

Abstract
Background:
Increased serum levels of procalcitonin (ProCT) and its component peptides have been reported in humans with sepsis. Using a hamster model of bacterial peritonitis, we investigated whether serum ProCT levels are elevated and correlate with mortality and hypocalcemia.
Results:
Incremental increases in doses of bacteria resulted in proportional increases in 72h mortality rates (0, 20, 70, and 100%) as well as increases in serum total immunoreactive calcitonin (iCT) levels at 12 h (250, 380, 1960, and 4020 pg/ml, respectively, vs control levels of 21 pg/ml). Gel filtration studies revealed that ProCT was the predominant (> 90%) molecular form of serum iCT secreted. In the metabolic experiments, total iCT peaked at 12 h concurrent with the maximal decrease in serum calcium.
Conclusions:
In this animal model, hyper-procalcitoninemia was an early systemic marker of sepsis which correlated closely with mortality and had an inverse correlation with serum calcium levels.
Keywords: inflammation, peritonitis, procalcitonin, prohormone, serum marker
Introduction
There are approximately 400000 cases of sepsis reported each year in the USA, leading to about 100000 deaths annually [1,2,3]. Indeed, mortality from sepsis in most series is reported to be between 25 and 40%, with Gram-negative bacteria being the most commonly encountered pathogens [3,4,5]. The severity of sepsis may distinguish those who may benefit from therapeutic blockade of their excessive and maladaptive immune response, from those who may not. Consequently, a practical way to determine the presence and severity of sepsis is essential. Although systems of evaluation based on clinical observations and physiologic parameters are helpful, they have been of limited use for predicting morbidity and mortality in individuals with inflammatory conditions, especially in surgical populations [6,7,8,9,10]. An early indicator of tissue injury should improve the predictive capability of these systems. Although several cytokines have been proposed as markers of disease severity, they are often transiently elevated, or detected only in local pools [7]. In this regard, recent studies in humans have revealed that the prohormone of calcitonin (CT), procalcitonin (ProCT), as well as its component peptides offer promise of being early and useful predictive markers of systemic inflammation [11,12,13].
CT is a neuroendocrine (NE) peptide that was once thought to be exclusively a hormone of thyroid origin. Its principal function appears to be the conservation of body calcium stores in certain physiologic states such as growth, pregnancy and lactation, and the maintenance of bone mineral in emergency situations by means of attenuation of the activity of osteoclasts [14]. Further study has revealed that CT is produced extrathyroidally by NE cells throughout the body, and may have multiple functions [15,16].
CT is initially biosynthesized as a larger ProCT polypeptide which is subsequently cleaved enzymatically into its components, including the mature, active hormone (Fig 1). Interestingly, in humans with severe systemic inflammation, very high serum levels of ProCT and its component peptides are accompanied by normal or only slightly increased levels of mature CT [17]. In order to investigate whether serum ProCT levels might correlate with the severity of illness in sepsis, and thus might provide a convenient marker, we employed a rodent model of quantifiable Escherichia coli peritonitis, modified for use in the hamster [18]. This model was then utilized to determine the metabolic perturbations associated with the procalcitonin peptide levels observed with sepsis.
Figure 1.
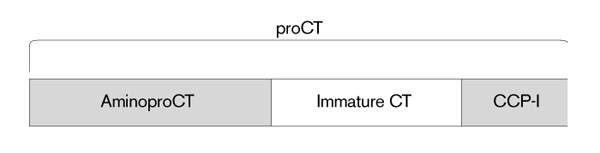
The procalcitonin (ProCT) molecule and its components. AminoproCT = amino terminus of procalcitonin; immature CT = the 33 amino acid, non-amidated CT; CCP-I = calcitonin carboxyterminus peptide-I. In normal people, in addition to the free, active, mature CT, small amounts of ProCT, aminoproCT, CCP-I, the conjoined CT:CCP-I peptide, and the immature CT circulate [18]. The amino acid sequence of the rat mature CT is very similar to that of humans, and the sequence of hamster CT, although not yet known, reveals, by immunoassay studies, a marked homology with the rat.
Materials and methods
Animals
Male Golden-Syrian hamsters weighing 80–140g (Harlan Animals, Indianapolis, Indiana, USA) were housed in a controlled environment and were exposed to 12 h light–dark cycles. The animals had unrestricted access to water and standard rodent chow throughout the experiments. This study was approved by the Institutional Animal Care and Use Committee at the Veterans Affairs Medical Center, Washington, DC.
Bacteria
Escherichia coli (O18:K1:H7) were obtained from Dr Alan S Cross, Division of Communicable Diseases and Immunology, Walter Reed Army Institute of Research, Washington, DC, USA. The bacteria were grown in 100 ml of LB Broth (Fisher Scientific, Pittsburgh, Pennsylvania, USA) at 37°C in a shaker water bath to log phase and stored in 250 μ l aliquots at -70°C until use.
On the day of an experiment, a 250 μ l aliquot of bacteria was thawed and grown in 100 ml LB broth at 37°C in a shaker water bath to log phase. The optical density of the specimen was measured at 600 nm on a Stasar III spectrophotometer (Gilford Instruments, Oberlin, Ohio, USA) and quantified by interpolation on a previously constructed curve of optical density plotted against colony forming units (cfu). Additional specimens were taken from the stock solution, and diluted and plated to confirm the counts estimated by spectrophotometry.
Intra-abdominal pellets
Bacterial suspensions of 2.0 × 108, 1.0 × 109, 2.0 × 109, or 4.0 × 109 cfu/ml E coli were pipetted in 0.5 ml aliquots into 8 mm plastic embedding molds (Shandon-Upshaw, War-rington, Pennsylvania, USA). Each pellet for implantation was made by adding 0.5 ml sterile molten agar at 50°C to the bacterial suspension, after which the mixture was allowed to solidify at room temperature. The final number of viable colony forming units of bacteria in each pellet was 1.0 × 108, 5.0 × 108, 1.0 × 109, or 2.0 × 109 cfu/pellet.
Experimental protocol
Mortality studies
Individual hamsters were assigned to four groups (n = 16/group) to receive progressively increasing inocula of bacteria. After adequate anesthesia with 50 mg/kg pentobarbital via intraperitoneal injection, the abdomen of each animal was prepared with 70% alcohol and incised in the midline. Bacterial sepsis was induced by implanting one pellet in the right lower quadrant of the peritoneal cavity of each animal. The abdominal incisions were then closed with non-absorbable suture. Animals were caged individually, given unrestricted access to water and rodent chow and monitored for mortality over a 72 h period.
Total iCT studies
After intraperitoneal implantation of agar pellets with progressively increasing doses of E coli, separate groups (n = 10/group) were killed for serum total immunoreactive (i)CT determinations. Since mortality was evident but not prohibitively high at 12 h, we chose this timepoint to determine serum total iCT levels. Therefore, 12 h after animals were challenged with E coli, they were anesthetized with intraperitoneal pentobarbital (50 mg/kg) and exsanguinated by open cardiac puncture. The blood was collected in individual tubes and centrifuged at 3000 rpm for 15 min. The serum specimens were transferred to individual glass tubes, sealed with parafilm and stored at -70°C until radioimmunoassay.
Serum was also obtained from a patient with documented Gram-negative sepsis and was stored at -70°C to be assayed with the hamster serum samples following G-75 Sephadex gel filtration for the purpose of comparison of molecular forms as described below.
Metabolic studies
Male hamsters (n = 16/group) underwent intraperitoneal implantation of agar pellets impregnated with 2 × 109 cfuE coli (O18: K1: H7), according to the above implantation protocol. This high dose was chosen for its ability to induce a significant increase of ProCT at 12 h in the proceeding experiments. Animals were killed in the previously described fashion at 3, 6, 12 or 24 h after septic challenge. Their sera were analyzed for serum total iCT per the radioimmunoassay described below, as well as for total serum total calcium and serum albumin using a standard serum multichannel analyzer.
Radioimmunoassay
The samples were allowed to warm to room temperature and were pipetted into labeled glass test tubes in 1.0 ml aliquots, to which 100 μ l dextran blue (B-2000, 2 000 000 Da; Sigma Chemical Co, St Louis, Missouri, USA) was added. Five milliliter glass columns were rinsed with 1M ammonium hydroxide:acetonitrile (1:1) and deionized water, after which fine-grade polyacrylamide gel columns (5ml) were prepared (BioGel P-2; 100–200 mesh; Bio-Rad Laboratories, St Louis, Missouri, USA) using a glass bead to support the gel. The samples were applied to the columns and eluted with 0.1 M ammonium bicarbonate containing 0.1% Triton X-100 (Pierce, Rockford, Illinois, USA). The specimens containing dextran blue were then recovered in their original test tubes, to which ethyl alcohol was added in a 2:1 volume ratio. These mixtures were then centrifuged at 3000 rpm for 30 min, after which the supernatant for each was decanted into new tubes and the pellet discarded. The solvent was removed using a Savant SpeedVac Plus (SC110A) over 2–4 h. The residue for each sample was then reconstituted to the original specimen volume using gelatin buffer (0.2% gelatin in borate buffer with 0.01% merthiolate and 0.1% Triton X-100). Using these techniques, peptide recovery is approximately 80%.
The radioimmunoassay design was similar to that previously reported [19]. Initially, hamster serum total iCT from gel filtration studies was determined by using an antiserum to the carboxyl-terminal portion of mature human CT, Ab4. This antiserum reacts with the CT molecule, whether it is in the free, amidated, 32-amino acid mature form, or within its precursor propeptides [ie procalcitonin, the conjoined calcitonin:calcitonin carboxypeptide-I (CT:CCP-1), or the free immature, unamidated CT]. Subsequent studies were performed with a new antibody, R1B4, which has ten times the crossreactivity of Ab-4 with the prohormone. The buffer was 0.2% gelatin (0.13 M H3BO3 containing 9 g NaCl, 2 g gelatin, 1 ml Triton-X 100 and 0.1 g merthiolate/l at pH = 7.5). The antiserum was preincubated with standards or unknowns (20–100 μ l) in 0.2 ml at 4°C for 4 days. After addition of 50 μ l 125I-hCT, and 200 μ l gelatin buffer, incubation was continued for 2 days. After adding 50 μ l goat anti-rabbit IgG bound to iron particles, incubation was continued in 0.5 ml for 1 day. Bound and free hormone were separated with magnetic tube racks. Maximum bound =37%; sensitivity =1g; 50% B/Bo =50pg.
Gel filtration
Similarly to work previously reported [20], constituted extracts, in 1–10 ml 0.2% gelatin or 0.2% HSA, were applied to calibrated 2.5 × 100 cm columns containing G-75 superfine Sephadex (Pharmacia Biotech, Piscataway, New Jersey, USA) suspended in 0.1% human serum albumin (1 g HSA, 0.1 mol NH4HCO3 and 0.1 g merthiolate/l at pH = 7.5) at 4°C. One hundred fractions (120 drops or 5.5 ml/tube) were collected during 48 h in 16× 100 mm borosilicate glass culture tubes. The void volume (VV) was based on the peak elution volume (EV) of blue dextran, and the salt volume (SV) was based on the peak EV of Na125I. The Kav for individual components was determined according to the formula: Kav = (EV–VV)/(SV–VV).
Results
Mortality
The mortality rates at 72 h for animals receiving progressively increasing doses of bacteria (n = 16/group) were 0, 20, 70, and 100%, respectively. Differences in mortality between all groups, including control animals (n = 17), were significant by Chi-square (P = 0.001). Furthermore, these values represent a direct relationship between the size of the inoculum of E coli and mortality (Fig 2).
Figure 2.
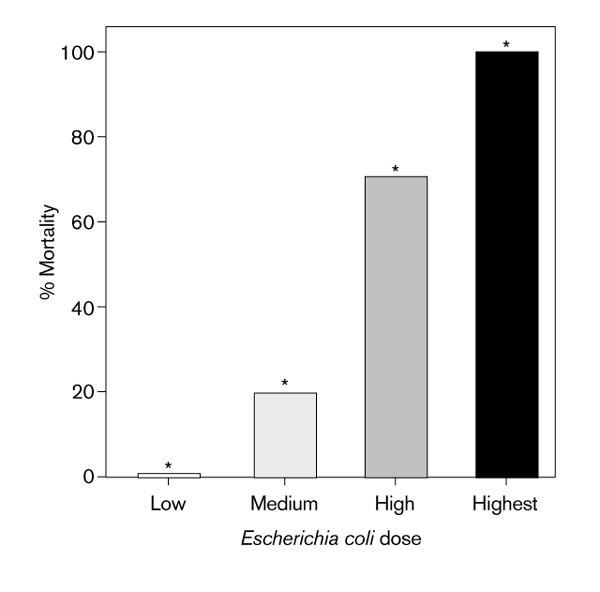
Relationship between inoculum of Escherichia coli and mortality. Low dose = 1.0 × 108 cfu/pellet, medium dose = 5.0 × 108 cfu/pellet, high dose = 1.0 × 109 cfu/pellet, and highest dose = 2.0 × 109 cfu/pellet. Mortality for low dose was 0%. *Significantly different from other groups per Chi-square analysis, P < 0.001.
Serum total iCT levels
Hamsters which were subjected to these graded doses of sepsis (n = 10/group) had serum total iCT levels at 12 h (mean ± SEM) of 250 ± 90, 380 ± 60, 1960 ± 490, and 4020 ± 510 pg/ml, respectively. Control animals (n =17) had serum total iCT levels of 21± 2 pg/ml. All groups were statistically distinct, except between 0 and 20% mortality (P = 0.001, Kruskal-Wallis one-way ANOVA; Fig 3).
Figure 3.
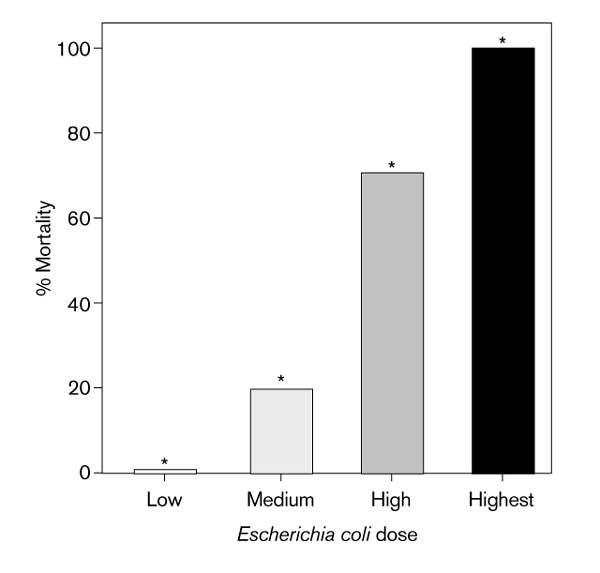
Relationship between inoculum of Escherichia coli and total immunoreactive calcitonin (iCT). Low dose = 1.0 × 108 cfu/pellet, medium dose = 5.0 × 108 cfu/pellet, high dose = 1.0 × 109 cfu/pellet, and highest dose = 2.0 × 109 cfu/pellet. *Statistically distinct, except between low and medium doses, per one-way ANOVA, P = 0.001.
Molecular species of the total serum iCT
The molecular species of the total serum iCT in the serum was determined by radioimmunoassay of fractions obtained from Sephadex gel filtration of pooled hamster sera as described above. The molecular mass of the predominant species of iCT measured (ie > 90%) was approximately 14 000 Da. From previous data [31], it is known that this fraction corresponds to ProCT, which in humans is 12795 Da. As shown in Fig 4, this molecular fraction in the hamster co-elutes with the ProCT fraction in the serum of a septic patient [17].
Figure 4.
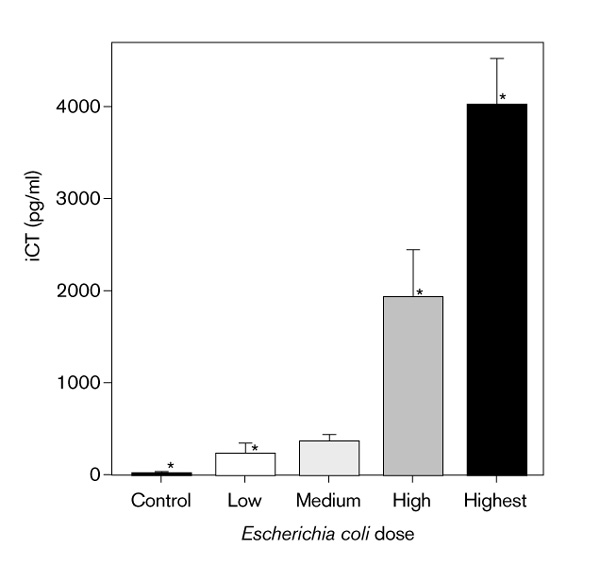
Comparison of chromatographs from septic human serum (a) and pooled septic hamster serum (b). The dominant peak in each graph has an estimated elution position of 0.2 KaV, which corresponds to the elution position of human procalcitonin (ProCT) [17]. CT, calcitonin; CT:CCP-1, conjoined calcitonin:calcitonin carboxypeptide-I.
Metabolic studies
Serum total iCT levels among groups exposed to a high dose of E coli (n = 13–15) and killed at 3, 6, 12 and 24 h increased from a baseline of 21 ± 2 pg/ml (mean ± SEM) to 78 ± 3, 542 ± 100, 3570 ± 920, and 4240 ± 1080 pg/ml, respectively. The changes in serum total iCT at all time points, except between 12 and 24 h, were statistically significant (one-way ANOVA, P = 0.001).
Total serum calcium levels at these timepoints were 11.6 ± 0.1, 12.1 ± 0.2, 9.4 ± 0.2, and 10.6 ± 0.4 mg/dl. The decrease at 12 h was statistically significant per MannWhitney rank sum test (P < 0.05). Simple linear regression reveals an inverse correlation between total calcium levels and total iCT (r = -0.81). Serum albumin levels varied minimally at 3, 6, 12 and 24 h from a baseline of 3.3 ± 0.1 g/dl, and therefore did not account for the decrease in measured calcium.
Discussion
The characteristics of the inflammatory response in sepsis suggest that successful treatment requires a clinically useful marker which can indicate the severity of illness and which is expressed early enough in the sepsis cascade to allow therapeutic interventions to be initiated in a timely manner [4]. Additionally, insights into the biosynthesis, regulation, and physiologic activity of such a marker may illuminate some of the causative factors in the pathophysiologic and clinical events of the sepsis syndrome. Furthermore, the marker itself may prove to be a therapeutic target.
Serum levels of ProCT as well as its component peptides are massively elevated in burns [11], heat stroke [21], systemic infections [13], and other inflammatory states [12,22]. Using an antiserum to CT which recognizes the free mature CT, the immature CT within the ProCT molecule, and the conjoined CT:CCP-1 peptide, we have demonstrated that levels of serum total iCT are also elevated in the septic hamster. Then, utilizing gel filtration techniques, we showed that much of this iCT was in the form of ProCT; this is similar to the human subject with sepsis [17,23]. Our findings indicate a positive correlation between ProCT component peptides and the degree of sepsis. In this model, the series of metabolic experiments furthermore reveal that ProCT is temporally associated with and inversely correlated with serum total calcium levels.
CT is a single chain, 32-amino acid peptide that originates from the CALC-I gene on chromosome 11 [16]. In humans the highest concentration of tissue iCT is in the parafollicular cells of the thyroid gland. However, iCT can be detected throughout the body in NE cells of various tissues. Indeed, in humans the lungs contain more total iCT than does the thyroid gland [24].
While mature CT has diverse effects on various target tissues, its overall physiologic significance in normal individuals is not well understood. In health, its principal role is to protect against excessive bone turnover during times of increased need by attenuating the activity of osteoclasts [25]. CT and its precursors, however, may exert other effects in health or in disease [16].
The polypeptide precursor of CT, pre-procalcitonin, undergoes cleavage of its leader sequence early in posttranslational processing to yield ProCT, and several constituent peptides (Fig 1). In normal, regulated secretion, ProCT is trafficked through the Golgi apparatus and then packaged into dense-core secretory vesicles [26,27]. Proteolytic processing within the trans-Golgi and the secretory vesicles culminates in the formation of the active, mature secretory product, CT, which is released by exocytosis at the apical surface of the NE cell. In the absence of an appropriate signal at the plasma membrane, these vesicles serve as storage repositories for mature CT.
In severe systemic inflammation in humans, however, enormous levels of ProCT and other component peptides appear in the serum, while mature serum CT remains normal or only minimally elevated [17]. The cellular source of this increase in serum levels, and the reasons that ProCT and its component peptides are not processed to the mature hormone, are unknown. In inflammatory states, ProCT and its related peptides appear to be secreted by a continuous bulk-flow constitutive pathway, in which only limited conversion to mature CT occurs [28]. One might postulate that severe inflammation causes such a profound hypersynthesis of the prohormone that the NE endoproteolytic machinery is overwhelmed. This may result in a marked shift to the constitutive pathway of secretion, resulting in an incomplete processing of precursors. In this respect, a shift to constitutive secretion has been reported to occur by the experimental induction of dysfunctional prohormone convertase enzymes or by injury to the plasma membrane [29]. Perhaps some cytokines may induce constitutive secretion by such a process [30,31]. It is also possible that ProCT and its component peptides are released by non-NE cells, which normally possess regulatory mechanisms limiting the expression of ProCT mRNA; these inhibitory mechanisms may be deregulated by unusually high levels of inflammatory mediators. Stimulation of synthesis in such non-NE cells would result in a preferential production of ProCT because these cells lack the enzymes for complete prohormonal processing.
It is unknown what impact, if any, this increase of ProCT and related peptides has on patients. Hypocalcemia is a common finding in critically ill and especially septic patients. Indeed, the development of hypocalcemia in the critically ill has been shown to be associated with a poor prognosis [32,33]. ProCT contains within its structure the immature CT molecule; therefore, very high and sustained levels of ProCT might mimic one of the pharmaco/physiologic activities of CT, which is the lowering of serum calcium levels. In our experiments we noted that total iCT levels peaked at 12h following the septic insult. This was concurrent with a significant decrease in serum total calcium. Nevertheless, this association does not prove a causal relationship between elevated ProCT levels and hypocalcemia. Also, the relationship with ionized calcium was not determined in this study.
The early and marked hypersecretion of ProCT and its component peptides in inflammatory states makes them promising serum markers for the sepsis syndrome. These peptides are released into the central circulation and may act systemically, as opposed to many of the known mediators of sepsis, which are released locally and often act in an autocrine or paracrine fashion. An important feature of ProCT and some of its component peptides are their long half-lives, which contribute to their potential usefulness as serum markers. Indeed, elevated levels of ProCT peptides have been found to persist at least 24 h following an appropriate stimulus, in contradistinction to other markers, such as tumor necrosis fctor-α, whose levels may be only transiently elevated after an inflammatory challenge [34,35]. Thus, they provide a long-lasting target to evaluate the effects of immunoneutralization. Accordingly, we recently reported that ProCT markedly contributes to mortality in experimental sepsis, and that immunoneutralization of this molecule diminishes mortality in our model of hamster sepsis [36].
In summary, our animal experiments demonstrate an association between levels of serum ProCT and its component peptides with the degree of sepsis, reinforcing clinical findings that these peptides are useful markers for this condition, and may predict mortality. Further experiments to examine the cellular source, pathophysiology and metabolic activity of ProCT and its component peptides are warranted. Such studies may determine the role of these hormonal peptides in inflammation and sepsis.
References
- American College of Chest Physicians — Society of Critical Care Consensus Conference Definitions of sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. Crit Care Med. 1992;20:864–874. [PubMed] [Google Scholar]
- Parillo JE. Septic shock in humans: advances in the understanding of pathogenesis, cardiovascular dysfunction, and therapy. . Ann Intern Med. 1990;113:227–242. doi: 10.7326/0003-4819-113-3-227. [DOI] [PubMed] [Google Scholar]
- Bone RC, Fisher CJ, Clemmer TP, et al. Sepsis syndrome: a valid clinical entity. Crit Care Med. 1989;17:389–393. [PubMed] [Google Scholar]
- Christman JW, Holden EP, Blackwell TS. Strategies for blocking the systemic effects of cytokines in the sepsis syndrome. Crit Care Med. 1995;25:955–963. doi: 10.1097/00003246-199505000-00027. [DOI] [PubMed] [Google Scholar]
- Zeigler EJ, Fisher CJ, Spriung CL, et al. Treatment of gram-negative bacteremia and septic shock with HA-IA human monoclonal antibody against endotoxin. N Engl J Med. 1991;324:429–436. doi: 10.1056/NEJM199102143240701. [DOI] [PubMed] [Google Scholar]
- Lowry SF. Sepsis and it complications: clinical definitions and therapeutic prospects. Crit Care Med. 1994;22:51–52. [PubMed] [Google Scholar]
- Bone RC. Toward a theory regarding the pathogenesis of the systemic inflammatory response syndrome. What we know and what we do not know about cytokine regulation. Crit Care Med. 1996;24:163–172. doi: 10.1097/00003246-199601000-00026. [DOI] [PubMed] [Google Scholar]
- Rutledge R, Fakhry S, Rutherford E, Muakkassa F, Meyer A. Comparison of APACHE II, Trauma Score and Injury Severity Score as predictors of outcome in critically injured patients. Am J Surg . 1993;166:244–247. doi: 10.1016/s0002-9610(05)80966-3. [DOI] [PubMed] [Google Scholar]
- Cerra FB, Negro F, Abrams J. APACHE II score does not predict multiple organ failure or mortality in postoperative surgical patients. . Arch Surg. 1990;125:519–522. doi: 10.1001/archsurg.1990.01410160105021. [DOI] [PubMed] [Google Scholar]
- Meyer AA, Messick J, Young P, et al. Prospective comparison of clinical judgment and APACHE II score in predicting the outcome in critically ill surgical patients. J Trauma. 1992;32:747–753. doi: 10.1097/00005373-199206000-00013. [DOI] [PubMed] [Google Scholar]
- Nylen E, O'Neill W, Jordan M, et al. Serum procalcitonin as an index of inhalation injury in burns. Horm Metab Res. 1992;24:439–443. doi: 10.1055/s-2007-1003354. [DOI] [PubMed] [Google Scholar]
- Assicot M, Gendrel D, Carsin H, et al. High serum calcitonin concentrations in patients with sepsis and infection. . Lancet. 1993;341:515–518. doi: 10.1016/0140-6736(93)90277-N. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whang KT, Steinwald PM, White JC, et al. Serum calcitonin percursors in sepsis and systemic inflammation. J Clin Endocrinol Metab. 1998;83:3296–3301. doi: 10.1210/jcem.83.9.5129. [DOI] [PubMed] [Google Scholar]
- Stevenson JC, Hillyard CJ, MacIntyre I, Cooper H, Whitehead MI. A physiologic role for calcitonin: protection of the maternal skeleton. Lancet. 1979;ii:769–770. doi: 10.1016/s0140-6736(79)92117-2. [DOI] [PubMed] [Google Scholar]
- Nylen ES, Linnoila I, Snider RH, Tabassian AR, Becker KL. Comparative studies of hamster calcitonin from pulmonary endocrine cells in vitro. . Peptides. 1987;8:977–982. doi: 10.1016/0196-9781(87)90124-0. [DOI] [PubMed] [Google Scholar]
- Becker KL, Nylen ES, Cohen R, Snider RH. Calcitonin: structure, molecular biology, and actions. Bilezikian J, Raisz LG, Rodan GA (editors). Principles of Bone Biology. San Diego, California: Academic Press, 1996. pp. 471–493.
- Snider RH, Nylen ES, Becker KL. Procalcitonin and its component peptides in systemic inflammation: immunochemical characterization. . J Invest Med. 1997;45:552–560. [PubMed] [Google Scholar]
- Dunne JR, Dunkin BJ, Nelson S, White JC. Effects of granulocyte colony stimulation factor in a non-neutropenic rodent model of Escherichia coli peritonitis. J Surg Res . 1996;61:348–354. doi: 10.1006/jsre.1996.0128. [DOI] [PubMed] [Google Scholar]
- Snider RH, Moore CF, Silva OL, Becker KL. Radioimmunoassay of calcitonin in normal human urine. Anal Chem. 1978;50:449–454. doi: 10.1021/ac50025a023. [DOI] [PubMed] [Google Scholar]
- Snider RH, Silva OL, Moore CF, Becker KL. Immunochemical heterogeneity of calcitonin in man: effect on radioimmunoassay. . Clin Chim Acta. 1977;76:1–14. doi: 10.1016/0009-8981(77)90113-9. [DOI] [PubMed] [Google Scholar]
- Nylen ES, Arifi A, Becker KL, Snider RH, Alzeer A. The effect of classical heat stroke on serum procalcitonin. Crit Care Med. 1997;25:1362–1365. doi: 10.1097/00003246-199708000-00024. [DOI] [PubMed] [Google Scholar]
- Davis TME, Assicot M, Bohuon C, et al. Serum procalcitonin concentrations in acute malaria. Trans Roy Soc Trop Med Hyg. 1994;88:670–671. doi: 10.1016/0035-9203(94)90221-6. [DOI] [PubMed] [Google Scholar]
- Nylen ES, Jordan M, O'Neil W, et al. Altered molecular heterogeneity of serum calcitonin: an index of inhalational burn injury 23rd Annual Meeting of the American Burn Association April Baltimore, MD [abstract 194] 1991.
- Becker KL, Snider RH, Moore CF, et al. Calcitonin in extrathyroidal tissues in man. Acta Endocrinol (Copenhagen) . 1979;92:746–751. doi: 10.1530/acta.0.0920746. [DOI] [PubMed] [Google Scholar]
- Zaidi M, Bax BE, Shankar VS, et al. Dimensional analysis of osteoclastic bone resorption and the measurement of biologically active calcitonin. Exp Physiol. 1994;79:387–399. doi: 10.1113/expphysiol.1994.sp003773. [DOI] [PubMed] [Google Scholar]
- Rothman JE, Orci L. Movement of proteins through the Golgi stack: a molecular dissection of vesicular transport. FASEB J . 1990;4:1460–1468. doi: 10.1096/fasebj.4.5.2407590. [DOI] [PubMed] [Google Scholar]
- Treilhou-Lahille F, Pidoux E, Day F, et al. Granular and extragranular forms of immunoreactive calcitonin in normal rat 'C' cells. Biology of the Cell. 1986;57:221–230. doi: 10.1111/j.1768-322x.1986.tb00477.x. [DOI] [PubMed] [Google Scholar]
- Burgess TL, Kelly RB. Constitutive and regulated secretion of proteins. Ann Rev Cell Biol. 1987;3:243–293. doi: 10.1146/annurev.cb.03.110187.001331. [DOI] [PubMed] [Google Scholar]
- Bershadsky AD, Futerman AH. Disruption of the Golgi apparatus by brefeldin A blocks polarization and inhibits directed cell migration. Proc Natl Acad Sci. 1994;91:5686–5689. doi: 10.1073/pnas.91.12.5686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd JC, Mollitt DL. Sepsis-induced alterations in the erythrocyte membrane. Am Surg. 1994;60:954–957. [PubMed] [Google Scholar]
- Madara JL, Stafford J. Interferon-γ directly affects barrier function of cultured intestinal epithelial monolayers. . J Clin Invest. 1989;83:724–727. doi: 10.1172/JCI113938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chernow B, Zaloga G, McFadden E, et al. Hypocalcemia in critically ill patients. Crit Care Med. 1982;10:848–851. doi: 10.1097/00003246-198212000-00008. [DOI] [PubMed] [Google Scholar]
- Desai TK, Carlson RW, Geheb MA. Prevalence and clinical implications of hypocalcemia in acutely ill patients in a medical intensive care setting. Am J Med. 1988;84:209–214. doi: 10.1016/0002-9343(88)90415-9. [DOI] [PubMed] [Google Scholar]
- Dandona P, Nix D, Wilson MF, et al. Procalcitonin increase after endotoxin injection in normal subjects. J Clin Endocrinol Metab. 1994;79:1605–1608. doi: 10.1210/jcem.79.6.7989463. [DOI] [PubMed] [Google Scholar]
- Becker KL, Whang KT, Snider RH, et al. Relation between cytokines and procalcitonin peptides in an animal model of infection. . Ann Endocrinol. 1998;59:170. [Google Scholar]
- Nylen ES, Whang KT, Snider RH, Steinwald PM, White JC, Becker KL. Mortality is increased by procalcitonin and decreased by an antiserum reactive to procalcitonin in experimental sepsis. Crit Care Med. 1998;26:1001–1006. doi: 10.1097/00003246-199806000-00015. [DOI] [PubMed] [Google Scholar]