

Abstract
Wls/Evi/Srt encoding a multipass transmembrane protein has been identified as a regulator for proper sorting and secretion of Wnt in flies. We have previously demonstrated that Gpr177 is the mouse orthologue required for axis determination. Gpr177 is a transcriptional target of Wnt which is activated to assist its subcellular distribution in a feedback regulatory loop. We therefore proposed that reciprocal regulation of Wnt and Gpr177 is essential for the Wnt-dependent developmental and pathogenic processes. Here we examine the expression pattern of Gpr177 in mouse development. Gpr177 is expressed in a variety of tissues and cell types during organogenesis. Furthermore, Gpr177 is a glycoprotein primarily accumulating in the Golgi apparatus in signal-producing cells. The glycosylation of Gpr177 is necessary for proper transportation in the secretory pathway. Our findings suggest that the Gpr177-mediated regulation of Wnt is crucial for organogenesis in health and disease.
Keywords: Gpr177, Wntless, Evi, Sprinter, β-catenin, Wnt production, Wnt signaling, organogenesis
Introduction
Members of the Wnt family trigger cellular signals essential for proper development of organisms (Logan and Nusse, 2004; Clevers, 2006). Aberrant regulation of an evolutionary conserved Wnt signal transduction pathway has been implicated in a variety of cancers and congenital diseases (van Amerongen and Berns, 2006; Grigoryan et al., 2008). Wnt signaling has been repetitively proven to be critical for human health and disease. Compared to the enormous wealth of knowledge on the events in signal-receiving cells, we know relatively little about the processes associated with Wnt maturation, sorting and secretion in signal-producing cells (Willert et al., 2003; Takada et al., 2006; Coudreuse and Korswagen, 2007; Hausmann et al., 2007). At the primary sequence level, Wnt proteins share a nearly invariant pattern of 23 Cys residues, an N-terminal signal sequence and several potential N-glycosylation sites (Miller, 2002). Although the role of glycosylation in Wnt secretion and function is not fully understood (Tanaka et al., 2002; Vincent and Dubois, 2002; Eaton, 2006), two lipid modifications present on the mature Wnt seem to be required for correct intracellular targeting of Wnt and signaling activity of the secreted protein (Willert et al., 2003; Zhai et al., 2004; Takada et al., 2006). Based on recent studies in the fly, Wnt proteins might be secreted bound to lipoprotein particles (Panakova et al., 2005; Eaton, 2006). Alternatively, Wnt proteins might form high order complexes, thereby overcoming their hydrophobic property and potentiating their signaling capacity (Miller, 2002).
The mechanism underlying the sorting and secretion of Wnt remains largely elusive. Studies in the fly suggest that Wnt accumulates in cells where nonconventional secretory routes may be utilized (Strigini and Cohen, 2000; Marois et al., 2006). A significant fraction of Wnt is present in endocytic vesicles which retrieve back to the surface (Pfeiffer et al., 2002). The function of this retrieval is not clear, but it might allow Wnt to gain access to cellular compartments from which standard secretory molecules are excluded. Alternatively, Wnt might directly enter these vesicles during the secretion process as the fusion of secretory vesicles from the Golgi with endosomes has been reported (Futter et al., 1995; Ang et al., 2004). The identification of Wntless (Wls/Evi/Srt) (Banziger et al., 2006; Bartscherer et al., 2006; Goodman et al., 2006) has shown that this multipass transmembrane protein is a transporting regulator for Wnt production in Drosophila. Given the extensive Wnt family in higher organisms, the essential role of Wls in production of all Wnt proteins remains to be determined.
We have previously demonstrated that Gpr177 is the mouse orthologue of Drosophila Wls, required for Wnt mediated embryonic axis determination (Fu et al., 2009). In the Gpr177 deficient mutant, Wnt production is impaired, leading to disruption of Wnt signaling in the establishment of the anterior-posterior axis. As a Wnt direct target, Gpr177 is activated by β-catenin and Lef/Tcf dependent transcription. This activation then modulates Wnt production in a positive feedback loop. Our study has led to a hypothesis in which reciprocal regulation of Wnt and Gpr177 is essential for normal developmental processes. Alterations of this regulatory circuit are causally linked to pathogenesis in human diseases.
In this study, we have examined the expression pattern of mouse Gpr177 during development of various organs. This comprehensive survey reveals that Gpr177 may modulate the Wnt pathway in a variety of tissues and cell types. Furthermore, Gpr177 is a glycoprotein predominantly localized in the Golgi apparatus. Disturbance of N-linked glycosylation prevents the Golgi accumulation of Gpr177. Given the established role of Wnt signaling in health and disease (http://www.stanford.edu/~rnusse/wntwindow.html), our findings imply that the Gpr177-mediated Wnt regulation is critical for organogenesis, including neural development, craniofacial morphogenesis and the other developmental and pathogenic processes, especially related to epithelial-mesenchymal interaction.
Results and Discussion
The expression pattern of Gpr177 mRNA and protein in mouse embryogenesis
To study the potential involvement of Gpr177 during mouse embryogenesis, we analyzed its expression by in situ hybridization. A majority of organs began to form in the developing mouse embryo after 13.5 dpc (days post coitum). We detected the Gpr177 transcript in various neural tissues, craniofacial prominences, developing skeleton and several internal organs (Figure 1A, B). Strong expression of Gpr177 was shown in certain regions of the brain which include forebrain, midbrain, hindbrain and Rathke’s pouch (future pituitary gland). Craniofacial tissues such as tooth primordium, tongue, olfactory epithelium, Meckel’s cartilage, inner ear, esophagus and lip, were also positive for the staining. In the trunk region, the stained signals were present in dorsal root ganglia, spinal cord and cartilage primordium. The Gpr177 transcript could also be found in lung, kidney, intestine, thymic primordium and urethra.
Figure 1.

Gpr177 transcript and protein are detected in multiple tissues and organs during mouse embryogenesis. In situ hybridization (A, B) and immunostaining (C, D) analyses show the expression pattern of Gpr177 in E14.5 and E13.5 embryos, respectively. Cb, cerebellum; ChP, choroid plexus; CC, cerebral cortex; CP, cartilage primordium; ccSC, central canal of spinal cord; DRG, dorsal root ganglia; Es, esophagus; Hc, hippocampus; Ht, hypothalamus; HB, hyoid bone; IE, inner ear; K, kidney; Lp, lip; Lu, lung; Mb, midbrain; MG, mid gut; MC, Meckel’s cartilage; OE, olfactory epithelium; Ov, ovary; PP, pancreatic primordium; RP, Rathke’s pouch; Tg, tongue; Th, thalamus; TP, tooth primordium; TyP; thymic primordium; Ur, urethra. Scale bar, 1 mm.
We next investigate the presence of Gpr177 protein during embryogenesis using an antibody, which we developed previously against the carboxyl terminus (Fu et al., 2009). In our previous report, this antibody recognized a ~60 Kd protein which is absent in the Gpr177 mutants, suggesting the specificity of this antibody (Fu et al., 2009). In addition, immunostaining was able to detect a strong presence of Gpr177 in the control mesoderm at E7.5 while the Gpr177-positive mesoderm was absent in the mutant (Fu et al., 2009). The protein expression analysis was in agreement with the in situ hybridization study (Figure 1C, D). Strongest levels of Gpr177 were identified in the brain regions with a very restricted expression pattern around hippocampus, thalamus, hypothalamus and ventricles. Similar to the in situ expression study, Rathke’s pouch, dorsal root ganglia, spinal cord, cartilage primordium, lung, kidney, intestine, thymic primordium, ovary and urethra were positive for the staining. Immunostained signals were also observed in the craniofacial regions, including tooth primordium, tongue, olfactory epithelium, Meckel’s cartilage, inner ear, esophagus and lip. The protein expression pattern coincides with that of the Gpr177 transcript, indicating the specificity of our Gpr177 antibody.
Early neural development
Closer examination of Gpr177 in neural development found that it is highly expressed in the neural epithelial cells of various tissues (Figure 2). Most prominent expression was detected in the dentate gyrate epithelium of hippocampus (Figure 2A, E, I, M) and epithelium of the thalamus surrounding the third ventricle (Figure 2I, M). The Gpr177 expression was distinguished in the neural epithelial cells of midbrain (Figure 2B, F, J, N), cerebellum (Figure 2C, G, K, O), pons (Figure 2D, H), medulla (Figure 2D, K, O), and spinal cord (Figure 2L, P) that line the aqueduct, forth ventricle, and central canal, especially in the ventral and dorsal regions. The choroid epithelial cells of the choroid plexus in the lateral ventricle (Figure 2I, M) and forth ventricle (Figure 2C, G, K, O) also exhibited the Gpr177 transcript and protein at high levels. The expression in the proliferating zones of these tissues suggests that Gpr177 may have a role in the expansion of neural stem/progenitor cells. Mouse genetic analyses have demonstrated that members of the Wnt family are essential for early neural development (McMahon and Bradley, 1990; Thomas and Capecchi, 1990; Hall et al., 2000; Lee et al., 2000). In addition, several Wnt signaling molecules, e.g. GSK3 and Axin1, are essential for development of neuroectoderm (Zeng et al., 1997; Kim et al., 2009). The deletion of Axin1 has been reported in human medulloblastoma (Dahmen et al., 2001). Therefore, it is highly possibly that the Gpr177 mediated Wnt production is critically linked to neural development in health and disease.
Figure 2.

Gpr177 gene is active in neural development. The expression pattern of Gpr177 in various neural tissues and cell types of the developing embryos is analyzed by in situ hybridization (A–D, I–L; E14.5) and immunostaining (E–H; E13.5, M–P; E14.5). Cb, cerebellum; ChP, choroid plexus; CC, cerebral cortex; Hc, hippocampus; Ht, hypothalamus; Mb, midbrain; Me, medulla; SC, spinal cord; Th, thalamus. Scale bar, 200 μm.
Craniofacial morphogenesis
During craniofacial morphogenesis, the expression of Gpr177 was detected in many cell types and tissues (Figure 3). In the developing incisors and molars, both epithelial cells and surrounding mesenchymal cells show expression of Gpr177 (Figure 3A–D). In humans, odonto-onycho-dermal dysplasis, a rare autosomal recessive inherited form of ectodermal dysplasia, was recently shown to be caused by a nonsense WNT10A mutation (Adaimy et al., 2007). The mutation of AXIN2, a gene encoding a negative regulator of Wnt essential for targeting β-catenin degradation, was linked to familial tooth agenesis (Lammi et al., 2004). Many previous studies (Jarvinen et al., 2006; Wang et al., 2009; Cobourne and Sharpe, 2010; Rooker et al., 2010) also support a potential role of Gpr177 in mediating Wnt production during tooth development.
Figure 3.

Expression of Gpr177 during craniofacial morphogenesis. In situ hybridization (A, C, E, G, I, K, M, O, Q, S) and immunostaining (B, D, F, H, J, L, N, P, R, T) analyses reveal that a variety of craniofacial tissues and cell types express Gpr177 at E14.5. Co, cochlea; HF, hair follicle; Ln, lens; OE, olfactory epithelium; OM, ocular muscle; ON, optic nerve; OR, optic recess; P, palate; SC, semicircular canal; SG, salivary gland; SL, sublingual duct; SM, submandibular duct; Tg, tongue; TP, tooth primordium. Scale bars, 100 μm (A–D, G, H, S, T); 50 μm (Q, R); 200 μm (E, F, I–P).
Non-syndromic cleft lip with or without cleft palate is one of the most common birth defects. Genetic variations in several Wnt family members were shown to be associated with this phenotype in humans (Chiquet et al., 2008). Mouse genetic analysis also indicated that Wnt5a (He et al., 2008) and Wnt9b (Juriloff et al., 2006) are essential for palate development. Indeed, we were able to identify the presence of Gpr177 mRNA and protein during embryonic development of the lip (Figure 1A, C) and palate (Figure 3E, F). Strong Gpr177 expression was also found in the tongue (Figure 3E, F) that maybe related to the role of Wnt signaling in formation of taste papilla (Iwatsuki et al., 2007).
The Wnt pathway has a well established role in development of the skin and hair follicles (Alonso and Fuchs, 2003; Haegebarth and Clevers, 2009). There was no surprise to identify the Gpr177 transcript and protein in these regions (Figure 3G, H). Other sensory organs, including inner ear (Figure 3I, J), olfactory epithelium (Figure 3K, L), eye (Figure 3M–P), were also positive for the expression of Gpr177. Both canonical and non canonical Wnt proteins were shown to control formation of the inner ear (Riccomagno et al., 2005; Qian et al., 2007). It has been suggested that Wnt3 acts as an axon guidance molecule to mediate medial-lateral retinotecal topography (Schmitt et al., 2006). The localization of the Gpr177 mRNA and protein supports the role of Wnt signaling in retinal and lens development (Van Raay and Vetter, 2004; Lovicu and McAvoy, 2005; Silver and Rebay, 2005). However, the important function of Gpr177 and Wnt-mediated development in the olfactory system remains largely to be explored. In the salivary gland, Wnt stimulation was linked to pleomorphic adenoma, the most common type of salivary gland tumor in humans (Zhao et al., 2006; Declercq et al., 2008). Strong expression of Gpr177 was detected in the epithelial components of major salivary glands to a lesser extent in the mesenchymal regions (Figure 3Q–T). Nevertheless, the requirement of Wnt signaling in normal development of the salivary glands remains to be determined.
Internal organs
We next examined if Gpr177 is also expressed in development of other major organs. In the kidney, the Gpr177 mRNA and protein were detected predominantly in the ureteric epithelium, but not the differentiated comma and S-shape bodies (Figure 4A–D). A weak mesenchymal expression was also observed, suggesting that Gpr177 may regulate the sorting and secretion of Wnt4, Wnt7b, Wnt9b and Wnt11, which have been implicated in kidney organogenesis (Stark et al., 1994; Majumdar et al., 2003; Park et al., 2007; Karner et al., 2009; Yu et al., 2009). In agreement with mouse genetic studies of the Wnt family proteins (Li et al., 2002; Shu et al., 2002; Rajagopal et al., 2008; Goss et al., 2009), our data showed that Gpr177 is expressed in both epithelial and mesenchymal cells, potentially involved in lung development (Figure 4E, F). A selective expression of Gpr177 was found in the thymic primordium (Figure 4G, H). Given the well established role of Wnt in hematopoiesis (Staal and Clevers, 2005; Grigoryan et al., 2008; Malhotra and Kincade, 2009), Gpr177 is likely to play a role in development of the hematopoietic lineages. In the developing gut, the expression was found in both epithelium and mesenchyme of the esophagus (Figure 4I, J), mid gut (Figure 1B), duodenum (Figure 4K, L), suggesting the Gpr177-dependent regulation of Wnt may be critical for development of the digestive system.
Figure 4.
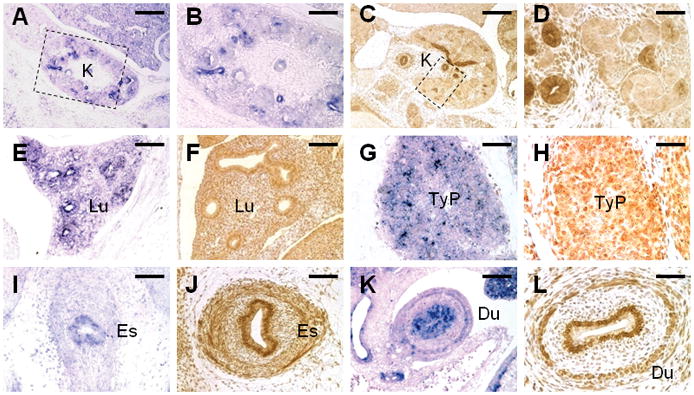
Expression of Gpr177 in development of multiple organs. The mRNA and protein expression of Gpr177 is examined by in situ hybridization (A, B, E, G, I, K) and immunostaining (C, D, F, H, J, L) of E14.5 embryos, respectively. Insets in A, C denote higher magnification in B, D, respectively. Du, duodenum; Es, esophagus; K, kidney; Lu, lung; TyP, thymic primordium. Scale bars, 200 μm (A, C); 100 μm (B, E, F, I, K); 50 μm (D, G, H, J, L).
Glycosylation of Gpr177
We previously showed that Gpr177 is differentially localized in the Wnt-producing and non Wnt-producing cells (Fu et al., 2009). The differential compartmentalization is dependent upon the positive feedback regulation of Wnt to activate the Gpr177 expression at the transcriptional level (Fu et al., 2009). The Golgi accumulation of Gpr177 can only be identified in the signal-producing cells, such as neural stem cells (Fu et al., 2009). Because membrane and secreted proteins are modified by saccharides, we examined whether inhibition of glycosylation interferes with the Golgi accumulation of Gpr177. In the culture of neurosphere cells, Gpr177 is co-localized with a Golgi marker GM130 (Figure 5A). The addition of tunicamycin, which inhibits the enzyme GlcNAc phosphotransferase involved in the first step of glycoprotein synthesis, affects concentrations of Gpr177 in the Golgi apparatus (Figure 5A). Treatment of tunicamycin prevented the co-localization of Gpr177 with GM130, suggesting that glycosylation is required for the Golgi accumulation. Immunoblot analysis indicated that a slow migrating band disappears after the tunicamycin treatment (Figure 5B). Furthermore, the addition of Endo Hf diminished the detection of Gpr177 with slow mobility (Figure 5C). The results suggest that Gpr177 is modified by N-linked glycosylation required for proper sorting within the cells.
Figure 5.
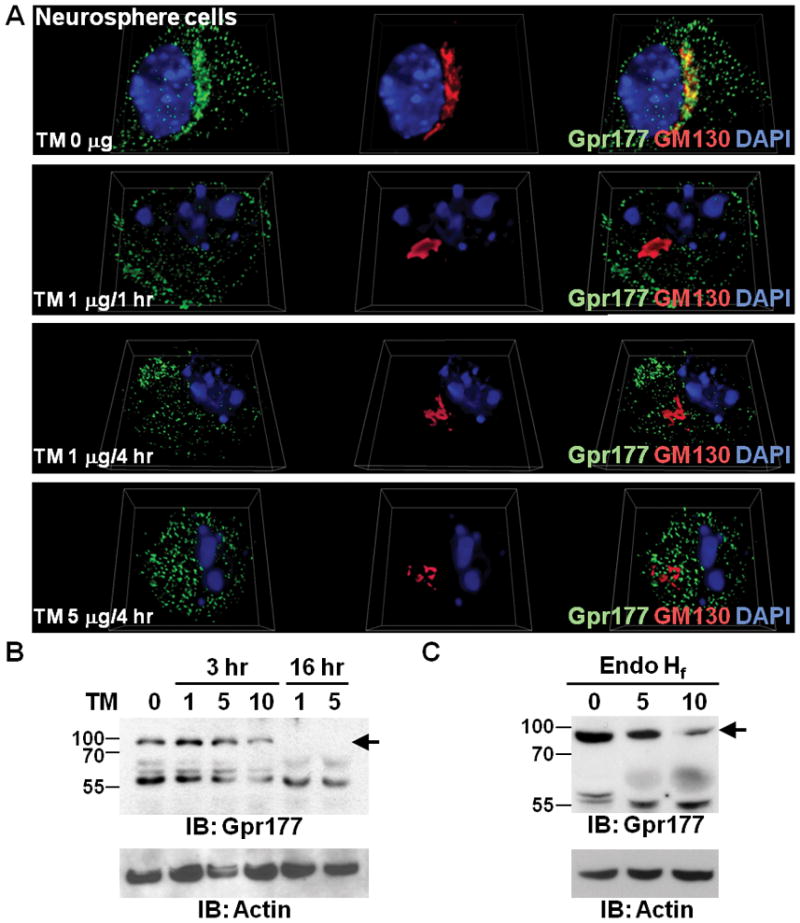
Glycosylation is required for Golgi distributions of Gpr177. (A) Neurosphere cells were treated with Tunicamycin (TM) for different concentrations and time periods as indicated. 3D imaging of the immunostained cells shows the localization of endogenous Gpr177 (green) and GM130 (red), counterstained with DAPI (blue). Note that the TM treatment disrupts the co-localization of Gpr177 and GM130 in Golgi. (B) Immunoblot analysis of Gpr177 reveals that TM abolishes the detection of Gpr177 with slow mobility (arrow) in neural stem cells. The number indicates the drug concentration (μg/ml) present in culture media. The expression level of Actin was analyzed as a loading control. (C) Immunoblot analysis shows that the slow migrating band (arrow) detected by the Gpr177 antibody disappears after the addition of Endo Hf. The number presents the amount of enzyme (μl: 1,000 units per μl) added to the in vitro assays. The expression level of Actin was analyzed as a loading control.
In summary, we have performed a comprehensive survey on the expression of Gpr177 using in situ hybridization and immunostaining analyses. The expression patterns of mRNA and protein are in agreement with each other, supporting the notion that Wnt signaling is critical for development of multiple organs. Our prior discovery of Gpr177 essential for modulating canonical Wnt in early embryogenesis implies that their reciprocal regulations are likely to be important for other developmental processes. Gpr177 might also be associated with the Wnt-mediated malignant transformation of these organs. The expression of Gpr177 in development of several organs requiring the epithelial-mesenchymal interaction adds another layer of regulation for signal-producing and signal-receiving cells. In addition, the Wnt-producing cells are able to initiate paracrine as well as autocrine signaling effects. Gpr177 seems to be expressed in places where non canonical Wnt proteins have essential functions. However, whether non canonical Wnt is regulated by Gpr177 remains an important question to be determined, especially to be assessed functionally by genetic analysis. Using large-scale meta-analysis of genome-wide association, a recent study has identified GPR177 as one of the twenty bone-mineral-density loci in humans (Rivadeneira et al., 2009). Among them, CTNNB1, encoding β-catenin, is another locus identified at the genome-wide study level. The finding implies that Gpr177 and β-catenin are master regulators for Wnt production and signaling, respectively. One can expect further discovery revealing the significance of the Gpr177-mediated regulation of Wnt in skeletal development and disease.
Methods
Mouse embryos were first fixed, paraffin embedded, sectioned, and stained with hematoxylin/eosin for histological evaluation as described (Yu et al., 2007; Hsu et al., 2010). Sections were then subject to in situ hybridization with digoxygenin labeled anti-sense RNA probes or immunological staining with avidin:biotinlylated enzyme complex as described (Yu et al., 2005a; Liu et al., 2007; Chiu et al., 2008). For in situ hybridization, RNA probes for detecting the Gpr177 transcript as well as the detailed method were described previously (Chiu et al., 2008; Fu et al., 2009). Rabbit polyclonal antibodies were generated for the immunostaining analysis of Gpr177 as described (Fu et al., 2009). Briefly, sections were deparaffinized and hydrated, followed by antigen unmasking (Vector Laboratories, Burlingame, CA). Endogenous peroxidase activity was blocked by incubating sections with 3% hydrogen peroxide. After addition of primary antibodies, sections were incubated with horseradish peroxidase-conjugated secondary antibodies. The staining was then visualized by enzymatic color reaction according to the manufacture’s specification (Vector Laboratories). Images were analyzed using Zeiss AXIO OBSERVER with AXIOCAM or Nikon SMZ-1500 with SPOT-RT microscope imaging systems (Yu et al., 2005b; Chiu et al., 2008; Liu et al., 2008).
Isolation and culture of primary neurospheres were performed as described (Fu et al., 2009). Briefly, telencephalons of E12.5 mouse embryos were recovered and mechanically dissociated in DMEM/F12 medium (1:1; Invitrogen, Carlsbad, CA), followed by filtering through a 70 μm nylon mesh (BD Biosciences, Bedford, MA). Cells were then cultured in DMEM/F12 medium, supplemented with Insulin-Transferrin-Selenium supplements (Invitrogen), EGF (20ng/ml; Sigma-Aldrich, St. Louis, MO), and antibiotic solution (Sigma-Aldrich), at 37°C in a humidified 5% CO2 incubator. After 5 days, neurospheres were harvested by centrifugation, dissociated by Trypsin-EDTA (Sigma-Aldrich), and continued to be cultured for next passages or used in experimental procedures.
Immunostaining of cultured neurosphere cells was performed using indirect fluorescent staining techniques described previously (Fu et al., 2009). Images were taken using AXIO OBSERVER with AXIOCAM microscope imaging system, followed by deconvolution, 3D imaging analysis. Immunoblot analysis was performed as described (Liu et al., 2008; Fu et al., 2009). Bound primary antibodies were detected with horseradish peroxidase-conjugated secondary antibodies, followed by enhanced chemical luminescence-mediated visualization (GE Healthcare, Piscataway, NJ) and autoradiography.
Acknowledgments
This work is support by National Institutes of Health: CA106308 and DE015654
We thank Anthony Mirando for preparation of the manuscript. This work is supported by National Institutes of Health grants CA106308 and DE015654 to W.H.
References
- Adaimy L, Chouery E, Megarbane H, Mroueh S, Delague V, Nicolas E, Belguith H, de Mazancourt P, Megarbane A. Mutation in WNT10A is associated with an autosomal recessive ectodermal dysplasia: the odonto-onycho-dermal dysplasia. Am J Hum Genet. 2007;81:821–828. doi: 10.1086/520064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alonso L, Fuchs E. Stem cells in the skin: waste not, Wnt not. Genes Dev. 2003;17:1189–1200. doi: 10.1101/gad.1086903. [DOI] [PubMed] [Google Scholar]
- Ang AL, Taguchi T, Francis S, Folsch H, Murrells LJ, Pypaert M, Warren G, Mellman I. Recycling endosomes can serve as intermediates during transport from the Golgi to the plasma membrane of MDCK cells. J Cell Biol. 2004;167:531–543. doi: 10.1083/jcb.200408165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banziger C, Soldini D, Schutt C, Zipperlen P, Hausmann G, Basler K. Wntless, a conserved membrane protein dedicated to the secretion of Wnt proteins from signaling cells. Cell. 2006;125:509–522. doi: 10.1016/j.cell.2006.02.049. [DOI] [PubMed] [Google Scholar]
- Bartscherer K, Pelte N, Ingelfinger D, Boutros M. Secretion of Wnt ligands requires Evi, a conserved transmembrane protein. Cell. 2006;125:523–533. doi: 10.1016/j.cell.2006.04.009. [DOI] [PubMed] [Google Scholar]
- Chiquet BT, Blanton SH, Burt A, Ma D, Stal S, Mulliken JB, Hecht JT. Variation in WNT genes is associated with non-syndromic cleft lip with or without cleft palate. Hum Mol Genet. 2008;17:2212–2218. doi: 10.1093/hmg/ddn121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiu SY, Asai N, Costantini F, Hsu W. SUMO-Specific Protease 2 Is Essential for Modulating p53-Mdm2 in Development of Trophoblast Stem Cell Niches and Lineages. PLoS Biol. 2008;6:e310. doi: 10.1371/journal.pbio.0060310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clevers H. Wnt/beta-catenin signaling in development and disease. Cell. 2006;127:469–480. doi: 10.1016/j.cell.2006.10.018. [DOI] [PubMed] [Google Scholar]
- Cobourne MT, Sharpe PT. Making up the numbers: The molecular control of mammalian dental formula. Semin Cell Dev Biol. 2010 doi: 10.1016/j.semcdb.2010.01.007. [DOI] [PubMed] [Google Scholar]
- Coudreuse D, Korswagen HC. The making of Wnt: new insights into Wnt maturation, sorting and secretion. Development. 2007;134:3–12. doi: 10.1242/dev.02699. [DOI] [PubMed] [Google Scholar]
- Dahmen RP, Koch A, Denkhaus D, Tonn JC, Sorensen N, Berthold F, Behrens J, Birchmeier W, Wiestler OD, Pietsch T. Deletions of AXIN1, a component of the WNT/wingless pathway, in sporadic medulloblastomas. Cancer Res. 2001;61:7039–7043. [PubMed] [Google Scholar]
- Declercq J, Van Dyck F, Van Damme B, Van de Ven WJ. Upregulation of Igf and Wnt signalling associated genes in pleomorphic adenomas of the salivary glands in PLAG1 transgenic mice. Int J Oncol. 2008;32:1041–1047. [PubMed] [Google Scholar]
- Eaton S. Release and trafficking of lipid-linked morphogens. Curr Opin Genet Dev. 2006;16:17–22. doi: 10.1016/j.gde.2005.12.006. [DOI] [PubMed] [Google Scholar]
- Fu J, Jiang M, Mirando AJ, Yu HM, Hsu W. Reciprocal regulation of Wnt and Gpr177/mouse Wntless is required for embryonic axis formation. Proc Natl Acad Sci U S A. 2009;106:18598–18603. doi: 10.1073/pnas.0904894106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Futter CE, Connolly CN, Cutler DF, Hopkins CR. Newly synthesized transferrin receptors can be detected in the endosome before they appear on the cell surface. J Biol Chem. 1995;270:10999–11003. doi: 10.1074/jbc.270.18.10999. [DOI] [PubMed] [Google Scholar]
- Goodman RM, Thombre S, Firtina Z, Gray D, Betts D, Roebuck J, Spana EP, Selva EM. Sprinter: a novel transmembrane protein required for Wg secretion and signaling. Development. 2006;133:4901–4911. doi: 10.1242/dev.02674. [DOI] [PubMed] [Google Scholar]
- Goss AM, Tian Y, Tsukiyama T, Cohen ED, Zhou D, Lu MM, Yamaguchi TP, Morrisey EE. Wnt2/2b and beta-catenin signaling are necessary and sufficient to specify lung progenitors in the foregut. Dev Cell. 2009;17:290–298. doi: 10.1016/j.devcel.2009.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grigoryan T, Wend P, Klaus A, Birchmeier W. Deciphering the function of canonical Wnt signals in development and disease: conditional loss- and gain-of-function mutations of beta-catenin in mice. Genes Dev. 2008;22:2308–2341. doi: 10.1101/gad.1686208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haegebarth A, Clevers H. Wnt signaling, lgr5, and stem cells in the intestine and skin. Am J Pathol. 2009;174:715–721. doi: 10.2353/ajpath.2009.080758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall AC, Lucas FR, Salinas PC. Axonal remodeling and synaptic differentiation in the cerebellum is regulated by WNT-7a signaling. Cell. 2000;100:525–535. doi: 10.1016/s0092-8674(00)80689-3. [DOI] [PubMed] [Google Scholar]
- Hausmann G, Banziger C, Basler K. Helping Wingless take flight: how WNT proteins are secreted. Nat Rev Mol Cell Biol. 2007;8:331–336. doi: 10.1038/nrm2141. [DOI] [PubMed] [Google Scholar]
- He F, Xiong W, Yu X, Espinoza-Lewis R, Liu C, Gu S, Nishita M, Suzuki K, Yamada G, Minami Y, Chen Y. Wnt5a regulates directional cell migration and cell proliferation via Ror2-mediated noncanonical pathway in mammalian palate development. Development. 2008;135:3871–3879. doi: 10.1242/dev.025767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu W, Mirando AJ, Yu HM. Manipulating gene activity in Wnt1-expressing precursors of neural epithelial and neural crest cells. Dev Dyn. 2010;239:338–345. doi: 10.1002/dvdy.22044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iwatsuki K, Liu HX, Gronder A, Singer MA, Lane TF, Grosschedl R, Mistretta CM, Margolskee RF. Wnt signaling interacts with Shh to regulate taste papilla development. Proc Natl Acad Sci U S A. 2007;104:2253–2258. doi: 10.1073/pnas.0607399104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarvinen E, Salazar-Ciudad I, Birchmeier W, Taketo MM, Jernvall J, Thesleff I. Continuous tooth generation in mouse is induced by activated epithelial Wnt/beta-catenin signaling. Proc Natl Acad Sci U S A. 2006;103:18627–18632. doi: 10.1073/pnas.0607289103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juriloff DM, Harris MJ, McMahon AP, Carroll TJ, Lidral AC. Wnt9b is the mutated gene involved in multifactorial nonsyndromic cleft lip with or without cleft palate in A/WySn mice, as confirmed by a genetic complementation test. Birth Defects Res A Clin Mol Teratol. 2006;76:574–579. doi: 10.1002/bdra.20302. [DOI] [PubMed] [Google Scholar]
- Karner CM, Chirumamilla R, Aoki S, Igarashi P, Wallingford JB, Carroll TJ. Wnt9b signaling regulates planar cell polarity and kidney tubule morphogenesis. Nat Genet. 2009;41:793–799. doi: 10.1038/ng.400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim WY, Wang X, Wu Y, Doble BW, Patel S, Woodgett JR, Snider WD. GSK-3 is a master regulator of neural progenitor homeostasis. Nat Neurosci. 2009;12:1390–1397. doi: 10.1038/nn.2408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lammi L, Arte S, Somer M, Jarvinen H, Lahermo P, Thesleff I, Pirinen S, Nieminen P. Mutations in AXIN2 cause familial tooth agenesis and predispose to colorectal cancer. Am J Hum Genet. 2004;74:1043–1050. doi: 10.1086/386293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee SM, Tole S, Grove E, McMahon AP. A local Wnt-3a signal is required for development of the mammalian hippocampus. Development. 2000;127:457–467. doi: 10.1242/dev.127.3.457. [DOI] [PubMed] [Google Scholar]
- Li C, Xiao J, Hormi K, Borok Z, Minoo P. Wnt5a participates in distal lung morphogenesis. Dev Biol. 2002;248:68–81. doi: 10.1006/dbio.2002.0729. [DOI] [PubMed] [Google Scholar]
- Liu B, Yu HM, Hsu W. Craniosynostosis caused by Axin2 deficiency is mediated through distinct functions of beta-catenin in proliferation and differentiation. Dev Biol. 2007;301:298–308. doi: 10.1016/j.physletb.2003.10.071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu B, Yu HM, Huang J, Hsu W. Co-opted JNK/SAPK signaling in Wnt/beta-catenin-induced tumorigenesis. Neoplasia. 2008;10:1004–1013. doi: 10.1593/neo.08548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol. 2004;20:781–810. doi: 10.1146/annurev.cellbio.20.010403.113126. [DOI] [PubMed] [Google Scholar]
- Lovicu FJ, McAvoy JW. Growth factor regulation of lens development. Dev Biol. 2005;280:1–14. doi: 10.1016/j.ydbio.2005.01.020. [DOI] [PubMed] [Google Scholar]
- Majumdar A, Vainio S, Kispert A, McMahon J, McMahon AP. Wnt11 and Ret/Gdnf pathways cooperate in regulating ureteric branching during metanephric kidney development. Development. 2003;130:3175–3185. doi: 10.1242/dev.00520. [DOI] [PubMed] [Google Scholar]
- Malhotra S, Kincade PW. Wnt-related molecules and signaling pathway equilibrium in hematopoiesis. Cell Stem Cell. 2009;4:27–36. doi: 10.1016/j.stem.2008.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marois E, Mahmoud A, Eaton S. The endocytic pathway and formation of the Wingless morphogen gradient. Development. 2006;133:307–317. doi: 10.1242/dev.02197. [DOI] [PubMed] [Google Scholar]
- McMahon AP, Bradley A. The Wnt-1 (int-1) proto-oncogene is required for development of a large region of the mouse brain. Cell. 1990;62:1073–1085. doi: 10.1016/0092-8674(90)90385-r. [DOI] [PubMed] [Google Scholar]
- Miller JR. The Wnts Genome Biol. 2002;3:REVIEWS3001. doi: 10.1186/gb-2001-3-1-reviews3001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panakova D, Sprong H, Marois E, Thiele C, Eaton S. Lipoprotein particles are required for Hedgehog and Wingless signalling. Nature. 2005;435:58–65. doi: 10.1038/nature03504. [DOI] [PubMed] [Google Scholar]
- Park JS, Valerius MT, McMahon AP. Wnt/beta-catenin signaling regulates nephron induction during mouse kidney development. Development. 2007;134:2533–2539. doi: 10.1242/dev.006155. [DOI] [PubMed] [Google Scholar]
- Pfeiffer S, Ricardo S, Manneville JB, Alexandre C, Vincent JP. Producing cells retain and recycle Wingless in Drosophila embryos. Curr Biol. 2002;12:957–962. doi: 10.1016/s0960-9822(02)00867-9. [DOI] [PubMed] [Google Scholar]
- Qian D, Jones C, Rzadzinska A, Mark S, Zhang X, Steel KP, Dai X, Chen P. Wnt5a functions in planar cell polarity regulation in mice. Dev Biol. 2007;306:121–133. doi: 10.1016/j.ydbio.2007.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajagopal J, Carroll TJ, Guseh JS, Bores SA, Blank LJ, Anderson WJ, Yu J, Zhou Q, McMahon AP, Melton DA. Wnt7b stimulates embryonic lung growth by coordinately increasing the replication of epithelium and mesenchyme. Development. 2008;135:1625–1634. doi: 10.1242/dev.015495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riccomagno MM, Takada S, Epstein DJ. Wnt-dependent regulation of inner ear morphogenesis is balanced by the opposing and supporting roles of Shh. Genes Dev. 2005;19:1612–1623. doi: 10.1101/gad.1303905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivadeneira F, Styrkarsdottir U, Estrada K, Halldorsson BV, Hsu YH, Richards JB, Zillikens MC, Kavvoura FK, Amin N, Aulchenko YS, Cupples LA, Deloukas P, Demissie S, Grundberg E, Hofman A, Kong A, Karasik D, van Meurs JB, Oostra B, Pastinen T, Pols HA, Sigurdsson G, Soranzo N, Thorleifsson G, Thorsteinsdottir U, Williams FM, Wilson SG, Zhou Y, Ralston SH, van Duijn CM, Spector T, Kiel DP, Stefansson K, Ioannidis JP, Uitterlinden AG. Twenty bone-mineral-density loci identified by large-scale meta-analysis of genome-wide association studies. Nat Genet. 2009;41:1199–1206. doi: 10.1038/ng.446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rooker SM, Liu B, Helms JA. Role of Wnt signaling in the biology of the periodontium. Dev Dyn. 2010;239:140–147. doi: 10.1002/dvdy.22003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmitt AM, Shi J, Wolf AM, Lu CC, King LA, Zou Y. Wnt-Ryk signalling mediates medial-lateral retinotectal topographic mapping. Nature. 2006;439:31–37. doi: 10.1038/nature04334. [DOI] [PubMed] [Google Scholar]
- Shu W, Jiang YQ, Lu MM, Morrisey EE. Wnt7b regulates mesenchymal proliferation and vascular development in the lung. Development. 2002;129:4831–4842. doi: 10.1242/dev.129.20.4831. [DOI] [PubMed] [Google Scholar]
- Silver SJ, Rebay I. Signaling circuitries in development: insights from the retinal determination gene network. Development. 2005;132:3–13. doi: 10.1242/dev.01539. [DOI] [PubMed] [Google Scholar]
- Staal FJ, Clevers HC. WNT signalling and haematopoiesis: a WNT-WNT situation. Nat Rev Immunol. 2005;5:21–30. doi: 10.1038/nri1529. [DOI] [PubMed] [Google Scholar]
- Stark K, Vainio S, Vassileva G, McMahon AP. Epithelial transformation of metanephric mesenchyme in the developing kidney regulated by Wnt-4. Nature. 1994;372:679–683. doi: 10.1038/372679a0. [DOI] [PubMed] [Google Scholar]
- Strigini M, Cohen SM. Wingless gradient formation in the Drosophila wing. Curr Biol. 2000;10:293–300. doi: 10.1016/s0960-9822(00)00378-x. [DOI] [PubMed] [Google Scholar]
- Takada R, Satomi Y, Kurata T, Ueno N, Norioka S, Kondoh H, Takao T, Takada S. Monounsaturated fatty acid modification of Wnt protein: its role in Wnt secretion. Dev Cell. 2006;11:791–801. doi: 10.1016/j.devcel.2006.10.003. [DOI] [PubMed] [Google Scholar]
- Tanaka K, Kitagawa Y, Kadowaki T. Drosophila segment polarity gene product porcupine stimulates the posttranslational N-glycosylation of wingless in the endoplasmic reticulum. J Biol Chem. 2002;277:12816–12823. doi: 10.1074/jbc.M200187200. [DOI] [PubMed] [Google Scholar]
- Thomas KR, Capecchi MR. Targeted disruption of the murine int-1 proto-oncogene resulting in severe abnormalities in midbrain and cerebellar development. Nature. 1990;346:847–850. doi: 10.1038/346847a0. [DOI] [PubMed] [Google Scholar]
- van Amerongen R, Berns A. Knockout mouse models to study Wnt signal transduction. Trends Genet. 2006;22:678–689. doi: 10.1016/j.tig.2006.10.001. [DOI] [PubMed] [Google Scholar]
- Van Raay TJ, Vetter ML. Wnt/frizzled signaling during vertebrate retinal development. Dev Neurosci. 2004;26:352–358. doi: 10.1159/000082277. [DOI] [PubMed] [Google Scholar]
- Vincent JP, Dubois L. Morphogen transport along epithelia, an integrated trafficking problem. Dev Cell. 2002;3:615–623. doi: 10.1016/s1534-5807(02)00323-4. [DOI] [PubMed] [Google Scholar]
- Wang XP, O’Connell DJ, Lund JJ, Saadi I, Kuraguchi M, Turbe-Doan A, Cavallesco R, Kim H, Park PJ, Harada H, Kucherlapati R, Maas RL. Apc inhibition of Wnt signaling regulates supernumerary tooth formation during embryogenesis and throughout adulthood. Development. 2009;136:1939–1949. doi: 10.1242/dev.033803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willert K, Brown JD, Danenberg E, Duncan AW, Weissman IL, Reya T, Yates JR, Nusse R. Wnt proteins are lipid-modified and can act as stem cell growth factors. Nature. 2003 doi: 10.1038/nature01611. [DOI] [PubMed] [Google Scholar]
- Yu HM, Jerchow B, Sheu TJ, Liu B, Costantini F, Puzas JE, Birchmeier W, Hsu W. The role of Axin2 in calvarial morphogenesis and craniosynostosis. Development. 2005a;132:1995–2005. doi: 10.1242/dev.01786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu HM, Liu B, Chiu SY, Costantini F, Hsu W. Development of a unique system for spatiotemporal and lineage-specific gene expression in mice. Proc Natl Acad Sci U S A. 2005b;102:8615–8620. doi: 10.1073/pnas.0500124102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu HM, Liu B, Costantini F, Hsu W. Impaired neural development caused by inducible expression of Axin in transgenic mice. Mech Dev. 2007;124:146–156. doi: 10.1016/j.mod.2006.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu J, Carroll TJ, Rajagopal J, Kobayashi A, Ren Q, McMahon AP. A Wnt7b-dependent pathway regulates the orientation of epithelial cell division and establishes the cortico-medullary axis of the mammalian kidney. Development. 2009;136:161–171. doi: 10.1242/dev.022087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeng L, Fagotto F, Zhang T, Hsu W, Vasicek TJ, Perry WL, 3rd, Lee JJ, Tilghman SM, Gumbiner BM, Costantini F. The mouse Fused locus encodes Axin, an inhibitor of the Wnt signaling pathway that regulates embryonic axis formation. Cell. 1997;90:181–192. doi: 10.1016/s0092-8674(00)80324-4. [DOI] [PubMed] [Google Scholar]
- Zhai L, Chaturvedi D, Cumberledge S. Drosophila wnt-1 undergoes a hydrophobic modification and is targeted to lipid rafts, a process that requires porcupine. J Biol Chem. 2004;279:33220–33227. doi: 10.1074/jbc.M403407200. [DOI] [PubMed] [Google Scholar]
- Zhao X, Ren W, Yang W, Wang Y, Kong H, Wang L, Yan L, Xu G, Fei J, Fu J, Zhang C, Wang Z. Wnt pathway is involved in pleomorphic adenomas induced by overexpression of PLAG1 in transgenic mice. Int J Cancer. 2006;118:643–648. doi: 10.1002/ijc.21400. [DOI] [PubMed] [Google Scholar]