journal of Memory and Language. 1991;30(5):580–602. [Google Scholar]
- King JW, Kutas M. Who did what and when? Using word- and clause-level ERPs to monitor working memory usage in reading. Journal of Cognitive Neuroscience. 1995;7(3):376–395. doi: 10.1162/jocn.1995.7.3.376. [DOI] [PubMed] [Google Scholar]
- Kutas M, Hillyard SA. Reading senseless sentences: Brain potentials reflect semantic incongruity. Science. 1980;207(4427):203–205. doi: 10.1126/science.7350657. [DOI] [PubMed] [Google Scholar]
- Linebarger MC, Schwartz MF, Saffran EM. Sensitivity to grammatical structure in so-called agrammatic aphasics. Cognition. 1983 May;13(3):361–392. doi: 10.1016/0010-0277(83)90015-x. [DOI] [PubMed] [Google Scholar]
- McCarthy G, Wood CC. Scalp distributions of event-related potentials: an ambiguity associated with analysis of variance models. Electroencephalogr Clin Neurophysiol. 1985;62(3):203–208. doi: 10.1016/0168-5597(85)90015-2. [DOI] [PubMed] [Google Scholar]
- Mecklinger A, Schriefers H, Steinhauer K, Friederici AD. Processing relative clauses varying on syntactic and semantic dimensions: An analysis with event-related potentials. Memory & Cognition. 1995;23(4):477–494. doi: 10.3758/bf03197249. [DOI] [PubMed] [Google Scholar]
- Mendelsohn AL, Mogilner LN, Dreyer BP, Forman JA, Weinstein SC, Broderick M, et al. The impact of a clinic-based literacy intervention on language development in inner-city preschool children. Pediatrics. 2001;107(1):130–134. doi: 10.1542/peds.107.1.130. [DOI] [PubMed] [Google Scholar]
- Mills DL, Coffey-Corina SA, Neville HJ. Language acquisition and cerebral specialization in 20-month-old infants. Journal of Cognitive Neuroscience. 1993;5:317–334. doi: 10.1162/jocn.1993.5.3.317. [DOI] [PubMed] [Google Scholar]
- Münte TF, Matzke M, Johannes S. Brain activity associated with syntactic incongruencies in words and pseudo-words. Journal of Cognitive Neuroscience. 1997;9(3):318–329. doi: 10.1162/jocn.1997.9.3.318. [DOI] [PubMed] [Google Scholar]
- Münte TJ, Heinze H, Mangun G. Dissociation of brain activity related to syntactic and semantic aspects of language. Journal of Cognitive Neuroscience. 1993;5(3):335–344. doi: 10.1162/jocn.1993.5.3.335. [DOI] [PubMed] [Google Scholar]
- Neville HJ, Coffey SA, Holcomb PJ, Tallal P. The neurobiology of sensory and language processing in language-impaired children. Journal of Cognitive Neuroscience. 1993 Spr;5(2):235–253. doi: 10.1162/jocn.1993.5.2.235. [DOI] [PubMed] [Google Scholar]
- Neville HJ, Coffey SA, Lawson DS, Fischer A, et al. Neural systems mediating American Sign Language: Effects of sensory experience and age of acquisition. Brain and Language. 1997 May;57(3):285–308. doi: 10.1006/brln.1997.1739. [DOI] [PubMed] [Google Scholar]
- Neville HJ, Mills DL, Lawson DS. Fractionating language: Different neural subsystems with different sensitive periods. Cerebral Cortex. 1992 May-Jun;2(3):244–258. doi: 10.1093/cercor/2.3.244. [DOI] [PubMed] [Google Scholar]
- Neville HJ, Nicol J, Barss A, Forster K, Garrett M. Syntactically based sentence processing classes: Evidence from event-related brain potentials. Journal of Cognitive Neuroscience. 1991;3:155–170. doi: 10.1162/jocn.1991.3.2.151. [DOI] [PubMed] [Google Scholar]
- Newman AJ, Bavelier D, Corina D, Jezzard P, Neville HJ. A critical period for right hemisphere recruitment in American Sign Language processing. Nature Neuroscience. 2002 Jan;5(1):76–80. doi: 10.1038/nn775. [DOI] [PubMed] [Google Scholar]
- Newport E. Maturational constraints on language learning. Cognitive Science. 1990;14:11–28. [Google Scholar]
- Noble KG, McCandliss BD, Farah MJ. Socioeconomic gradients predict individual differences in neurocognitive abilities. Developmental Science. 2007 Jul;10(4):464–480. doi: 10.1111/j.1467-7687.2007.00600.x. [DOI] [PubMed] [Google Scholar]
- Noble KG, Norman MF, Farah MJ. Neurocognitive correlates of socioeconomic status in kindergarten children. Dev Sci. 2005;8(1):74–87. doi: 10.1111/j.1467-7687.2005.00394.x. [DOI] [PubMed] [Google Scholar]
- Osterhout L, Holcomb PJ. Event-related brain potentials elicited by syntactic anomaly. Journal of Memory & Language. 1992;31(6):785–806. [Google Scholar]
- Osterhout L, Holcomb PJ. Event-related potentials and syntactic anomaly: Evidence of anomaly detection during the perception of continuous speech. Language & Cognitive Processes. 1993;8(4):413–437. [Google Scholar]
- Osterhout L, Holcomb PJ, Swinney DA. Brain potentials elicited by garden-path sentences: Evidence of the application of verb information during parsing. JEP:LMC. 1995;20:786–803. doi: 10.1037//0278-7393.20.4.786. [DOI] [PubMed] [Google Scholar]
- Osterhout L, Mobley LA. Event-related brain potentials elicited by failure to agree. Journal of Memory & Language. 1995;34(6):739–773. [Google Scholar]
- Pakulak E. Unpublished Doctoral dissertation. University of Oregon; Eugene: 2008. An investigation of the effects of proficiency and age of acquisition on neural organization for syntactic processing using ERPs and fMRI. [Google Scholar]
- Pakulak E, Dow M, Neville HJ. An investigation of the neural generators of ERP indicies of syntactic processing using proficiency-related ERP modulations in an ERP-fMRI paradigm. Paper presented at the Cognitive Neuroscience Society; San Francisco, CA. 2009. [Google Scholar]
- Payne AC, et al. The Role of Home Literacy Environment in the Development of Language Ability in Preschool Children from Low-Income Families. Early Childhood Research Quarterly. 1994;9(3-4):427–440. [Google Scholar]
- Rossi S, Gugler MF, Friederici AD, Hahne A. The impact of proficiency on syntactic second-language processing of German and Italian: evidence from event-related potentials. Journal of cognitive neuroscience. 2006;18(12):2030–2048. doi: 10.1162/jocn.2006.18.12.2030. [DOI] [PubMed] [Google Scholar]
- Scarborough HS, Dobrich W. On the efficacy of reading to preschoolers. Developmental Review. 1994 Sep;14(3):245–302. [Google Scholar]
- Sharif I, Reiber S, Ozuah PO. Exposure to Reach Out and Read and vocabulary outocmes in inner city preschoolers. Journal of the National Medical Association. 2002 Mar;94(3):171–177. [PMC free article] [PubMed] [Google Scholar]
- Stromswold K, Caplan D, Alpert N, Rauch S. Localization of syntactic comprehension by positron emission tomography. Brain & Language. 1996;52(3):452–473. doi: 10.1006/brln.1996.0024. [DOI] [PubMed] [Google Scholar]
- van den Brink D, Hagoort P. The influence of semantic and syntactic context constraints on lexical selection and integration in spoken-word comprehension as revealed by ERPs. Journal of Cognitive Neuroscience. 2004 Jul-Aug;16(6):1068–1084. doi: 10.1162/0898929041502670. [DOI] [PubMed] [Google Scholar]
- Vos SH, Friederici AD. Intersentential syntactic context effects on comprehension: The role of working memory. Cognitive Brain Research. 2003;16(1):111–122. doi: 10.1016/s0926-6410(02)00226-4. [DOI] [PubMed] [Google Scholar]
- Vos SH, Gunter TC, Kolk HHJ, Mulder G. Working memory constraints on syntactic processing: An electrophysiological investigation. Psychophysiology. 2001;38(1):41–63. [PubMed] [Google Scholar]
- Vos SH, Gunter TC, Schriefers H, Friederici AD. Syntactic parsing and working memory: The effects of syntactic complexity, reading span, and concurrent load. Language & Cognitive Processes. 2001;16(1):65–103. [Google Scholar]
- Weber-Fox C, Davis LJ, Cuadrado E. Event-related brain potential markers of high-language proficiency in adults. Brain and Language. 2003;85:231–244. doi: 10.1016/s0093-934x(02)00587-4. [DOI] [PubMed] [Google Scholar]
- Weber-Fox C, Neville HJ. Maturational constraints on functional specializations for language processing: ERP and behavioral evidence in bilingual speakers. Journal of Cognitive Neuroscience. 1996;8(3):231–256. doi: 10.1162/jocn.1996.8.3.231. [DOI] [PubMed] [Google Scholar]
- Yamada Y, Neville HJ. An ERP study of syntactic processing in English and nonsense sentences. Brain Research. 2007 Jan;1130:167–180. doi: 10.1016/j.brainres.2006.10.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye Z, Luo Yj, Friederici AD, Zhou X. Semantic and syntactic processing in Chinese sentence comprehension: Evidence from event-related potentials. Brain Research. 2006;1071(1):186–196. doi: 10.1016/j.brainres.2005.11.085. [DOI] [PubMed] [Google Scholar]