

Abstract
Humans have five members of the well conserved RecQ helicase family: RecQ1, Bloom syndrome protein (BLM), Werner syndrome protein (WRN), RecQ4, and RecQ5, which are all known for their roles in maintaining genome stability. BLM, WRN, and RecQ4 are associated with premature aging and cancer predisposition. Of the three, RecQ4's biological and cellular roles have been least thoroughly characterized. Here we tested the helicase activity of purified human RecQ4 on various substrates. Consistent with recent results, we detected ATP-dependent RecQ4 unwinding of forked duplexes. However, our results provide the first evidence that human RecQ4's unwinding is independent of strand annealing, and that it does not require the presence of excess ssDNA. Moreover, we demonstrate that a point mutation of the conserved lysine in the Walker A motif abolished helicase activity, implying that not the N-terminal portion, but the helicase domain is solely responsible for the enzyme's unwinding activity. In addition, we demonstrate a novel stimulation of RecQ4's helicase activity by replication protein A, similar to that of RecQ1, BLM, WRN, and RecQ5. Together, these data indicate that specific biochemical activities and protein partners of RecQ4 are conserved with those of the other RecQ helicases.
1. INTRODUCTION
The RecQ family represents a group of helicases well conserved from bacteria to humans. Unlike bacteria and yeast, which have only one family member, humans have five distinct helicases: RecQ1, Bloom syndrome protein (BLM), Werner syndrome protein (WRN), RecQ4, and RecQ5. To date, three of these, BLM, WRN, and RecQ4, have been linked to premature aging and cancer predisposition. While the roles of BLM and WRN in DNA repair, DNA replication, and telomere maintenance have been characterized extensively, relatively little is known about the biological and cellular functions of RecQ4 [1-5].
RecQ4 deficiencies have been linked to three rare autosomal recessive diseases – Baller-Gerold syndrome, RAPADILINO syndrome, and Rothmund-Thomson syndrome (RTS). RTS symptoms include developmental abnormalities, growth deficiencies, proneness to cancer, predominantly osteosarcomas, and premature aging, including development of cataracts and hair loss [6, 7]. Cells from RTS patients display chromosomal instability and aneuploidy [7-11], in addition to sensitivity to replication inhibitors and oxidative stress [12, 13]. Although RTS is not exclusively caused by defects in the RECQ4 gene, a majority of RTS patients have mutations in RECQ4, most within the conserved helicase domain [14-16]. Together these results are indicative of a role for RecQ4 in processes that maintain genome stability and suggest that the helicase activity is important for RecQ4's biological and cellular roles.
More recently the cellular roles of RecQ4 in DNA replication and repair have been explored. Studies of the RecQ4 homolog in Xenopus revealed that it is important for loading replication factors at origins of replication [17]. Consistent with this work, subsequent reports showed that Xenopus RecQ4 promotes association of polymerase α with chromatin during replication initiation [18].
Additionally, Xenopus RecQ4 is loaded in a replication-independent manner onto chromatin containing double strand breaks, suggesting a role for RecQ4 in DNA repair processes, as well [19]. DrosophilaRECQ4 mutants display sensitivity to gamma irradiation, along with deficiency in repair of double strand breaks in vivo, in support of RecQ4 participation in double strand break repair [20]. Moreover, following treatment with the double strand break-inducing agent etoposide, RecQ4 co-localized with both Rad51 foci and regions of single-stranded DNA (ssDNA), implying its involvement in the homologous recombination pathway of double strand break repair [21].
RecQ4 participation in DNA repair also extends to other pathways. A recent study demonstrated that, following UV irradiation, RecQ4 co-localized and interacted directly with xeroderma pigmentosum group A protein, which mediates the associations of repair machinery around damaged DNA during nucleotide excision repair (NER) [22]. In addition, roles of RecQ4 in base excision repair (BER) have been recently discovered. RecQ4 physically interacts with poly-(ADP ribose) polymerase-1, which has an important role in regulation of BER through (ADP-ribosyl)ation of component proteins [23]. RecQ4 stimulation of key players in BER, namely DNA polymerase β, apurinic endonuclease 1 (APE1), and flap endonuclease 1 (FEN1) is evidence of a direct functional involvement of RecQ4 in BER, as well [24]. Furthermore, studies with RTS cells showed increased formation of strand breaks upon exposure to hydrogen peroxide and accumulation of XRCC1 foci following oxidative stress [24]. Also, treatment with the single strand break-inducing agent paraquat resulted in decreased survival in Drosophila RECQ4 mutants [20]. Together these results signify a role of RecQ4 in single strand break repair.
Although the precise mechanisms of RecQ4 functions in these cellular replication and repair processes have yet to be elucidated, they are likely linked to its biochemical activities. Generally, the RecQ helicases RecQ1, BLM, WRN, and RecQ5 have similar biochemical activities, including 3′,-5′, ATP-dependent DNA unwinding and strand annealing [4]. Initial biochemical characterization of RecQ4 demonstrated ATPase and strand annealing activities without detectable unwinding activity [25, 26]. However, in recent studies, purified human RecQ4 displayed helicase activity [27, 28]. Xu and Liu proposed that the helicase activity of RecQ4 is relatively weak compared to its strand annealing activity, which regenerates the native substrate following unwinding. RecQ4's helicase activity was only revealed when excess of ssDNA was used to trap the released strand [28]. Based on its homology to the other RecQ helicases and on the conserved strand annealing and ATPase activities, it is not surprising that RecQ4 would also display helicase activity. Yet, the unwinding activity of RecQ4 has not been fully characterized. In the current report we set out to further analyze the helicase activity of purified human RecQ4 through studies in vitro.
2. MATERIALS AND METHODS
2.1 Enzyme expression and purification
Wild-type (WT) human RecQ4 with a C-terminal 9-histidine tag in the pGEX6p1 vector (GE Healthcare) was expressed and purified from E. coli Rosetta2 (DE3) (Novagen) as described previously, with the following modifications [25]. Cells were lysed by sonication pulses 30 sec on then 30 sec off for a total of 7 min at 50% power in lysis buffer containing 50 mM Tris-HCl pH 7.5, 200 mM KCl, 10% sucrose, 2 mM EDTA, 1 mM DTT, 0.01% Igepal (Sigma), and 5 μg/ml each of aprotinin, chymostatin, leupeptin, and pepstatin A protease inhibitors. The extract was clarified by low speed centrifugation at 8,000 rpm (rotor JA-12, Beckman Coulter) for 15 min followed by ultracentrifugation at 40,000 rpm (rotor 60 Ti, Beckman Coulter) for 30 min. Lysate was passed through a 70 ml Q Sepharose column (GE Healthcare) and then onto a 40 ml SP Sepharose column (GE Healthcare). Protein was eluted by a gradient of 200-660 mM KCl in K Buffer (20 mM KH2PO4, 10 % glycerol, 0.5 mM EDTA, 0.01% Igepal, and 1 mM DTT). The glutathione-sepharose fast flow matrix (GE Healthcare) was washed with three times 30 ml K buffer containing 500 mM KCl. Following elution from the nickel-NTA agarose (Novagen) with 300 mM imidazole in K buffer containing 500 mM KCl, the RecQ4 was identified by electrophoresis and Coomassie Blue staining of a polyacrylamide gel. The RecQ4 was pooled, dialyzed in K buffer containing 500 mM KCl overnight at 4°C to remove the imidazole, aliquoted, and stored at −80°C.
Human RecQ4 mutant K508M (KM) was generated by PCR amplification of the WT RecQ4 construct using forward and reverse primers 5′-ATGTCCCTGTGCTACCAGCTC-3′ and 5′-GCCGGCACCTGTAGGCAGCAC-3′, respectively. PCR product was ligated and amplified in bacteria. Final sequence-verified construct was expressed in E. coli Rosetta2 (DE3) (Novagen). Expression and purification of the KM RecQ4 was as described above for WT. Final fractions of both WT and KM RecQ4 were analyzed by SDS-PAGE and Coomassie Blue gel staining. Human RPA was expressed from plasmid vector p11d-tRPA (gift from Dr. Marc S. Wold) [29] in E.coli BL21 (Novagen) and purified as previously described [30]. Human WRN was expressed and purified using a baculovirus/Sf9 insect cell system as previously described [31].
2.2 Oligonucleotide substrates
Oligonucleotides were synthesized by Integrated DNA Technologies (Coralville, IA), and sequences are listed in Table 1 and Supplementary Table 1. Primers T1-9, HJ1, and G1 were radiolabeled at the 5′ end with [γ-32P] ATP (PerkinElmer Life Sciences) and T4 polynucleotide kinase (New England Biolabs). Following labeling, unincorporated [γ-32P] ATP was removed from T1-9 using MicroSpin G-25 columns (GE Healthcare). Then to anneal the fork and full duplex substrates, corresponding oligonucleotides were combined in annealing buffer (40 mM Tris-HCl pH 8.0 and 50 mM NaCl) in a 1:2 ratio (labeled to unlabeled oligonucleotide), heated at 95°C for 5 minutes, and cooled gradually to room temperature. The Holliday junction [32, 33], and G-quadruplex [34, 35] substrates were prepared as previously described. Substrates used for individual experiments are indicated in the figure legends and depicted in each figure.
Table 1.
Oligonucleotide sequences
| Primer | Sequence |
|---|---|
| Top (5′-3′) | |
| T1 | GACGCTGCCGAATTCTACCAGTGCCTTGCTAGGACATCTTTGCCCACCTG CAGGTTCACCC |
| T2 | GGAATTCTACCAGTGCCTTGCTAGGACATCTTTGCCCA |
| T3 | GTAGTGCATGTACACCACACTCTTTTTTTTTTTTTTT |
| T4 | GACGCTGCCGAATTCTACCAGTGCCTTGCTAGGACATCTTTGCCCA |
| Bottom (3′-5′) | |
| B1 | TGCGACGGCTTAAGATGGTCACGGAACGATGTACCTCGACAGATCTCCT AGGCTGATAGCTA |
| B2 | CTTAAGATGGTCACGGAACGATGTACCTCGACAGATC |
| B3 | CATCACGTACATGTGGTGTGAGTTTTTTTTTTTTTTT |
| B4 | CTAGACAGCTCCATGTAGCAAGGCACTGGTAGAATTCGGCAGCGT |
2.3 Enzyme Assays
All assays were performed at least in triplicate. For helicase assays, RecQ4 (amount indicated in figure legend) was incubated with substrate (0.5 nM) for 30 minutes at 37°C in 10 μl helicase buffer containing 30 mM Tris pH 7.4, 5 mM MgCl2, 1 mM DTT, 100 μg/ml BSA, 10% glycerol, and 5 mM ATP (unless noted otherwise in figure legend). Helicase reactions were stopped by addition of 10 μl of 3X native stop dye (50mM EDTA, 38% glycerol, 0.9% SDS, 0.05% bromophenol blue, and 0.05% xylene cyanol). As indicated in the figure legends, following addition of stop dye, select heat denatured substrates were generated by incubation at 95°C for 5 minutes. Products were separated by electrophoresis on 12% native polyacrylamide gels and exposed to a PhosphorImager plate (GE Healthcare). Results were analyzed using ImageQuant version 5.2 (GE Healthcare). Following quantitation of each experiment, error bars were calculated as the standard deviation of the data.
For strand annealing assays, RecQ4 (amount indicated in figure legend) was incubated with single-stranded radiolabeled top oligonucleotide (0.5 nM) and the unlabeled complimentary bottom oligonucleotide (1 nM) for 10 minutes at 37°C in 10 μl buffer containing 30 mM Tris pH 7.4, 5 mM MgCl2, 1 mM DTT, 100μg/ml BSA, and 10% glycerol. Reactions were stopped and analyzed as described above.
For ATPase assays, RecQ4 (amount indicated in figure legend) was incubated with 8 pmol of a 61 nucleotide ssDNA primer (primer B10, Supplementary Table 1) and 1.25 μCi [γ-32P] ATP (PerkinElmer Life Sciences) in 10 μl helicase assay buffer with 50 μM cold ATP for 1 hour at 37°C. Reactions were stopped by addition of 5 μl of 0.5 M EDTA and placed on ice. 2 μl from each reaction was spotted on a cellulose polyethyleneimine thin-layer chromatography sheet (JT-Baker), and resolved in 0.8 M LiCl/1 M formic acid solution. The chromatography sheet was exposed to a PhosphorImager plate (GE Healthcare) and analyzed as described above.
2.4 Generation of RecQ4 Antibodies
A fragment of the RECQL4 gene corresponding to amino acids 26-176 was PCR-amplified and cloned into pTXB1 plasmid (New England Biolabs). The resulting construct was sequenced and transformed into BL21(DE3) CodonPlus RIPL E. coli strain (Stratagene). Bacteria were grown to OD600=0.7 in LB medium supplemented with 100 μg/ml ampicillin and 50 μg/ml chloramphenicol. Protein expression was induced with 0.2 mM IPTG for 3 hours at 37°C. Cells were collected and protein purification performed on chitin resin (New England Biolabs) according to manufacturer's instructions. Purified N-terminal fragment of RecQL4 was sent to Covance Inc. (Denver, PA) for polyclonal antibody production in rabbits. Antibodies were affinity-purified using AminoLink Plus Immobilization Kit (Thermo Scientific).
2.5 Immunofluorescence
HeLa cells were plated on Lab-Tek II chambered glass slides (Thermo-Fisher Scientific) at a density of 20,000 cells per chamber and grown overnight. Cells were washed with PBS pH 7.4, fixed with 4 % formaldehyde in PBS for 10 minutes, and permeabilized with 0.2 % Triton X-100 in PBS for 5 minutes. Next, cells were incubated with primary antibodies for RecQ4 (1:200) and RPA32 (Santa Cruz sc81372, 1:1000) in PBS with 3 % BSA for 1 hour. Cells were washed five times for 2 minutes with PBS and then incubated with secondary antibodies (goat anti-mouse AlexaFluor 568 and goat anti-rabbit AlexaFluor 647, 1:1000, Invitrogen) in PBS with 3 % BSA for 1 hour. Finally, cells were washed five times for 4 minutes and mounted in Vectashield with DAPI. Images were captured with a Nikon Eclipse TE2000 confocal microscope and analyzed using Volocity-5 software (Perkin Elmer).
3. RESULTS
3.1 Purified RecQ4 possesses helicase activity
A recently published study demonstrated that human RecQ4, like the other members of the RecQ helicase family, is an active helicase [28]. Under normal reaction conditions, though, this helicase activity was masked by a robust strand annealing activity. In the Xu and Liu study, unwinding of a 30 base pair (bp) duplex forked DNA substrate with 30 nucleotide (nt) single-stranded arms occurred only in the presence of excess ssDNA [28]. The ssDNA was used to trap the complementary strand released during the helicase unwinding.
To further explore the helicase activity of human RecQ4, we purified a C-terminal 9-histidine tagged RecQ4 [25]. The protein was judged to be at least 95% pure by SDS-PAGE analysis (Fig. 1A, lane 2). We also generated a long 30 bp duplex forked DNA substrate with 30 nt single-stranded arms, referred to as fork-1, which was utilized to test the helicase activity of our purified RecQ4. Consistent with the published results, we were unable to see unwinding of fork-1 in the absence of excess ssDNA (Fig. 1B, upper panel, lanes 2-6). Yet, upon addition of 25-fold excess ssDNA, we were able to unmask the helicase activity to some extent (Fig. 1B, upper panel, lanes 8-12). This indicates that the RecQ4 possesses helicase activity, in agreement with the previous study [28].
Figure 1. Purified RecQ4 possesses helicase activity.

A) Coomassie blue stained SDS-PAGE gel of purified wild-type (WT) RecQ4 (250 ng ; lane 2). B) The helicase activity of purified wild-type RecQ4 (10, 20, 50, 100, and 200 nM) was assayed on 0.5 nM fork-1 (T1:B1; upper panel) and fork-2 (T2:B2; lower panel) in the absence (lanes 2-6) and presence (lanes 8-12) of unlabeled single-stranded primers (12.5 nM) T1 or T2, respectively. The Δ (lane 13) indicates heat denatured substrate. Asterisk indicates the position of the radiolabel. C) The strand annealing activity of RecQ4 (5, 10, 20, and 50 nM) measured with the radiolabeled ssDNA T1 and T2 and their complimentary primers B1 (lanes 2-5) and B2 (lanes 7-10), respectively. D) Quantitation of RecQ4's strand annealing. The values represent an average of four independent experiments corrected for substrate annealing in the absence of RecQ4. Error bars represent standard deviation.
However, even in the presence of the trap DNA, the RecQ4 helicase activity on fork-1 was relatively weak, only 12% unwinding at the maximum concentration of RecQ4. Addition of higher concentrations of RecQ4 were not successful in significantly increasing the amount of helicase unwinding due to the inhibitory effect of high salt present in the purified RecQ4. The data suggests that RecQ4 is not a robust helicase nor a highly processive helicase. Characterization of RecQ1, BLM, WRN, and RecQ5 revealed that alone they are not effective in unwinding long duplex DNA [36-41]. Therefore, we tested the helicase activity of RecQ4 on a fork substrate with a smaller 22 bp duplex region and 15 nt single-stranded arms. This substrate is a truncated version of fork-1 with the same sequence, and will be referred to as fork-2. Surprisingly, RecQ4 was effective in unwinding fork-2, up to 28% with the highest concentration of RecQ4, even in the absence of excess ssDNA (Fig. 1B, lower panel, lanes 2-6). We saw some additional unwinding, approximately 1.7-fold more, in the presence of 25-fold excess ssDNA (Fig. 1B, lower panel, lanes 8-12). Together, these results indicate that although on the longer duplex (fork-1) RecQ4 helicase activity appears to be masked by strand annealing activity (Fig. 1 A, upper panel and [28]), RecQ4 is functional in unwinding both long and short duplexes. Notably, this is the first observation that human RecQ4 is capable of unwinding short forked duplexes in the complete absence of excess single-stranded DNA.
Nevertheless, on each substrate, fork-1 and fork-2, variation in RecQ4 helicase activity in the absence and the presence of ssDNA suggests differences in the strand annealing of long versus short duplex forks. Because RecQ4 unwound fork-1 less effectively, we hypothesized that RecQ4 strand annealing activity should be more active on that substrate compared to fork-2. To test this, we examined RecQ4 strand annealing activity on both the long and short fork substrates (Fig. 1C). Fork-1 had a higher level of strand annealing in the absence of protein as compared to fork-2 (Fig. 1C, lane 1 versus lane 6). Still, in the presence of increasing amounts of RecQ4, the strand annealing activity appears to be higher on fork-1 as compared to fork-2 (Fig. 1C, lanes 2-5 versus lanes 7-10). Quantitation of the amount of strand annealing, corrected for annealing in the absence of protein, revealed that RecQ4-mediated annealing of fork-1 was notably higher than that of fork-2 (Fig. 1D). Consistently, we also saw a higher binding affinity of RecQ4 for long single-stranded DNA compared to short single-stranded DNA (data not shown). The results suggest that inefficient RecQ4 unwinding of the fork-1 is due to a more effective strand annealing activity with this substrate.
3.2 RecQ4 helicase activity is intrinsic to RecQ4
SDS-PAGE analysis of the RecQ4 used in our study indicates that the protein used in the helicase assays was at least 95% pure (Fig. 1A). It is worth noting that the amount of protein analyzed by SDS-PAGE (Fig. 1A, lane 2) was comparable to the highest amount used in the helicase assay where RecQ4 helicase activity was detected (Fig. 1B, lower panel, lanes 6 and 12). However, there is a possibility that the helicase activity could be attributed to proteins which co-purified with the RecQ4 and were undetectable by our SDS-PAGE analysis. To address this issue, we generated a K508M (KM) point mutation in the full length RecQ4 (Fig. 2A). This conserved lysine, required for ATP hydrolysis, is located within the Walker A motif of the superfamily II (SFII) helicase domain. The purified KM RecQ4 was of equal purity when compared to the wild-type (WT) protein (Fig. 2B, lanes 2 and 3).
Figure 2. Helicase activity is intrinsic to RecQ4.

A) Diagram of the SFII helicase domain of RecQ4. Amino acid residues surrounding the conserved lysine 508 (bold) are noted for the wild-type protein. The point mutation K508M (KM) is indicated in bold italics. B) Coomassie blue stained SDS-PAGE gel of purified K508M mutant (KM; lane 2) and wild-type (WT; lane 3) RecQ4 (250 ng/lane). C) The strand annealing activity of KM RecQ4 (5, 10, 20, and 50 nM; lanes 3-6) versus WT (50 nM; lane 2) measured with the radiolabeled ssDNA T1 and its complimentary primer B1. D) Quantitation of RecQ4 strand annealing. The values represent an average of four independent experiments corrected for substrate annealing in the absence of RecQ4. The WT data is taken from Figure 1D. Error bars represent standard deviation. E) The helicase activity of KM (50, 100, and 200 nM; lanes 2-4) versus WT (200 nM ; lane 5) RecQ4 assayed on fork-2 (T2:B2). The Δ (lane 6) indicates heat denatured substrate. Asterisk indicates the position of the radiolabel. F) ATP hydrolysis by KM (2, 20, and 200nM; lanes 2-4) versus WT RecQ4 (2, 20, and 200nM; lanes 5-7) in the presence of ssDNA co-factor evaluated by thin-layer chromatography.
Moreover, to validate that the protein was properly folded and otherwise functional, we tested the strand annealing activity, which does not require ATP hydrolysis, of the KM versus WT RecQ4. Although some strand annealing of fork-1 occurred in the absence of protein (Fig. 2C, lane 1), the KM RecQ4 was effective in complete strand annealing of this substrate (Fig. 2C, lanes 3-6), as was the WT (Fig. 2C, lane 2). Quantitation of the strand annealing, corrected for annealing in the absence of protein revealed that the strand annealing of KM RecQ4 is equivalent to that of the WT (Fig. 2D).
Subsequently, we tested KM RecQ4 on a fork substrate and failed to detect helicase activity (Fig. 2E, lanes 2-4). However, under the same conditions, the WT RecQ4 was able to unwind 23% of the substrate (Fig. 2E, lane 5). This indicates that the helicase activity seen in the RecQ4 preparation is not likely caused by co-purification of unknown proteins, and is instead intrinsic to RecQ4. In addition, the KM mutant was unable to hydrolyze ATP (Fig. 2F, lanes 2-4 compared to 5-7) confirming that the conserved lysine in the Walker A motif is necessary for ATP hydrolysis and helicase activity in RecQ4, as it is in the other human RecQ helicases and in Drosophila RecQ4 [36, 40, 42-44]. Our results indicate that the unwinding activity of RecQ4 is confined to the conserved helicase domain. They also are in contrast to the previous report by Xu and Liu, which suggested that the conserved helicase domain is not solely responsible for ATP-dependent helicase activity of RecQ4 [28]. The authors proposed that the N-terminus of RecQ4 also posseses ATP-dependent unwinding activity, but they failed to demonstrate its ability to hydrolyze ATP.
3.3 Helicase activity is ATPase-dependent and sequence-independent
To verify that the RecQ4 unwinding is in fact driven by ATPase activity, we compared the helicase activity of RecQ4 in the absence of ATP, as well as, in the presence of ATP or the non-hydrolyzable analog ATPγS Consistent with ATP-dependent helicase activity, we were only able to see unwinding of fork-2 in the presence of ATP (Fig. 3, upper panel, lanes 2-3), but not in the presence of ATPγS or in the absence of ATP (Fig. 3, upper panel, lanes 4-5 and 6-7, respectively).
Figure 3. RecQ4 helicase activity is ATPase-dependent and sequence-independent.
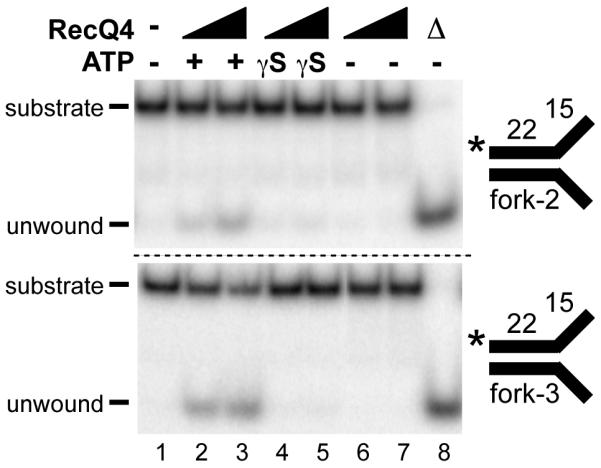
The helicase activity of RecQ4 (100 and 200 nM) was measured on fork-1 (T2:B2; upper panel) and fork-3 (T3:B3; lower panel) in the presence of 5 mM ATP (lanes 2-3), 5 mM non-hydrolyzable ATPγS (lanes 4-5), or absence of ATP (lanes 6-7). The Δ (lane 8) indicates heat denatured substrate. Asterisk indicates the position of the radiolabel.
Additionally, we were interested in whether the ATPase-dependent helicase activity of RecQ4 was restricted to a particular substrate sequence, since unwinding was seen on fork substrates composed of the same basic sequence. To investigate this, we tested RecQ4 unwinding of a short duplex fork substrate of an alternate sequence. Like fork-2, fork-3 substrate (22 bp duplex region with 15 nt arms) was unwound by RecQ4 in the presence of ATP (Fig. 3, lower panel, lanes 2-3). We were unable to see unwinding of fork-3 in the presence of ATPγS or in the absence of ATP (Fig. 3, lower panel, lanes 4-5 and lanes 6-7). Both substrates were unwound by RecQ4 only in the presence of ATP, indicating that the ATPase-dependent helicase activity of RecQ4 is likely to be sequence-independent.
We further tested RecQ4 helicase activity on a series of substrates with varying structures, sizes, and sequence contexts (Table 2). RecQ4 was effective in resolving fork substrates containing a 22 bp duplex with either 15 or 6 nt single-stranded arms with mixed, or with telomeric duplex DNA sequence. Under the same conditions, fork substrates with duplex regions greater than 26 bp were not unwound by RecQ4 in the absence of excess ssDNA, regardless of the sequence and length of the single-stranded arms. Unwinding of the long duplex forks (greater than 26 bp), with the exception of the telomeric fork, was observed upon addition of ssDNA. Perhaps the stable base-pairing of G-rich sequence in the long telomeric duplex was more difficult for RecQ4 to resolve even in the presence of ssDNA. Additionally, RecQ4 was unable to unwind a full 61 bp duplex, a Holliday junction, or G-quadruplex structures. It is worth noting that these alternate structures were only tested with one sequence. Although, unwinding of fork substrates is likely sequence-independent, we cannot rule out the effect of sequence context on RecQ4 resolution of more complex DNA structures.
Table 2. RecQ4 helicase activity on multiple substrates in the absence and presence of excess single-stranded DNA (ssDNA).
Substrate composition is detailed in Supplementary Table 1. Helicase activity on each substrate was measured as described in the Materials and Methods. Forks are named by the lengths of the duplex region (a) and of the single-stranded arms (b). Forks listed above the dotted line all contain the same basic sequence as fork-1 (described in Figure 1) but are truncated in the length of the duplex and/or arms. Forks listed below the dotted line are all composed of sequences varying from fork-1 and varying from each other. Each of the alternate conformation substrates was tested with one sequence. (NA: not applicable)
| Structure | Substrate | without ssDNA |
with ssDNA |
|---|---|---|---|
| Forks | (a/b) | ||
|
30/30 (fork-1) | − | + |
| 30/15 (fork-4) | − | + | |
| 22/15 (fork-2) | + | + | |
| 22/6 | + | + | |
|
22/15 (fork-3) | + | + |
| 26/23 | − | + | |
| 34/15 | − | + | |
| 22/15, telomeric | + | + | |
| 34/15, telomeric | − | − | |
| Alternate Conformations | |||
|
|
Full Duplex, 61 base pairs | − | − |

|
Holliday Junction | − | NA |

|
G-quadruplex | − | NA |
3.4 Replication protein A (RPA) stimulates RecQ4 helicase activity
Our results thus far demonstrate that purified human RecQ4 possesses an ATP-dependent helicase activity, similar to those of the previously characterized human RecQ helicases. RecQ1, BLM, WRN, and RecQ5 helicase activities are stimulated by the ssDNA binding protein RPA [36-41]. Based on the similarities in biochemical activities and interacting protein partners [2, 4], we predicted that, like other members of the RecQ family, RecQ4's helicase activity would be stimulated by RPA. Our results indicate that addition of increasing amounts of RPA stimulates RecQ4 unwinding of both fork-2 and fork-3 (Fig. 4A, upper and lower panels, respectively, lane 2 compared to 3-6). Since RPA had been previously shown to destabilize DNA duplexes [45-48], stimulation of RecQ4 was tested at concentrations of RPA that generate very minimal amounts of single-stranded radiolabeled DNA (Fig. 4A). Consequently, we were unable to test higher concentrations of RPA due to destabilization of the fork substrates. Nevertheless, quantitation of the unwinding, corrected for RPA substrate destabilization, revealed that RPA stimulated RecQ4 helicase activity approximately twofold on fork-2 and fork-3 (Fig. 4B). In an effort to see a greater amount of RecQ4 helicase stimulation by RPA, we adjusted the helicase assay reaction conditions by changing the amount of KCl in the buffer. However, we were unable to stimulate unwinding more than two-fold on fork-2 and fork-3 (data not shown). Additionally, we tested RecQ4 helicase activity on the longer 30 bp duplex fork substrate, which displayed less RPA destabilization than the short duplex fork. RecQ4 was unable to unwind the longer 30 bp duplex fork even in presence of RPA (Table 2 and data not shown). To determine whether the mild RPA stimulation of RecQ4 fork unwinding was in keeping with that of other RecQ helicases, we compared it to the RPA stimulation of WRN helicase activity. WRN was highly efficient in unwinding the short 22 bp duplex forks. Thus, at a concentration of WRN low enough to detect stimulation, we were unable to further increase WRN's unwinding with addition of RPA and saw instead a slight inhibition (Fig. 4C, upper panel, lanes 2-5 and Fig. 4D). Yet, on a 30 bp duplex fork substrate, which was unwound less efficiently by WRN alone, RPA was able to stimulate helicase activity approximately four-fold (Fig. 4C, lower panel, lane 2 compared to 5, and Fig. 4E). Our results indicate that RPA stimulation of unwinding is optimally observed on short duplex fork for RecQ4 and a long duplex fork for WRN. This difference is likely due to the activity and processivity of each respective helicase. Nevertheless, although the RPA stimulation of RecQ4 helicase activity was mild, it is in the same range as the stimulation of WRN helicase on an optimal forked duplex substrate.
Figure 4. RPA stimulates RecQ4 helicase activity.

A) The helicase activity of RecQ4 (34 nM ; lane 2) was measured on fork-2 (T2:B2; upper panel) and fork-3 (T3:B3; lower panel) in the presence of increasing concentrations of RPA (0.5, 1, 2, and 5 nM; lanes 3-6). The Δ (lane 11) indicates heat denatured substrate. Asterisk indicates the position of the radiolabel. B) Quantitation of RecQ4 unwinding in the presence of RPA. The values represent an average of four independent experiments corrected for destabilization of the substrate by RPA in the absence of RecQ4. Error bars represent standard deviation. C) The helicase activity of WRN (0.125 nM and 0.5 nM; lane 2 upper and lower panels, respectively) was measured on fork-2 (T2:B2; upper panel) and fork-4 (T4:B4; lower panel) in the presence of increasing concentrations of RPA (1, 2, and 5 nM, lanes 3-6 upper panel; 5, 10, and 20 nM lanes 3-6 lower panel). The Δ (lane 11) indicates heat denatured substrate. Asterisk indicates the position of the radiolabel. D and E) Quantitation of WRN unwinding of fork-2 and fork-4, respectively, in the presence of RPA. The values represent an average of three independent experiments corrected for destabilization of the substrate by RPA in the absence of WRN. Error bars represent standard deviation.
3.5 RecQ4 and RPA co-localize within the nucleus
To determine a potential biological association between RecQ4 and RPA, we used immunofluorescent staining to look for cellular co-localization of these two proteins. In HeLa cells, both RecQ4 and RPA individually demonstrated nuclear localization (Supplementary Fig. 1, upper panels). When examining the merged image (Supplementary Fig. 1, lower left panel), there are some areas of co-localization within the nucleus, as indicated by the yellow areas of the image. A Pearson's correlation of 0.819 was calculated using the default settings in the co-localization module of Volocity-5 software for image analysis. This co-localization is suggestive of a biologically relevant interaction between RecQ4 and RPA. Currently, we are conducting additional experiments to further examine the details and the biological consequences of this interaction. Additionally, we are pursuing the significance of cytoplasmic localization of RecQ4 (Supplementary Fig. 1).
4. DISCUSSION
Although RecQ4 strand annealing and ATPase activities have been shown, helicase activity was previously undetectable [25, 26]. Yet, using the same construct and purification of RecQ4 from Macris et al. [25] we were able to detect DNA unwinding by purified human RecQ4, through studies in vitro, consistent with recent results [27, 28]. Xu and Liu suggested that robust strand annealing activity by RecQ4 prevented visualization of helicase activity [28]. As RecQ4 unwinds duplex DNA and releases a strand, its strand annealing activity reforms the duplex. Thus, the helicase activity must be unmasked by addition of ssDNA which traps the released strand. In their study, Xu and Liu could only detect helicase activity in the presence of excess ssDNA [28]. We also observed RecQ4 unwinding only in the presence of the single-stranded trap DNA on a fork substrate containing 30 bp duplex (Fig. 1). Notably, however, on a fork substrate with a short 22 bp duplex region, RecQ4 unwinding was seen, regardless of sequence context, in both the absence and presence of the ssDNA (Figs. 1 and 3 and Table 2). Our novel observation of RecQ4's ability to unwind short duplexes in the absence of excess ssDNA could be attributed to the low processivity of RecQ4. A screen of helicase activity on varying sized fork substrates further demonstrated that fork unwinding by RecQ4 was likely related to the short length of the duplex and not the length of the single-stranded arms (Table 2). Thus, our preparation of RecQ4 appears to be effective in unwinding short regions of duplex DNA without excess of ssDNA, and is therefore, like the other RecQ helicase family members, not highly processive [4]. Effective unwinding of only short duplexes in the absence of ssDNA also provides explanation of visible RecQ4 DNA unwinding with a protein construct formerly lacking helicase activity. The substrates used in those early experiments contained duplex regions 40 bp in length, which were tested in the absence of excess ssDNA [25]. Our results show that in the absence of excess ssDNA RecQ4 was only able to unwind duplexes 22 bp in length. Helicase activity on duplexes greater than 26 bp was only seen with the ssDNA trap. Therefore, the 40 bp duplexes used by Macris et al. were most likely too large to be efficiently unwound by RecQ4 [25]. In adddition, it is possible that our buffer conditions are more optimal for RecQ4 than those used previously.
Alternatively, as mentioned above, differences in the RecQ4 strand annealing could explain the variation in unwinding of the two substrates. Unlike on the long duplex fork, the strand annealing did not appear to mask RecQ4 helicase activity on the short duplex fork. We hypothesized that strand annealing activity was more active on the long substrate, obscuring the helicase activity, and less active on the short substrate, allowing for the helicase activity to be revealed. Consistent with our hypothesis, RecQ4 strand annealing was in fact higher for the long duplex fork (Fig. 1). Together, these results suggest a competition between RecQ4's strand annealing and helicase activities. Unlike the proposal set forth by Xu and Liu where RecQ4's strand annealing overshadows its helicase activity [28], our in vitro studies suggest that there must be an equilibrium between strand annealing and helicase activities that is dependent on the substrate length. Although the conditions of this equilibrium require further investigation, it is possible that the substrate preference modulates RecQ4's biological roles. For example, RecQ4 may unwind short regions of duplex DNA that can form as secondary structure in flaps generated during strand displacement of Okazaki fragments. These foldback structures are inhibitory to FEN1 cleavage and subsequent ligation to complete Okazaki fragment maturation [49, 50]. It is conceivable that RecQ4 may resolve the secondary structure allowing FEN1 cleavage and progression of lagging strand synthesis, similar to what has been proposed for BLM [51]. In this way, RecQ4 unwinding of short duplexes may have a role in DNA replication. Conversely, RecQ4 strand annealing of longer lengths of DNA may function during DNA repair. One possibility is that RecQ4 strand annealing mediates the strand pairing during homologous recombination [52]. Still, as it has been observed with WRN, BLM, and RecQ5, the strand annealing and helicase activities may be coordinated to perform strand exchange reactions facilitating regression of stalled replication forks via branch migration or disruption of illegitimate recombination intermediates [53-55].
Nevertheless, the helicase activity of RecQ4 is separable from the strand annealing activity. We generated RecQ4 with a K508M point mutation of the conserved lysine in the Walker A motif of the SFII helicase domain, which is important for ATP hydrolysis, and compared its activity to the WT RecQ4. As expected, the K508M mutation eliminated RecQ4 helicase and ATPase activity (Fig. 2), consistent with the notion that helicase activity is ATPase-dependent. A recent analysis of purified RecQ4 also showed ATPase-dependent unwinding of duplex DNA [27]. However, it was not clear in that study whether the helicase activity was due to co-purification of other proteins and not simply due to the RecQ4. Significantly, with the K508M RecQ4 we demonstrated that the ATPase-dependent helicase activity seen with the purified protein is intrinsic to human RecQ4 (Figs. 2 and 3). Additionally, the K508M RecQ4 retained its strand annealing equivalent to WT (Fig. 2), providing the first indication that human RecQ4 strand annealing and helicase activities are independent of one another. More importantly, absence of helicase activity in the K508M RecQ4 indicates that the conserved lysine of SFII helicase domain is essential for RecQ4 unwinding activity. In contrast, the Xu and Liu report suggests the N-terminus of RecQ4, which has weak homology to yeast replication initiation factor Sld2, possesses ATP-dependent DNA unwinding activity independent of the SFII domain [28]. Although, the authors fail to demonstrate that the N-terminal domain is actually able to hydrolyze ATP. It is possible that unwinding seen with the N-terminal domain of RecQ4 is not true ATP-dependent unwinding, but it is rather due to protein binding-induced destabilization of the duplex DNA. On the other hand, we demonstrate that the ATPase-dependent DNA unwinding by human RecQ4, which requires the lysine in the Walker A motif of the SFII helicase domain, is conserved among the RecQ family members [36, 40, 42, 43]. Our results are consistent with recent work demonstrating that the conserved lysine is also essential for helicase activity of Drosophila RecQ4 [44].
Based on the fact that the strand annealing and ATPase-dependent helicase activities are shared between the RecQ helicases, we were interested in whether some of their interacting protein partners were conserved as well. RecQ1, BLM, WRN, and RecQ5, are stimulated by RPA [36-41]. Accordingly, we tested the stimulation of RecQ4 helicase activity by RPA (Fig. 4). In our assays, we saw an approximate two-fold stimulation of RecQ4 unwinding. The stimulation occurred on two different substrates, indicating that RecQ4 helicase activity, as well as the RPA stimulation, is sequence independent. Additionally, it is worth noting that we were unable to see significant RPA stimulation of unwinding of a short duplex fork when we tested another RecQ helicase, WRN (Fig. 4). This is consistent with previous work from our laboratory showing that WRN and BLM are highly efficient in unwinding short duplexes. RPA stimulation is most useful for unwinding of long duplex substrates, which are not readily processed by WRN and BLM helicases [36, 37]. RecQ4 appears to be generally less active as a helicase, allowing for some amount of stimulation by RPA. Although the stimulation was mild, these results are the first evidence of a conserved RPA interaction among all the RecQ helicases. Additionally, we detected co-localization of RecQ4 and RPA within the nucleus, suggestive of biologically relevant association between RecQ4 and RPA (Supplementary Fig. 1). Since RPA is involved in multiple DNA replication and repair processes [56], the interaction with RPA may facilitate RecQ4 function in a variety of DNA transactions. Functional interaction of RecQ4 with RPA also suggests that other common protein partners may be shared among the RecQ helicases, which merits additional investigation.
Previous biological and cellular characterization of RecQ4 revealed roles in DNA replication [17, 18], double strand break repair [19-21], NER [22], and BER [20, 23, 24]. These roles, similar to the other RecQ helicases, are likely accomplished through the conserved strand annealing and ATPase-dependent helicase activities. Since humans possess five distinct RecQ family members, the apparent redundancy of function raises the question under which conditions do each of the RecQ helicases work. Presumably, each helicase has different substrate specificity. BLM and WRN, for example, are effective in unwinding G-quadruplex DNA, while RecQ1 is not [35, 57-59]. Conversely, while RecQ1 can mediate resolution of immobile Holliday junction structures, BLM is only capable of doing so in the presence of RPA [59]. In addition, although the RecQ helicases share some protein-protein interactions, including RPA [36-41] and FEN1 [24, 60, 61], unique and as of yet undiscovered protein partners may modulate functions of the RecQ helicases in alternate repair pathways. Furthermore, post-translational modifications may affect differential functions of the RecQ helicases. For instance, WRN helicase and exonuclease activities are inhibited by phosphorylation [62-64], yet stimulated by acetylation [65]. Also, recent results indicate that RecQ4 is acetylated by p300, which affects its localization within the cell [66]. The current study provides novel advances in the understanding of RecQ4's biochemical activities indicating that intrinsic ATPase-dependent helicase activity of human RecQ4 is solely due to its conserved helicase domain and not to the N-terminal Sld2-like domain, as suggested previously [28]. To date, RecQ4 remains one of the least well characterized RecQ helicases [67], and further examination of substrate specificity, protein-protein interactions, and post-translational modifications will be necessary to elucidate its unique biological and cellular roles.
Supplementary Material
ACKNOWLEDGEMENTS
We are particularly grateful to Dr. Patrick Sung for providing the WT RecQ4 plasmid construct and Dr. Marc S. Wold for providing the RPA plasmid construct. Also, we would like to acknowledge the members of the Laboratory of Molecular Gerontology at the National Institutes of Health, National Institute on Aging for helpful discussions and technical support, especially Dr. Dharmendra K. Singh, Dr. Takashi Tadokoro, Lale Dawut, and Guido Keijzers for assistance with the protein purification and Alfred May for assistance with the microscopy. Additionally, we thank Drs. Suhasini Avvaru and Chandrika Canugovi for critical reading of this manuscript.
FUNDING
This work was supported entirely by funds from the Intramural Research Program of the National Institutes of Health, National Institute on Aging.
The abbreviations used are
- APE1
apurinic endonuclease 1
- BER
base excision repair
- BLM
Bloom syndrome protein
- bp
base pair
- FEN1
flap endonuclease 1
- NER
nucleotide excision repair
- nt
nucleotide
- RPA
replication protein A
- RTS
Rothmund-Thomson syndrome
- SFII
superfamily II
- ssDNA
single-stranded DNA
- WT
wild-type
- WRN
Werner syndrome protein
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
REFERENCES
- 1.Bachrati CZ, Hickson ID. RecQ helicases: guardian angels of the DNA replication fork. Chromosoma. 2008;117:219–233. doi: 10.1007/s00412-007-0142-4. [DOI] [PubMed] [Google Scholar]
- 2.Brosh RM, Jr., Bohr VA. Human premature aging, DNA repair and RecQ helicases. Nucleic Acids Res. 2007;35:7527–7544. doi: 10.1093/nar/gkm1008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hanada K, Hickson ID. Molecular genetics of RecQ helicase disorders. Cell Mol. Life Sci. 2007;64:2306–2322. doi: 10.1007/s00018-007-7121-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sharma S, Doherty KM, Brosh RM., Jr. Mechanisms of RecQ helicases in pathways of DNA metabolism and maintenance of genomic stability. Biochem. J. 2006;398:319–337. doi: 10.1042/BJ20060450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Singh DK, Ahn B, Bohr VA. Roles of RECQ helicases in recombination based DNA repair, genomic stability and aging. Biogerontology. 2009;10:235–252. doi: 10.1007/s10522-008-9205-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Stinco G, Governatori G, Mattighello P, Patrone P. Multiple cutaneous neoplasms in a patient with Rothmund-Thomson syndrome: case report and published work review. J. Dermatol. 2008;35:154–161. doi: 10.1111/j.1346-8138.2008.00436.x. [DOI] [PubMed] [Google Scholar]
- 7.Vennos EM, Collins M, James WD. Rothmund-Thomson syndrome: review of the world literature. J. Am. Acad. Dermatol. 1992;27:750–762. doi: 10.1016/0190-9622(92)70249-f. [DOI] [PubMed] [Google Scholar]
- 8.Anbari KK, Ierardi-Curto LA, Silber JS, Asada N, Spinner N, Zackai EH, Belasco J, Morrissette JD, Dormans JP. Two primary osteosarcomas in a patient with Rothmund-Thomson syndrome. Clin. Orthop. Relat Res. 2000:213–223. doi: 10.1097/00003086-200009000-00032. [DOI] [PubMed] [Google Scholar]
- 9.Der, V K, McGill JJ, Vekemans M, Kopelman HR. Clonal lines of aneuploid cells in Rothmund-Thomson syndrome. Am. J. Med. Genet. 1990;37:336–339. doi: 10.1002/ajmg.1320370308. [DOI] [PubMed] [Google Scholar]
- 10.Durand F, Castorina P, Morant C, Delobel B, Barouk E, Modiano P. Rothmund-Thomson syndrome, trisomy 8 mosaicism and RECQ4 gene mutation. Ann. Dermatol. Venereol. 2002;129:892–895. [PubMed] [Google Scholar]
- 11.Orstavik KH, McFadden N, Hagelsteen J, Ormerod E, van der Hagen CB. Instability of lymphocyte chromosomes in a girl with Rothmund-Thomson syndrome. J. Med. Genet. 1994;31:570–572. doi: 10.1136/jmg.31.7.570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jin W, Liu H, Zhang Y, Otta SK, Plon SE, Wang LL. Sensitivity of RECQL4-deficient fibroblasts from Rothmund-Thomson syndrome patients to genotoxic agents. Hum. Genet. 2008;123:643–653. doi: 10.1007/s00439-008-0518-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Werner SR, Prahalad AK, Yang J, Hock JM. RECQL4-deficient cells are hypersensitive to oxidative stress/damage: Insights for osteosarcoma prevalence and heterogeneity in Rothmund-Thomson syndrome. Biochem. Biophys. Res. Commun. 2006;345:403–409. doi: 10.1016/j.bbrc.2006.04.093. [DOI] [PubMed] [Google Scholar]
- 14.Balraj P, Concannon P, Jamal R, Beghini A, Hoe TS, Khoo AS, Volpi L. An unusual mutation in RECQ4 gene leading to Rothmund-Thomson syndrome. Mutat. Res. 2002;508:99–105. doi: 10.1016/s0027-5107(02)00189-6. [DOI] [PubMed] [Google Scholar]
- 15.Kitao S, Shimamoto A, Goto M, Miller RW, Smithson WA, Lindor NM, Furuichi Y. Mutations in RECQL4 cause a subset of cases of Rothmund-Thomson syndrome. Nat. Genet. 1999;22:82–84. doi: 10.1038/8788. [DOI] [PubMed] [Google Scholar]
- 16.Lindor NM, Furuichi Y, Kitao S, Shimamoto A, Arndt C, Jalal S. Rothmund-Thomson syndrome due to RECQ4 helicase mutations: report and clinical and molecular comparisons with Bloom syndrome and Werner syndrome. Am. J. Med. Genet. 2000;90:223–228. doi: 10.1002/(sici)1096-8628(20000131)90:3<223::aid-ajmg7>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- 17.Sangrithi MN, Bernal JA, Madine M, Philpott A, Lee J, Dunphy WG, Venkitaraman AR. Initiation of DNA replication requires the RECQL4 protein mutated in Rothmund-Thomson syndrome. Cell. 2005;121:887–898. doi: 10.1016/j.cell.2005.05.015. [DOI] [PubMed] [Google Scholar]
- 18.Matsuno K, Kumano M, Kubota Y, Hashimoto Y, Takisawa H. The N-terminal noncatalytic region of Xenopus RecQ4 is required for chromatin binding of DNA polymerase alpha in the initiation of DNA replication. Mol. Cell Biol. 2006;26:4843–4852. doi: 10.1128/MCB.02267-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kumata Y, Tada S, Yamanada Y, Tsuyama T, Kobayashi T, Dong YP, Ikegami K, Murofushi H, Seki M, Enomoto T. Possible involvement of RecQL4 in the repair of double-strand DNA breaks in Xenopus egg extracts. Biochim. Biophys. Acta. 2007;1773:556–564. doi: 10.1016/j.bbamcr.2007.01.005. [DOI] [PubMed] [Google Scholar]
- 20.Xu Y, Lei Z, Huang H, Dui W, Liang X, Ma J, Jiao R. dRecQ4 is required for DNA synthesis and essential for cell proliferation in Drosophila. PLoS. One. 2009;4:e6107. doi: 10.1371/journal.pone.0006107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Petkovic M, Dietschy T, Freire R, Jiao R, Stagljar I. The human Rothmund-Thomson syndrome gene product, RECQL4, localizes to distinct nuclear foci that coincide with proteins involved in the maintenance of genome stability. J. Cell Sci. 2005;118:4261–4269. doi: 10.1242/jcs.02556. [DOI] [PubMed] [Google Scholar]
- 22.Fan W, Luo J. RecQ4 facilitates UV light-induced DNA damage repair through interaction with nucleotide excision repair factor xeroderma pigmentosum group A (XPA) J. Biol. Chem. 2008;283:29037–29044. doi: 10.1074/jbc.M801928200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Woo LL, Futami K, Shimamoto A, Furuichi Y, Frank KM. The Rothmund-Thomson gene product RECQL4 localizes to the nucleolus in response to oxidative stress. Exp. Cell Res. 2006;312:3443–3457. doi: 10.1016/j.yexcr.2006.07.023. [DOI] [PubMed] [Google Scholar]
- 24.Schurman SH, Hedayati M, Wang Z, Singh DK, Speina E, Zhang Y, Becker K, Macris M, Sung P, Wilson DM, III, Croteau DL, Bohr VA. Direct and indirect roles of RECQL4 in modulating base excision repair capacity. Hum. Mol. Genet. 2009;18:3470–3483. doi: 10.1093/hmg/ddp291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Macris MA, Krejci L, Bussen W, Shimamoto A, Sung P. Biochemical characterization of the RECQ4 protein, mutated in Rothmund-Thomson syndrome. DNA Repair (Amst) 2006;5:172–180. doi: 10.1016/j.dnarep.2005.09.005. [DOI] [PubMed] [Google Scholar]
- 26.Yin J, Kwon YT, Varshavsky A, Wang W. RECQL4, mutated in the Rothmund-Thomson and RAPADILINO syndromes, interacts with ubiquitin ligases UBR1 and UBR2 of the N-end rule pathway. Hum. Mol. Genet. 2004;13:2421–2430. doi: 10.1093/hmg/ddh269. [DOI] [PubMed] [Google Scholar]
- 27.Suzuki T, Kohno T, Ishimi Y. DNA helicase activity in purified human RECQL4 protein. J. Biochem. 2009;146:327–335. doi: 10.1093/jb/mvp074. [DOI] [PubMed] [Google Scholar]
- 28.Xu X, Liu Y. Dual DNA unwinding activities of the Rothmund-Thomson syndrome protein. RECQ4, EMBO J. 2009;28:568–577. doi: 10.1038/emboj.2009.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Henricksen LA, Umbricht CB, Wold MS. Recombinant replication protein A: expression, complex formation, and functional characterization. J. Biol. Chem. 1994;269:11121–11132. [PubMed] [Google Scholar]
- 30.Araki M, Masutani C, Takemura M, Uchida A, Sugasawa K, Kondoh J, Ohkuma Y, Hanaoka F. Centrosome protein centrin 2/caltractin 1 is part of the xeroderma pigmentosum group C complex that initiates global genome nucleotide excision repair. J. Biol. Chem. 2001;276:18665–18672. doi: 10.1074/jbc.M100855200. [DOI] [PubMed] [Google Scholar]
- 31.Orren DK, Brosh RM, Jr., Nehlin JO, Machwe A, Gray MD, Bohr VA. Enzymatic and DNA binding properties of purified WRN protein: high affinity binding to single-stranded DNA but not to DNA damage induced by 4NQO. Nucleic Acids Res. 1999;27:3557–3566. doi: 10.1093/nar/27.17.3557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lloyd RG, Sharples GJ. Processing of recombination intermediates by the RecG and RuvAB proteins of Escherichia coli. Nucleic Acids Res. 1993;21:1719–1725. doi: 10.1093/nar/21.8.1719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Karow JK, Constantinou A, Li JL, West SC, Hickson ID. The Bloom's syndrome gene product promotes branch migration of holliday junctions. Proc. Natl. Acad. Sci. U. S. A. 2000;97:6504–6508. doi: 10.1073/pnas.100448097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Huber MD, Lee DC, Maizels N. G4 DNA unwinding by BLM and Sgs1p: substrate specificity and substrate-specific inhibition. Nucleic Acids Res. 2002;30:3954–3961. doi: 10.1093/nar/gkf530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sun H, Karow JK, Hickson ID, Maizels N. The Bloom's syndrome helicase unwinds G4 DNA. J. Biol. Chem. 1998;273:27587–27592. doi: 10.1074/jbc.273.42.27587. [DOI] [PubMed] [Google Scholar]
- 36.Brosh RM, Jr., Orren DK, Nehlin JO, Ravn PH, Kenny MK, Machwe A, Bohr VA. Functional and physical interaction between WRN helicase and human replication protein A. J. Biol. Chem. 1999;274:18341–18350. doi: 10.1074/jbc.274.26.18341. [DOI] [PubMed] [Google Scholar]
- 37.Brosh RM, Jr., Li JL, Kenny MK, Karow JK, Cooper MP, Kureekattil RP, Hickson ID, Bohr VA. Replication protein A physically interacts with the Bloom's syndrome protein and stimulates its helicase activity. J. Biol. Chem. 2000;275:23500–23508. doi: 10.1074/jbc.M001557200. [DOI] [PubMed] [Google Scholar]
- 38.Cui S, Arosio D, Doherty KM, Brosh RM, Jr., Falaschi A, Vindigni A. Analysis of the unwinding activity of the dimeric RECQ1 helicase in the presence of human replication protein A. Nucleic Acids Res. 2004;32:2158–2170. doi: 10.1093/nar/gkh540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Doherty KM, Sommers JA, Gray MD, Lee JW, von KC, Thoma NH, Kureekattil RP, Kenny MK, Brosh RM., Jr. Physical and functional mapping of the replication protein a interaction domain of the werner and bloom syndrome helicases. J. Biol. Chem. 2005;280:29494–29505. doi: 10.1074/jbc.M500653200. [DOI] [PubMed] [Google Scholar]
- 40.Garcia PL, Liu Y, Jiricny J, West SC, Janscak P. Human RECQ5beta, a protein with DNA helicase and strand-annealing activities in a single polypeptide. EMBO J. 2004;23:2882–2891. doi: 10.1038/sj.emboj.7600301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Shen JC, Gray MD, Oshima J, Loeb LA. Characterization of Werner syndrome protein DNA helicase activity: directionality, substrate dependence and stimulation by replication protein A. Nucleic Acids Res. 1998;26:2879–2885. doi: 10.1093/nar/26.12.2879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Neff NF, Ellis NA, Ye TZ, Noonan J, Huang K, Sanz M, Proytcheva M. The DNA helicase activity of BLM is necessary for the correction of the genomic instability of bloom syndrome cells. Mol. Biol. Cell. 1999;10:665–676. doi: 10.1091/mbc.10.3.665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sharma S, Sommers JA, Choudhary S, Faulkner JK, Cui S, Andreoli L, Muzzolini L, Vindigni A, Brosh RM., Jr. Biochemical analysis of the DNA unwinding and strand annealing activities catalyzed by human RECQ1. J. Biol. Chem. 2005;280:28072–28084. doi: 10.1074/jbc.M500264200. [DOI] [PubMed] [Google Scholar]
- 44.Capp C, Wu J, Hsieh TS. Drosophila RecQ4 has a 3′-5′ DNA helicase activity that is essential for viability. J. Biol. Chem. 2009;284:30845–30852. doi: 10.1074/jbc.M109.008052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Georgaki A, Strack B, Podust V, Hubscher U. DNA unwinding activity of replication protein A. FEBS Lett. 1992;308:240–244. doi: 10.1016/0014-5793(92)81283-r. [DOI] [PubMed] [Google Scholar]
- 46.Georgaki A, Hubscher U. DNA unwinding by replication protein A is a property of the 70 kDa subunit and is facilitated by phosphorylation of the 32 kDa subunit. Nucleic Acids Res. 1993;21:3659–3665. doi: 10.1093/nar/21.16.3659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Lao Y, Lee CG, Wold MS. Replication protein A interactions with DNA. 2. Characterization of double-stranded DNA-binding/helix-destabilization activities and the role of the zinc-finger domain in DNA interactions. Biochemistry. 1999;38:3974–3984. doi: 10.1021/bi982371m. [DOI] [PubMed] [Google Scholar]
- 48.Treuner K, Ramsperger U, Knippers R. Replication protein A induces the unwinding of long double-stranded DNA regions. J. Mol. Biol. 1996;259:104–112. doi: 10.1006/jmbi.1996.0305. [DOI] [PubMed] [Google Scholar]
- 49.Henricksen LA, Tom S, Liu Y, Bambara RA. Inhibition of flap endonuclease 1 by flap secondary structure and relevance to repeat sequence expansion. J. Biol. Chem. 2000;275:16420–16427. doi: 10.1074/jbc.M909635199. [DOI] [PubMed] [Google Scholar]
- 50.Kao HI, Veeraraghavan J, Polaczek P, Campbell JL, Bambara RA. On the roles of Saccharomyces cerevisiae Dna2p and Flap endonuclease 1 in Okazaki fragment processing. J. Biol. Chem. 2004;279:15014–15024. doi: 10.1074/jbc.M313216200. [DOI] [PubMed] [Google Scholar]
- 51.Wang W, Bambara RA. Human Bloom protein stimulates flap endonuclease 1 activity by resolving DNA secondary structure. J. Biol. Chem. 2005;280:5391–5399. doi: 10.1074/jbc.M412359200. [DOI] [PubMed] [Google Scholar]
- 52.Ouyang KJ, Woo LL, Ellis NA. Homologous recombination and maintenance of genome integrity: cancer and aging through the prism of human RecQ helicases. Mech. Ageing Dev. 2008;129:425–440. doi: 10.1016/j.mad.2008.03.003. [DOI] [PubMed] [Google Scholar]
- 53.Bartos JD, Wang W, E.Pike J, Bambara RA. Mechanisms by which Bloom protein can disrupt recombination intermediates of Okazaki fragment maturation. J. Biol. Chem. 2006;281:32227–32239. doi: 10.1074/jbc.M606310200. [DOI] [PubMed] [Google Scholar]
- 54.Kanagaraj R, Saydam N, Garcia PL, Zheng L, Janscak P. Human RECQ5beta helicase promotes strand exchange on synthetic DNA structures resembling a stalled replication fork. Nucleic Acids Res. 2006;34:5217–5231. doi: 10.1093/nar/gkl677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Machwe A, Xiao L, Groden J, Matson SW, Orren DK. RecQ family members combine strand pairing and unwinding activities to catalyze strand exchange. J. Biol. Chem. 2005;280:23397–23407. doi: 10.1074/jbc.M414130200. [DOI] [PubMed] [Google Scholar]
- 56.Wold MS. Replication protein A: a heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu. Rev. Biochem. 1997;66:61–92. doi: 10.1146/annurev.biochem.66.1.61. [DOI] [PubMed] [Google Scholar]
- 57.Fry M, Loeb LA. Human werner syndrome DNA helicase unwinds tetrahelical structures of the fragile X syndrome repeat sequence d(CGG)n. J. Biol. Chem. 1999;274:12797–12802. doi: 10.1074/jbc.274.18.12797. [DOI] [PubMed] [Google Scholar]
- 58.Mohaghegh P, Karow JK, Brosh JR, Jr., Bohr VA, Hickson ID. The Bloom's and Werner's syndrome proteins are DNA structure-specific helicases. Nucleic Acids Res. 2001;29:2843–2849. doi: 10.1093/nar/29.13.2843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Popuri V, Bachrati CZ, Muzzolini L, Mosedale G, Costantini S, Giacomini E, Hickson ID, Vindigni A. The Human RecQ helicases, BLM and RECQ1, display distinct DNA substrate specificities. J. Biol. Chem. 2008;283:17766–17776. doi: 10.1074/jbc.M709749200. [DOI] [PubMed] [Google Scholar]
- 60.Brosh RM, Jr., von KC, Sommers JA, Karmakar P, Opresko PL, Piotrowski J, Dianova I, Dianov GL, Bohr VA. Werner syndrome protein interacts with human flap endonuclease 1 and stimulates its cleavage activity. EMBO J. 2001;20:5791–5801. doi: 10.1093/emboj/20.20.5791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Sharma S, Sommers JA, Wu L, Bohr VA, Hickson ID, Brosh RM., Jr. Stimulation of flap endonuclease-1 by the Bloom's syndrome protein. J. Biol. Chem. 2004;279:9847–9856. doi: 10.1074/jbc.M309898200. [DOI] [PubMed] [Google Scholar]
- 62.Cheng WH, von KC, Opresko PL, Fields KM, Ren J, Kufe D, Bohr VA. Werner syndrome protein phosphorylation by abl tyrosine kinase regulates its activity and distribution. Mol. Cell Biol. 2003;23:6385–6395. doi: 10.1128/MCB.23.18.6385-6395.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Karmakar P, Piotrowski J, Brosh RM, Jr., Sommers JA, Miller SP, Cheng WH, Snowden CM, Ramsden DA, Bohr VA. Werner protein is a target of DNA-dependent protein kinase in vivo and in vitro, and its catalytic activities are regulated by phosphorylation. J. Biol. Chem. 2002;277:18291–18302. doi: 10.1074/jbc.M111523200. [DOI] [PubMed] [Google Scholar]
- 64.Yannone SM, Roy S, Chan DW, Murphy MB, Huang S, Campisi J, Chen DJ. Werner syndrome protein is regulated and phosphorylated by DNA-dependent protein kinase. J. Biol. Chem. 2001;276:38242–38248. doi: 10.1074/jbc.M101913200. [DOI] [PubMed] [Google Scholar]
- 65.Muftuoglu M, Kusumoto R, Speina E, Beck G, Cheng WH, Bohr VA. Acetylation regulates WRN catalytic activities and affects base excision DNA repair. PLoS. One. 2008;3:e1918. doi: 10.1371/journal.pone.0001918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Dietschy T, Shevelev I, Pena-Diaz J, Huhn D, Kuenzle S, Mak R, Miah MF, Hess D, Fey M, Hottiger MO, Janscak P, Stagljar I. p300-mediated acetylation of the Rothmund-Thomson-syndrome gene product RECQL4 regulates its subcellular localization. J. Cell Sci. 2009;122:1258–1267. doi: 10.1242/jcs.037747. [DOI] [PubMed] [Google Scholar]
- 67.Dietschy T, Shevelev I, Stagljar I. The molecular role of the Rothmund-Thomson-, RAPADILINO- and Baller-Gerold-gene product, RECQL4: recent progress. Cell Mol. Life Sci. 2007;64:796–802. doi: 10.1007/s00018-007-6468-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.