

Abstract
When embryonic stem cells (ESCs) differentiate, they must both silence the ESC self-renewal program as well as activate new tissue specific programs. In the absence of DGCR8 (Dgcr8 -/-), a protein required for microRNA (miRNA) biogenesis, mouse ESCs are unable to silence self-renewal. Here, we find that the introduction of let-7 miRNAs, a family of miRNAs highly expressed in somatic cells, can suppress self-renewal in Dgcr8 -/-, but not wild-type ESCs. Introduction of ESC cell cycle regulating (ESCC) miRNAs into the Dgcr8 -/- ESCs, blocks the capacity of let-7 to suppress self-renewal. Profiling and bioinformatic analyses show that let-7 inhibits while ESCC miRNAs indirectly activate numerous self-renewal genes. Furthermore, inhibition of the let-7 family promotes de-differentiation of somatic cells to induced pluripotent stem (iPS) cells. Together, these findings show how the ESCC and let-7 miRNAs act through common pathways to alternatively stabilize the self-renewing versus differentiated cell fates.
Mammalian development follows a carefully orchestrated unfolding of cell fate transitions leading to a complex set of highly specialized cell types. These cell fate transitions involve the silencing of previously active molecular programs along with the activation of new ones. MiRNAs are small non-coding RNAs that are well suited to suppress previously active programs and, thereby, provide robustness to cell fate decisions1,2. MiRNAs identify their targets via base pairing of nucleotides 2-8 of the miRNA (the seed sequence) with complementary sequences within the target mRNA's open reading frame (ORF) and 3′ untranslated region (UTR)1. This targeting is carried out in coordination with the RNA-induced silencing complex (RISC) and often results in both destabilization and translational inhibition of the targets. While inhibition of any one target is usually only partial, each miRNA binds and suppresses hundreds of mRNA targets, resulting in large overall changes in the molecular constitution of cells.
Removal of genes required for maturation of all miRNAs has shown that miRNAs play essential roles in the proliferation and differentiation of ESCs3-5. For example, the loss of the RNA binding protein DGCR8, which is required for the production of all canonical miRNAs, results in a cell cycle defect and an inability to silence the self-renewal program of ESCs when they are placed in differentiation-inducing conditions3. The introduction of individual members of a family of miRNAs, the ESCC miRNAs, into Dgcr8 -/- ESCs can rescue the cell cycle defect6. These same miRNAs are able to enhance the de-differentiation of somatic cells to iPS cells7. Here, we report the identification of another large family of miRNAs, the let-7 family, which performs the opposite role to the ESCC family. When introduced into Dgcr8 -/- ESCs, let-7 silences self-renewal by suppressing many of the same downstream targets that are indirectly activated by the ESCC family. Indeed, co-introduction of the ESCC miRNAs inhibits the capacity of let-7 to silence self-renewal, and suppression of the let-7 family in somatic cells promotes de-differentiation.
Let-7 and ESCCs regulate self-renewal
The let-7 miRNAs are broadly expressed across differentiated tissues8,9 and are tightly regulated during ESC differentiation10-14. Therefore, we hypothesized that the let-7 miRNAs could rescue the capacity of Dgcr8 -/- ESCs to silence ESC self-renewal when induced to differentiate. To test this hypothesis we introduced mimics of a representative let-7 family member, let-7c, into the Dgcr8 -/- ESCs (Fig. 1a). Let-7c silenced the ESC self-renewal program even when the ESCs were maintained in ESC culture conditions. Three days after treatment with let-7c, Dgcr8 -/- cells downregulated ESC associated markers including alkaline phosphatase activity (Fig. S1, panel i), Pou5f1/Oct4 immunofluorescence staining (Fig. 1b, panel i), and mRNA expression of Pou5f1/Oct4, Sox2, and Nanog (Fig. 1c, panel i). Furthermore, the transfected cells showed a diminished capacity to reform ESC colonies in replating assays, a functional test of ESC self-renewal capacity (Fig. 1d, panel i). Similar effects were observed with the introduction of let-7a, let-7b, let-7d, and let-7g (Fig. S2) and these effects were observed over a range of concentrations, including levels normally found in more differentiated cell types (Fig. S3).
Figure 1.
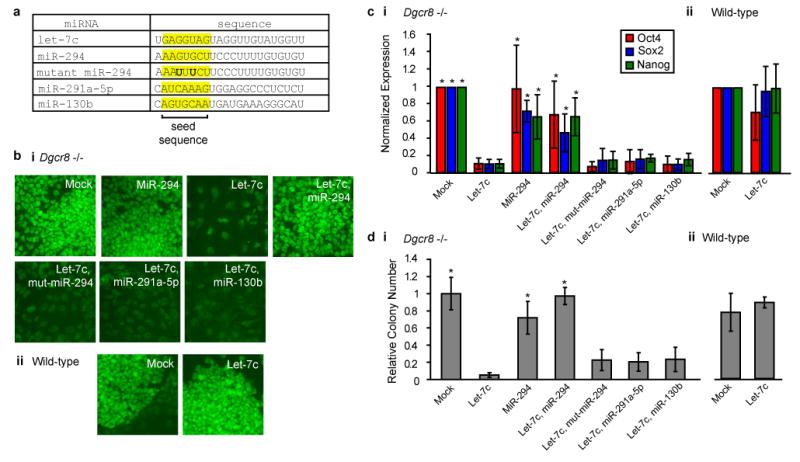
The let-7 and ESCC miRNA families have opposing roles in regulating ESC self-renewal. (a) Transfected miRNAs with the seed sequence highlighted. (b) Pou5f1/Oct4 immunofluorescence staining after transfection of let-7c, miR-294 and combinations of let-7c with miR-294, mutant-miR-294, miR-291a-5p, or miR-130b in Dgcr8 -/- (i) and wild-type (ii) ESCs. Representative images, n = 3. (c) qRT-PCR for Pou5f1/Oct4, Sox2, and Nanog normalized to beta-actin after miRNA introduction as in b. n = 3-8. * indicates p < 0.02. (d) Colony reforming assays after miRNA introduction as in b and c. n = 3. * indicates p < 0.05. All p-values generated by Bonferroni corrected t-test of comparisons to let-7c treated. Error bars represent standard deviation.
In contrast to the Dgcr8 -/- ESCs, wild-type ESCs were resistant to let-7c (Fig. S1, panel ii & 1b-d, panel ii). This finding suggested that other miRNAs normally expressed in wild-type ESCs inhibit let-7c-induced suppression of self-renewal. The ESCC miRNAs are likely candidates as they make up a majority of miRNA molecules in mouse ESCs15,16, they are rapidly downregulated upon differentiation coincident with the upregulation of mature let-7 (Fig. S4), and they promote the ESC fate6,7,17,18. Therefore, we introduced a representative member of this family, miR-294, to test if it could block let-7c-induced suppression of Dgcr8 -/- ESC self-renewal. Three days after co-introduction of miR-294 and let-7c, Dgcr8 -/- ESCs retained alkaline phosphatase activity (Fig. S1, panel i), Pou5f1/Oct4 immunofluorescence staining (Fig. 1b, panel i), and mRNA expression of Pou5f1/Oct4, Sox2, and Nanog (Fig. 1c, panel i). Furthermore, miR-294 rescued the colony forming capacity of the Dgcr8 -/- ESCs (Fig. 1d, panel i). Control miRNAs (miR-294 with a seed mutation and other ESC expressed miRNAs, miR-291a-5p and miR-130b, that do not contain the ESCC miRNA seed sequence) did not antagonize the effects of let-7c (Fig. 1a-d) showing that miR-294's effect is not simply secondary to competition for RISC complexes. Other members of the ESCC family miR-291a-3p, miR-291b-3p, and miR-295 were similarly able to block the effects of let-7c (Fig. S5). These data indicate that the let-7 and ESCC families of miRNAs have opposing roles in the maintenance of ESC self-renewal.
Targeting through ORFs and 3′UTRs
The functional antagonism between let-7c and miR-294 on ESC self-renewal suggested opposing roles for these miRNAs on downstream molecular targets. To test this prediction, we sought to globally identify these targets using mRNA microarrays following the introduction of let-7c or miR-294 into Dgcr8 -/- ESCs. The introduction of the let-7c mimic led to downregulation of 693 and upregulation of 208 transcripts relative to mock treated cells with a false discovery rate (FDR) less than 5% (Fig. 2a, Table S1). Of the 693 downregulated transcripts, 294 contained a let-7c 7mer seed match in the 3′UTR, 287 contained a 7mer seed match in the ORF, and 113 contained both 3′UTR and ORF seed matches (Table S1). The presence of these seed matches in the downregulated transcripts was highly enriched compared to the entire gene set (Fig. 2b, Fig. S6a). Similarly, the introduction of miR-294 led to a large number of upregulated and downregulated transcripts (Fig. 2c, Table S1). Again, downregulated transcripts were enriched for seed matches in the 3′UTR and ORF. In contrast, upregulated transcripts were depleted for seed matches in the 3′UTR and ORF (Fig. 2d, Fig. S6b). These findings suggest that miR-294 and let-7c functionally act through the downregulation of many targets by binding their ORF and/or 3′UTR.
Figure 2.

The let-7 and ESCC miRNAs suppress hundreds of transcripts by binding their ORF and/or 3′UTR. (a) Microarray analysis following introduction of let-7c alone. Upregulated transcripts are shown in dark grey, downregulated transcripts in black (FDR < 0.05). (b) Analysis of seed matches in the promoter, 5′UTR, ORF, and 3′UTR of let-7c-downregulated and upregulated transcripts. Presented are the mean number of seeds matches per kb of sequence for the listed groups of altered genes described in a. P-values calculated by the Wilcoxon Rank Sum Test and Bonferroni corrected are shown for p < 0.01 . (c) Microarray analysis following introduction of miR-294 alone. Color labeling, as in a. (d) Seed analysis as in b for miR-294 up and downregulated transcripts.
Impact on ESC transcriptional network
To further investigate the mechanism for the opposing roles of let-7c and miR-294 on ESC self-renewal, we performed pathway analysis on the miRNA regulated transcript sets. Specifically, we searched for overlaps between the miRNA-regulated transcripts and genes identified by chromatin immunoprecipitation (ChIP) of pluripotency associated transcription factors15,19. This analysis measures whether there is any influence of the let-7 or ESCC miRNAs on the transcription factors themselves (Fig. 3a, i&ii, & Supplementary Methods) or the transcripts originating from the genes bound by the transcription factors (Fig. 3a, iii, & Supplementary Methods).
Figure 3.

Enrichment/depletion of transcription factor bound genes among miRNA-regulated transcripts. (a) A schematic of hypothetical miRNA regulation of a transcription factor or its targets. Corresponding expected enrichment/ depletion of the transcription factor ChIP targets in miRNA-induced upregulated or downregulated transcript sets are displayed in a heat map. A key of color coding representing relative enrichment is given in b. (b) A heat map showing enrichment of the ChIP targets among the different sets of miRNA-regulated transcripts on the horizontal axis. Vertical axis represents the different ChIP data sets with first author and factor that was immunoprecipitated.
In ESCs, two Myc family members—nMyc and cMyc—are highly expressed and have largely overlapping ChIP target genes19. cMyc has previously been identified as a let-7 target in cancer cells20, and we find that nMyc is significantly downregulated by let-7c in our array data (Table S1). Consistent with let-7 directly targeting the Myc family, overlapping let-7c-regulated transcripts with Myc-bound genes showed an enrichment of Myc target genes in the let-7c-downregulated transcript set and a depletion in the let-7c-upregulated transcript set (Fig. 3b, Box I). Furthermore, the enrichment was independent of the presence of seed sequence matches within the ORF or 3′UTR. This finding suggests that let-7 is acting directly through Myc (cMyc and/or nMyc) rather than through Myc's downstream target genes (Fig 3a, i).
Performing a similar analysis overlapping miR-294-regulated transcripts and Myc target genes showed the exact opposite pattern as the analysis with let-7c-regulated transcripts. There was a depletion for Myc targets in the miR-294-downregulated transcript set and an enrichment in the miR-294-upregulated transcript set (Fig. 3b, Box II). This pattern suggests that miR-294 upregulates Myc activity (Fig 3a, ii). Indeed, microarray data showed that miR-294 dramatically increased cMyc levels (Table S1). As miR-294 itself suppresses its downstream targets (Fig 2d), the upregulation of cMyc must be indirect, through an unknown intermediate repressor (Fig. 3a, ii). These data show that the let-7 and ESCC families of miRNAs have opposing effects on Myc activity.
Overlap of the let-7c-regulated transcripts with ChIP target genes for the pluripotency transcription factors, Pou5f1/Oct4, Sox2, Nanog, and Tcf3 once again showed an enrichment among let-7c-downregulated transcript set (Fig. 3b, Box III). However, this enrichment was limited to the downregulated transcripts with seed matches in their ORF or 3′UTR. These data suggest that rather than directly regulating the pluripotency transcription factors, let-7 targets transcripts originating from the genes bound by them (Fig. 3a, iii). This pattern of enrichment is most clear for the ChIP target genes bound by Tcf3, cobound by Pou5f1/Oct4, Sox2, and Nanog, or bound by Chen et al.'s pluripotency cluster (a group of targets bound by Pou5f1/Oct4, Sox2, Nanog, Smad1, and STAT3). The latter results agree with recent reports showing that genes bound by multiple pluripotency transcription factors are more likely to be transcriptionally activated19,21. There was no enrichment in the overlap between the miR-294-regulated transcripts and Pou5f1/Oct4, Sox2, Nanog, and Tcf3 bound genes. These data suggest that let-7c inhibits downstream targets of these pluripotency factors while miR-294 has no obvious effects on either the transcription factors themselves or on their downstream targets.
Opposing regulation of Myc, Lin28, and Sall4
Having discovered that Myc activity was alternatively downregulated and upregulated by let-7c and miR-294, we sought to identify other factors that might be similarly regulated by these miRNAs. Indeed, gene ontology analysis showed an enrichment for ESC enriched genes among the let-7c-downregulated and miR-294-upregulated transcript sets (Fig. S7, Table S2). 88 transcripts were regulated in opposing directions by let-7c and miR-294, of which 44 contained a let-7c seed match (Fig. S8, Table S3). Notably, this set of transcripts included the well-known pluripotency genes Lin28 and Sall4. Lin28 is an RNA binding protein that inhibits let-7 processing10-13,22, but not transfected let-7 mimic (Fig. S9). Sall4 is a transcription factor that promotes ESC self-renewal23-25. These findings show that the let-7 and ESCC families antagonistically regulate multiple genes with described roles in ESC self-renewal.
To verify our genomic analysis, we performed qRT-PCR, Western analysis, and reporter assays for a subset of the genes. qRT-PCR confirmed the opposing effects of let-7c and miR-294 on Lin28, Sall4, nMyc, and cMyc mRNA levels with a combination of the two miRNAs showing intermediate levels (Fig. 4a). Western analysis showed similar results (Fig. 4b, Fig. S10). Of note, cMyc protein was dramatically reduced in Dgcr8 -/- versus wild-type ESCs and was brought back to wild-type levels with the introduction of miR-294. MiR-294 had little effect on nMyc levels. In contrast, let-7c had little effect on cMyc, yet dramatically reduced nMyc levels. Therefore, the cumulative effect of the miRNAs on total Myc (cMyc + nMyc) protein levels followed a strong pattern of opposing regulation. Similarly, the miRNAs showed significant opposing effects on Lin28 and Sall4 protein levels. Lin28 and cMyc are known targets of let-710,20, and luciferase assays confirmed that nMyc and Sall4 are also direct targets (Fig. 4c).
Figure 4.
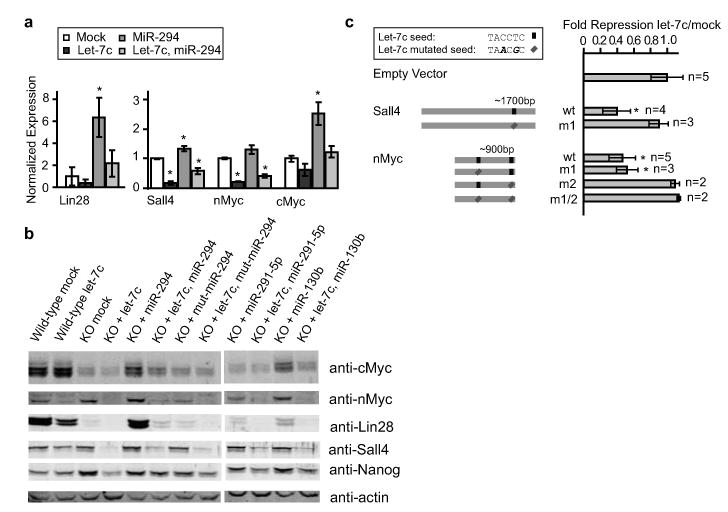
Let-7c and miR-294 regulate Lin28, Sall4, cMyc, and nMyc. (a) qRT-PCR for Lin28, Sall4, nMyc, and cMyc 12 hours after transfection with let-7c, miR-294, or a combination of the two. n = 3. (b) Representative Western blot analysis 48 hours after transfection with miRNAs. Quantitation shown in Fig. S10 n = 3. (c) Luciferase analysis of Sall4 and nMyc 3′UTRs. Seed matches for let-7c in the 3′UTRs along with different mutant constructs are diagrammatically represented in the left panel. Luciferase results after co-transfection with let-7c mimic releative to mock transfected are shown in the right panel. All data are represented as mean +/- standard deviation. * indicates p < 0.05 by Bonferroni corrected t-test.
Considering that cMyc was dramatically reduced in Dgcr8 -/- cells and then increased with miR-294, we considered the possibility that the loss of cMyc alone could largely explain the sensitivity of Dgcr8 -/- cells to let-7-induced silencing of ESC self-renewal. To test this possibility, we generated and evaluated cMyc -/- ESCs (Fig. S11). The loss of cMyc led to decreased expression of Pou5f1/Oct4 relative to the parental cell line (Fig. S12a). Introduction of let-7c into the cMyc -/- cells decreased the expression levels of Sox2 and Nanog (Fig. S12b&c). However, levels were not reduced to the same degree as seen with the introduction of let-7c into Dgcr8 -/- cells. These results indicate that the decrease of cMyc in Dgcr8 -/- cells alone cannot explain the sensitivity of these cells to let-7-induced silencing of ESC self-renewal.
Inhibition of let-7 promotes de-differentiation
Having identified a pro-differentiation function of the let-7 family of miRNAs, we hypothesized that inhibition of this miRNA family would enhance reprogramming of somatic cells to iPS cells. Indeed, Lin28, among other activities26-29, inhibits let-7 biogenesis10-13,22 and promotes de-differentiation of human somatic cells to iPS cells30. Reprogramming to iPS cells is typically achieved by the introduction of virally expressed Pou5f1/Oct4, Sox2, and Klf4 with or without Myc into somatic cells such as mouse embryonic fibroblasts (MEFs). While Myc dramatically increases the efficiency of reprogramming, it is not essential31,32. To test the impact of let-7 family on reprogramming, we used a let-7 antisense inhibitor. This inhibitor was able to suppress multiple let-7 family members simultaneously (Fig. S13).
MEFs express high levels of mature let-715 and, therefore, these cells should be responsive to any pro-reprogramming effects of let-7 downregulation. We used Oct4∷GFP transgenic MEFs in order to quantify changes in reprogramming efficiencies as Oct4∷GFP is activated late in the reprogramming process33,34. MEFs were transduced with retroviral vectors expressing Pou5f1/Oct4, Sox2, Klf4, with or without cMyc on day 0 as well as transfected with let-7 or a control inhibitor on days 0 and 6. When 3 transcription factors were used (minus cMyc), let-7 inhibition increased the number of GFP positive colonies on day 10 by 4.3 fold compared to mock whereas a control inhibitor had no significant effect (Fig. 5a, left panel). In the presence of all 4 transcription factors, let-7 inhibition resulted in a 1.75 fold increase (Fig. 5a, right panel). Immunofluorescence confirmed expression of Nanog in reprogrammed cells (Fig. S14). Furthermore, the resulting iPS cells expressed endogenous pluripotency markers at levels similar to wild-type ESCs and did not express the exogenously introduced factors (Fig. S15&S16), as expected for fully reprogrammed cells35. The impact of the let-7 inhibitor is not due to enhanced proliferation of the MEFs as there was actually a subtle decrease in proliferation following transfection of either the let-7 or control inhibitor (Fig. S17). These findings show that inhibition of let-7 family of miRNAs enhances the reprogramming of somatic cells. The finding that the enhancement was greater in absence of Myc is consistent with Myc activity being one, but not the only important downstream target of let-7 in stabilizing the somatic cell fate.
Figure 5.

Inhibition of let-7 miRNAs promotes reprogramming to induced pluripotency (a) Fold increase of Oct4∷GFP positive colonies in reprogramming with transduction of 3TFs (Pou5f1/Oct4, Sox2, and Klf4) or 4TFs (+ cMyc) after mock, let-7 inhibitor, or control inhibitor transfection. P-values are indicated for p < 0.01 calculated by Bonferroni corrected t-test. n = 10 for mock and let-7 inhibitor samples and n = 6 for control inhibitor samples (b) A model of the antagonism between the miR-294 and let-7c in the stabilization of the self-renewing and differentiated states. Bold and enlarged genes and arrows are active in the indicated state. Mechanisms of ESCC upregulation of Lin28 and cMyc are unknown and represented by a question mark.
Discussion
Our findings show that the let-7 and ESCC miRNA families have opposing effects on ESC self-renewal. We propose that they act in self-reinforcing loops to maintain the ESC self-renewing versus differentiated cell states (Fig. 5b). In the self-renewing state, ESCC miRNAs indirectly increase expression of Lin28 and cMyc. Lin28 functions to block the maturation of let-710-13. Therefore, the ESCC miRNAs prevent co-expression of let-7 miRNAs. Additionally, ESCC-induced upregulation of cMyc forms a positive feedback loop in which cMyc and nMyc, along with Pou5f1/Oct4, Sox2, and Nanog, bind and activate expression of the ESCC miRNAs in the miR-290 miRNA cluster7,15. As ESCs differentiate, Pou5f1/Oct4, Sox2, and Nanog are downregulated, resulting in the loss of ESCC and Lin28 expression. With the loss of Lin28, mature let-7 rapidly increases. This increase in let-7 is enhanced by a positive feedback loop in which let-7 suppresses its own negative regulator Lin28. In the differentiated state, downregulation of Myc activity by let-7 prevents co-expression of the ESCC miRNAs. Furthermore, let-7 inhibits downstream targets of Pou5f1/Oct4, Sox2, Nanog, and Tcf3 to stabilize the differentiated state. Sall4, like Myc and Lin28, is positively regulated by the ESCC family and negatively regulated by let-7 family. Decreases in Myc, Sall4, and Lin28 all promote ESC differentiation23,25,26,36.
In the model we propose, the function of let-7 in repressing the self-renewing state is restricted to cells that do not express high levels of ESCC miRNAs. In fact, our model suggests that let-7 and ESCC miRNAs are never co-expressed at high levels. For this reason, we propose that the let-7 family does not function to initiate differentiation, but rather the antagonism between the let-7 and ESCC families stabilizes the switch between self-renewal and differentiation. Consistent with this model, the introduction of either ESCC miRNAs7 or let-7 inhibitors into somatic cells promotes their de-differentiation into iPS cells. Additionally, the ESCC and let-7 miRNAs make up a preponderance of the miRNAs in self-renewing ESCs and somatic cells respectively15, supporting a major role in influencing these alternative cell fates.
Other miRNAs have been reported to target the ESC transcriptional network37-39. Unlike the let-7 family, these other miRNAs have a more limited tissue distribution8,9, suggesting that they may suppress self-renewal during differentiation along specific developmental pathways. Alternatively, these miRNAs may be involved in the early and transient stages of ESC differentiation while the let-7 miRNAs are involved in stabilizing the resulting differentiated cell fate. miRNAs related to the ESCC family (miR-17, miR-20, miR-93, and miR-106) and let-7 miRNAs play analogous roles in cancer with the ESCC related miRNAs promoting and the let-7 miRNAs inhibiting cancer growth40,41. It will be interesting to determine whether these miRNAs act through similar opposing pathways in cancer as in ESCs.
Methods Summary
Dgcr8 -/- and wild-type V6.5 ESCs were cultured as previously described3. miRNA mimics and inhibitors were obtained from ThermoFisher. mRNA profiling was performed on Affymetrix Mouse Gene 1.0 ST arrays. Bioinformatic analysis was performed using significance analysis of microarrays (SAM), R packages, and custom Python scripts. Reprogramming with Oct4-GFP MEFs was performed as previously described7.
Methods
Tissue culture, transfection, and AP staining
ES cell lines and culture conditions were previously described3. ES cells were weaned off MEFs and maintained in MEF conditioned media. For ES cell differentiation assays 40,000 Dgcr8 -/- or 12,000 wild-type ES cells were plated in gelatinized 12 well plates (or half the number of cells were plated on 24 well plates) on day 0 in LIF media. On day 1, miRIDIAN miRNA mimics (Dharmacon, ThermoFisher) were transfected at a concentration of 50nM using Dharmafect1 (Dharmacon, ThermoFisher) following the manufacturer's protocol. Media was changed daily. On the third day after transfection, cells were either lysed in Trizol (Invitrogen) for qRT-PCR analysis or fixed in 4% PFA for AP staining. AP staining was performed per the manufacturer's instructions (Vector Labs). iPS lines were maintained in ES media + 15% knock-out serum on irradiated MEF feeders. Colony reformation assays were performed as previously described3. Briefly, cells were exposed to miRNA mimics for 3 days then trypsinized and counted. A defined number of cells were replated on MEFs to form colonies for 5-7 days. The efficiency of colony reformation was determined by counting the number of AP positive colonies divided by the number of cells plated. Neural progenitor cells used in Fig. S3 were generated by in vitro differentiation of ES cells as described previously42.
Animal Use
All animal experiments described in this article have been approved by UCSF's Institutional Animal Care and Use Committee.
ES Cell Derivation
Timed matings where set up for cMyc f/f mice43. ES cells were derived from embryos isolated at E3.5. These embryos were cultured on an irradiated MEF feeder layer in ES cell media supplemented with 50uM PD9805944 and disassociated onto fresh feeders. ES cells were PCR genotyped as previously described43. A flox/flox line was grown out, infected with Ad5 Cre-IRES-GFP virus, sorted by FACs, and plated back onto MEF feeders. cMyc -/- colonies were grown out and verified by PCR genotyping and Western.
mRNA arrays
qRT-PCR showed that mRNA levels of a known let-7 target, Lin2810, was maximally reduced 12 hours post-transfection prior to a large decline in Oct4 and Nanog (Fig. S18). Therefore, we chose twelve hours for all microarray analysis to minimize secondary effects of let-7c-induced differentiation. 150,000 cells were plated in a 3.5cm dish on day 0. miRIDIAN miRNA mimics (Dharmacon, ThermoFisher) were transfected at a concentration of 50nM in media in the absence of LIF. At 12 hours post transfection cells were lysed in Trizol (Invitrogen) and RNA was prepared according to the manufacturer's protocol. Affymetrix Mouse Gene 1.0 ST arrays were probed by the Gladstone Genomics Core (www.gladstone.ucsf.edu/gladstone/site/genomicscore). Three biological samples were assayed for each treatment. Data were analyzed by Affymetrix Expression Console software. The Robust Multichip Analysis (RMA) algorithm was used to normalize the array signal across chips. SAM (http://www-stat.stanford.edu/∼tibs/SAM/) was used to determine FDR cutoffs for significantly altered genes.
qRT-PCR analysis
RNA for all qRT-PCR analysis was prepared using Trizol (Invitrogen) and quantified on a Nanodrop Spectrophotometer (ThermoFisher). 500ng of RNA was DNAse treated using DNAseI amplification grade (Invitrogen). For qRT-PCR of mRNAs, DNAse treated samples were reverse transcribed using the Superscript™ III first-strand synthesis system for RT-PCR (Invitrogen). qPCR reactions on resulting cDNAs were performed on either an ABI Prism 7100 or ABI 7900HT (Applied Biosystems). Primers are listed in Table S4. For miRNAs, qRT-PCR was performed either by using TaqMan® miRNA assays (Applied Biosystems) or by polyadenylating the miRNAs and then using a modified oligodT reverse transcription primer as described previously45.
Lin28 and GFP Expression in 293T cells
Lin28 was cloned into an expression vector under the EF1alpha promoter and upstream of IRES Pac (puromycin resistance). A similarly constructed GFP expression construct was previously generated6. 293T cells were transfected with 5ug of each construct and selected with 0.6ug/mL puromycin for 12 days.
Luciferase Reporter Assays
Constructs were produced as follows. The nMyc and Sall4 3′ UTRs were amplified from ES cell cDNA and cloned into the NotI and XhoI sites in psiCheck™-2 vector (Promega). Mutant UTRs were generated by a two-step PCR strategy with overlapping mutated PCR primers. Products of two PCRs with mutations were used in a second PCR reaction to generate full-length mutated inserts that were cut and ligated into cut empty vector. For transfections, 8000 Dgcr8 -/- ES cells were plated in ES cell media in a 96 well plate pretreated with 0.2% gelatin. The next day, miRIDIAN miRNA mimics (Dharmacon, ThermoFisher) were transfected with Dharmafect1 (Dharmacon, ThermoFisher) following the manufacturer's protocol at a concentration of 100nM. Simultaneously, luciferase constructs were transfected into ES cells at a concentration of 200ng per well using FUGENE® 6 (Roche) transfection reagent following the manufacturers protocol. The following day, 14-18 hours later, cells were lysed and luciferase assays were performed using a Dual-Luciferase® Reporter Assay System (Promega) on a single automatic injection Mithras (Berthold technologies) luminometer following the manufacturer's protocol. Transfection of each construct was performed in triplicate in each assay. Ratios of Renilla luciferase readings to Firefly luciferase readings were averaged for each experiment. Replicates performed on separate days were mean centered with the common readings from the individual days.
Seed match analysis
Promoter (1000 base pairs from the transcriptional start), 5′ UTR, ORF, and 3′ UTRs for Ensembl Transcripts (mm9) and known genes (mm8) were downloaded separately from the UCSC Genome Browser Table Browser. Seed match analysis was performed on these transcripts using a custom Python script. 7mer seeds were defined as either 7mer-1A or 7mer-m846. Seed match results were mapped to Affymetrix IDs. A Python script was then implemented to eliminate redundant transcripts as transcripts often mapped >1:1 with Affymetrix IDs. The transcript with the most 7mer seed matches was chosen to produce a 1:1 transcript to Affymetrix ID mapping. This mapping was done separately for the promoters, 5′ UTRs, ORFs, and 3′ UTRs. In rare cases, duplicate Affymetrix IDs exist for the same gene. These were retained in our analyses. Microsoft Access (Microsoft) was used to generate list overlaps for analyses. P-values were calculated in Fig. 2b&d with the # of seed matches per kb of transcript using the Wilcoxon Rank Sum test in R. P-values were calculated in Fig. S6a&b using a binary 0 for no seed matches or 1 for a seed match using the hypergeometric distribution function in R.
ChIP target overlap analysis
ChIP targets were downloaded from the supplementary tables15,19. Scripts were written to convert provided transcript IDs to a non-redundant list of Affymetrix IDs. Microsoft Access (Microsoft) and custom Python scripts were used to perform comparisons between gene lists and ChIP gene target lists. ChIP data from Chen et al. was downloaded as an association score between any particular gene and the transcription factor of interest. These scores were used directly for enrichment. For the Oct4, Sox2, Nanog bound group from Chen et al. any score above 0 was counted as bound. For all data, enrichment for ChIP gene target sets in miRNA-regulated gene sets was performed relative to all genes analyzed to produce the miRNA-regulated gene sets (i.e. all genes with Affymetrix IDs mapping to coding transcripts). The enrichments for any given ChIP target set were median normalized with all the miRNA-regulated genes sets in Fig. 3b. We performed this normalization because both the ChIP targets of the transcription factors and the miRNA-regulated gene sets in our analysis are enriched for more highly expressed genes19. We get a similar pattern of results without this normalization although all comparisons appear more highly enriched due to the expression levels (data not shown). Unnormalized enrichment is defined as (Genes in overlap of miRNA altered group and ChIP group/All genes in miRNA altered group)/(All genes in ChIP group/All genes used in analysis to generate miRNA altered groups).
Our enrichment analysis could yield a number of possible outcomes dependent on whether the miRNA targeted the transcription factor directly versus targeted transcripts downstream of the transcription factor. The following outcomes are presented in Fig. 3a: (i) If a miRNA directly targets a specific transcriptional activator, this activator will be downregulated and thus its ChIP target genes will likewise tend to be downregulated. This will result in an enrichment of ChIP target genes within the miRNA's downregulated gene set independent of there being a seed match in these targets. Likewise, the ChIP target genes should be depleted in the miRNA's upregulated gene set (Fig. 3a, i). (ii) If a miRNA directly targets a transcriptional repressor, there would be the inverse outcome; that is, the ChIP target genes should be enriched in the miRNA's upregulated gene set and depleted in the miRNA's downregulated gene set regardless of seed match (Fig. 3a, ii). (iii) If a miRNA targets an activating transcription factor's downstream targets, but not the transcription factor itself, ChIP target genes would be enriched in the downregulated gene set with a seed match but not without a seed match. Furthermore, there should not be an enrichment in the upregulated transcripts (Fig. 3a, iii).
Gene Ontology
Stem cell associated genes (genes upregulated in ESCs relative to brain and bone marrow) were generated from data in Ramalho-Santos et al. 200247 and were downloaded as a list from MySigDB (http://www.broad.mit.edu/gsea/msigdb). Enrichment of these stem cell associated genes in miRNA altered gene sets was performed, and p-values were calculated by Fischer's exact test.
Immunohistochemistry
Cells were fixed with 4% paraformaldehyde and washed twice in 1×PBS with 0.1% Triton x-100 (PBT). PBT with 2% BSA and 1% goat-serum was used to block for one hour before addition of primary antibody against Oct4 (Santa Cruz, rabbit polyclonal, product # sc-9081) or Nanog (Calbiochem, rabbit polyclonal, product # sc-1000) which was incubated overnight at 4°C or at room temperature for approximately 2 hours. Cells were washed with PBT, blocked with PBT with 2% BSA and 10% goat-serum for 1 hour before addition of secondary antibodies (Invitrogen: Alexa Fluor 488 goat anti-rabbit IgG).
Western Blots
On day 0, approximately 200,000 Dgcr8 -/- or 50,000 wild-type ES cells were plated in a 6 well plate. The following day miRIDIAN miRNA mimics (Dharmacon, ThermoFisher) were transfected at a concentration of 50nM. Lysates were collected two days after transfection in EBC buffer (50mM Tris-HCl, pH 8.0, 120mM NaCl, 0.5% Nonidet P-40, 1mM EDTA) containing 1× protease inhibitor cocktail (Roche). Lysates were incubated at 4 degrees C for 45 minutes rocking then spun at 4 degrees and 15,000rpm in a table top centrifuge. Protein was quantified using a Bio-Rad protein assay (Bio-Rad). 30ug of protein was resolved on an 8% SDS-PAGE gel. Proteins were transferred to Immobilon-FL (Millipore) and processed for immunodetection. Blots were scanned on a Licor Odyssey Scanner (Licor). The actin antibody was used at a 1:1000 dilution (Sigma, mouse monoclonal clone AC-40, Cat# A4700), the cMyc antibody was used at 1:500 (Epitomics, N-term rabbit monoclonal, Cat# 1472-1), the nMyc antibody was used at 1:500 (Calbiochem, mouse monoclonal, Cat# OP13), the Nanog antibody was used at 1:1000 (Abcam, rabbit polyclonal, Cat# ab21603), the Sall4 antibody was used at 1:500 (Abcam, rabbit polyclonal, Cat# ab29112), the Lin28 antibody was used at 1:1000 (Abcam, rabbit polyclonal, Cat# ab46020). Secondary IR antibodies from Licor were used at 1:10,000. Data were exported from the Licor Odyssey as jpg and quantified using ImageJ software (NIH).
MEF Isolation
E13.5 embryos from Oct4∷GFP/Rosa-26∷β-galactosidase transgenic crosses were isolated by Caesarean section and washed in HBSS. Heads and visceral tissues were removed. Remaining tissue was washed in fresh HBSS, briefly rinsed with 70% ethanol, then submerged in 0.05mM trypsin / 1mM EDTA HBSS solution and incubated at 37°C for 10 minutes. Tissue was pipetted repeatedly to aid in tissue dissociation, then added to MEF media containing 10% FBS and plated (passage 0).
Retrovirus infection
The retroviral packaging vector pCL-ECO was transfected into 293T cells simultaneously with pMXs vectors containing Oct4, Sox2, Klf4, or cMyc cDNA (Addgene) using Fugene 6 (Roche)48. At 24 hours, the media was changed, and at 48 hours, the media was collected, filtered (0.45μM), and frozen in aliquots at -80°C. Retrovirus was never thawed more than once. To induce reprogramming, passage 3 Oct4-GFP, Rosa26-β-galactosidase/neo MEFs49 were plated on gelatin-coated 12-well plates at 12 thousand cells per well. Retrovirus-containing media was added 24 hours later (Day 0). Cells were transfected with 16nM microRNA inhibitors (Dharmacon, ThermoFisher, Cat# I-310106-04 for let-7 inhibitor & Cat# IN-001000-01-05 for control inhibitor). Cells in reprogramming assays were transfected on days 0 and 6 post-retroviral-infection. Media was changed daily. Media was replaced with ES media + 15% FBS + LIF on day 2, and ES media + 15% knock-out serum replacement (Invitrogen) + LIF on day 6. GFP+ colonies were counted on day 10. Individual iPS colonies were picked and expanded for analysis between days 10 and 15.
Supplementary Material
Acknowledgments
We would like to thank Dr. Miguel Ramalho-Santos, Dr. Susan Fisher, Dr. Marco Conti, and members of the Blelloch Laboratory for critical reading of the manuscript. We would like to thank Dr. Alex Amiet at ThermoFisher for let-7 and control inhibitors, Dr. Michael Bishop for cMyc floxed mice and Archana Shenoy for ESC derived NPC cultures. Additionally we would like to acknowledge Dr. Adam Olshen and Dr. Ritu Roy of the UCSF Cancer Center Biostatistics and Computational Biology Core for their helpful advice concerning our statistical methods. This work was supported by funds to RB from NIH (K08 NS48118 and R01 NS057221), California Institute of Regenerative Medicine (CIRM) (Seed Grant RS1-00161, New Faculty Award RN2-00906), the American Health Assistance Foundation (formerly Stem Cell Research Foundation), and the Pew Charitable Trust. CM and RJ are supported by NSF graduate research fellowships.
Footnotes
Supplementary Information: Supplementary Information is linked to the online version of the paper at www.nature.com/nature. Supplementary data include 18 Figures and 4 Tables.
Author Contributions: CM contributed to Fig 1-4, 5b, and S1-13 and S18. RJ contributed to Fig5a-c and Fig S13-17. CM, RJ, and RB conceived the experiments, analyzed the data, and wrote the manuscript.
Author Information: Our microarray profiling data can be found at the GEO database, ID number GSE18840.
Reprints and permissions information is available at npg.nature.com/reprintsandpermissions. The authors declare no competing financial interests.
Correspondence and requests for materials should be addressed to RB (blellochr@stemcell.ucsf.edu).
References
- 1.Babiarz JE, Blelloch R. Small RNAs – their biogenesis, regulation and function in embryonic stem cells. The Stem Cell Research Community, StemBook. (StemBook) 2009 doi: 10.3824/stembook.1.47.1. [DOI] [PubMed] [Google Scholar]
- 2.Hornstein E, Shomron N. Canalization of development by microRNAs. Nat Genet. 2006;38:S20–24. doi: 10.1038/ng1803. [DOI] [PubMed] [Google Scholar]
- 3.Wang Y, Medvid R, Melton C, Jaenisch R, Blelloch R. DGCR8 is essential for microRNA biogenesis and silencing of embryonic stem cell self-renewal. Nat Genet. 2007;39:380–5. doi: 10.1038/ng1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kanellopoulou C, et al. Dicer-deficient mouse embryonic stem cells are defective in differentiation and centromeric silencing. Genes Dev. 2005;19:489–501. doi: 10.1101/gad.1248505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Murchison EP, Partridge JF, Tam OH, Cheloufi S, Hannon GJ. Characterization of Dicer-deficient murine embryonic stem cells. Proc Natl Acad Sci U S A. 2005;102:12135–40. doi: 10.1073/pnas.0505479102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Wang Y, et al. Embryonic stem cell-specific microRNAs regulate the G1-S transition and promote rapid proliferation. Nat Genet. 2008;40:1478–83. doi: 10.1038/ng.250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Judson R, Babiarz JE, Venere M, Blelloch R. Embryonic stem cell specific microRNAs promote induced pluripotency. Nat Biotech. 2009 doi: 10.1038/nbt.1535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Landgraf P, et al. A Mammalian microRNA Expression Atlas Based on Small RNA Library Sequencing. Cell. 2007;129:1401–1414. doi: 10.1016/j.cell.2007.04.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chen C, et al. Defining embryonic stem cell identity using differentiation-related microRNAs and their potential targets. Mamm Genome. 2007;18:316–327. doi: 10.1007/s00335-007-9032-6. [DOI] [PubMed] [Google Scholar]
- 10.Rybak A, et al. A feedback loop comprising lin-28 and let-7 controls pre-let-7 maturation during neural stem-cell commitment. Nat Cell Biol. 2008;10:987–93. doi: 10.1038/ncb1759. [DOI] [PubMed] [Google Scholar]
- 11.Viswanathan SR, Daley GQ, Gregory RI. Selective blockade of microRNA processing by Lin28. Science. 2008;320:97–100. doi: 10.1126/science.1154040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Heo I, et al. Lin28 mediates the terminal uridylation of let-7 precursor MicroRNA. Mol Cell. 2008;32:276–84. doi: 10.1016/j.molcel.2008.09.014. [DOI] [PubMed] [Google Scholar]
- 13.Newman MA, Thomson JM, Hammond SM. Lin-28 interaction with the Let-7 precursor loop mediates regulated microRNA processing. RNA. 2008;14:1539–49. doi: 10.1261/rna.1155108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Thomson JM, et al. Extensive post-transcriptional regulation of microRNAs and its implications for cancer. Genes Dev. 2006;20:2202–7. doi: 10.1101/gad.1444406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Marson A, et al. Connecting microRNA genes to the core transcriptional regulatory circuitry of embryonic stem cells. Cell. 2008;134:521–33. doi: 10.1016/j.cell.2008.07.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Calabrese JM, Seila AC, Yeo GW, Sharp PA. RNA sequence analysis defines Dicer's role in mouse embryonic stem cells. Proc Natl Acad Sci U S A. 2007;104:18097–18102. doi: 10.1073/pnas.0709193104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Benetti R, et al. A mammalian microRNA cluster controls DNA methylation and telomere recombination via Rbl2-dependent regulation of DNA methyltransferases. Nat Struct Mol Biol. 2008;15(3):268–279. doi: 10.1038/nsmb.1399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sinkkonen L, et al. MicroRNAs control de novo DNA methylation through regulation of transcriptional repressors in mouse embryonic stem cells. Nat Struct Mol Biol. 2008;15(3):259–267. doi: 10.1038/nsmb.1391. [DOI] [PubMed] [Google Scholar]
- 19.Chen X, et al. Integration of external signaling pathways with the core transcriptional network in embryonic stem cells. Cell. 2008;133:1106–17. doi: 10.1016/j.cell.2008.04.043. [DOI] [PubMed] [Google Scholar]
- 20.Kumar MS, Lu J, Mercer KL, Golub TR, Jacks T. Impaired microRNA processing enhances cellular transformation and tumorigenesis. Nat Genet. 2007;39:673–677. doi: 10.1038/ng2003. [DOI] [PubMed] [Google Scholar]
- 21.Kim J, Chu J, Shen X, Wang J, Orkin SH. An extended transcriptional network for pluripotency of embryonic stem cells. Cell. 2008;132:1049–61. doi: 10.1016/j.cell.2008.02.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Piskounova E, et al. Determinants of microRNA processing inhibition by the developmentally regulated RNA-binding protein Lin28. J Biol Chem. 2008;283:21310–4. doi: 10.1074/jbc.C800108200. [DOI] [PubMed] [Google Scholar]
- 23.Lim CY, et al. Sall4 regulates distinct transcription circuitries in different blastocyst-derived stem cell lineages. Cell Stem Cell. 2008;3:543–54. doi: 10.1016/j.stem.2008.08.004. [DOI] [PubMed] [Google Scholar]
- 24.Wu Q, et al. Sall4 interacts with Nanog and co-occupies Nanog genomic sites in embryonic stem cells. J Biol Chem. 2006;281:24090–4. doi: 10.1074/jbc.C600122200. [DOI] [PubMed] [Google Scholar]
- 25.Zhang J, et al. Sall4 modulates embryonic stem cell pluripotency and early embryonic development by the transcriptional regulation of Pou5f1. Nat Cell Biol. 2006;8:1114–23. doi: 10.1038/ncb1481. [DOI] [PubMed] [Google Scholar]
- 26.Heo I, et al. TUT4 in concert with Lin28 suppresses microRNA biogenesis through pre-microRNA uridylation. Cell. 2009;138:696–708. doi: 10.1016/j.cell.2009.08.002. [DOI] [PubMed] [Google Scholar]
- 27.Xu B, Zhang K, Huang Y. Lin28 modulates cell growth and associates with a subset of cell cycle regulator mRNAs in mouse embryonic stem cells. RNA. 2009;15:357–361. doi: 10.1261/rna.1368009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jones MR, et al. Zcchc11-dependent uridylation of microRNA directs cytokine expression. Nat Cell Biol. 2009;11:1157–1163. doi: 10.1038/ncb1931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Polesskaya A, et al. Lin-28 binds IGF-2 mRNA and participates in skeletal myogenesis by increasing translation efficiency. Genes Dev. 2007;21:1125–1138. doi: 10.1101/gad.415007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Yu J, et al. Induced Pluripotent Stem Cell Lines Derived from Human Somatic Cells. Science. 2007;318:1917–1920. doi: 10.1126/science.1151526. [DOI] [PubMed] [Google Scholar]
- 31.Nakagawa M, et al. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat Biotechnol. 2008;26:101–106. doi: 10.1038/nbt1374. [DOI] [PubMed] [Google Scholar]
- 32.Wernig M, Meissner A, Cassady JP, Jaenisch R. c-Myc is dispensable for direct reprogramming of mouse fibroblasts. Cell Stem Cell. 2008;2:10–12. doi: 10.1016/j.stem.2007.12.001. [DOI] [PubMed] [Google Scholar]
- 33.Stadtfeld M, Maherali N, Breault DT, Hochedlinger K. Defining molecular cornerstones during fibroblast to iPS cell reprogramming in mouse. Cell Stem Cell. 2008;2:230–240. doi: 10.1016/j.stem.2008.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Brambrink T, et al. Sequential expression of pluripotency markers during direct reprogramming of mouse somatic cells. Cell Stem Cell. 2008;2:151–159. doi: 10.1016/j.stem.2008.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hochedlinger K, Plath K. Epigenetic reprogramming and induced pluripotency. Development. 2009;136(4):509–523. doi: 10.1242/dev.020867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cartwright P, et al. LIF/STAT3 controls ES cell self-renewal and pluripotency by a Myc-dependent mechanism. Development. 2005;132:885–96. doi: 10.1242/dev.01670. [DOI] [PubMed] [Google Scholar]
- 37.Tay YM, et al. MicroRNA-134 modulates the differentiation of mouse embryonic stem cells, where it causes post-transcriptional attenuation of Nanog and LRH1. Stem Cells. 2008;26:17–29. doi: 10.1634/stemcells.2007-0295. [DOI] [PubMed] [Google Scholar]
- 38.Tay Y, Zhang J, Thomson AM, Lim B, Rigoutsos I. MicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiation. Nature. 2008;455:1124–8. doi: 10.1038/nature07299. [DOI] [PubMed] [Google Scholar]
- 39.Xu N, Papagiannakopoulos T, Pan G, Thomson JA, Kosik KS. MicroRNA-145 regulates OCT4, SOX2, and KLF4 and represses pluripotency in human embryonic stem cells. Cell. 2009;137:647–658. doi: 10.1016/j.cell.2009.02.038. [DOI] [PubMed] [Google Scholar]
- 40.Mendell JT. miRiad Roles for the miR-17-92 Cluster in Development and Disease. Cell. 2008;133:217–222. doi: 10.1016/j.cell.2008.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bussing I, Slack FJ, Grobhans H. let-7 microRNAs in development, stem cells and cancer. Trends in Molecular Medicine. 2008;14:400–409. doi: 10.1016/j.molmed.2008.07.001. [DOI] [PubMed] [Google Scholar]
- 42.Okabe S, Forsberg-Nilsson K, Spiro AC, Segal M, McKay RD. Development of neuronal precursor cells and functional postmitotic neurons from embryonic stem cells in vitro. Mechanisms of Development. 1996;59:89–102. doi: 10.1016/0925-4773(96)00572-2. [DOI] [PubMed] [Google Scholar]
- 43.Trumpp A, et al. c-Myc regulates mammalian body size by controlling cell number but not cell size. Nature. 2001;414:768–773. doi: 10.1038/414768a. [DOI] [PubMed] [Google Scholar]
- 44.Buehr M, Smith A. Genesis of Embryonic Stem Cells. Philosophical Transactions: Biological Sciences. 2003;358:1397–1402. doi: 10.1098/rstb.2003.1327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Shi R, Chiang VL. Facile means for quantifying microRNA expression by real-time PCR. Biotechniques. 2005;39:519–25. doi: 10.2144/000112010. [DOI] [PubMed] [Google Scholar]
- 46.Lewis BP, Burge CB, Bartel DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell. 2005;120:15–20. doi: 10.1016/j.cell.2004.12.035. [DOI] [PubMed] [Google Scholar]
- 47.Ramalho-Santos M. & Yoon, S., Matsuzaki, Y., Mulligan, R. C., Melton, D. A. Stemness': transcriptional profiling of embryonic and adult stem cells. Science. 2002;298:597–600. doi: 10.1126/science.1072530. [DOI] [PubMed] [Google Scholar]
- 48.Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–676. doi: 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- 49.Blelloch R, Venere M, Yen J, Ramalho-Santos M. Generation of induced pluripotent stem cells in the absence of drug selection. Cell Stem Cell. 2007;1:245–247. doi: 10.1016/j.stem.2007.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.