

Abstract
Procedural skills such as riding a bicycle and playing a musical instrument play a central role in daily life. Such skills are learned gradually and are retained throughout life. The present study investigated one-year retention of procedural skill in a version of the widely used serial reaction time task (SRTT) in young and older motor-skill experts and older controls in two experiments. The young experts were college-age piano and action video-game players, and the older experts were piano players. Previous studies have reported sequence-specific skill retention in the SRTT as long as two weeks but not at one year. Results indicated that both young and older experts and older non-experts revealed sequence-specific skill retention after one year with some evidence that general motor skill was retained as well. These findings are consistent with theoretical accounts of procedural skill learning such as the procedural reinstatement theory as well as with previous studies of retention of other motor skills.
Keywords: Implicit learning, sequence learning, aging, motor skill expertise, piano players, video-game players
Over time, people acquire a wide range of implicit skills including riding a bicycle, playing a musical instrument, and displaying fluency in language. A prominent characteristic of such skills is that they are learned only gradually through extended experience, but once learned, they are retained throughout life. This property of skill learning has both practical and theoretical significance. The practical interest stems from the fact that rehabilitation following neurological damage or brain insult, such as stroke, often requires re-learning basic life skills such as walking and speaking (e.g., Boyd, Vidoni, & Daly, 2007), and expertise in specialties such as laparoscopic surgery is developed in part through procedural learning (Stefanidis, Korndorffer, Sierra, Touchard, Dunne, & Scott, 2005). The theoretical interest derives from contemporary multi-system theories that posit that distinct brain regions support non-declarative learning, which occurs without awareness, and declarative learning, which occurs with awareness. The non-declarative procedural memory system, viewed as engaging cortico-striatial circuits, is often contrasted with the medial temporal lobe-dependent declarative system, which acquires new information quickly and flexibly but is less long-lived (Clark, Manns, & Squire, 2002; Poldrack & Packard, 2003; Reber & Squire, 1998; Squire, Clark, & Bayley, 2004). Many laboratory studies have investigated how declarative and non-declarative skills are learned, and many have also examined long-term retention of declarative skills. However, surprisingly few studies have examined long-term retention of non-declarative procedural skills. Healy and colleagues have shown retention of target detection over fifteen months (Healy, Fendrich, & Proctor, 1990), Morse code reception (Clawson, Healy, Ericsson, & Bourne, 2001) and entry of digit lists (Fendrich, Healy, & Bourne, 1991) after one month, and temporal duration production (Healy, Wohldmann, Parker, & Bourne, 2005) and speeded aiming (Healy, Wohldmann, Sutton, & Bourne, 2006) after one week. In addition, Cooke, Durso and Schvaneveldt (1994) have shown retention of skilled letter and digit search after nine years, and Allen and Reber (1980) have shown retention of an artificial grammar learning task after two years. The present study expands upon previous work by demonstrating long-term retention of a sequential skill in which people learn the predictive relationships among temporally ordered events. Specifically, we use a variation of the widely used serial reaction time task (SRTT) to demonstrate one-year retention of an implicitly acquired sequential skill in both young and older adults.
The SRTT, which has been shown to depend primarily on cortico-striatal networks, has been widely used as a laboratory assay of procedural skill learning (for reviews, see Forkstam & Petersson, 2005; Robertson, 2007). In the SRTT, events, such as asterisks, appear in one of four different locations on a computer screen. Participants are instructed to press the key corresponding to each event, as quickly as possible when it appears, while maintaining a high level of accuracy. The positions at which the events appear on the screen occur in a specific repeating pattern unbeknownst to the participant. Learning is assessed by comparing performance on the pattern trial blocks to performance on random event blocks, which occur either within or at the end of the session. Sequence learning is said to occur when individuals are faster on pattern blocks compared to random blocks.
Both young and older adults show learning of both simple repeating (D. V. Howard & Howard, 1989, 1992) and subtle probabilistic regularities (D. V. Howard, Howard, Japikse, DiYanni, Thompson, & Somberg, 2004; J. H. Howard, Jr. & Howard, 1997; Schendan, Searl, Melrose, & Stern, 2003; Schvaneveldt & Gomez, 1998) in the SRTT. Although some studies have demonstrated retention of sequence-specific skill for periods up to 16 months in other sequential response tasks (Hikosaka, Rand, Nakamura, Miyachi, Kitaguchi, Sakai, et al., 2002; Savion-Lemieux & Penhune, 2005), no published study to date has demonstrated retention of sequence-specific procedural skill in the SRTT for longer than two months (Feeney, 2000; Knopman, 1991; Nissen, Willingham, & Hartman, 1989).
The present experiment examines one-year retention of procedural skill in a version of the SRTT that can be practiced over many trials without acquiring declarative knowledge. In the Alternating SRTT (ASRTT), a four-element repeating sequence of pattern stimuli alternate with random stimuli (J. H. Howard, Jr. & Howard, 1997). This regularity is subtler than the more obvious pattern in the traditional SRTT. For example, in the traditional SRTT, a typical pattern might be 1432134123 (where 1, 2, 3, and 4 refer to four targets on a computer screen, left to right, respectively), and this 10-element pattern is repeated over many blocks. General skill learning is measured by an overall reduction in RT over the pattern blocks, and sequence-specific learning is measured by comparing performance on the pattern-trial blocks to performance on a random-trial block that is introduced later in the session. People often acquire declarative knowledge of these simple repeating patterns. In contrast, in the ASRTT, a typical pattern might be 1r4r3r2r, where r refers to a randomly chosen target, and this 8-element pattern is repeated throughout the session. General skill learning is measured by an overall reduction in RT, and sequence-specific learning is demonstrated by the pattern and random trials diverging over time. People do not acquire declarative knowledge, as the simplest predictive relationship in an ASRTT sequence is between non-adjacent events on pattern trials (trial n-2 predicts the target on pattern trial n) (J. H. Howard, Jr., & Howard, 1997).
The primary goal of the present study was to investigate long-term retention of sequence-specific and general-procedural skills in the ASRTT by inviting participants from two earlier experiments to return to the lab for further testing after approximately one year. The earlier experiments examined learning in young and older adult perceptual-motor skill experts – the younger experts in Experiment 1 were piano players or action video-game players, whereas the older adults in Experiment 2 were piano players or age-matched non-musician controls. Music and video-game expert populations have been shown to have enhanced performance on a number of perceptual, cognitive, and motor tasks (Castel, Pratt, & Drummond, 2005; Elbert, Pantev, Wienbruch, Rockstroh, & Taub, 1995; Feng, Spence, & Pratt, 2007; Green & Bavelier, 2003), and the skill learning in these groups typically results in improvement in perceptual, cognitive and motor functioning that persists over time (Green & Bavelier, 2008). Thus, we were interested in examining skill retention in young and older motor-skill experts as well as non-expert older adults.
Experiment 1
Method
Participants
Introductory Psychology students at The Catholic University of America participated in the original ASRTT study for course credit (Romano, Kranz, Kramer, Howard, & Howard, 2005). In the original study, implicit sequence learning and contextual cueing were examined in college-age video-game players and piano players. The video-game players reported playing fast-action video games for at least four hours a week. The piano players reported that they had at least four years of formal music training, had played for at least 10 hours in the last month and were able to read sheet music. We contacted all of the original participants for whom we had contact information and 12 (six video-game players and six piano players) of the 36 motor skill experts contacted agreed to return to the lab for the present study. The mean retention interval was 420 days (342 – 489 days). There was no difference in implicit learning or in standardized neuropsychological scores at Time 1 between the people who returned and those who did not. None of the participants were told at the initial experiment that they would be asked to return for a follow-up study. Mean age, education, and standardized scores are reported in Table 1.
Table 1.
Participant Mean (and Standard Deviation) Characteristics at Time 2
| Experiment 1: Young adults |
Experiment 2: Older adults |
||
|---|---|---|---|
| Video/piano players | Piano players | Control | |
| Gender | 5 F/7 M | 2 F/4 M | 5 F/3 M |
| Age (in years) | 19.80 (1.22) | 73.50 (7.04) | 75.50 (6.66) |
| Education (in years) | 12.92 (0.99) | 16.33 (2.88) | 15.50 (2.93) |
| Self-rated health1 | 4.33 (0.65) | 4.17 (0.98) | 3.86 (1.07) |
| WAIS-III2 vocabulary* | 36.58 (7.68) | 56.33 (1.63) | 46.63 (12.88) |
| WAIS-III digit symbol* | 80.92 (13.99) | 73.17 (10.44) | 63.00 (12.01) |
| WAIS-III digit span* | 18.25 (3.72) | 20.83 (2.23) | 15 (4.14) |
| WMS-III3 spatial span | 17.33 (4.74) | 13.33 (1.63) | 11.38 (4.44) |
Piano Players > Controls, p < .01 for Experiment 2.
Self-rated health responses ranged from 1 (poor) to 5 (excellent).
WAIS-III = Wechsler Adult Intelligence Scale, 3rd Edition.
WMS-III = Wechsler Memory Scale, 3rd Edition.
Procedure
The same testing procedure was used at Time 2 as at Time 1. After signing an informed consent form approved by the Catholic University of America IRB, participants completed the ASRTT and several standardized neuropsychological tasks.
The ASRTT was performed on an iMac computer with a 38 cm monitor. The stimuli, apparatus, and procedure were similar to those used in previous studies (e.g., J. H. Howard, Jr. & Howard, 1997). Participants were instructed to place their middle and index fingers of each hand on the keys marked “z”, “x”, “.”, and “/”, respectively. The keys corresponded to four equally-spaced circles on the computer screen. On each trial, one circle became black and remained so until the participant pressed the key corresponding to this target. After a delay of 120 ms, the next target appeared. At both Time 1 and Time 2, participants completed 20 blocks, each consisting of 10 random trials followed by 80 learning trials. These 80 trials consisted of an eight-element sequence, in which random trials alternated with pattern trials (e.g. 1r2r3r4r), that was repeated 10 times in each block. Thus, altogether, at each Time, participants responded to 200 repetitions of the pattern or 1600 trials. Participants took a brief break in between each block, and a longer break in between blocks 5 and 6 to complete the standardized neuropsychological tasks (see Table 1). Six patterns were counterbalanced across participants (1r2r3r4r, 1r2r4r3r, 1r3r2r4r, 1r3r4r2r, 1r4r2r3r, and 1r4r3r2r, where 1 – 4 indicate circle positions from left to right and r indicates a randomly selected event. Since the repeating pattern forms a continuous sequence, these six patterns include all permutations of the 4 alternating events.). The computer guided participants to an accuracy level of about 92% via an end-of-block visual display. If accuracy for a block was above 93%, the computer displayed “focus more on speed;” if accuracy was below 91%, the computer displayed “focus more on accuracy.”
Upon completion, participants were asked questions designed to probe for declarative knowledge as in previous studies with this task (D. V. Howard et al., 2004). The experimenter read the following questions aloud one at a time and recorded participants’ responses. (1) “Do you have anything to report regarding the task?” (2) “Did you notice anything special about the task or the materials?” (3) “Did you notice any regularity in the way the stimulus was moving on the screen?” If participants answered “yes” to question 3, they were asked (4) “Did you attempt to take advantage of any regularities you noticed in order to anticipate subsequent targets? If so, did this help?” (5) “In fact, there was some regularity to the sequences you responded to. What do you think it was? That is, try to describe any regularity you think there might have been.”
To further examine declarative knowledge after learning, participants performed a card-sorting task in which they sorted 7.6 cm by 12.7 cm white index cards that displayed three rows of four circles. Each line represented the four circles seen on the screen throughout the experiment. One circle on each line was black, so that each card represented three successive trials (a triplet). Participants placed each of the 64 cards into one of two piles, labeled “Frequent” and “Rare”, depending on how often they believed the triplet sequence had occurred during the ASRTT.
In addition, participants completed a computerized recognition task, in which they observed a sequence of circles turning black one at a time. They then rated the certainty of each sequence’s occurrence during the learning trials using a 4-point scale with 1 being certain the sequence did not occur and 4 being certain the sequence did occur. Participants performed 20 trials, each consisting of a 16-element sequence – two repetitions of an alternating eight-element pattern from a random starting point. Ten of these trials contained targets: pattern-consistent sequences that occurred in the experiment; ten contained foils: new sequences produced by reversing the sequence structure.
Following the ASRTT, participants completed a biographical and health screening questionnaire. Upon completion, participants were debriefed and told that they had responded to an alternating pattern. They were not told the exact pattern or that different participants received different patterns.
Results and Discussion
Data reduction
The 20 blocks were grouped into 10 epochs, each containing two blocks. The data from the first 10 random trials of each block were not analyzed. For the remaining 80 trials, each person’s sequence was parsed into a series of overlapping triplets using a sliding three-trial window. Because predictable (pattern) and unpredictable (random) events alternate, some successive runs of three events, or triplets, occur more often than others. For example, for the sequence 1r2r3r4r, the triplets 132 and 431 occur often, whereas the triplets 231 and 134 occur only rarely, ending only on random trials. We refer to the former as high-frequency triplets and the latter as low-frequency triplets and the third event of each as high- and low-frequency trials, respectively. Previous studies have shown that people learn triplet frequencies implicitly, responding more quickly and accurately to the third event of high- than low-frequency triplets with practice, despite the fact that people do not become aware of the alternating nature of the sequence (D. V. Howard et al., 2004).
Two kinds of low frequency triplets, repetitions and trills, were excluded from the analyses since, as is typical, people revealed pre-existing response tendencies when responding to these triplets (Soetens, Melis, & Notebaert, 2004). Repetitions are those triplets that repeat a single element (111, 222, 333 and 444), and trills are those that begin and end with the same element, with a different element in the middle (121, 343, 414, etc.). In general, from the outset, people responded exceptionally fast to repetitions and exceptionally slowly to trills. These initial response tendencies did not occur for other triplets.
Triplets that ended on pattern trials were always high frequency, and 25% of those that ended on random trials were high frequency by chance; thus, the 16 different high-frequency triplets made up 62.5% of the total triplets. The 32 low-frequency triplets (excluding the four repetitions and 12 trills described above) occurred 25% of the time. As a result, each high-frequency triplet occurred five times more often than each low-frequency triplet. People responded to each high-frequency triplet 62.4 times and each low-frequency triplet (excluding reps and trills) 12.6 times, on average, in each of the two sessions.
Is learning declarative?
Consistent with previous ASRTT studies (e.g., D. V. Howard et al., 2004), responses on the post-experimental questionnaire, as well as the card-sorting and recognition tasks, revealed that participants did not have declarative knowledge of patterns either after original learning at Time 1 or after the retention test at Time 2. When asked whether they noticed any regularity, none of the participants reported a pattern or described the alternating nature of the sequence. This occurred, even though during the Time 1 debriefing, participants had been told that there was a pattern.
For the card-sorting data, to determine whether individuals were able to explicitly judge the relative frequency with which each triplet occurred, the mean proportion of times the low-and high-frequency triplets were placed into the frequent category was calculated for each group at Time 1 and Time 2. See Table 2 for mean proportion of “frequent” responses and standard errors. A 2 × 2 × 2 analysis of variance (ANOVA) comparing these proportions for video-game players and piano players across the two sessions (Time 1 and 2) for each triplet type (high and low frequency) revealed no significant main effects of session or triplet type and no group × triplet type, triplet type × session, group × session, and group × triplet type × session interactions, indicating that the two groups sorted the high- and low-frequency triplets the same at both Time 1 and Time 2. However, there was a significant main effect of group, F (1, 10) = 25.93, p < 0.001, reffect = 0.59, which reflected the fact that piano players sorted more cards overall as “frequent.” Thus, consistent with the questionnaire responses, the card-sorting data confirm that participants in both groups were unable to explicitly distinguish low- and high-frequency triplets at either testing session.
Table 2.
Mean (and Standard Error) Proportion of Each Triplet Type Rated as Frequent for Card-Sort Data for Experiment 1
| Time 1 |
Time 2 |
|||
|---|---|---|---|---|
| High frequency | Low frequency | High frequency | Low frequency | |
| Video-game players | .55 (.07) | .53 (.05) | .52 (.06) | .55 (.03) |
| Piano players | .73 (.03) | .72 (.05) | .79 (.08) | .68 (.05) |
For the computerized recognition task, a mean familiarity rating was determined for target and foil sequences for each group. See Table 3 for mean ratings and standard errors. A 2 × 2 × 2 ANOVA revealed no significant main effects or interactions, indicating that participants in both groups rated the target and foil sequences as equally familiar at both sessions. These findings are consistent with the other declarative measures in revealing that participants in both groups were unable to explicitly judge how often specific sequences occurred. Thus, we conclude that participants did not acquire declarative knowledge of the patterns either at Time 1 or Time 2.
Table 3.
Participant Mean (and Standard Error) Familiar Ratings for Computerized Recognition Data for Experiment 1
| Time 1 |
Time 2 |
|||
|---|---|---|---|---|
| Target sequences | Foil sequences | Target sequences | Foil sequences | |
| Video-game players | 2.73 (.08) | 2.63 (.09) | 2.63 (.11) | 2.73 (.13) |
| Piano players | 2.67 (.13) | 2.88 (.05) | 2.83 (.09) | 2.65 (.17) |
Is there learning at time 1?
A median RT was determined separately for correct high-frequency and low-frequency triplets for each participant on each block. The median RTs were then averaged across blocks to obtain a mean of the median RT for each epoch. A similar procedure was used for accuracy. Preliminary 2 × 2 × 10 ANOVAs on the RT and accuracy data comparing the performance of video-game players and piano players for the two triplet types (high and low frequency) across Time 1 (10 epochs) revealed that although the piano players were marginally faster than the video-game players were overall (363 ms vs. 399 ms, respectively), F (1, 10) = 4.64, MSE = 16802, p = 0.06, reffect = 0.56, no interactions involving group were significant. It is possible that the small sample size and resulting low power made it difficult to detect small group effects (a power analysis revealed that all interactions with group were below 0.62). However, since the focus of the present study is on the retention of procedural skill rather than on differences between the two groups, the data were collapsed across groups as shown in the first 10 epochs plotted in Figure 1 for both RT (upper panel) and accuracy (lower panel).
Figure 1.
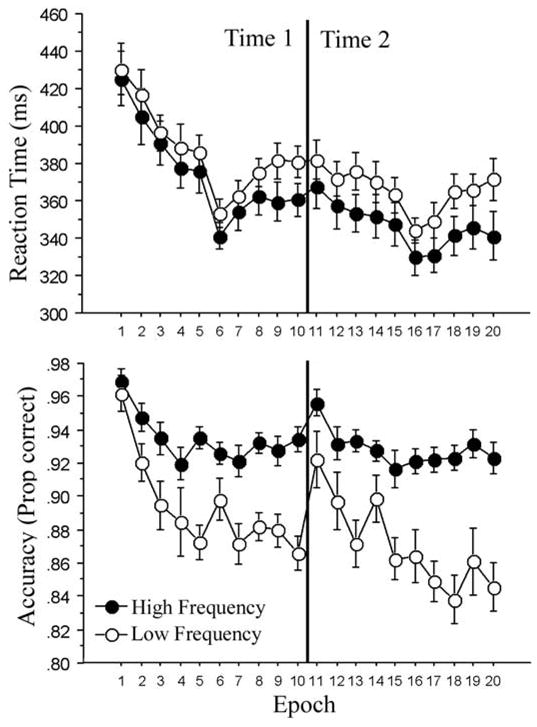
RTs for correct responses (upper panel) and accuracy (lower panel) as a function of epoch and trial type. Time 1: epochs 1–10; time 2 (1 year later): epochs 11–20. Each epoch consists of 20 blocks of 80 trials, for a total of 1600 trials per epoch. Participants rested for a few minutes in between each epoch, except for between epochs 5 and 6 and between epochs 14 and 15, in which they completed the neuropsychological tasks displayed in Table 1. The dips between epochs 5 and 6 and between epochs 14 and 15 reflect a consolidation that occurred during the break. This pattern is typical in ASRTT studies (cf. Howard and Howard, 1997; 2004). Experiment 1.
A 2 × 10 ANOVA on the combined Time 1 data revealed a significant main effect of epoch for RT, F (9, 90) = 31.10, MSE = 452, p < 0.0001, reffect = 0.56, such that people became faster overall with practice, reflecting general skill learning. The main effect of triplet type, F (1, 10) = 23.87, MSE = 352, p < 0.001, reffect = 0.84, and the triplet type × epoch interaction, F (9, 90) = 2.05, MSE = 92, p < 0.05, reffect = 0.41, were also significant, revealing sequence-specific learning: people responded 12 ms faster on high-frequency trials than they did on low-frequency trials overall (375 vs. 387 ms, respectively), and this difference was substantially greater by the end of training on Epoch 10 (20 ms) than it was on Epoch 1 (5 ms).
The accuracy analysis also produced significant main effects of epoch, F (9, 90) = 7.51, MSE = 0.001, p < 0.0001, reffect = 0.66, and triplet type, F (1, 10) = 84.33, MSE = .001, p < 0.0001, reffect = 0.95, as well as a significant epoch × triplet type interaction, F (9, 90) = 2.68, MSE = 0.001, p < 0.01, reffect = 0.46. Overall accuracy declined with practice from an initial 96% in Epoch 1 to 90% in Epoch 10, likely influenced in part by the end-of-block feedback that encouraged overall responding at 92% correct. Sequence-specific learning was reflected in a greater decline in accuracy on low- than high-frequency triplets with practice, such that people learned the sequence structure and tried to force the unstructured items into the structure with more practice, which led to more errors on the random trials completing low-frequency triplets. This pattern is typical of probabilistic sequence learning in which people make increasingly more errors on the unpredictable trials as they learn the sequence structure (D. V. Howard et al., 2004; Schvaneveldt & Gomez, 1998).
Thus, there was clear evidence at Time 1 of both general skill learning, in that people responded more quickly with practice, and sequence-specific skill learning, in that high-frequency trials were faster and more accurate than low-frequency trials, and these trial-type effects increased with practice.
Is there retention of general skill?
To investigate whether general skill learning was preserved over the one-year retention period, we carried out two planned t-test comparisons, corrected using Bonferroni correction, on the RT data. We focused on RT since the overall accuracy was strongly influenced by the computer feedback to encourage responding at 92% accuracy. Figure 2 shows the mean RTs used in these comparisons. As is evident in Figure 2, overall RT was significantly faster at the start of Time 2 (epoch 11) than it was at the start of Time 1 (epoch 1), one year earlier (375 vs. 428 ms, respectively), t (11) = 7.74, p < 0.001, reffect = 0.51, but there was no difference in mean RT between the end of Time 1 (epoch 10) and the start of Time 2 (epoch 11) (371 vs. 375 ms, respectively), t (11) = 0.51, p = 0.621, reffect = 0.06 (power to detect the epoch 10 versus epoch 11 difference was 0.05, while the epoch 1 versus epoch 11 power was 0.36). These results demonstrate at least partial incidental retention of general procedural motor skill over a one-year period in which there was no opportunity to practice the task.
Figure 2.
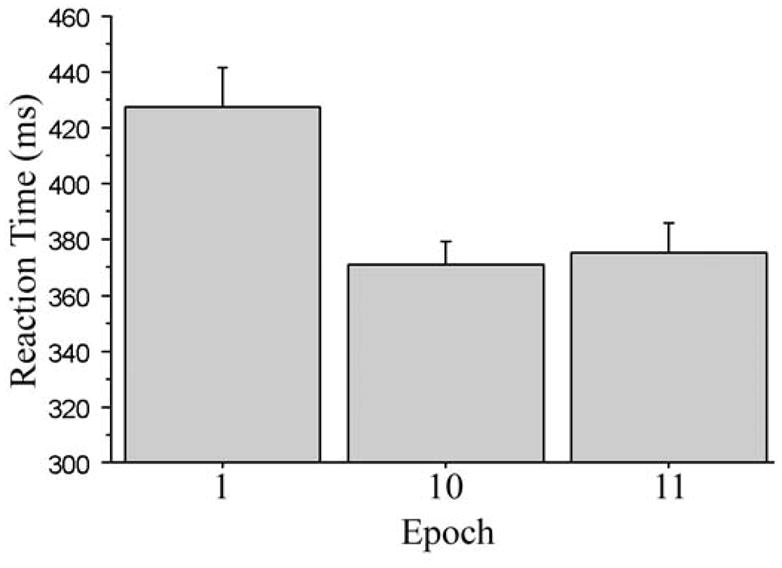
RT scores for the first and last epoch at time 1 and the first epoch at time 2. Experiment 1.
Is there retention of sequence-specific skill?
One could look for evidence of sequence-specific skill retention in various ways, and we consider a number of such methods here. Figure 3 depicts the mean estimate of learning, or trial-type effects, for RT (upper panel) and accuracy (lower panel) for the epochs used in the following planned comparisons (corrected using Bonferroni correction). Trial-type effects are defined as the difference between responses to the high and low frequency triplets (low – high for RT; high – low for accuracy).
Figure 3.
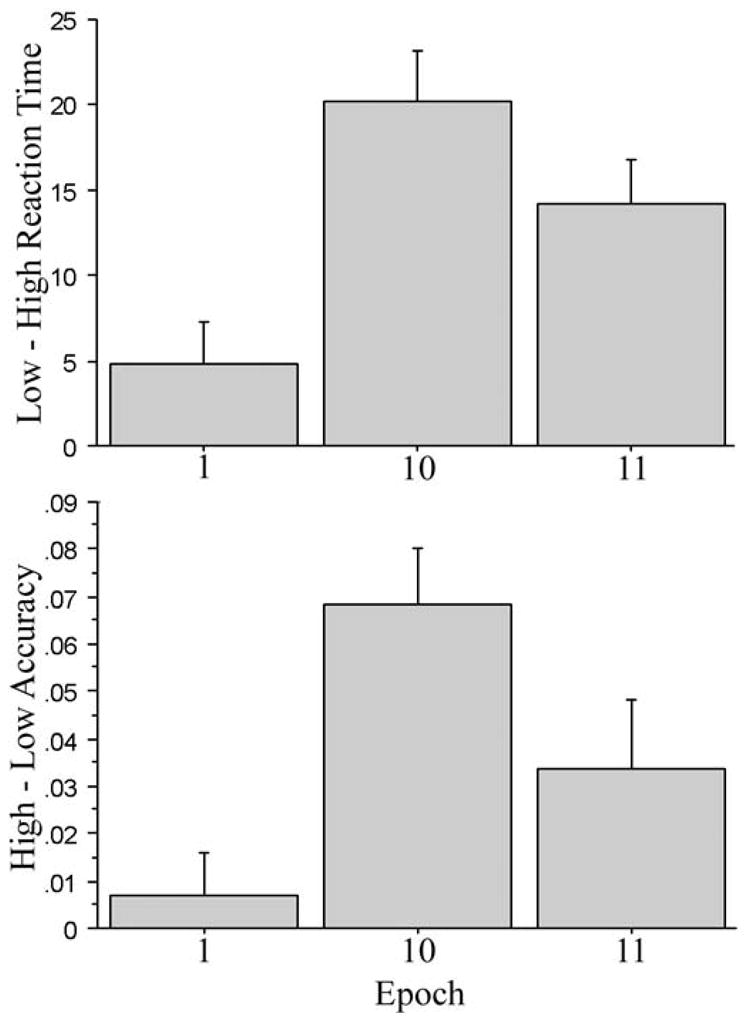
RT trial-type effects (upper panel) and accuracy trial-type effects (lower panel) for the first and last epoch at time 1 and the first epoch at time 2. Experiment 1.
One way to examine retention of sequence-specific skill is to compare the trial-type effects at the beginning of both testing sessions (Knopman, 1991). As may be seen in Figure 3, there were significantly greater trial-type effects at the start of Time 2 (epoch 11) than at the start of Time 1 (epoch 1) for both RT (14.1 ms vs. 4.8 ms, respectively), t (11) = 2.83, p < 0.02, reffect = 0.47, and accuracy (0.03 vs. 0.01, respectively), t (11) = 3.00, p = 0.01, reffect = 0.31 (power for the RT comparison = 0.32, while the accuracy comparison = 0.17). Prior research has referred to this as an “exposure effect” in that it indicates that the sequence-specific skill acquired at Time 1 carried over to the beginning of Time 2 (Knopman, 1991).
Retention can also be examined by comparing learning at the end of Time 1 to learning at the beginning of Time 2. If retention has occurred, there should not be a large difference between performance at the end of Time 1 and the beginning of Time 2. A comparison of trial-type effects for the final epoch of Time 1 (epoch 10) and the first epoch of Time 2 (epoch 11) revealed no significant difference for RT (20.2 ms vs. 14.1 ms, respectively), t (11) = 2.14, p = 0.06, reffect = 0.30, and no significant difference for accuracy (0.07 vs. 0.03, respectively), t (11) = 1.52, p = 0.16, reffect = 0.35 (power for the RT comparison = 0.16, while the accuracy comparison = 0.20). Thus, consistent with the significant “exposure effect” described above, the retention analyses indicate that participants retained sequence-specific skill over the one-year interval.
Experiment 2
Overall, the findings of Experiment 1 indicate that young motor-skill experts retained both general motor and sequence-specific skills in the ASRTT for at least one year. In addition, participants did not reveal declarative knowledge of the sequence structure on the card-sorting and recognition tests or in interviews at either Time 1 or Time 2. In the second experiment, we extend these findings to healthy adults, over the age of 65. Our labs have shown in earlier studies that while healthy older adults show general and sequence-specific skill learning in the ASRTT, they learn less than young adults and are slower overall (e.g., D. V. Howard et al., 2004, Howard & Howard, 1997). To date, there are no studies of long-term retention in older adults with the ASRTT, though, as mentioned above, two studies have shown general and sequence-specific skill retention in the traditional SRTT for two weeks in older controls (Knopman, 1991; Nissen et al., 1989). Studies using different skill-learning tasks have reported long-term retention in older adults despite age deficits in initial learning and overall performance. For example, two studies demonstrated two-year retention of a novel fine-motor skill (Smith, Walton, Loveland, Umberger, Kryscio, & Gash, 2005; Walton, Scheib, McLean, Zhang, & Grondin, 2008), and another study reported five-year retention of a mirror-tracing skill (Rodrigue, Kennedy, & Raz, 2005) in older adults. However, the continuous motor tasks investigated in these studies are quite different from the ASRTT and, in addition, none of these studies distinguished between retention of general and sequence-specific skills. In the second experiment, we investigate retention of general and sequence-specific procedural skills in the ASRTT with adults over the age of 65.
In addition to extending the investigation to healthy older adults, we also examined the role of motor-skill expertise on long-term retention. Previous studies have shown that musical and video-game expertise contributes to enhanced perceptual-motor ability in other visual perception and attention domains (see review by Green & Bavelier, 2008). To date, there are no studies of long-term skill retention in these expert groups, but it is possible that their expertise contributed to the long-term retention we observed in Experiment 1. Thus, we studied musicians, as well age-matched, non-musician controls in Experiment 2 to examine this possibility.
Method
Participants
Older individuals were recruited by advertisements in local newspapers for participation in the original ASRTT study that compared implicit learning in older piano players and controls (Romano, Burns, Kranz, Howard, & Howard, 2008). The piano players had been playing piano recreationally for an average of 67.61 years (range 60 – 77 years). All had some formal training, could read sheet music and currently played regularly. We contacted all of the original participants, and 14 (7 male; 6 piano players) of the 26 agreed to return for the present follow-up study. There was no difference in implicit learning at Time 1 between the participants who returned and those who did not. None of the participants had been told at the initial experiment that they would be asked to return for a follow-up portion of the study. The mean retention interval was 367 days (362 – 390 days).
Mean age, education and standardized cognitive scores for the participants at Time 2 are reported in Table 1. The piano players had greater digit span, faster processing speed (as measured by Digit Symbol Coding) and performed better on the vocabulary task, compared to controls. However, there was no difference in visual short-term or working memory (as measured by the WMS-III Spatial Span) between the piano players and controls at Time 2. The identical pattern was found in standardized cognitive scores at Time 1.
Procedure
The stimuli, apparatus, procedure, and data reduction were identical to those in Experiment 1.
Results and Discussion
Is learning declarative?
Responses on the post-experimental questionnaire, the card-sorting task, and the computerized recognition task provide consistent evidence that participants did not acquire declarative knowledge of the sequence structure, either after original learning at Time 1 or after the retention test at Time 2.
When asked whether they noticed any regularity, no one reported a pattern or described the alternating nature of the sequence, even though during the Time 1 debriefing, they had been told that there was a pattern. In fact, as also observed by Smith and colleagues (Smith et al., 2005), a number of our older participants did not remember that they had previously performed the task.
For the card-sorting task, as in Experiment 1, the mean proportion of times the low- and high-frequency triplets were placed into the frequent category was calculated for each group at Time 1 and Time 2. See Table 4 for proportion of “frequent” responses and standard errors. A 2 × 2 × 2 ANOVA comparing the performance of piano players to controls across the two sessions (Time 1 and 2) for both triplet types (high and low frequency) revealed no main effects or interactions, indicating that people in both groups sorted the rare and frequent triplets the same at both sessions.
Table 4.
Participant Mean (and Standard Error) Proportion of Each Triplet Type Rated as Frequent for Card-Sort Data for Experiment 2
| Time 1 |
Time 2 |
|||
|---|---|---|---|---|
| High frequency | Low frequency | High frequency | Low frequency | |
| Piano players | .66 (.04) | .61 (.03) | .42 (.09) | .47 (.08) |
| Controls | .58 (.10) | .56 (.06) | .58 (.05) | .54 (.05) |
For the computerized recognition task, a mean familiarity rating was determined for target and foil sequences for each group. See Table 5 for mean ratings and standard errors. A 2 × 2 × 2 ANOVA revealed no main effects and no interactions, indicating no difference between the groups in sequence-type ratings at either session. These results, consistent with questionnaire and card-sorting data, further demonstrate that participants did not acquire declarative knowledge of the sequence structure.
Table 5.
Participant Mean (and Standard Error) Familiar Ratings for Computerized Recognition Data for Experiment 2
| Time 1 |
Time 2 |
|||
|---|---|---|---|---|
| Target sequences | Foil sequences | Target sequences | Foil sequences | |
| Piano players | 2.55 (.12) | 2.75 (.09) | 2.85 (.19) | 2.88 (.14) |
| Controls | 2.73 (.12) | 2.71 (.07) | 2.51 (.14) | 2.64 (.07) |
Is there learning at Time 1?
Preliminary 2 × 2 × 10 ANOVAs comparing the performance of piano players and controls for the two triplet types (high and low frequency) across Time 1 (10 epochs) revealed no significant differences between the groups and no interactions involving group for either RT or accuracy. As in Experiment 1, the relatively low power resulting from the small sample size here may have made it difficult to detect small group effects (a power analysis revealed that all interactions with group were at or below 0.55). As in Experiment 1, for subsequent Time 1 analyses, the data were collapsed across groups as shown in Figure 4.
Figure 4.
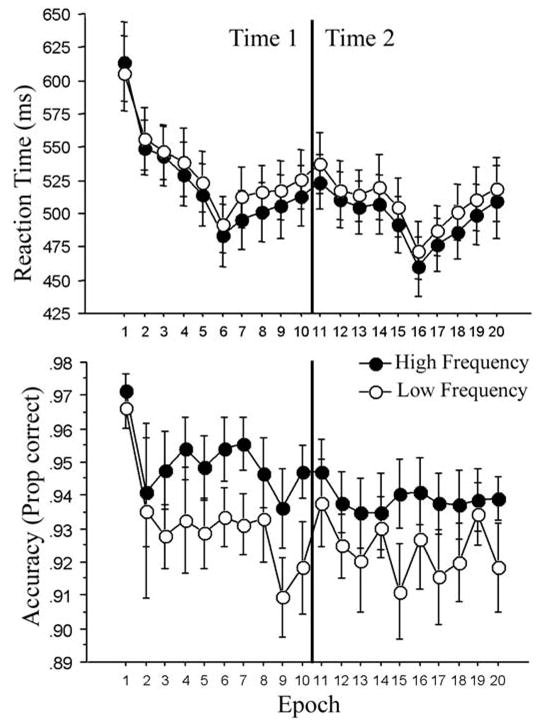
RTs for correct responses (upper panel) and accuracy (lower panel) as a function of epoch and trial type. Time 1: epochs 1–10; time 2 (1 year later): epochs 11–20. Each epoch consists of 20 blocks of 80 trials, for a total of 1600 trials per epoch. Participants rested for a few minutes in between each epoch, except for between epochs 5 and 6 and between epochs 14 and 15, in which they completed the neuropsychological tasks displayed in Table 1. The dips between epochs 5 and 6 and between epochs 14 and 15 reflect a consolidation that occurred during the break. This pattern is typical in ASRTT studies (see Figure 1). Experiment 2.
A 2 × 10 ANOVA on the combined Time 1 data revealed a significant main effect of epoch for RT, F (9, 108) = 18.93, MSE = 1606, p < 0.001, reffect = 0.78, such that people became faster overall, revealing general skill learning. The main effect of triplet type, F (1, 12) = 33.03, MSE = 143, p < 0.001, reffect = 0.86, and the triplet type x epoch interaction, F (9, 108) = 2.78, MSE = 128, p < 0.01, reffect = 0.43, were also significant for RT, reflecting sequence-specific learning: participants responded 9 ms faster on high-frequency trials than on low-frequency trials overall (525 ms vs. 534 ms, respectively), and this difference was greater by the end of training on Epoch 10 (13 ms) than it was at the beginning on Epoch 1 (8 ms, in the opposite direction, such that people began by responding faster to the low- than high-frequency trials).
The accuracy analysis also produced significant main effects of epoch, F (9, 108) = 2.86, MSE = .001, p < 0.01, reffect = 0.44, and triplet type, F (1, 12) = 32.17, MSE = 0.001, p = 0.0001, reffect = 0.85. However, the epoch × triplet type interaction did not reach significance for accuracy, F (9, 108) = 0.74, MSE = 0.001, p = 0.67, reffect = 0.24. Overall accuracy declined with practice from an initial 97% in Epoch 1 to 93% in Epoch 10, likely reflecting the feedback that encouraged responding at 92% as well as sequence-specific learning, as described above.
Thus, older adults, like young adults, showed both general skill learning and sequence-specific skill learning at Time 1 on both the RT and accuracy measures, in that high-frequency trials were faster and more accurate than were the low-frequency trials, and the RT trial-type effect increased with practice. This finding replicates previous findings with older adults (e.g., D. V. Howard et al., 2004, Howard & Howard, 1997).
Is there retention of general skill?
Although initial learning did not differ between the two groups, we wanted to examine whether or not expertise had an effect on retention. This was investigated as prior research has shown that motor-skill experts not only improve in perceptual, cognitive, and/or motor performance, but that their improvement is a result of lasting change, as opposed to differences due to short-term effects (Green & Bavelier, 2008). So for the following comparisons, we included group (piano players versus controls) as an independent variable.
First, we investigated whether or not there was retention of general skill learning. As in Experiment 1, we focused on RT, and Figure 5 shows the means used in the analysis. A group × epoch ANOVA produced a significant main effect of epoch, F (2, 24) = 17.86, MSE = 1794, p < 0.0001, reffect = 0.82, but neither the main effect of group nor the group × epoch interaction approached significance. The epoch effect was investigated further in two planned t-test comparisons, corrected using Bonferroni correction. As shown in Figure 5, like the young adults, overall RT was significantly faster at the start of Time 2 (epoch 11) than at the start of Time 1 (epoch 1), one year earlier, t (13) = 4.87, p < 0.001, reffect = 0.38 (power = 0.26). There also was no difference in RT between the end of Time 1 (epoch 10) and the start of Time 2 (epoch 11), t (13) = 1.09, p = 0.30, reffect = 0.07 (power = 0.06). Thus, these results parallel those of Experiment 1 with young adults in demonstrating at least partial incidental retention of general procedural skill over a period of one year for older adults.
Figure 5.
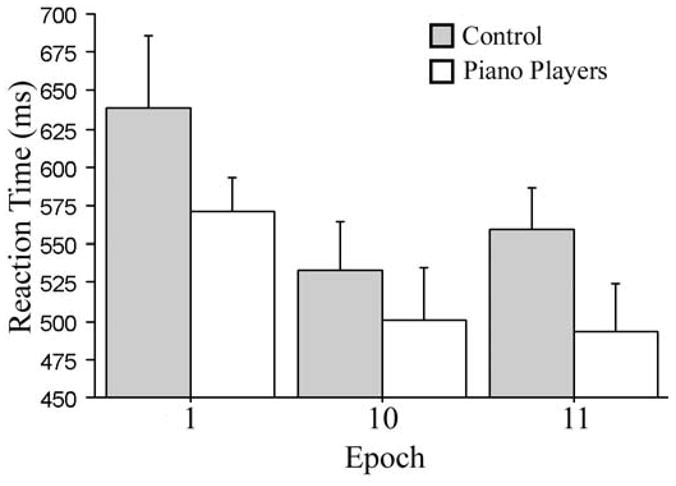
RT scores for the first and last epoch at time 1 and the first epoch at time 2 for piano players and controls. Experiment 2.
Is there retention of sequence-specific skill?
Figure 6 displays the trial-type effects for RT and accuracy (upper and lower panels, respectively) for the epochs used in the following analyses. A group × epoch ANOVA on the RT data revealed a significant main effect of epoch, F (2, 24) = 6.91, MSE = 285, p < 0.01, reffect = 0.78, but neither the main effect of group nor the group × epoch interaction reached significance.
Figure 6.
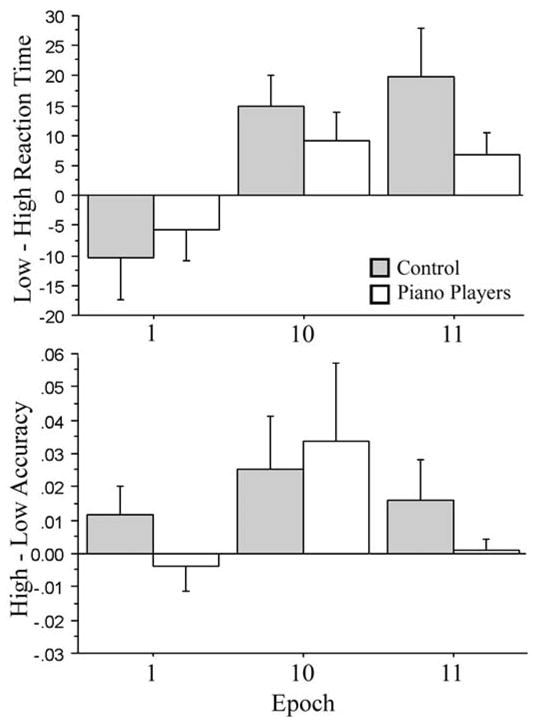
RT trial-type effects (upper panel) and accuracy trial-type effects (lower panel) for the first and last epoch at time 1 and the first epoch at time 2 for piano players and controls. Experiment 2.
The epoch effect was investigated further in two planned comparisons, corrected using Bonferroni correction to investigate retention. As seen in Figure 6, there was a significant “exposure effect” (Knopman, 1991) as revealed in a significantly greater RT trial-type effect at the start of Time 2 (epoch 11) than at the start of Time 1 (epoch 1), t (13) = 3.08, p < 0.01, reffect = 0.53 (power = 0.45). There was also clear evidence of retention as revealed in no significant difference between the RT trial-type effects at the end of Time 1 (epoch 10) and the start of Time 2 (epoch 11), t (13) = 0.35, p = 0.73, reffect = 0.05 (power = 0.05). Although the accuracy trial-type effects (lower panel of Figure 6) show a similar pattern to the RT effects, the accuracy effects are small, and no effects in the group × epoch ANOVA on accuracy reached significance.
Thus, the RT data reveal that older adults showed sequence-specific retention over one year. In addition, there were no significant differences in retention between the piano players and controls.
General Discussion
The present findings demonstrate that both general and sequence-specific skills acquired in the ASRTT can be retained for periods as long as one year. In addition, we have shown that long-term retention occurs in healthy older adults as well as in college-age adults and that these effects are not limited to motor-skill experts.
A caveat is that low statistical power may have played a role in our findings. Group differences were not detected in either experiment, and it is possible that group differences would have emerged with a larger sample. However, the focus of this study was on the retention of procedural skills rather than on group differences or the role of expertise per se. Evidence for retention involved two comparisons for each condition, one where retention would be revealed by a significant difference (Epoch 1 vs Epoch 11) and another where retention would be revealed by no difference (Epoch 10 vs Epoch 11). Our results followed this pattern suggesting that low power did not play an important role in these analyses.
In addition, some forgetting may have occurred over the one-year interval but was not detected with this small sample. A second caveat is that fatigue may have masked continued learning at Time 1, and what is seen as “retention of general motor skill” in this study could actually reflect a release of fatigue over the retention interval (Rickard, Cai, Rieth, Jones, & Ard, 2008). Indeed, participants in both experiments showed v-shaped learning curves as seen in Figures 1 and 4. This finding is common in ASRTT studies (cf. D.V. Howard & Howard, 2004) and may reflect the presence of performance fatigue that at least partially masked continued learning (Rickard et al, 2008). However, in this study, overall RT (Figures 2 and 5) was faster at the beginning of Time 2 than at the beginning of Time 1, consistent with the conclusion that at least some retention of general motor skill occurred.
Despite these concerns, our findings provide evidence that sequence-specific skill is retained over a one-year period. These findings are consistent with the ‘procedural reinstatement theory’ that posits that skills are well retained to the extent that the perceptual, motor and cognitive procedures underlying their learning are reinstated at test (Fendrich, Healy, & Bourne, 1991; Healy, Wohldmann, & Bourne, 2005; Marmie & Healy, 1995). Thus, one would expect high levels of retention in the present study as the identical task was used at test. However, in contrast, when Willingham and Dumas (1997) examined one-year retention of the SRTT in young adults, they found no sequence-specific retention, but they did find retention of general skill as measured by faster overall RT compared to a naïve control group. Some of the participants displayed declarative knowledge, but as in another study (Knopman, 1991), when these individuals were excluded, results remained the same. So although other studies (Knopman, 1991; Nissen et al., 1989) conclude that implicit sequence-specific learning is retained after one or two weeks, Willingham and Dumas (1997) conclude that it is not retained over a longer one-year interval, even when the original procedures were reinstated at retest. However, the present results indicate that implicit sequence-specific learning is retained for one year.
Willingham and Dumas (1997) outline several possible reasons why retention of sequence-specific knowledge was not revealed in their study, two of which are considered here. First, since a number of participants showed evidence of declarative knowledge, it is possible that learning the simple repeating pattern tapped into the declarative system which, although developing more quickly, lasts for a shorter time than procedural memory does (Tunney, 2003). Thus, declarative learning of the simple repeating pattern may have interfered with procedural learning (c.f., Poldrack & Rodriguez, 2004). However, since Willingham and Dumas (1997) found no sequence-specific retention, even when the individuals with declarative knowledge were removed, this cannot fully account for their null findings.
Second, a more likely explanation for the lack of retention that Willingham and Dumas (1997) consider is that their participants may not have had enough training for long-term retention to occur. In their experiment, individuals received only 40 repetitions of the pattern in the initial session. They point out that this is much less practice than had been used where long-term retention of implicit skill is observed, and it may not be sufficient for retention of sequence-specific knowledge after one year. In order to test this hypothesis, participants could receive more training. However, in the simple repeating SRTT, with extended practice, individuals become increasingly aware of the regularity and the explicit declarative system plays a larger role in learning. Even with only 40 pattern repetitions (480 trials), Willingham and Dumas (1997) reported that over half of the participants had some level of declarative knowledge, as assessed by post-learning free-recall and sequence-recognition tests. Therefore, simply extending practice does not allow unbiased evaluation of the long-term effect of practice in the SRTT context.
In the present study, using the ASRTT paradigm, participants in both of our experiments responded to a subtle pattern that repeated 200 times (1600 trials) during Time 1. Our findings are consistent with their conjecture that long-term retention of sequence-specific skill in the SRTT requires extensive practice prior to the retention interval. This possibility is consistent with evidence from other well-remembered procedural tasks that often require extensive practice to learn and remember (Karni & Sagi, 1993; Rodrigue et al., 2005; Savion-Lemieux & Penhune, 2005; Shadmehr & Brashers-Krug, 1997; Smith et al., 2005; Walton et al., 2008) as well as with the procedural skills we learn in everyday life, such as riding a bike or learning natural language grammar.
Although we cannot attribute long-term retention in the ASRTT specifically to extensive practice, we can rule out a number of possible alternative explanations for our findings. First, we can rule out the possibility that declarative rather than procedural knowledge underlies the long-term retention. Previous studies have shown that people do not spontaneously acquire declarative knowledge of the sequence regularity in the ASRTT (D. V. Howard et al., 2004) even with more extensive training than given here. In fact, it is difficult for people to gain declarative knowledge of the regularity even when they are given very specific prompts about its nature, such as presenting the pattern- and random-trial elements in different colors (Song, Howard, & Howard, 2007). In the present study, we used three very different tests and found no evidence of declarative knowledge.
Second, it is very unlikely that our findings simply reflect relearning at Time 2. In contrast to previous studies that have used the traditional SRTT in which learning is measured in random probe trials that occur only after many pattern trials (Knopman, 1991; Nissen et al., 1989; Willingham & Dumas, 1997), in the ASRTT, learning is assessed by comparing responses to pattern and random trials within the first few blocks at Time 2. Although it is likely that some additional learning did occur at Time 2, the retention effects we report cannot be attributed to relearning alone.
Third, we are confident that we did not simply recruit a biased sample of particularly strong learners at initial testing who were more likely to show long-term retention. As we reported in the methods section, the participants who returned in both experiments did not differ from those who did not, either in their initial learning or in their demographic or neuropsychological profiles.
Finally, it is not likely that people continued to practice the ASRTT during the retention interval. No one (including the experimenters) knew that they would be brought back a year later for testing. Even if participants moved their fingers to practice the general motor skill in the interim, the ASRTT task involves responding to stimuli and, without declarative knowledge of the sequence, people would not have known what to practice.
In conclusion, we have shown one-year retention of sequence-specific procedural skills in the ASRTT for older and younger adults as well as in motor-skill experts and controls. This finding is at odds with the results reported by Willingham and Dumas (1997) with the traditional SRTT, but consistent with theoretical accounts of the procedural memory system and the procedural reinstatement theory as well as with previous demonstrations of long-term motor skill retention using other tasks. Future studies should investigate the factors and neural mechanisms that contribute to such long-term retention of procedural skill.
Acknowledgments
This research was supported by grant R37AG15450 from the National Institute on Aging. We thank Eileen Kranz, Lindsey Kramer, Matthew Conetta, Andrew Kelly, Danielle Overpeck and Noel Burns for help with data collection. We thank Hadley Bergstrom, Betty Murphy, and Deborah Clawson for helpful comments. A preliminary report of these findings was presented at the May 2006 Association for Psychological Sciences Annual Convention.
Footnotes
We replicated all t-tests with non-parametric Wilcoxon signed-rank tests, and the results were the same as those reported in the manuscript.
Contributor Information
Jennifer C. Romano, Department of Psychology, The Catholic University of America
James H. Howard, Jr., Department of Psychology, The Catholic University of America & Department of Neurology, Georgetown University
Darlene V. Howard, Department of Psychology, Georgetown University
References
- Boyd LA, Vidoni ED, Daly JJ. Answering the call: The influence of neuroimaging and electrophysiological evidence on rehabilitation. Physical Therapy. 2007;87(6):684–703. doi: 10.2522/ptj.20060164. [DOI] [PubMed] [Google Scholar]
- Castel AD, Pratt J, Drummond E. The effects of action video game experience on the time course of inhibition of return and the efficiency of visual search. Acta Psychologica. 2005;119(2):217–230. doi: 10.1016/j.actpsy.2005.02.004. [DOI] [PubMed] [Google Scholar]
- Clark RE, Manns JR, Squire LR. Classical conditioning, awareness, and brain systems. Trends in Cognitive Science. 2002;6(12):524–531. doi: 10.1016/s1364-6613(02)02041-7. [DOI] [PubMed] [Google Scholar]
- Clawson DM, Healy AF, Ericsson KA, Bourne LE., Jr Retention and transfer of Morse code reception skill by novices: Part-whole training. Journal of Experimental Psychology: Applied. 2001;7:129–142. doi: 10.1037//1076-898x.7.2.129. [DOI] [PubMed] [Google Scholar]
- Cooke NJ, Durso FT, Schvaneveldt Retention of skilled search after nine years. Human Factors. 1994;36:597–605. [Google Scholar]
- Elbert T, Pantev C, Wienbruch C, Rockstroh B, Taub E. Increased cortical representation of the fingers of the left hand in string players. Science. 1995;270(5234):305–307. doi: 10.1126/science.270.5234.305. [DOI] [PubMed] [Google Scholar]
- Feeney JJ. Implicit learning and retention of serial patterns in middle aged adults: A dissertation. The Catholic University of America; Washington, DC: 2000. [Google Scholar]
- Fendrich DW, Healy AF, Bourne LE., Jr Long-term repetition effects for motoric and perceptual processes. Journal of Experimental Psychology: Learning, Memory and Cognition. 1991;17(1):137–151. doi: 10.1037//0278-7393.17.1.137. [DOI] [PubMed] [Google Scholar]
- Feng J, Spence I, Pratt J. Playing an action video game reduces gender differences in spatial cognition. Psychological Science. 2007;18(10):850–855. doi: 10.1111/j.1467-9280.2007.01990.x. [DOI] [PubMed] [Google Scholar]
- Forkstam C, Petersson KM. Towards an explicit account of implicit learning. Current Opinions in Neurology. 2005;18(4):435–441. doi: 10.1097/01.wco.0000171951.82995.c4. [DOI] [PubMed] [Google Scholar]
- Green CS, Bavelier D. Action video game modifies visual selective attention. Nature. 2003;423(6939):534–537. doi: 10.1038/nature01647. [DOI] [PubMed] [Google Scholar]
- Green CS, Bavelier D. Exercising your brain: A review of human brain plasticity and training-induced learning. Psychology and Aging. 2008;23(4):692–701. doi: 10.1037/a0014345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Healy AF, Fendrich DW, Proctor JD. Acquisition and retention of a letter-detection skill. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1990;16:270–281. doi: 10.1037//0278-7393.16.2.270. [DOI] [PubMed] [Google Scholar]
- Healy AF, Wohldmann EL, Bourne LE., Jr . The procedural reinstatement principle: Studies on training, retention, and transfer. In: Healy AF, editor. Experimental cognitive psychology and its applications. Washington, DC: APA; 2005. pp. 59–71. [Google Scholar]
- Healy AF, Wohldmann EL, Parker JT, Bourne LE., Jr Skill training, retention, and transfer: The effects of a concurrent secondary task. Memory & Cognition. 2005;33:1457–1471. doi: 10.3758/bf03193378. [DOI] [PubMed] [Google Scholar]
- Healy AF, Wohldmann EL, Sutton EM, Bourne LE., Jr Specificity effects in training and transfer of speeded responses. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2006;32:534–546. doi: 10.1037/0278-7393.32.3.534. [DOI] [PubMed] [Google Scholar]
- Hikosaka O, Rand MK, Nakamura K, Miyachi S, Kitaguchi K, Sakai K, et al. Long-term retention of motor skill in macaque monkeys and humans. Experimental Brain Research. 2002;147(4):494–504. doi: 10.1007/s00221-002-1258-7. [DOI] [PubMed] [Google Scholar]
- Howard DV, Howard JH., Jr Age differences in learning serial patterns: Direct versus indirect measures. Psychology and Aging. 1989;4(3):357–364. doi: 10.1037//0882-7974.4.3.357. [DOI] [PubMed] [Google Scholar]
- Howard DV, Howard JH., Jr Adult age differences in the rate of learning serial patterns: Evidence from direct and indirect tests. Psychology and Aging. 1992;7(2):232–241. doi: 10.1037//0882-7974.7.2.232. [DOI] [PubMed] [Google Scholar]
- Howard DV, Howard JH, Jr, Japikse K, DiYanni C, Thompson A, Somberg R. Implicit sequence learning: Effects of level of structure, adult age, and extended practice. Psychology and Aging. 2004;19(1):79–92. doi: 10.1037/0882-7974.19.1.79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howard JH, Jr, Howard DV. Age differences in implicit learning of higher order dependencies in serial patterns. Psychology and Aging. 1997;12(4):634–656. doi: 10.1037//0882-7974.12.4.634. [DOI] [PubMed] [Google Scholar]
- Karni A, Sagi D. The time course of learning a visual skill. Nature. 1993;365(6443):250–252. doi: 10.1038/365250a0. [DOI] [PubMed] [Google Scholar]
- Knopman D. Long-term retention of implicitly acquired learning in patients with alzheimer’s disease. Journal of Clinical and Experimental Neuropsychology. 1991;13(6):880–894. doi: 10.1080/01688639108405105. [DOI] [PubMed] [Google Scholar]
- Marmie WR, Healy AF. The long-term retention of a complex skill. In: Healy AF, Bourne LE Jr, editors. Learning and memory of knowledge and skills: Durability and specificity. Thousand Oaks, CA: Sage; 1995. pp. 30–65. [Google Scholar]
- Nissen MJ, Willingham D, Hartman M. Explicit and implicit remembering: When is learning preserved in amnesia? Neuropsychologia. 1989;27(3):341–352. doi: 10.1016/0028-3932(89)90023-7. [DOI] [PubMed] [Google Scholar]
- Poldrack RA, Packard MG. Competition among multiple memory systems: Converging evidence from animal and human brain studies. Neuropsychologia. 2003;41(3):245–251. doi: 10.1016/s0028-3932(02)00157-4. [DOI] [PubMed] [Google Scholar]
- Poldrack RA, Rodriguez P. How do memory systems interact? Evidence from human classification learning. Neurobiology of Learning and Memory. 2004;82(3):324–332. doi: 10.1016/j.nlm.2004.05.003. [DOI] [PubMed] [Google Scholar]
- Reber PJ, Squire LR. Encapsulation of implicit and explicit memory in sequence learning. Journal of Cognitive Neuroscience. 1998;10(2):248–263. doi: 10.1162/089892998562681. [DOI] [PubMed] [Google Scholar]
- Rickard TC, Cai DJ, Rieth CA, Jones J, Ard MC. Sleep does not enhance motor sequence learning. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2008;34:834–842. doi: 10.1037/0278-7393.34.4.834. [DOI] [PubMed] [Google Scholar]
- Robertson EM. The serial reaction time task: Implicit motor skill learning? Journal of Neuroscience. 2007;27(38):10073–10075. doi: 10.1523/JNEUROSCI.2747-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodrigue KM, Kennedy KM, Raz N. Aging and longitudinal change in perceptual-motor skill acquisition in healthy adults. Journal of Gerontology: B Psychological Sciences and Social Sciences. 2005;60(4):P174–181. doi: 10.1093/geronb/60.4.p174. [DOI] [PubMed] [Google Scholar]
- Romano JC, Burns NP, Kranz EM, Howard DV, Howard JH., Jr Old pianists have better working memory, speed of processing and vocabulary, but not implicit sequence learning. Paper presented at the Cognitive Aging Conference.2008. [Google Scholar]
- Romano JC, Kranz EM, Kramer LE, Howard DV, Howard JH., Jr Implicit sequence learning and spatial contextual cueing in experienced college-age musicians and video gamers. Paper presented at the Cognitive Neuroscience Society.2005. [Google Scholar]
- Savion-Lemieux T, Penhune VB. The effects of practice and delay on motor skill learning and retention. Experimental Brain Research. 2005;161(4):423–431. doi: 10.1007/s00221-004-2085-9. [DOI] [PubMed] [Google Scholar]
- Schendan HE, Searl MM, Melrose RJ, Stern CE. An fMRI study of the role of the medial temporal lobe in implicit and explicit sequence learning. Neuron. 2003;37:1013–1025. doi: 10.1016/s0896-6273(03)00123-5. [DOI] [PubMed] [Google Scholar]
- Schvaneveldt RW, Gomez RL. Attention and probabilistic sequence learning. Psychological Research. 1998;61(3):175–190. [Google Scholar]
- Shadmehr R, Brashers-Krug T. Functional stages in the formation of human long-term motor memory. Journal of Neuroscience. 1997;17(1):409–419. doi: 10.1523/JNEUROSCI.17-01-00409.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith CD, Walton A, Loveland AD, Umberger GH, Kryscio RJ, Gash DM. Memories that last in old age: Motor skill learning and memory preservation. Neurobiology of Aging. 2005;26(6):883–890. doi: 10.1016/j.neurobiolaging.2004.08.014. [DOI] [PubMed] [Google Scholar]
- Soetens E, Melis A, Notebaert W. Sequence learning and sequential effects. Psychological Research. 2004;69:124–137. doi: 10.1007/s00426-003-0163-4. [DOI] [PubMed] [Google Scholar]
- Song S, Howard JH, Jr, Howard DV. Implicit probabilistic sequence learning is independent of explicit awareness. Learning and Memory. 2007;14(3):167–176. doi: 10.1101/lm.437407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Squire LR, Clark RE, Bayley PJ. Medial temporal lobe function and memory. In: Gazzaniga M, editor. The Cognitive Neurosciences III. Cambridge, MA: MIT Press; 2004. pp. 691–705. [Google Scholar]
- Stefanidis D, Korndorffer JR, Jr, Sierra R, Touchard C, Dunne JB, Scott DJ. Skill retention following proficiency-based laparoscopic simulator training. Surgery. 2005;138(2):165–170. doi: 10.1016/j.surg.2005.06.002. [DOI] [PubMed] [Google Scholar]
- Tunney RJ. Implicit and explicit knowledge decay at different rates: A dissociation between priming and recognition in artificial grammar learning. Experimental Psychology. 2003;50(2):124–130. doi: 10.1026//1618-3169.50.2.124. [DOI] [PubMed] [Google Scholar]
- Walton A, Scheib JL, McLean S, Zhang Z, Grondin R. Motor memory preservation in aged monkeys mirrors that of aged humans on a similar task. Neurobiology of Aging. 2008;29(10):1556–1562. doi: 10.1016/j.neurobiolaging.2007.03.016. [DOI] [PubMed] [Google Scholar]
- Willingham DB, Dumas JA. Long-term retention of a motor skill: Implicit sequence knowledge is not retained after a one-year delay. Psychological Research. 1997;60:113–119. [Google Scholar]