

Abstract
Background
Support for the role of transmembrane and membrane-proximal domains of αIIbβ3 integrin in the maintenance of receptor low affinity comes from mutational studies showing that activating mutations can induce constitutive bi-directional transmembrane signaling.
Design and Methods
We report the functional characterization of a mutant αIIbβ3 integrin carrying the Leu718Pro mutation in the membrane-proximal region of the β3 cytoplasmic domain, identified in heterozygosis in a patient with a severe bleeding phenotype and defective platelet aggregation and adhesion.
Results
Transiently transfected cells expressed similar levels of normal and mutant αIIbβ3, but surface expression of mutant αvβ3 was reduced due to its retention in intracellular compartments. Cells stably expressing mutant αIIbβ3 showed constitutive binding to soluble multivalent ligands as well as spontaneous fibrinogen-dependent aggregation, but their response to DTT was markedly reduced. Fibrinogen-adherent cells exhibited a peculiar spreading phenotype with long protrusions. Immunofluorescence analysis revealed the formation of αIIbβ3 clusters underneath the entire cell body and the presence of atypical high-density patches of clustered αIIbβ3 containing encircled areas devoid of integrin that showed decreased affinity for the fluorescent lipid analog DiIC16 and were disrupted in cholesterol-depleted cells.
Conclusions
These findings are consistent with an important role of the membrane-proximal region of β3 in modulating αIIbβ3 clustering and lateral redistribution of membrane lipids. Since the β3 mutant was associated with a thrombasthenic phenotype in a patient carrying one normal β3 allele, these results support a dominant role of clustering in regulating integrin αIIbβ3 functions in vivo.
Introduction
Integrins function as transducers of extra- and intracellular signals, and integrin transmembrane domains are involved in these bi-directional signaling processes mediating the transmission of conformational rearrangements between the cytoplasmic and extracellular regions.1 The evidence from mutational studies suggests that a helical interface between α and β subunit transmembrane domains restrains the integrin in the resting state.2 In addition, hydrophobic and electrostatic contacts within the membrane-proximal helices of αIIb and β3 cytoplasmic tails also have a role in maintaining αIIbβ3 in a low affinity state.3,4 Full activation of integrin signaling requires both ligand occupancy and receptor clustering. Recent data suggest that integrin conformational changes and clustering are complementary and may be mechanistically linked,5 but the molecular base involved in the lateral association of integrins is a matter of debate.6,7
Glanzmann’s thrombasthenia is a rare autosomal recessive disorder characterized by a lifelong bleeding tendency due to absent or severely reduced platelet aggregation.8 The disease is caused by quantitative or qualitative defects in integrin αIIbβ3, the main platelet fibrinogen receptor. More than 100 mutations causing Glanzmann’s thrombasthenia have been found in ITGA2B and ITGB3 genes (http://sinaicentral.mssm.edu/intranet/research/glanzmann). Most frequently, αIIbβ3 expression on the platelet surface is absent or very low due to mutations resulting in reduced stability of the mRNA or defective post-translational processing. Mutations associated with near normal or low receptor expression have provided useful information on the molecular basis of receptor function.9,10 So far, five natural mutations have been reported in the cytoplasmic tail of αIIb or β3. Two of them, R995Q in αIIb11 and D723H in β3,12 disrupt a conserved salt bridge that stabilizes the interaction between the membrane-proximal regions of the α and β subunits.13 Three other mutations in the β3 cytoplasmic domain, R724STOP,14 S752P,15 and IVS14-3C>G,16 were found in homozygosis associated with variant forms of Glanzmann’s thrombasthenia caused by defective activation of αIIbβ3.
It is widely accepted that the plasma membrane is organized into liquid ordered (Lo) microdomains, sphingolipid-and sterol-enriched, which coexist with a more fluid or liquid disordered (Ld) phase having lower resistance to extraction with non-ionic detergents.17 Lo domains, named membrane rafts, compartmentalize cellular processes and can sometimes stabilize to form larger platforms through protein-protein and protein-lipid interactions.18–20 In living cell membranes, cholesterol depletion has been shown to induce macroscopic separation of Lo and Ld phases into large domains that are clearly visible with fluorescent dyes that partition differentially into the two phases.21
In the present work, we used transfected cell models to study the functional consequences of a new natural mutation, L718P, in the membrane-proximal region of the β3 cytoplasmic tail. L718P had been identified in heterozygosis as a de novo mutation in a patient with a severe thrombasthenia-like phenotype carrying one normal β3 allele.
The results revealed that the L718P mutation in the proximal-membrane region of β3 promotes abnormal integrin clustering and alters lipid order in the plasma membrane, exerting a dominant negative effect on integrin function.
Design and Methods
Case report
Written informed consent to the use of blood samples was obtained from the patient, her family members and controls in accordance with the Declaration of Helsinki. The patient is a Caucasian 43-year old female from Canary Island (Spain) with a phenotype of severe and frequent mucocutaneous hemorrhages which started when she was 12 months old, menorrhagia since the age of 14 years and two episodes of spontaneous intra-peritoneal bleeding. Analytical studies revealed moderate thrombocytopenia (127×109/L) and platelet anisocytosis. Platelet aggregation, investigated in platelet-rich plasma, was markedly reduced (10–20% of control) in response to 10 μM ADP, 10 μM epinephrine, 10 μg/mL collagen, and 1.25 mM arachidonic acid, but agglutination induced by 1.2 mg/mL ristocetin was normal. P-selectin and CD63 expression upon activation with 25 μM TRAP was 7% of control. A bone marrow examination was not performed in the patient. Exhaustive questioning did not reveal a significant bleeding tendency in the mother, one brother, or three sisters of the patient. The patient’s deceased father was also asymptomatic.
Flow cytometry analysis of surface receptors
Membrane receptor content was measured using the monoclonal antibodies 2BC1 (anti-αIIb) and H1AG11 (anti-β3),22 P2 (anti-αIIbβ3, Immunotech, Marseille, France), LM609 (anti-αvβ3, Chemicon, CA, USA), and AK2 (anti-GPIbα, Serotec, Oxford, UK).
Washed platelets (106/100 μL) or transfected cells (2.5×105/100 μL) were incubated with the specific monoclonal antibody for 30 min, washed, and treated with a 1:500 dilution of Alexa Fluor 488-anti-mouse IgG (Molecular Probes, Eugene, Oregon, USA) for 20 min. Samples were analyzed with a Coulter flow cytometer model EPICS XL.
Fibrinogen and PAC-1 binding assays
Purified human fibrinogen (Calbiochem) was labeled with fluorescein isothiocyanate (FITC) as previously described.23 FITC-conjugated PAC-1 monoclonal antibody was from BD Biosciences (San José, CA, USA).
Washed platelets were resuspended in Hepes buffer, pH 7.4 (10 mM HEPES, 136 mM NaCl, 2.7 mM KCl, 1 mM CaCl2, 1 mM MgCl2, 0.1% glucose) at a final concentration of approximately 5×107/mL. Aliquots of 100 μL were treated for 5 min with 100 μM ADP plus 1 mM epinephrine, 40 μM TRAP-6, or 10 nM PMA, in the presence of 50 μg/mL FITC-Fg or 0.5 μg of FITC-PAC-1. After 20 min, platelets were washed and analyzed by flow cytometry. Numerical data are calculated as a product of the percent of gated positive cells and the value of the mean channel of fluorescence intensity.
Chinese hamster ovary (CHO) cells stably expressing wild or mutant αIIbβ3 were resuspended in Hepes buffer at 4×106/mL and treated for 5 min with 1 mM RGDS or 25 mM DTT, and incubated with FITC-PAC-1 (5 μg/mL). When indicated, cells were first incubated with the monoclonal antibody 2BC1 at 4ºC for 30 min, washed, and labeled with Alexa Fluor 647-anti-mouse IgG. Cells were then washed, incubated with 80 μg/mL FITC-fibrinogen in the absence or presence of 40 mU/mL thrombin, washed again, and analyzed with a Coulter flow cytometer model FC500.
Sequence analysis
Screening for mutations was carried out by direct sequencing of PCR-amplified overlapping fragments of reversed transcribed αIIb and β3 mRNA. Genomic DNA was extracted from peripheral blood cells and the sequence of PCR amplification products containing exons and whole or flanking regions of introns were analyzed using standard procedures.
Transient and stable transfections
To generate the mutant β3-P718 construct, the Afl II-EcoR I cDNA fragment from the patient was exchanged for the corresponding segment in the wild β3-cDNA vector.22 K562 cells were transiently transfected with wild or mutant β3-cDNA plasmid alone or together with wild αIIb-cDNA vector, using the Amaxa nucleofection system (Koeln, Germany). With this procedure, cell viability and transfection efficiency were higher than 90%.
CHO cells were transfected with αIIb-cDNA and either normal or mutant β3-cDNA vectors using Lipofectamine reagent (Invitrogen, CA, USA). Transfected cells were grown in medium containing G418 for 3 weeks before being sorted twice for cells expressing high levels of αIIbβ3 using a FACS Vantage cell sorter (Becton Dickinson). Experiments were performed with cell populations expressing similar levels of wild or mutant αIIbβ3.
Western blot analysis
Western blot analysis of total αIIb and β3 platelet content was carried out using 1 μg/mL of mixed 2BC1 and H1AG11 monoclonal antibodies. Tyrosine phosphorylation of focal adhesion kinase (FAK) in stably transfected CHO cells was determined by western analysis using anti-FAK [pY576] (Biosource, Nivelles, Belgium).
Soluble fibrinogen-dependent aggregation of Chinese hamster ovary cell transfectants
Stably transfected CHO cells (1.2×106) in 0.5 mL of DMEM medium containing 1 mg/mL fibrinogen were plated on 24-well dishes blocked with 1% BSA and incubated for 15 min at 37ºC in the absence or presence of 15 mM DTT. Cell aggregates were examined and microphotographed using a phase-contrast microscope with x4 objective.
Cell and platelet spreading and immunofluorescence microscopy
Aliquots of 500 μL of stably transfected CHO cells (4×104/mL) or washed platelets (2×108/mL) were plated on glass coverslips coated with 50 μg/mL fibrinogen or 10% FCS in 24-well dishes. After different periods of time, they were fixed, blocked and per-meabilized with 0.5% Triton X-100 in PBS-0.5% BSA for 20 min at room temperature. The cells were then incubated with anti-αvβ3 (LM609), anti-αIIb (2BC1), anti-β3 (H1AG11), or anti-αIIbβ3 (P2) monoclonal antibodies, and/or with an anti-FAK[pY576] or anti-protein disulfide-isomerase (PDI) (Upstate, Lake Placid, NY) polyclonal antibodies in PBS containing 0.1% Triton X-100 and 10% normal goat serum for 1 h. After washing, they were incubated with Alexa Fluor 488-anti-mouse and/or Alexa Fluor 546-anti-rabbit antibodies and then washed as above. When indicated, Alexa Fluor 568 conjugated to phalloidin was added to simultaneously visualize integrins and the actin cytoskeleton. In some experiments, cells were incubated with 5 μM dialkylindocarbocyanine (DiIC16, AnaSpec, San José, CA, USA) at room temperature for 15 min and then thoroughly washed before mounting. The preparations were mounted on Mowiol-DABCO (Sigma) and visualized with a x63 objective using a Zeiss Axioplan fluorescence microscope (Göttingen, Germany) equipped with a cooled CCD camera (Leica DFC 350 FX), or the Leica TCS-SP2-AOBS confocal microscope system. To detect DilC16 labeling, cells were excited with a laser emitting at 543 nm, and a 560-nm long-pass filter was used for collecting emissions. For cholesterol depletion, cells were harvested and incubated in suspension for 15 min at 37ºC with 5 mM methyl-β-cyclodextrin (MβCD, Sigma) in the absence of serum, followed by plating on fibrinogen-coated coverslips.
Results
Platelet glycoprotein expression and functional analysis
The platelet content of αIIbβ3 was estimated by western and flow cytometry (Figure 1A–B). The total αIIbβ3 content in the patient was similar to that in the controls. However, the surface detection of platelet αIIbβ3 was found to be 78%, 69% or 47% of the control depending on whether antibodies recognizing β3, αIIb or αIIbβ3 epitopes were used, respectively. This analysis also revealed a 25% decrease of GPIbα and a reduced mean value of side scatter in the patient’s platelets.
Figure 1.

Platelet expression of αIIbβ3 and binding of fibrinogen to stimulated platelets. (A) Flow cytometry analysis of surface αIIbβ3 and GPIbα. Washed platelets were incubated with monoclonal antibodies against β3 (H1AG11), αIIb (2BC1), αIIbβ3 (P2) and GPIbα (AK2) as described in the Design and Methods section. (B) Western analysis of platelet αIIb and β3 content. (C) Flow cytometry analysis of fibrinogen binding. Washed platelets were stimulated for 5 min with 100 μM ADP plus 1 mM epinephrine, 40 μM TRAP-6, or 10 nM PMA in the presence of FITC-fibrinogen. Numbers in the figure panels represent the fluorescence values calculated as a product of the percent of gated positive cells and the value of the mean channel of fluorescence intensity. (D) Adhesion and spreading of platelets on immobilized fibrinogen. Washed platelets were seeded on fibrinogen-coated coverslips for 30 min at 37ºC and, then, fixed, labeled with anti-αIIbβ3 monoclonal antibody 2BC1, and analyzed with an epifluorescence microscope. Bars: 15 μm. All data are representative of determinations with platelets from two different extractions of blood samples.
As shown in Figure 1C, a small population of the patient’s platelets was found to bind soluble fibrinogen under basal conditions, but the binding response to ADP plus epinephrine, TRAP, and PMA was significantly reduced, concordant with the deficient aggregation. Binding to the activation-dependent PAC-1 antibody following agonist stimulation was also markedly decreased compared to control (values of 1.42, 0.36, and 1.42 versus 102, 14.4, and 333 in response to ADP plus epinephrine, TRAP, and PMA, respectively). In addition, when plated on fibrinogen, platelets showed size heterogeneity and a high proportion displayed impaired spreading and defective lamellipodia formation (Figure 1D).
Platelets from the other members of the family showed glycoprotein levels, soluble ligand binding and spreading on immobilized fibrinogen similar to control platelets.
Identification of the L718P mutation in β3
The defective platelet function and reduced surface detection of αIIbβ3 complex in the patient prompted the sequence analysis of the ITGA2B and ITGB3 genes. RT-PCR analysis revealed similar amounts of two different β3 transcripts: one normal, and the other carrying a novel non-synonymous single-nucleotide variation in exon 14, T2231C, which would change Leu to Pro at position 718 in the mature β3 chain (Figure 2A). The heterozygous status of the mutation was confirmed by analyzing the genomic DNA. The mutation was not found in the other family members and the carrier status of the IVS14+9C>T polymorphism was very useful to infer the absence of the T2231C mutation in the deceased father (Figure 2B). It was, therefore, concluded that T2231C started as a de novo mutation in the patient. The L718P mutation in β3 was not found in more than 50 DNA samples from control donors and healthy relatives of patients suffering from other platelet-related diseases.
Figure 2.
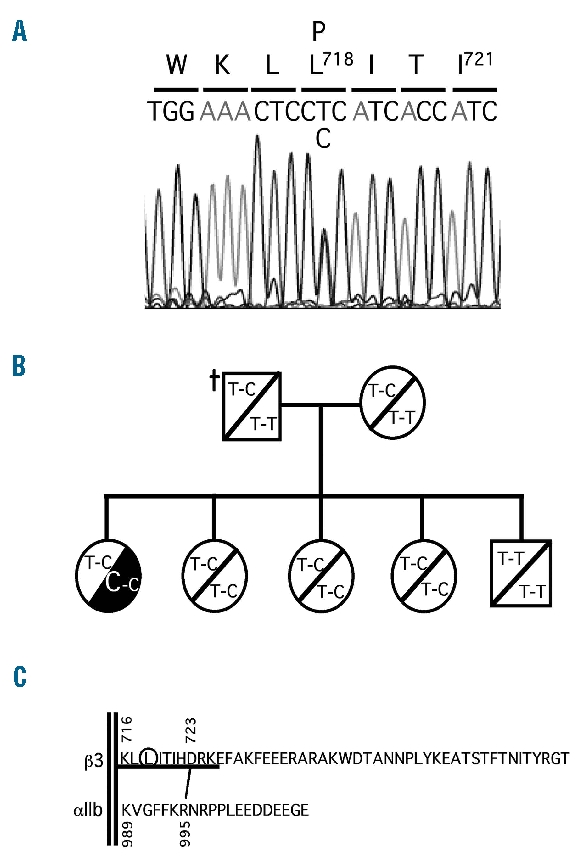
Identification of a de novo Leu718Pro mutation in β3. (A) Direct sequencing of the sense strand of the 3’ overlapping RT-PCR product showing a heterozygous T to C substitution that changes Leu718 to Pro in the β3 subunit of the thrombasthenic patient. (B) Detection of the T2231C mutation and the IVS14+9C>T polymorphism in family members. The haplotype of the deceased father was deduced from those of the other members. (C) Localization of the Leu718Pro mutation (encircled) in the membrane-proximal region (underlined) of the β3 cytoplasmic tail.
The analysis of αIIb-cDNA identified two homozygous polymorphic variations: I843S (HPA-3) in exon 26, and the synonymous transition c.3063C>T in the first codon of exon 30. Further sequencing of genomic DNA confirmed their homozygous status and revealed the presence of three other polymorphisms in intron 21: IVS21+36delCAGGGGCTC, IVS21-7C>G, and IVS21+254A>G (Online Supplementary Figure S1A). All five variations were found in heterozygosis in a control from the same geographical region. The first four variants have already been reported to be reciprocally linked and found in homozygosis in individuals with normal platelet aggregation responses.24 Exon-trapping analysis of a genomic DNA fragment containing the variant intron 21 indicated that it does not have a significant impact on αIIb gene expression at the transcription level. Both alleles generated similar amounts of transcripts, but a scarce additional RT-PCR product was obtained from the variant αIIb allele (Online Supplementary Figure S1B). Sequence analysis revealed that it contains aberrant αIIb transcripts with incorporation of variable length fragments of intron 22.
Surface expression of αIIbβ3-P718 and αvβ3-P718 mutants
Transient transfection of K562 cells showed that surface expression of αIIbβ3 complexes was slightly reduced in cells transfected with mutant β3-cDNA. However, expression of the β3-P718 mutant complexed to endogenous αv chains was markedly decreased, independently of the absence or presence of αIIb (Online Supplementary Figure S2A). Immunoprecipitation analysis of total αIIbβ3 and αvβ3 complexes suggested that the mutant β3 subunit is synthesized and forms dimers with αIIb at near normal rates, but that it prevents some of the steps involved in the maturation and traffic of the αvβ3 complex (Online Supplementary Figure S2B).
CHO cells stably expressing similar levels of wild or mutant αIIbβ3 complexes were obtained by successive cell sorting. Fluorescence microscopy analysis of adherent CHO cells showed that normal αvβ3 integrin detected with both H1AG11 and LM609 monoclonal antibodies was evenly dispersed within the plasma membrane, with some bright fluorescent clusters mainly at the cell periphery. In contrast, in cells expressing mutant β3-P718, the anti-β3 staining showed a heterogeneous reticular pattern that matches the endoplasmic reticulum distribution of protein disulfide isomerase. Furthermore, the immunofluorescence labeling of αvβ3 was strongly concentrated on a precise location, suggesting intracellular retention of this mutant integrin (Online Supplementary Figure S2C). The anti-β3 monoclonal antibody recognizes both β3 integrins, though the near normal surface expression of mutant αIIbβ3 integrin suggests that the retention pattern revealed by the anti-β3 monoclonal antibody corresponds basically to mutant αvβ3 integrin.
The αIIbβ3-P718 receptor is constitutively active
L718P is located in the membrane-proximal region of the β3 cytoplasmic domain (Figure 2C), which has a role in maintaining αIIbβ3 in a low affinity state.3 To explore whether the L718P substitution could render αIIbβ3 constitutively active, we estimated its ability to bind soluble lig-ands. A subpopulation (55%) corresponding to cells expressing the higher levels of the mutant integrin displayed significant spontaneous binding of the multivalent ligand-mimetic PAC-1 monoclonal antibody which was prevented in the presence of fibrinogen or the tetrapeptide RGDS. However, DTT-induced ligand binding was significantly reduced in cells expressing the mutant integrin (Figure 3A), suggesting that β3-P718 induced conformational changes altering the accessibility of the integrin disulfide bonds to the reducing agent. Constitutive binding of labeled fibrinogen and polymerizing fibrin was also detected in the subpopulation of cells with the highest expression levels of mutant receptor (Figure 3B). In agreement with these findings, CHO cells expressing mutant αIIbβ3 formed spontaneous cell aggregates in the presence of 1 mg/mL fibrinogen, but DTT failed to enhance cell aggregation (Figure 3C).
Figure 3.
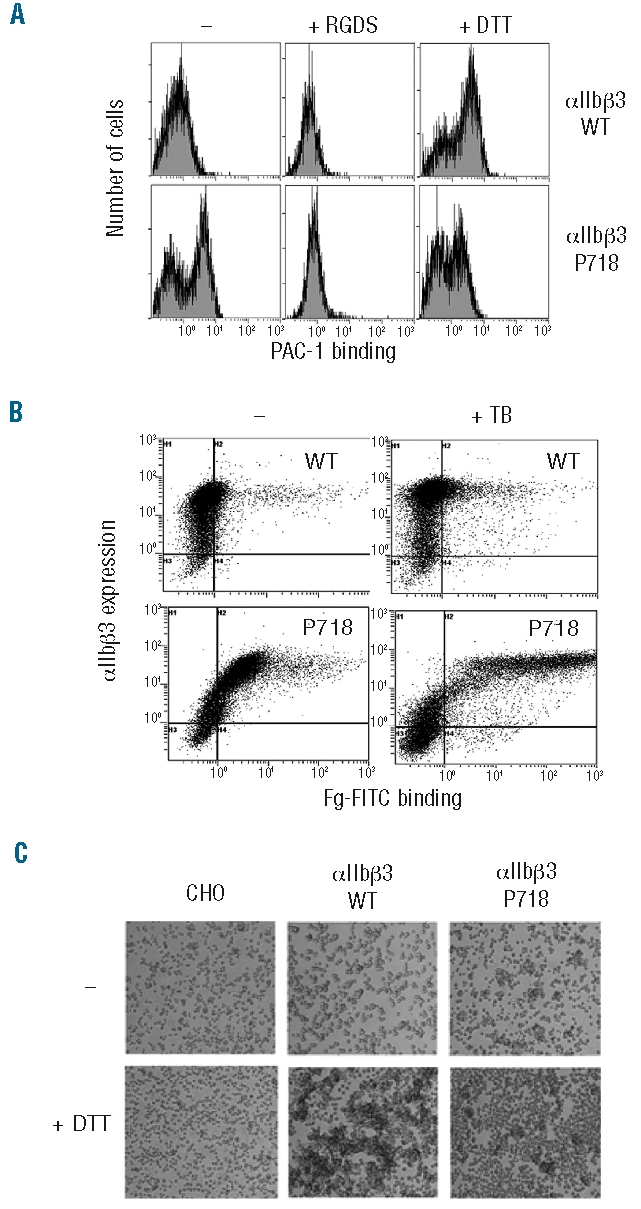
Binding of fibrinogen and PAC-1 to normal or mutant αIIbβ3 expressed in CHO cells. (A) CHO cells stably expressing wild type or mutant αIIbβ3-P718 were preincubated in the absence or presence of 1 mM RGDS or 25 mM DTT for 5 min at room temperature. Cells were then incubated with FITC-PAC-1 for 30 min, washed, and analyzed by flow cytometry. (B) CHO cells were first labeled with anti-αIIb monoclonal antibody and then incubated with FITC-Fg in the absence or presence of thrombin as described in the Design and Methods section. (C) Soluble fibrinogen-dependent aggregation of CHO cell transfectants. Non-transfected cells (CHO) or αIIbβ3 expressing cells were incubated in serum-free medium containing 1 mg/mL fibrinogen in the absence or presence of 15 mM DTT. Aggregates were examined with a phase contrast microscope using a x4 objective. All the results are representative of, at least, three separate experiments.
β3-P718 induces an atypical pattern of αIIbβ3 clustering in immobilized fibrinogen-adherent cells
β3 integrins mediate cell spreading on fibrinogen-coated surfaces.25 When plated on fibrinogen, CHO cells stably expressing mutant αIIbβ3-P718 showed a peculiar spreading phenotype with extensions. A number of cells also exhibited one or two long protrusions largely exceeding the cell borders and often with marked swelling at the tip. This extension pattern was not observed in cells attached to plates coated with serum proteins (Figure 4A–B). Although no differences were observed in static adhesion assays, cells expressing αIIbβ3-P718 integrin showed enhanced adhesion under flow (Online Supplementary Figure S3).
Figure 4.
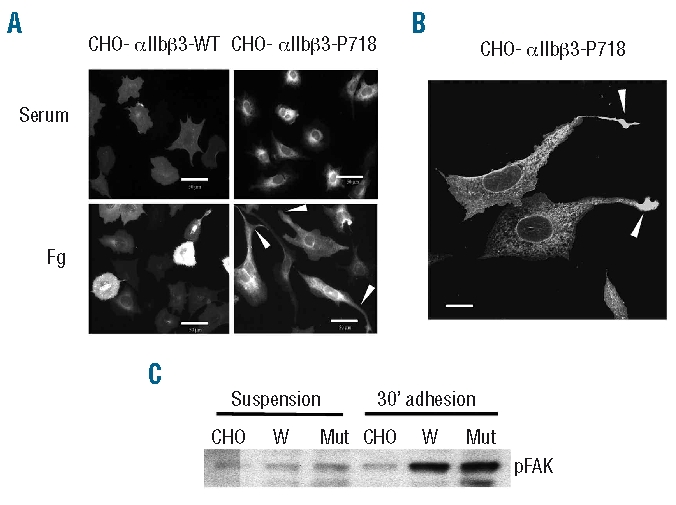
Fibrinogen-mediated spreading of CHO cell transfectants. (A) Fluorescence microphotographs of CHO cell transfectants seeded for 16 h on fibrinogen or serum proteins. Cells were fixed, labeled with anti-β3 monoclonal antibody H1AG11 and examined in an epifluorescence microscope with a x40 objective. Arrowheads point to the extending processes in cells expressing mutant αIIbβ3-P718. Bars: 50 μm. (B) Arrows point to the swelling at the tip of the extending protrusion in CHO-αIIbβ3-P718 cells examined with a x63 objective. Bar: 25 μm. (C) Tyrosine phosphorylation of FAK in CHO cells. Non-transfected (CHO) or cells expressing wild-type (W) or mutant αIIbβ3-P718 were maintained in suspension or adhered to fibrinogen for 30 min. Cell lysates were analyzed by western blot. All the experiments were performed at least three times and similar results were obtained.
In order to determine whether the active L718P mutation in β3 could affect ligand-induced integrin redistribution, we explored the recruitment of αIIbβ3 to focal adhesions. In our experimental conditions, wild type integrins were found uniformly diffused throughout the cell surface, or grouped in small clusters at the periphery in fibrinogen-adherent cells (Figure 5A). However, when mutant β3-expressing cells were plated on fibrinogen-coated surfaces we observed the formation of fluorescent integrin clusters evenly distributed underneath the entire cell body. They also displayed an abnormal clustering pattern consisting of patches of very high receptor density that contained integrin-free round areas. Confocal microscopy analysis localized integrin clusters in membrane sections close to the adherent surface in contact with the substratum. These atypical clusters of mutant αIIbβ3 were detected using both anti-αIIb (Figure 5A–B) and anti-αIIbβ3 (Figure 5B) monoclonal antibodies, and were not observed when cells expressing wild αIIbβ3 were stimulated with 1 mM Mn2+ (not shown). Double staining with phalloidin and anti-αIIb revealed that the abnormal clusters did not correspond to focal adhesions because they did not locate near the termini of actin stress fibers (Figure 5B). Interestingly, the patches were initially localized underneath the main cell body but, after long-term cell spreading, they adopted a polarized position at the opposite side of emerging protrusions (Figure 5C).
Figure 5.
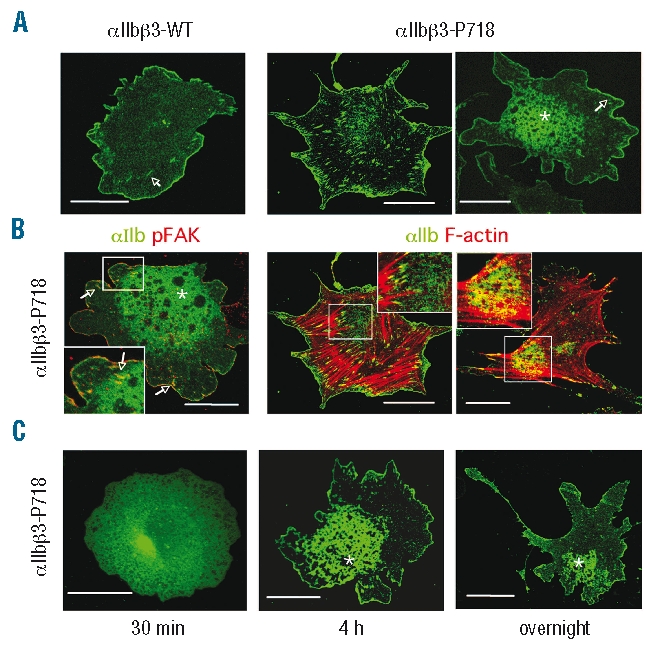
Clustering of mutant αIIbβ3-718P stably expressed in CHO cells. (A) CHO cells stably expressing wild-type or mutant αIIbβ3-P718 were seeded on fibrinogen-coated coverslips for 4 h and then fixed and labeled with the anti-αIIb monoclonal antibody 2BC1, and examined by confocal microscopy. (B) CHO cell transfectants expressing αIIbβ3-P718 were seeded on fibrinogen-coated cover-slips for 4 h and then labeled with anti-αIIbβ3 monoclonal antibody P2, and anti-FAK[pY576] polyclonal antibody or phalloidin, and analyzed by confocal microscopy. (C) CHO cells expressing αIIbβ3-P718 were plated on fibrinogen-coated coverslips for the indicated periods of time, fixed and labeled with 2BC1, and analyzed by confocal microscopy. The images are single confocal sections near the adherent surface of the cells. Bars: 30 μm. Arrowheads point to functional adhesion complexes. Asterisks point to cell regions where the atypical integrin clusters locate. These experiments were repeatedly performed (n>4) with the same results.
Consistent with the atypical clustering pattern of mutant αIIbβ3 integrin in transfected cells, confocal immunofluorescence analysis of platelets from the thrombasthenic patient revealed a non-homogeneous distribution of αIIbβ3 in the adherent surface in contact with the substratum (Online Supplementary Figure S4).
Cell lines expressing other αIIbβ3 activating mutants have been shown to exhibit constitutive phosphorylation of the focal adhesion adaptor protein FAK when in suspension.6,19 However, FAK phosphorylation in cells expressing mutant αIIbβ3-P718 was similar to that of control cells, regardless of whether the cells were in suspension or adhered to fibrinogen-coated plates (Figure 4C). Moreover, immunolabeling of cells adhered to fibrinogen showed that phosphorylated FAK colocalized with mutant αIIbβ3 in focal adhesions but was not detected within the atypical patches of clustered integrin (Figure 5B).
The proximity between αIIbβ3 molecules was further analyzed by treating cells with disuccinimidyl suberate (DSS), a cross-linker with spacer arm length of 11.4 Å. In cells expressing β3-P718, immunoprecipitation of αIIbβ3 with either anti-β3 or anti-αIIb monoclonal antibodies showed a significant increase of protein complexes of approximately 200 kDa that should correspond to combinations of αIIb and β3 subunits since they were detected with both antibodies (Online Supplementary Figure S5). Given that both wild-type and mutant cells express similar receptor levels, it is reasonable to deduce that such combinations do not correspond to the native αIIbβ3 heterodimers. Rather, they could result from crosslinking of α and β subunits from two different integrin molecules that would be closer than in cells transfected with the normal β3 subunit. In addition, the different patterns of cross-linked proteins obtained with some of the antibody pairs may reflect the fact that the molecules surrounding integrins are not the same in the two types of cells.
Clustering of αIIbβ3-P718 associates with disruption of ordered lipid domains in the plasma membrane
Current models of biological membranes show that protein clustering or cross-linking could affect the formation or distribution of different lipid phases.17 Cholesterol is thought to contribute to the tight packing of lipids in ordered domains;26 in addition, these domains can be formed only within certain ranges of cholesterol concentration.27 We hypothesized that the round areas devoid of α IIbβ3 observed within the atypical clustering patches may correspond to coalescence of lipid domains and could, therefore, be sensitive to cellular cholesterol depletion. Thus, we incubated the cells with MβCD, a cholesterol-sequestering agent widely used to study the effect of cholesterol depletion and lipid raft disassembly. In cells expressing mutant αIIbβ3-P718 integrin, MβCD disrupted the atypical clustering patches, fusing the round areas into larger and more irregular integrin-free domains (Figure 6A). A very incipient pattern of integrin-free round areas could also be observed in some control cells when treated with a high concentration of MβCD.
Figure 6.

Clustering of mutant αIIbβ3-718P induces coalescence of lipid domains. (A) Effect of cholesterol depletion on the clustering pattern induced by β3-P718. CHO cells were treated for 15 min at 37ºC with MβCD followed by plating on fibrinogen-coated coverslips. Cells were then fixed, labeled with 2BC1 and analyzed by confocal microscopy. In the bottom panels, MβCD-induced disruption of αIIbβ3-P718 integrin clustering is shown under high magnification. (B) Distribution of the fluorescent lipid analog DiLC16 in the plasma membrane of CHO cells expressing normal or mutant αIIbβ3 integrin. CHO cells stably expressing wild type or mutant αIIbβ3 were seeded on fibrinogen-coated coverslips for 4 h and then labeled with the anti-αIIbβ3 monoclonal antibody 2BC1 and the fluorescent lipid-analog DiIC16 as described in the Design and Methods section, and analyzed by confocal microscopy. Bars: 30 μm. Representative pictures of more than four separate experiments are shown.
Lipids with long and saturated acyl chains, such as the synthetic fluorescent lipid mimetic DiIC16, preferentially incorporate into the cholesterol-enriched Lo microdomains and have been used to observe the formation and dynamics of membrane domains in model plasma membranes28 and living cell membranes.29,30 To further corroborate a role of mutant αIIbβ3-P718 in membrane lipid phase behavior, cells were stained with DiIC16 and analyzed by confocal microscopy. As shown in Figure 6B, in cells expressing normal αIIbβ3, DiIC16 labeling was uniformly distributed in confocal sections near the cell surface in contact with the substratum. In contrast, cells expressing the mutant integrin showed a “Swiss cheese“ pattern with non-stained round areas that match the integrin-devoid holes.
Discussion
We have studied the functional consequences of a novel de novo mutation, L718P, located in the membrane-proximal region of the β3 cytoplasmic domain, and associated with a severe thrombasthenia-like phenotype in a patient carrying one normal β3 allele. The 50% reduction of reactivity in the flow cytometry analysis with a complex-dependent monoclonal antibody contrasted with the expression detected using other antibodies and with the normal content of αIIb and β3 in the western blot analysis, and suggested an almost normal amount but aberrant structure of native αIIbβ3 in the patient’s platelets. Consistent with this, the surface expression of mutant αIIbβ3 in transiently transfected cells was only faintly diminished. However, the surface content of mutant β3-P718 complexed to αv was markedly reduced due to intracellular retention of mutant αvβ3. This distinctive effect of the mutation on the processing of β3 integrins is unexpected in view of the conservation of the membrane-proximal sequences among the α subunits. Mutant αvβ3 integrin was recognized by a specific monoclonal antibody within some Golgi-related structures, but accumulation of more immature complexes within the endoplasmic reticulum was only detected with the anti-β3 monoclonal antibody, which also recognizes αIIbβ3. Thus, it cannot be excluded that a partial retention of mutant αIIbβ3 integrins at the level of the endoplasmic reticulum contributed to the decreased surface detection in the patient’s platelets.
A minor subpopulation of the patient’s platelets elicited spontaneous binding to soluble fibrinogen but platelet aggregation as well as fibrinogen binding induced by agonists were severely deficient. On the other hand, cells stably expressing the αIIbβ3 mutant receptor showed spontaneous binding and aggregation in the presence of multivalent ligands, but failed to respond to DTT. Taken together, these data suggest that the intracellular defect induces a conformational change that is propagated to the extracellular domain, rendering partially activated receptors unable to support normal aggregation. L718P substitution in β3 alters the second leucine in the membrane-proximal sequence LLXXXHDRR, highly conserved in all integrin β subunits. On the basis of mutational analysis and structural modeling, the interaction between the membrane-proximal domains of αIIb and β3 cytoplasmic tails has been proposed to play an important role in maintaining the αIIbβ3 receptor in a low affinity state.3,4,31 Thus, the L718P mutation may disrupt such an interaction rendering partially activated αIIbβ3. In support of this notion, activating L718H was identified in a random mutagenesis assay.2 Furthermore, it cannot be excluded that the P718-induced structural change facilitated the access to talin, which contributes substantially to integrin activation.32,33
Half of the normal αIIbβ3 content is sufficient to support normal platelet aggregation in heterozygous carriers of Glanzmann’s thrombasthenia mutations.8 Thus, another mechanism presumably contributes to the defective receptor function and thrombasthenia-like phenotype in the patient. Our results indicate that mutant β3-P718 promotes aberrant αIIbβ3 distribution in both platelets and transfected cells which, in turn, results in altered outside-in signaling as measured by the abnormal adhesion and/or spreading phenotype of platelets and adherent transfected cells on immobilized fibrinogen. Consistent with this, significant differences were found in the immunoprecipitation pattern of cross-linked wild-type and mutant receptors, suggesting that lateral interactions of mutant integrins with other proteins may be altered. To date, the intermolecular forces driving integrin clustering are poorly understood.34 Increasing evidence suggests that the orientation of transmembrane helices and their tendency to self-associate can be modulated by the lipid environment into which they are embedded, and it is clear that even a minor change in, for instance, the orientation of the transmembrane segments in the lipid bilayer can have important structural and functional consequences for the intact membrane protein.35 The recently determined high-resolution structure of the β3 integrin transmembrane segment and flanking sequences in phospholipid bicelles showed that the membrane-proximal hydrophobic residues L717-721 appear to be immersed in the membrane, thus ensuring a pronounced transmembrane helix tilt relative to the lipid bilayer.36 As reported for other transmembrane proteins,37 the introduced proline residue may form a hinge in the alpha helix that, in turn, may induce a hydrophobic mismatch with the membrane bilayer. Integrin clustering at the cell surface may, therefore, be a direct consequence of hydrophobic mismatching between mutant β3 and lipids in the membrane due to the presence of proline at position 718.
It is widely accepted that the plasma membrane is a laterally non-homogeneous mixture of lipids, with an organized structure in which lipid molecules segregate and form Lo microdomains – sphingolipid- and sterol-enriched lipid rafts – which coexist with more fluid domains having lower resistance to extraction with non-ionic detergents.38 Protein recruitment at the interface between lipid domains and the surrounding membrane is thought to contribute to a reduction in line tension at the phase boundary, changing the propensity of the domains to fuse.17 In our study, the effect of cholesterol depletion and the differential labeling of a lipid analog that preferentially incorporates into ordered domains, support the notion that the atypical αIIbβ3 recruitment induced by the L718P-β3 mutation promotes coalescence of apparently liquid-disordered or more fluid microdomains. One of the more important roles of lipid microdomains at the cell surface is their contribution to regulating the efficacy of signaling by concentrating selected proteins at specific sites.18–20 Hence, αIIbβ3 integrin clustering, by promoting lipid coalescence, may redistribute signaling molecules and, as a result, transmit aberrant outside-in signals. In our case, defective outside-in signaling can be inferred from abnormal spreading and clustering patterns in both CHO cells and platelets.
Another natural mutation in the membrane-proximal region of β3, D723H, has recently been reported in heterozygosis in a patient with an autosomal platelet disorder consisting of mild thrombocytopenia, platelet anisocytosis, and giant platelets.12 This mutation is predicted to disrupt a conserved salt bridge that stabilizes the interaction between the membrane-proximal regions of the α and β subunits13 and, similar to L718P, was found to activate αIIbβ3 and to induce long protrusions in transfected CHO cells plated on fibrinogen. In CHO-αIIbβ3-P718 cells, the atypical integrin-enriched patch was uniformly distributed upon adhesion but, during cell spreading, it relocated outside the membrane portion that emits protruding extensions. This polarized location may reflect that the aberrant clustering patch impedes normal retraction/detachment from the substratum and, consequently, deregulates cell spreading. The mild thrombocytopenia and platelet anisocytosis in the present case are, therefore, consistent with the recent proposal that outside-in signaling via αIIbβ3 after fibrinogen engagement is an important component of platelet formation.39 In accordance with this, dominant inheritance of an in-frame deletion in the ectodomain of β3 has been associated with macrothrombocytopenia and platelet dysfunction in two Italian families.40 It is possible that, as previously suggested,41 platelet anisocytosis arises from a physical effect through which increased proplatelet adhesion to fibrinogen alters platelet release in bone marrow sinusoids.
In summary, we report the functional characterization of L718P mutation in β3, identified in heterozygosis as the only αIIbβ3 pathogenic mutation in a patient with a thromboasthenia-like phenotype. The findings support a role for integrin clustering in regulating outside-in signaling through lipid microdomain coalescence. According to these findings, we propose aberrant integrin clustering as the mechanism underlying the thrombasthenic phenotype in the patient. Nevertheless, it cannot be ruled out that the patient has a second unidentified defect in another gene accounting for at least part of the platelet functional defects. Our study also yields insights into the role of the proximal-membrane region of β3 in the function of αIIbβ3 in vivo, and supports the notion that αIIbβ3 receptor clustering is necessary coupled to its conformational activation.
Acknowledgments
we thank María Teresa Seisdedos and Silvia Hernández for technical assistance, with the confocal microscopy.
Footnotes
Funding: this work was supported by grants from the Dirección General de Investigación del Ministerio de Educación y Ciencia (BFU2006-00914) and Fundación Rodríguez Pascual. AJ was recipient of a fellowship from the Ministerio de Educación y Ciencia. IC holds a research contract from the Consejo Superior de Investigaciones Científicas.
The online version of this article has a Supplementary Appendix.
Authorship and Disclosures
AJ and IC performed a major part of the experiments and analyzed data. PL was involved in the design of the experiments and contributed to the analysis of data. JR, CM, and VV performed the initial analysis of the patient’s platelets. CG-M designed the research, performed experiments and wrote the paper.
The authors declare no competing financial interests.
References
- 1.Hynes RO. Integrins: bidirectional, allosteric signaling machines. Cell. 2002;110(6):673–87. doi: 10.1016/s0092-8674(02)00971-6. [DOI] [PubMed] [Google Scholar]
- 2.Partridge AW, Liu S, Kim S, Bowie JU, Ginsberg MH. Transmembrane domain helix packing stabilizes integrin αIIbβ3 in the low affinity state. J Biol Chem. 2005;280(8):7294–300. doi: 10.1074/jbc.M412701200. [DOI] [PubMed] [Google Scholar]
- 3.Hughes P, O’Toole TE, Ylänne J, Shattil SJ, Ginsberg MH. The conserved membrane-proximal region of an integrin cytoplasmic domain specifies ligand binding affinity. J Biol Chem. 1995;270(21):12411–7. doi: 10.1074/jbc.270.21.12411. [DOI] [PubMed] [Google Scholar]
- 4.Ma YQ, Yang J, Pesho MM, Vinogradova O, Qin J, Plow EF. Regulation of integrin alphaIIbbeta3 activation by distinct regions of its cytoplasmic tails. Biochemistry. 2006;45(21):6656–62. doi: 10.1021/bi060279h. [DOI] [PubMed] [Google Scholar]
- 5.Hato T, Pampori N, Shattil SJ. Complementary roles for receptor clustering and conformational change in the adhesive and signaling functions of integrin alphaIIbbeta3. J Cell Biol. 1998;141(7):1685–95. doi: 10.1083/jcb.141.7.1685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Li R, Mitra N, Gratkowski H, Vilaire G, Litvinov R, Nagasami C, et al. Activation of integrin αIIbβ3 by modulation of transmembrane helix associations. Science. 2003;300(5620):795–8. doi: 10.1126/science.1079441. [DOI] [PubMed] [Google Scholar]
- 7.Luo B-H, Carman CV, Takagi J, Springer TA. Disrupting integrin transmembrane heterodimerization increases ligand binding affinity, not valency or clustering. Proc Natl Acad Sci. 2005;102(109):3679–84. doi: 10.1073/pnas.0409440102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.George JN, Caen JP, Nurden AT. Glanzmann’s thrombasthenia: the spectrum of clinical disease. Blood. 1990;75(7):1383–95. [PubMed] [Google Scholar]
- 9.Nair S, Ghosh K, Kulkarni B, Shetty S, Mohanty D. Glanzmann’s thrombasthenia: update. Platelets. 2002;13(7):387–93. doi: 10.1080/0953710021000024394. [DOI] [PubMed] [Google Scholar]
- 10.Nurden AT. Glanzmann thrombasthenia. Orphanet J Rare Dis. 2006;1:10–7. doi: 10.1186/1750-1172-1-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Peyruchaud O, Nurden AT, Milet S, Macchi L, Pannochia A, Bray PF, et al. R to Q amino acid substitution in the GFFKR sequence of the cytoplasmic domain of the integrin IIb subunit in a patient with a Glanzmann’s thrombasthenia-like syndrome. Blood. 1998;92(11):4178–87. [PubMed] [Google Scholar]
- 12.Ghevaert C, Salsmann A, Watkins NA, Schaffner-Reckinger E, Rankin A, Garner SF, et al. A nonsynonymous SNP in the ITGB3 gene disrupts the conserved membrane-proximal cytoplasmic salt bridge in the αIIbβ3 integrin and cosegregates dominantly with abnormal proplatelet formation and macrothrombocytopenia. Blood. 2008;111(7):3407–14. doi: 10.1182/blood-2007-09-112615. [DOI] [PubMed] [Google Scholar]
- 13.Hughes PE, Diaz-Gonzalez F, Leong L, Wu C, McDonald JA, Shattil SJ, Ginsberg MH. Breaking the integrin hinge. J Biol Chem. 1996;271(12):6571–4. doi: 10.1074/jbc.271.12.6571. [DOI] [PubMed] [Google Scholar]
- 14.Wang R, Shattil SJ, Ambruso DR, Newman PJ. Truncation of the cytoplasmic domain of beta3 in a variant form of Glanzmann thrombasthenia abrogates signaling through the integrin alphaIIbbeta complex. J Clin Invest. 1997;100(9):2393–403. doi: 10.1172/JCI119780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chen YP, Djaffar I, Pidard D, Steiner B, Cieutat AM, Caen JP, Rosa JP. Ser-752>Pro mutation in the cytoplasmic domain of integrin beta3 subunit and defective activation of platelet integrin alphaIIbbeta3 (glycoprotein IIb–IIIa) in a variant of Glanzmann thrombasthenia. Proc Natl Acad Sci USA. 1992;89(21):10169–73. doi: 10.1073/pnas.89.21.10169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Rosenberg N, Hauschner H, Peretz H, Mor-Cohen R, Landau M, Shenkman B, et al. A 13-bp deletion in alpha(IIb) gene is a founder mutation that predominates in Palestian-Arab patients with Glanzmann thrombasthenia. J Thromb Haemost. 2005;3(12):2764–72. doi: 10.1111/j.1538-7836.2005.01618.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pike LJ. The challenge of lipid rafts. J Lipid Res. 2009;50:S323–8. doi: 10.1194/jlr.R800040-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sengupta P, Baird B, Holowka D. Lipid raft, fluid/fluid phase separation, and their relevance to plasma membrane structure and function. Semin Cell Dev Biol. 2007;18(5):583–90. doi: 10.1016/j.semcdb.2007.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhang AY, Yi F, Gulbins E, Li P-L. Lipid raft clustering and redox signaling platform formation in coronary arterial endothelial cells. Hypertension. 2006;47(1):74–80. doi: 10.1161/10.1161/01.HYP.0000196727.53300.62. [DOI] [PubMed] [Google Scholar]
- 20.Hofman EG, Ruonala MO, Bader AN, van den Heuvel D, Voortman J, Roovers RC, et al. EGF induces coalescence of different lipid rafts. J Cell Sci. 2008;121(pt 15):2519–28. doi: 10.1242/jcs.028753. [DOI] [PubMed] [Google Scholar]
- 21.Hao M, Mukherjee S, Maxfield FR. Cholesterol depletion induces large scale domain segregation in living cell membranes. Proc Natl Acad Sci USA. 2001;98(23):13072–7. doi: 10.1073/pnas.231377398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Butta N, Arias-Salgado EG, González-Manchón C, Ferrer M, Larrucea S, Ayuso MS, Parrilla R. Disruption of the beta3 663–687 disulfide bridge confers constitutive activity to beta3 integrins. Blood. 2003;102(7):2491–7. doi: 10.1182/blood-2003-01-0213. [DOI] [PubMed] [Google Scholar]
- 23.Pabón D, Jayo A, Lastres P, González-Manchón C. Thrombin induces GPIb-IX-mediated fibrin binding to αIIbβ3 in a reconstituted Chinese hamster ovary cell model. J Thromb Haemost. 2006;4(10):2238–47. doi: 10.1111/j.1538-7836.2006.02141.x. [DOI] [PubMed] [Google Scholar]
- 24.Ruan J, Peyruchaud O, Nurden A, Bourre F. Linkage of four polymorphisms on the alphaIIb gene. Br J Haematol. 1998;102(2):622–5. doi: 10.1046/j.1365-2141.1998.00810.x. [DOI] [PubMed] [Google Scholar]
- 25.Ylanne J, Chen Y, O’Toole TE, Loftus JC, Takada Y, Ginsberg MH. Distinct functions of integrin alpha and beta subunit cytoplasmic domains in cell spreading and formation of focal adhesions. J Cell Biol. 1993;122(1):223–33. doi: 10.1083/jcb.122.1.223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Brown RE. Sphingolipid organization in biomembranes: what physical studies of model membranes reveal. J Cell Sci. 1998;111(pt 1):1–9. doi: 10.1242/jcs.111.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Brown DA, London E. Structure and function of sphingolipid- and cholestero-rich membrane rafts. J Biol Chem. 2000;275(23):17221–4. doi: 10.1074/jbc.R000005200. [DOI] [PubMed] [Google Scholar]
- 28.Hammond AT, Heberle FA, Baumgart T, Holowka D, Baird B, Feigenson GW. Crosslinking a lipid raft component triggers liquid ordered-liquid disordered phase separation in model plasma membrane. Proc Natl Acad Sci USA. 2005;102(18):6320–5. doi: 10.1073/pnas.0405654102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mukherjee S, Soe TT, Maxfield FR. Endocytic sorting of lipid analogues differing solely in the chemistry of their hydrophobic tails. J Cell Biol. 1999;144(6):1271–84. doi: 10.1083/jcb.144.6.1271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Adachi S, Nagao T, Ingolfsson HI, Maxfield FR, Andersen OS, Kopelovich L, Weinstein IB. The inhibitory effect of (−)-epigallocatechin gallate on activation of the epidermal growth factor receptor is associated with altered lipid order in HT29 colon cancer cells. Cancer Res. 2007;67(13):6493–501. doi: 10.1158/0008-5472.CAN-07-0411. [DOI] [PubMed] [Google Scholar]
- 31.Ma Y-Q, Qin J, Plow EF. Platelet integrin αIIbβ3: activation mechanisms. J Thromb Haemost. 2007;5(7):1345–52. doi: 10.1111/j.1538-7836.2007.02537.x. [DOI] [PubMed] [Google Scholar]
- 32.Tadokoro S, Shattil SJ, Eto K, Tai V, Liddington RC, de Pereda JM, et al. Talin binding to integrin β tails: a final common step in integrin activation. Science. 2003;302(5642):103–6. doi: 10.1126/science.1086652. [DOI] [PubMed] [Google Scholar]
- 33.Hato T, Yamanouchi J, Tamura T, Yakushijin Y, Sakai I, Yasukawa M. Cooperative role of the membrane-proximal and -distal residues of the integrin β3 cytoplasmic domain in regulation of talinmediated αIIbβ3 activation. J Biol Chem. 2008;283(9):5662–8. doi: 10.1074/jbc.M707246200. [DOI] [PubMed] [Google Scholar]
- 34.Shattil SJ, Newman PJ. Integrins: dynamic scaffolds for adhesion and signaling in platelets. Blood. 2004;104(6):1606–15. doi: 10.1182/blood-2004-04-1257. [DOI] [PubMed] [Google Scholar]
- 35.Nyholm TK, Özdirekcan S, Killian JA. How protein transmembrane segments sense lipid environment. Biochemistry. 2007;46(6):1457–65. doi: 10.1021/bi061941c. [DOI] [PubMed] [Google Scholar]
- 36.Lau T-L, Partridge AW, Ginsberg MH, Ulmer TS. Structure of the integrin β3 transmembrane segment in phospholipid bicelles and detergent micelles. Biochemistry. 2008;47(13):4008–16. doi: 10.1021/bi800107a. [DOI] [PubMed] [Google Scholar]
- 37.Yeagle PL, Bennett M, Lemaître V, Watts A. Transmembrane helices of membrane proteins may flex to satisfy hydrophobic mismatch. Biochim Biophys Acta. 2007;1768(3):530–7. doi: 10.1016/j.bbamem.2006.11.018. [DOI] [PubMed] [Google Scholar]
- 38.Simons K, Vaz WL. Model systems, lipid rafts, and cell membranes. Annu Rev Biophys Biomol Struct. 2004;33:269–95. doi: 10.1146/annurev.biophys.32.110601.141803. [DOI] [PubMed] [Google Scholar]
- 39.Larson MK, Watson SP. Regulation of pro-platelet formation and platelet release by integrin αIIbβ3. Blood. 2006;108(5):1509–14. doi: 10.1182/blood-2005-11-011957. [DOI] [PubMed] [Google Scholar]
- 40.Gresele P, Falcinelli E, Giannini S, D’Adamo P, D’Eustacchio A, Corazzi T, et al. Dominant inheritance of a novel integrin β3 mutation associated with a hereditary macrothrombocytopenia and platelet dysfunction in two Italian families. Haematologica. 2009;94(5):663–9. doi: 10.3324/haematol.2008.002246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bray PF. Sizing up platelet defects. Blood. 2008;111(7):3302–3. [Google Scholar]