

Abstract
TDP-43 is characteristically accumulated in TDP-43 proteinopathies such as frontotemporal lobar degeneration and motor neurone disease, but is also present in some tauopathies, including Alzheimer’s disease, argyrophilic grain disease, and corticobasal degeneration (CBD). However, several studies have suggested that cases of progressive supranuclear palsy (PSP) lack TDP-43 pathology. We have therefore examined limbic regions of the brain in 19 PSP cases, as well as in 12 CBD cases, using phosphorylation-dependent anti-TDP-43 antibodies. We observed TDP-43-positive inclusions in five PSP cases (26%), as well as in two CBD cases (17%). The amygdala and hippocampal dentate gyrus were most frequently affected in PSP. Regional tau burden tended to be higher in TDP-43-positive PSP cases, and a significant correlation between tau and TDP-43 burden was noted in the occipitotemporal gyrus. Hippocampal sclerosis (HS) was found in 3/5 TDP-43-positive PSP cases, but HS was significantly more frequent in TDP-43-positive than TDP-43 negative PSP cases. Dementia was present in 13/19 (58%) of the PSP cases, in 4/5 TDP-43-positive cases, in all 3 TDP-43-positive cases with HS, in 1/2 TDP-43-positive cases without HS, and 7/14 cases lacking both. TDP-43 and tau were frequently colocalized in the amygdala, but not in the hippocampal dentate gyrus. Immunoblotting demonstrated the characteristic (for TDP-43 proteinopathies) 45 and 25 kDa bands and high molecular weight smear in the TDP-43-positive PSP case. These findings suggest that (1) although PSP is nominally a tauopathy, pathological TDP-43 can accumulate in the limbic system in some cases, and (2) TDP-43 pathology may be concurrent with HS.
Keywords: Argyrophilic grains, Hippocampal sclerosis, Progressive supranuclear palsy, Tau, TDP-43
Introduction
Transactivation-responsive DNA-binding protein of Mr 43 kDa (TDP-43) is a nuclear protein involved in transcriptional repression and alternative splicing. It was originally identified as a major component of ubiquitin-positive and tau-negative inclusions in the frontotemporal cortex and motor neurons in frontotemporal lobar degeneration (FTLD-U), with or without progranulin gene mutations, and in amyotrophic lateral sclerosis (ALS) [3, 12, 31]. Subsequent studies revealed that TDP-43 is also abnormally accumulated in familial FTLD-U with mutations in the valosin-containing protein gene [32], in familial FTLD with motor neuron disease linked to chromosome 9p [10], and in ALS with TDP-43 gene mutations [25, 38, 41, 44]. TDP-43 is considered to play an essential pathogenic role in these diseases, now-called TDP-43 proteinopathies.
Although TDP-43 accumulation was originally considered to be a specific disease marker for FTLD-U and ALS, subsequent studies demonstrated that abnormal TDP-43 accumulation in some cases of other neurodegenerative diseases, such as Alzheimer’s disease (AD) [2], Parkinson’s disease with and without dementia [30], dementia with Lewy bodies (DLB) + AD [4, 30], ALS/parkinson-dementia complex of Guam (ALS/PDC of Guam) [15, 16], argyrophilic grain disease (AGD) [14], and Huntington disease [37]. However, the pathophysiological significance of concurrent TDP-43 accumulation, and its impact on clinical phenotype in these diseases remain unclear.
Several previous studies have suggested that cases of progressive supranuclear palsy (PSP) lack abnormal TDP-43 accumulation [3, 18, 40]. In these early studies, phosphorylation-independent antibodies were employed in TDP-43 immunohistochemistry and immunoblot analysis. We have made polyclonal and monoclonal antibodies specific for phophorylated TDP-43, which identify phosphorylation sites in the C-terminus of the TDP-43 accumulated in FTLD-TDP brains [17, 20], and selectively immunolabel pathological inclusions and dystrophic neurites without physiological nuclear staining in FTLD-TDP, ALS, AD with TDP-43 pathology, and in DLB with TDP-43 [4, 17]. They also recognize hyperphosphorylated TDP-43 at 45 kDa and additional 18–26 kDa fragments in sarkosyl-insoluble fractions on immunoblotting.
The principal aim of this study was to revisit the presence or absence, and the frequency, of TDP-43 pathology in PSP cases using a phosphorylation-dependent anti-TDP-43 antibody. In contrast to previous reports, we demonstrated that a significant proportion of PSP cases had variable degrees of TDP-43 pathology in the limbic system. We subsequently examined the relationships between TDP-43 pathology, tau pathology, and hippocampal sclerosis, as well as biochemical nature of the abnormally accumulated TDP-43, in PSP.
Materials and methods
Subjects
We investigated 19 pathologically confirmed PSP cases, 12 pathologically confirmed corticobasal degeneration (CBD) cases and 4 pathologically normal control subjects (Table 1). These cases were obtained from UK Parkinson’s Disease Society Tissue Bank (7 PSP and 4 control cases), Department of Pathology, Northwestern University Feinberg School of Medicine Cognitive Neurology and Alzheimer Disease Center (5 PSP and 7 CBD cases), and Department of Neuropsychiatry, Okayama University Graduate School of Medicine, Dentistry and Pharmaceutical Sciences (7 PSP and 5 CBD cases). All brains had been collected with Local Research Ethical Committee approval. All PSP cases showed characteristic tufted astrocytes, and all CBD cases astrocytic plaques, as revealed by Gallyas-Braak silver methods and tau immunohistochemistry.
Table 1.
Demographic data in PSP and CBD cases with and without TDP-43 pathologies
| PSP |
CBD |
|||||
|---|---|---|---|---|---|---|
| All | TDP-43-positive PSP | TDP-43-negative PSP | All | TDP-43-positive CBD | TDP-43-negative CBD | |
| N (%) | 19 | 5 (26.3) | 14 (73.7) | 12 | 2 (16.7) | 10 (83.3) |
| Male [N (%)] | 16 (84.2) | 4 (80.0) | 12 (85.7) | 7 (58.3) | 1 (50.0) | 6 (60.0) |
| Age at onset [mean (SD)] | 68.3 (9.8) | 75.0 (9.4) | 65.7 (9.0) | 55.2 (10.2) | 49.0 (12.7) | 56.6 (9.9) |
| Age at death [mean (SD)] | 76.3 (10.7) | 82.4 (11.7) | 74.1 (9.8) | 62.8 (11.2) | 56.0 (15.6) | 64.1 (10.7) |
| Duration [mean (SD)] | 7.4 (4.4) | 7.4 (4.6) | 7.5 (4.6) | 7.3 (2.9) | 7.0 (2.8) | 7.3 (3.1) |
| Dementia (%) | 11 (57.9) | 4 (80.0) | 7 (50.0) | 11 (91.7) | 2 (100.0) | 9 (90.0) |
| Brain weight [g, mean (SD)] | 1,202 (142) | 1,234 (180) | 1,190 (132) | 1,174 (146) | 1,008 (152) | 1,215 (120) |
| Argyrophilic grains [N (%)] | 4 (21.1) | 1 (20.0) | 3 (21.4) | 3 (25.0) | 1 (50.0) | 2 (20.0) |
| Hippocampal sclerosis [N (%)] | 3 (15.8) | 3 (60.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) |
Immunohistochemistry
Sections cut at 5-μm thickness to include the amygdala, entorhinal cortex, hippocampus, occipitotemporal cortex in all cases, as well as the substantia nigra in two cases for which tissue was available, were stained with antibodies against phosphorylated TDP-43 (pAb pS409/410, rabbit, polyclonal, 1:1,000 [17]), phosphorylated tau (AT8, mouse, monoclonal, 1:3,000, Innogenetics, Ghent, Belgium), phosphorylated α-synuclein (#1175, rabbit, polyclonal, 1:1,000, [33]), and Aβ (4G8, mouse, monoclonal, 1:2,000, Covance Research Products Inc., Dedham, MA, USA). Deparaffinized sections were incubated with 1% H2O2 in methanol for 20 min to eliminate endogenous peroxidase activity in the tissue. When using anti-α-synuclein and anti-TDP-43 antibodies, sections were pretreated to enhance immunoreactivity in a microwave oven for 5 min in 10 mM sodium citrate buffer, pH 6.0, at 100°C. After blocking with 10% normal serum, sections were incubated 1 h at room temperature with the primary antibody. After three 5-min washes in phosphate-buffered saline (PBS), sections were incubated in biotinylated secondary antibody for 30 min, and then in avidin-biotinylated horseradish peroxidase complex (ABC Elite kit, Vector, Burlingame, CA, USA) for 30 min. The peroxidase labeling was visualized with 0.2% 3,3′-diaminobenzidine (DAB) as chromogen. Sections were lightly counterstained with hematoxylin.
Semiquantitative assessment
TDP-43, tau, and Aβ pathologies in the amygdala, anterior and posterior portions of the entorhinal cortex, hippocampal dentate gyrus, CA1, 2, 3, and 4 regions, subiculum, fusiform gyrus, occipitotemporal gyrus were semiquantitatively evaluated using the following grading system blinded to any clinical or pathological information:
The total number of TDP-43-positive neuronal cytoplasmic inclusions (NCIs) in each anatomical region was assessed as follows: − no lesion, + one inclusion, ++ two or three inclusions, +++ four or five inclusions, ++++ 6–10 inclusions, +++++ 11 or over inclusions. In addition, the presence or absence of neuronal intranuclear inclusions (NIIs) and dystrophic neurites was also assessed. Then, we classified the topographic distribution of TDP-43 pathological changes using following system, which is similar to that reported by Amador-Ortiz et al. [2]: the amygdala type: inclusions were present only in the amygdala; the limbic type: inclusions extend to the amygdala, hippocampal dentate gyrus, CA1-4, entorhinal cortex, and fusiform gyrus, but not in the occipitotemporal gyrus; the temporal type: inclusions are present in the limbic system and also the in the occipitotemporal gyrus.
Tau-positive neuronal inclusions were counted in low power microscopic fields: 0, no tau-positive lesions; 1, one neuronal inclusion per few microscopic fields; 2, one inclusion in every field; 3, 4–30 inclusions in every field; 4, over 30 inclusions associated with numerous neurites in every field.
Aβ deposits were counted in low power microscopic fields: 0, no Aβ deposits; 1, two to three Aβ plaques in each field; 2, 4–10 Aβ plaques in each field; 3, 11–20 Aβ plaques in each field; 4, more than 20 Aβ deposits in each field.
Hippocampal sclerosis (HS) was defined by neuronal loss with gliosis in the hippocampal CA1 and/or subiculum, with relatively preserved neurons in the CA4, 3, and two regions and absence of intracellular and extracellular NFTs, or ischaemic changes that might explain neuronal loss in the CA1 and subiculum. HS was assessed blind to any clinical or pathological information.
Statistical analysis
The Mann–Whitney U test and Fisher’s exact test were used to compare the demographic and pathological data between TDP-43-positive and TDP-43-negative groups in PSP and CBD series, respectively. Correlations between ratings of TDP-43 pathology and demographic data, or ratings of tau and Aβ pathologies in each anatomical region were assessed with Spearman’s rank-order correlation statistic. Statistical analysis was performed using StatView for Macintosh program, version J-4.5. A value of p < 0.05 was accepted as significant.
Confocal laser scanning microscopy
Double-labeling immunofluorescence was performed with the combination of phosphorylation-dependent anti-TDP-43 (pAb pS409/410, rabbit, polyclonal, 1:1,000 [17]) and anti-tau antibodies (AT8, mouse, monoclonal, 1:500, Innogenetics, Ghent, Belgium). Sections from the amygdala and hippocampus in some PSP cases with TDP-43 pathology were pretreated by heating in a microwave oven for 5 min in 10 mM sodium citrate buffer, pH 6.0, at 100°C, allowed to cool then permeabilized with 0.2% (v/v) Triton X-100 in PBS. Following washing in PBS, non-specific antibody binding was blocked with normal sera and sections were incubated with a mixture of the two primary antibodies for 1 h at room temperature. After washing in PBS, sections were incubated with fluorescence-labeled secondary antibodies [AlexaFluor 488 anti-rabbit IgG (1:200) and AlexaFluor 555 anti-mouse IgG (1:200), Molecular Probes, Invitrogen, Paisley. UK]. After washing with PBS, sections were incubated with Toto-3 Iodide (Molecular Probes, Invitrogen, Paisley, UK) with 1 mg/ml RNase (Roche Diagnostics GmbH, Manheim, Germany) at 37°C. To quench (lipofuscin) autofluorescence, sections were incubated in 0.1% Sudan Black B for 10 min at room temperature and washed with 0.1% Tx-PBS for 30 min. Sections were coverslipped with Vectashield mounting media (Vector Laboratories Inc., Burlingame, CA, USA). Images were collected on a Leica TCS SP5 AOBS upright confocal (Leica Microsystems, Milton Keynes, UK) using the 488 nm (19%), 543 nm (30%) and 633 nm (60%) laser lines, respectively. To eliminate cross-talk between channels, the images were collected sequentially.
Immunoblotting
Frozen tissue from the amygdala, hippocampus, and frontal, temporal, and occipital cortices in one PSP case with TDP-43 pathology, one FTLD-TDP case (as a positive control) and eight negative controls (six PSP, one LBD, and one pathologically normal case) were prepared for western blotting according to methods previously described by Neumann et al. [31]. Briefly, 1 g of fresh frozen brain was homogenized in 5 ml/g (w/v) of low salt (LS) buffer-containing 10 mM Tris pH 7.5, 5 mM EDTA pH 8.0, 1 mM DTT, 10% (w/v) sucrose and Roche complete EDTA-free protease inhibitor. Homogenates were sequentially extracted with increasing strength buffers [Triton X-100 buffer (LS buffer + 1% Triton X-100 + 0.5 M NaCl), Triton X-100 buffer with 30% sucrose to float myelin, Sarkosyl buffer (LS buffer + 1% N-lauroyl-sarcosine + 0.5 M NaCl)]. Detergent-insoluble pellets were extracted in 0.25 ml/g Urea buffer (7 M Urea, 2 M Thiourea, 4% 3-[(3-Cholamidopropyl) dimethylammonio]-1-propanesulfonate (CHAPS), 30 mM Tris–HCl pH 8.5, Roche complete EDTA free protease inhibitor. Prior to SDS-PAGE immunoblot analysis, urea fractions were added in 1:1 ratio to SDS sample buffer (10 mM Tris pH 6.8, 1 mM EDTA, pH 8.0, 40 mM DTT, 1% SDS, 10% Sucrose, 0.01% Bromophenol Blue). Protein was resolved on 12% Tris–Glycine SDS-PAGE gels along with size standard (Bio-Rad kaleidoscope broad-range marker; Bio-Rad, Hercules, CA, USA). Proteins were transferred onto nitrocellulose membrane (Hybond ECL, GE Life Sciences, UK) and blocked for 1 h at 4°C in 5% (w/v) milk solution [5% powdered milk in Tris-buffered saline containing 0.1% Tween-20 (TBS-T)]. Membranes were incubated in phosphorylation-dependent mouse monoclonal antibody (mAb pS409/410, mouse, 1:1,000 [20]) for 1 h at room temperature followed by HRP-conjugated goat anti-mouse secondary antibody (Santa Cruz Biotechnology Inc, CA, USA). Antibodies were visualized by incubating in enhanced chemiluminescent reagent (ECL, GE Life Sciences) and imaged using the ImageQuant 350 system fitted with a F0,95 25 mm Fixed Lens (GE Healthcare, Life Sciences, UK). TDP-43 probed membranes were exposed for 5 min at different timeframes to obtain multiple images of differing intensity. Images were processed using Image-Quant TL software (GE Healthcare, Life Sciences, UK).
Results
Frequency and distribution of TDP-43 pathology
Clinical and pathological features for all subjects are shown in Table 1. TDP-43 pathology was noted in 5 of 19 PSP cases (26%) and in 2 of 12 CBD cases (17%). Disease duration, gender ratio and brain weight were not statistically different between PSP cases with and without TDP-43 pathology, or between CBD cases with and without TDP-43 pathology, respectively. Age at onset of disease (75 vs. 66 years) and age at death (82 vs. 74 years) tended to be higher, and dementia occurred more often, in PSP cases with TDP-43 pathology than in PSP cases without it (80 vs. 50%), although these differences did not reach statistical significance. One PSP case without TDP-43 pathology also had Lewy body pathology corresponding to brainstem-predominant type [26]. Ten PSP cases (3 TDP-43-positive and 7 TDP-43-negative cases) and four CBD cases (all were TDP-43-negative) had Aβ-positive diffuse plaques in the amygdala, hippocampus, and/or temporal cortex. Of the three TDP-43-positive PSP cases, one case had only a few neuritic plaques in the occipitotemporal gyrus. None of the PSP or CBD cases in our series fit the pathological criteria of AD [9, 28, 39].
In PSP cases, TDP-43-positive NCIs were most frequently noted in the amygdala and dentate gyrus granule cells in the hippocampus (5 cases, 100% of TDP-43-positive PSP cases), followed by the anterior portion of the entorhinal cortex (4 cases, 80%), subiculum (3 cases, 60%), posterior portion of the entorhinal cortex (3 cases, 60%), occipitotemporal gyrus (2 cases, 50%), fusiform gyrus (2 cases, 40%), and CA1 region (2 cases, 20%) (Table 2, Fig. 1a–d). In addition to the rounded inclusions noted in FTLD-TDP, all PSP cases had many irregular shaped NCIs, such as flame-shape NFT-like, globose-type NFT-like, and horseshoe-like inclusions (Fig. 1e–j). One PSP case (PSP 2 in Table 2) showed a few NII in the subiculum (Fig. 1k, l). Two cases (PSP4 and PSP5) had abundant fine, short, thread-like structures immunopositive for TDP-43 from the CA1 to subiculum (Fig. 1m). TDP-43-positive thread-like structures were also observed in the amygdala (3 cases), entorhinal cortex (2 cases), CA1 (one case), and subiculum (one case) (Fig. 1n–p).
Table 2.
Distribution of TDP-43 pathology in PSP and CBD cases
| TDP-43 pathology |
Hippocampal sclerosis (CA1/Subiculum) | Argyrophilic grainsb | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | Amygdala | ant.EC | DG | CA3/4 | CA2 | CA1 | SB | post.EC | FG | OTG | TDP-43 distributiona | ||
| PSP cases | |||||||||||||
| PSP1 | ++++ | – | + | – | – | – | – | – | – | – | Limbic | – | – |
| PSP2 | +++++ | +++++ | +++++ | – | – | – | + | – | – | n | Limbic | + | – |
| PSP3 | +++++ | +++++ | +++ | – | – | – | ++ | ++++ | – | – | Limbic | – | – |
| PSP4 | +++++ | +++++ | ++ | – | – | + | +++ | +++++ | +++++ | + | Temporal | + | – |
| PSP5 | +++++ | +++ | +++++ | – | – | ++ | – | ++ | +++ | +++ | Temporal | + | Stage III |
| % | 100.0 | 80.0 | 100.0 | 0.0 | 0.0 | 20.0 | 60.0 | 60.0 | 40.0 | 50.0 | 60.0 | 20.0 | |
| CBD cases | |||||||||||||
| CBD1 | +++ | ++ | – | – | – | – | – | n.a. | n.a. | n.a. | Limbic | – | – |
| CBD2 | +++++ | ++ | ++++ | ++ | – | +++++ | +++++ | ++ | – | – | Limbic | – | Stage II |
| % | 100.0 | 100.0 | 50.0 | 50.0 | 0.0 | 50.0 | 50.0 | 50.0 | 0.0 | 0.0 | 0.0 | 50.0 | |
The stages of TDP-43 pathology: –, no lesion in the anatomical region; +, 1 inclusion in the anatomical region; ++, 2–3 inclusions in the anatomical region; +++, 4–5 inclusion in the anatomical region; ++++, 6–10 inclusions in the anatomical region; +++++, 11 or over inclusions in the anatomical region. The stage of hippocampal sclerosis: –, no; +, mild; ++, moderate; +++, severe. The stage of argyrophilic grains: –, absent; +, present. ant.EC, the anterior portion of the entorhinal cortex; DG, hippocampal dentate gyrus; SB, subiculum; post.EC, the posterior portion of the entorhinal cortex; FG, fusiform gyrus; OTG, occipitotemporal gyrus
The amygdala type: inclusions were present only in the amygdala; the limbic type: inclusions extend to the limbic system, but not in the occipitotemporal gyrus; the temporal type: inclusions are present in the limbic system and occipitotemporal gyrus as well
The distribution of argyrophilic grains are assessed using a staging system proposed by Saito et al. [35]
Fig. 1.

TDP-43-positive lesions in PSP. a Neuronal cytoplasmic inclusions (NCIs) in the hippocampal dentate gyrus. b–d NCIs in the entorhinal cortex. Irregular shaped NCIs in the entorhinal cortex (e), fusiform gyrus (f), and subiculum (g). These inclusions have weakly stained or unstained regions. Small dot-like structures are also seen in the neuronal cytoplasm (g). Horseshoe-shaped (h, i) and NFT-like (j) NCIs in the entorhinal cortex. Intranuclear inclusions in the amygdala (k) and in the subiculum (l), cases PSP3 and PSP2, respectively. m Massive short threads-like structures in the subiculum, case PSP3. n Thick, thread-like structures in the amygdala. o, p Long, thin thread-like structures in the amygdala. pAb pS409/410 immunohistochemistry. All scale bars 20 μm
In two CBD cases, TDP-43-positive NCIs were observed in the amygdala, entorhinal cortex, hippocampal dentate gyrus, CA1, CA3/4, and subiculum (Table 2, Fig. 2a). The distribution of TDP-43 pathology was roughly consistent with that observed in PSP cases. NIIs were found in the subiculum and amygdala in one CBD case with severe TDP-43 pathology (Fig. 2b, c). Short thread-like structures immunopositive for TDP-43 were found in the amygdala, entorhinal cortex, CA1, CA3, and/or subiculum in both CBD cases with TDP-43 pathology. One CBD case had TDP-43-positive coiled body-like structures and thread-like structures in the alveus in the subiculum (Fig. 2d–f). Abnormal accumulation of TDP-43 was not found in the white matter of the temporal lobe and substantia nigra in any of the TDP-43-positive PSP or CBD cases.
Fig. 2.
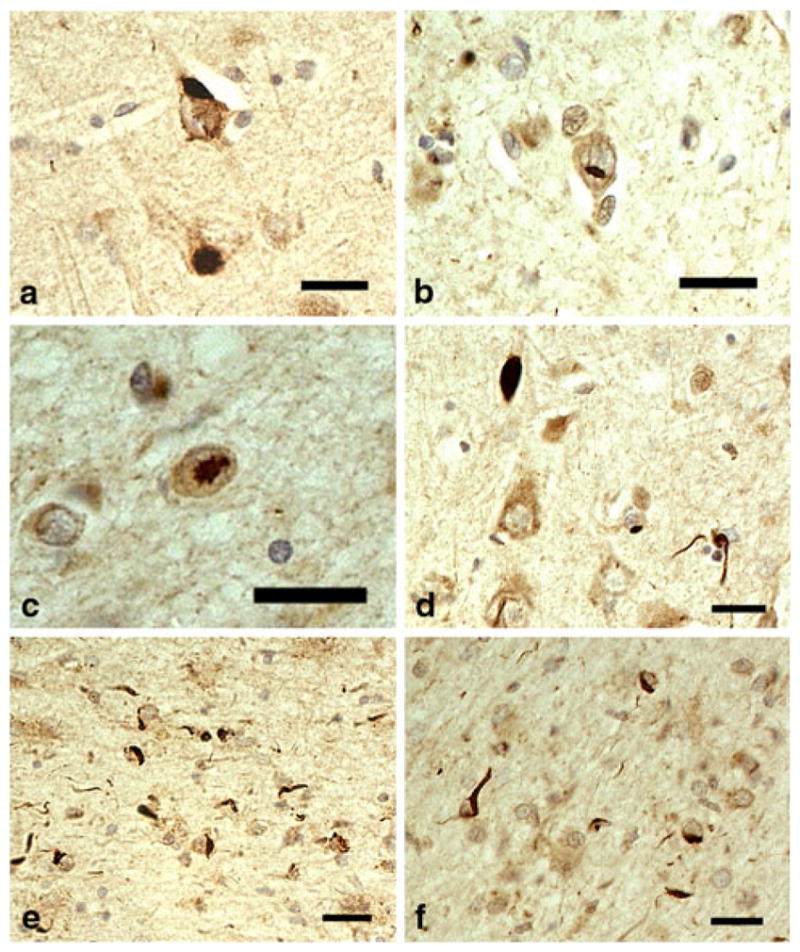
TDP-43-positive lesions in CBD. a Neuronal cytoplasmic inclusions (NCIs) in CA3 region of hippocampus. b, c Neuronal intranuclear inclusions in the amygdala. d A thick neurite and thin, thread-like structures in the amygdala. e Short thread-like structures and glial cytoplasmic inclusions (GCIs) in the alveus in the entorhinal cortex. f Coiled body-like structures and GCis in the alveus in the entorhinal cortex. pAb pS409/410 immunohistochemistry. All scale bars 20 μm
Relationship between TDP-43 pathology and tau or Aβ burden
The ratings for tau burden in the TDP-43-positive PSP cases tended to be higher (but not significantly so) than those in the TDP-43-negative PSP cases, in almost all regions examined (i.e., including amygdala, entorhinal cortex, hippocampal dentate gyrus, CA1-4, fusiform gyrus, and occipitotemporal gyrus) (Fig. 3). In the PSP cases overall, rating for tau pathology in the occipitotemporal gyrus was significantly correlated with that of TDP-43 pathology (r = 0.504, p < 0.05), but no significant correlations between tau and TDP-43 ratings were found in any other regions. There were no significant differences in the degree of Aβ burden in any region between TDP-43-positive and TDP-43-negative PSP cases, and ratings for TDP-43 pathology did not correlate with those for Aβ burden in any region. Of three TDP-43-positive PSP cases having Aβ deposits, only one case had a few neuritic plaques in the occipitotemporal gyrus; however, this case did not have any TDP-43-positive inclusions in the region.
Fig. 3.

Tau burden in the limbic system in PSP cases with and without TDP-43 pathology. In all regions but the subiculum, tau burden in PSP cases with TDP-43 pathology is more severe than that in PSP cases without TDP-43 pathology. Stage 0–1, no to mild tau deposition; stages 2–3, moderate to severe tau deposition; stage 4, very severe tau deposition (see detailed definition in the text). TDP-43+ TDP-43-positive, TDP-43− TDP-43-negative, Amy amygdala, antERC the anterior portion of the entorhinal cortex, DG hippocampal dentate gyrus, Subi subiculum, postERC the posterior portion of the entorhinal cortex, FG fusiform gyrus, OTG occipitotemporal gyrus
In the CBD cases, there were no significant differences in tau or Aβ burden in any region between TDP-43-positive and TDP-43-negative cases, and ratings for TDP-43 pathology did not correlate with those for tau or Aβ burden in any region.
Relationship of HS, argyrophilic grains, TDP-43 accumulation, and dementia
In 3 of 19 PSP cases (16%), evident neuronal loss in the CA1 and subiculum consistent with HS was noted (Fig. 4a, b). No CBD case showed HS. All three PSP cases with HS had a various degrees of TDP-43 pathology in the CA1 and/or subiculum (Fig. 4e–h), and two had extensive TDP-43 pathology in the limbic system: one case had both TDP-43 pathology and argyrophilic grains (Table 2). Two of the three PSP cases with HS had a few AT8-positive pretangles and argyrophilic grains in the CA1 and subiculum (Fig. 4b–d). Neurofibrillary tangles were rare in these regions in all PSP cases with HS (Fig. 4b). No significant ischemic changes in the hippocampal pyramidal neurons, or neuronal loss in the end plate, suggestive of a past history of severe epilepsy was noted in any of the PSP cases with HS. The frequency of HS in the TDP-43-positive PSP cases was significantly higher than that in TDP-43-negative PSP cases (60 vs. 0%, p = 0.021). Dementia was present in all of the 3 TDP-43-positive PSP cases with HS (100%), 4 of the 5 TDP-43-positive PSP cases with and without HS (80%), 1 of 2 TDP-43-positive PSP cases without HS (50%), and 7 of 14 PSP cases lacking both (50%). The frequency of dementia was not significantly different between PSP cases with and without HS (p = 0.170).
Fig. 4.

Pathological features in the hippocampus in a PSP case with TDP-43, HS, and argyrophilic grains (PSP5). a A low power view of the hippocampal CA1 to subiculum. Severe reduction of the width with tissue rarefaction is noted in the subiculum (arrow) and to a lesser degree in the adjacent CA1 region (arrowhead). b A moderate power view of the subiculum on the same section as that shown in a. Severe neuronal loss associated with gliosis is evident. Argyrophilic threads and grains are scattered, but tangles are rare. c The subiculum on an adjacent section of b. A moderate number of tau-positive threads and grains, but only a few tangles, are seen. d Argyrophilic grains in CA1 region. e, f TDP-43-positive cytoplasmic inclusions in CA1 region. g An irregular shaped TDP-43 accumulation in the subiculum. h A coiled body-like TDP-43-positive inclusion in the subiculum. a, b, d Gallyas-Braak hematoxylin-eosin stain. c AT-8 immunohistochemistry. e–h pAb pS409/410 immunohistochemistry. Scale bars a 400 μm, b, c 25 μm, d 50 μm, e–h 20 μm
Concomitant argyrophilic grains were observed in four PSP (21%) and three CBD cases (25%) (Fig. 4d). Among these cases, one PSP and one CBD case had TDP-43 pathology (Table 2, Fig. 4e–h). There was no significant difference in the frequency of argyrophilic grains between TDP-43-positive and TDP-43-negative PSP cases, or between CBD cases with and without TDP-43 pathology, respectively. However, in the TDP-43-positive PSP and CBD cases, argyrophilic grains were found in those cases with the most severe TDP-43 pathology (Table 2).
Double immunofluorescence labeling in PSP cases
In the PSP cases examined, TDP-43 and tau pathologies were independently present in the perikarya of granular cells in the hippocampal dentate gyrus with no coexistence of these proteins (Fig. 5a–f). In contrast, in the amygdala, TDP-43 accumulation was often intermingled with tau accumulation in NCIs and dystrophic neurites, and colocalization was frequent (Fig. 5g–o). In the entorhinal cortex and parahippocampal gyrus in one PSP case with argyrophilic grains, many tau-positive grain-like structures were demonstrated, and TDP-43 was colocalized with tau in some of these structures (data not shown).
Fig. 5.

Confocal double-imunofluorescence of TDP-43 (a, d, g, j, m) and tau (b, e, h, k, n) in PSP cases. Merged images are shown in c, f, i, l, and o. Blue fluorescence in merged images are nuclei. a–f In the hippocampal dentate gyrus, TDP-43 accumulation (arrows) is not colocalized with tau labeling. g–i In the amygdala, TDP-43 accumulation is often intermingled and colocalized with neuronal tau accumulation. j–o TDP-43-positive neurites (j, m) and many tau-positive neurites and granules (k, n) are seen in the amygdala. Coexistence of TDP-43 and tau is noted in some neurites (l, o). AT8 and pAb pS409/410 double immunofluorescence. Scale bars a–c 25 μm, d–f 25 μm, g–i 2.5 μm, j–l 7.5 μm, m–o 7.5 μm
Biochemical analyses of TDP-43 in PSP cases
Immunoblot analysis of the sarkosyl-insoluble, urea-soluble fraction with mAb pS409/410 demonstrated distinct bands at (approximately) 45 and 25 kDa, as well as high molecular weight smears in the amygdala of a PSP case having TDP-43 pathology (Fig. 6, lane 6) and in the frontal cortex of a FTLD-TDP case (lane 5). Weak 25 and 45 kDa bands were also observed in the hippocampus in a PSP case, which had very mild TDP-43 pathology at this site (lane 7). Pathological TDP-43 bands and smear were not demonstrated in any of the other cases lacking TDP-43 pathology, including those with PSP (lanes 1 and 2) or Lewy body disease (lane 4), or in normal control cases (lane 3).
Fig. 6.

Immunoblot analysis of the sarkosyl-insoluble fraction in representative PSP cases with phosphorylation-dependent monoclonal anti-TDP-43 antibody (mAb pS409/410). The 45 kDa full length TDP-43, 25 kDa fragments, and high molecular weight smear are strongly labeled in the amygdala of a PSP case with TDP-43 pathology (lane 6) and in the frontal cortex of a FTLD-TDP case (lane 5). Weakly stained 45 and 25 kDa bands are noted in the hippocampus of a PSP case (lane 7), in which TDP-43 pathology was mild. Similar 45 and 25 kDa bands and smears were not immunolabeled in any of the other cases without detectable TDP-43 pathology by immunohistochemistry (lanes 1–4). Normal 43 kDa TDP-43 is not stained by this phophorylation-dependent antibody in any case. PSP progressive supranuclear palsy, LBD Lewy body disease, NC normal control, AM amygdala, HP hippocampus, F frontal cortex, T temporal cortex, IHC pAb pS409/410 immunohistochemistry
Discussion
This is the first study demonstrating abnormal accumulations of phosphorylated TDP-43 in the limbic system in a significant proportion (26%) of patients with PSP. Immunoblot analysis also demonstrated biochemical alterations in TDP-43 in tissue samples from a PSP case with TDP-43 pathology, similar to those in FTLD-TDP and ALS. Regional tau burden in PSP cases with TDP-43 pathology was higher than that in PSP cases without it, and TDP-43 burden was significantly correlated with that of tau in the occipitotemporal cortex. The frequency of HS in PSP cases with TDP-43 pathology was significantly higher than that in PSP cases without it. Collectively, these findings suggest that (1) PSP is one of the tauopathies in which pathological TDP-43 accumulation can occur in the limbic system, and (2) TDP-43 pathology may be associated with the occurrence of HS in PSP cases.
Previous studies have demonstrated variable frequencies of concurrent TDP-43 pathology in many tauopathies: 23–56% in AD cases [2, 4, 40], 31–60% in DLB + AD cases [4, 30], 15% in CBD cases [40], and 60% in AGD cases [14]. Why no cases of PSP with TDP-43 pathology have previously been described is not clear. Our present findings show that, at least some, PSP cases may share a common pathophysiological background involving TDP-43 accumulation with other tauopathies with TDP-43 pathology. Several studies demonstrated that concurrent AD-type pathology was associated with the development of TDP-43 pathology in some neurodegenerative diseases [2, 4, 7, 14, 30]. However, it was unlikely that the development of TDP-43 pathology in our PSP series can be explained by the influence of Aβ deposits or neuritic plaques. For example, of all ten PSP cases having Aβ deposits, nine cases had only diffuse plaques, and the degree of Aβ deposition was not significantly different between TDP-43-positive and TDP-43-negative PSP cases and was not correlated with that of TDP-43 pathology in any regions. Although only one PSP case had a few neuritic plaques in the occipitotemporal gyrus, no TDP-43-positive inclusion was noted in the region.
Our findings are inconsistent with previous studies that failed to demonstrate immunohistochemical or biochemical abnormalities of TDP-43 in PSP cases [2, 3, 18, 40]. Considering that the sample size investigated in one of these previous studies [40] was far larger than that in our own study, the most plausible cause of the discrepancy may be the difference of the sensitivities of anti-TDP-43 antibodies employed: phosphorylation-dependent anti-TDP-43 antibodies do not stain normal nuclei, making true TDP-43-positive inclusions more readily identifiable [17, 37]. The distribution of TDP-43 pathology observed in our PSP cases was very similar to that reported previously in AD [2, 4, 18, 19, 40], DLB + AD [4, 30], and CBD [40], but tended to be more restricted than that in ALS/PDC of Guam [15, 16, 27]. Most frequently affected sites in these tauopathies are the amygdala and hippocampal dentate gyrus. Given these findings, it is plausible that the frequent TDP-43 accumulation in these sites in tauopathies is associated with some region-specific, rather than disease-specific, mechanism. On the other hand, it remains unclear whether TDP-43 is abnormally accumulated through an identical pathophysiological mechanism in various anatomical regions. For example, it was reported that abnormal TDP-43 accumulation was significantly correlated with the severity of tau pathology in AD cases [4] and Lewy body disease including many DLB + AD cases [30]. This same statistical relationship was observed in our PSP cases. Furthermore, in our present studies, TDP-43 was often colocalized with tau in NCIs and dystrophic neurites in the amygdala, although there were also TDP-43-positive but tau-negative lesions in this site. A coexistence of TDP-43 and tau in the same neuron in the amygdala and temporal cortex was also reported in AD and DLB cases in previous studies [4, 18]. However, in contrast to the amygdala, a coexistence of TDP-43 and tau in the same neuron in the hippocampal dentate gyrus was not seen in our PSP cases. This trend regarding non-colocalization of these two proteins was also noticed in the dentate granular cells in AD [40] and AGD brains [14]. This suggests that the mechanism underlying the accumulation of TDP-43 is different at least between the amygdala and hippocampal dentate gyrus, or that there is some unknown factor that can influence the occurrence of both TDP-43 and tau pathologies. In addition, considering the potential relationship between tau and TDP-43 in PSP presented in this paper, whether TDP-43 pathology is also noted in several other regions that are often involved by tau-associated lesions (e.g., the frontal cortex and basal ganglia) needs to be investigated in the future studies.
There is little known about the relationship between PSP and HS. In our series, 3 of 19 PSP cases (16%) had evident neuronal loss in the CA1 and/or subiculum consistent with the definition of HS. Furthermore, all of the cases with HS had TDP-43 pathology, and one of the three cases also had argyrophilic grains. It has been reported that HS cases have variable underling pathologies, including the ‘pure form’ of HS [1, 21, 34], FTLD-TDP [23], FTLD with motor neuron disease [29], AD [2], CBD [36], DLB [13], and AGD [5, 13]. Present findings support the possibility that the development of HS, at least in some PSP cases, may occur in association with concurrent TDP-43 pathology. On the other hand, whether the development of HS in PSP cases is correlated with the severity of tau or TDP-43 pathology remains unclear. Considering the relatively small size of the samples examined in the present study, the relationship between HS and TDP-43 accumulation in PSP, as well as the frequencies of these pathological features, needs to be confirmed in a larger case series.
Although influence of concurrent TDP-43 pathology on clinical features in tauopathies is not fully understood, some previous studies in AD, have demonstrated a comorbidity such that a concomitant TDP-43 pathology was associated with a later age at onset and death [4, 24], and significantly poorer cognitive function [24]. On the other hand, a study investigating a relatively small series of AGD did not demonstrate any significant difference in the age at death or disease duration between cases with and without TDP-43 pathology [14]. It is known that patients with PSP frequently exhibit psychiatric and behavioral disturbances, and that cognitive decline in PSP is associated with the atrophy in the orbitofrontal cortex [11] and more severe tau burden in the neocortex and hippocampus [6, 8, 22]. More recently, it was also reported that clinical presentation, including the occurrence of dementia, is influenced by the distribution and severity of tau pathology [42, 43]. In our PSP series, although not statistically significantly, the frequency of dementia in PSP cases with both TDP-43 and HS (100%), and that in all PSP cases with TDP-43 pathology (80%), were higher than that in PSP cases lacking both (50%). The potential co-morbid effect of concurrent TDP-43 pathology and/or HS on cognitive impairment in patients with PSP needs to be explored by further clinicopathological studies.
Acknowledgments
This study was supported in part by a research grant from the Uehara Memorial Foundation and Grant No. AG13854 from the National Institutes of Health. We thank Ms. M. Onbe (Department of Neuropsychiatry. Okayama University Graduate School of Medicine, Dentistry and Pharmaceutical Sciences) for their excellent technical assistance, and the Parkinson’s Disease Society Brain Bank for making available tissue samples for this study. A part of this study was presented at the 111th annual meeting of the British Neuropathological Society in January 2010.
Contributor Information
Osamu Yokota, Neurodegeneration and Mental Health Research Group, Faculty of Medical and Human Sciences, School of Community Based Medicine, Greater Manchester Neurosciences Centre, Hope Hospital, University of Manchester, Salford M6 8HD, UK, Department of Neuropsychiatry, Okayama University Graduate School of Medicine, Dentistry and Pharmaceutical Sciences, 2-5-1 Shikata-cho, Okayama 700-8558, Japan.
Yvonne Davidson, Neurodegeneration and Mental Health Research Group, Faculty of Medical and Human Sciences, School of Community Based Medicine, Greater Manchester Neurosciences Centre, Hope Hospital, University of Manchester, Salford M6 8HD, UK.
Eileen H. Bigio, Department of Pathology, Northwestern University Feinberg School of Medicine, Chicago, IL 60619, USA
Hideki Ishizu, Department of Neuropsychiatry, Okayama University Graduate School of Medicine, Dentistry and Pharmaceutical Sciences, 2-5-1 Shikata-cho, Okayama 700-8558, Japan, Zikei Institute of Psychiatry, 100-2, Urayasu-honcho, Okayama 702-8508, Japan.
Seishi Terada, Department of Neuropsychiatry, Okayama University Graduate School of Medicine, Dentistry and Pharmaceutical Sciences, 2-5-1 Shikata-cho, Okayama 700-8558, Japan.
Tetsuaki Arai, Department of Psychogeriatrics, Tokyo Institute of Psychiatry, 2-1-8 Kamikitazawa, Setagaya-ku, Tokyo 156-8585, Japan.
Masato Hasegawa, Department of Molecular Neurobiology, Tokyo Institute of Psychiatry, 2-1-8 Kamikitazawa, Setagaya-ku, Tokyo 156-8585, Japan.
Haruhiko Akiyama, Department of Psychogeriatrics, Tokyo Institute of Psychiatry, 2-1-8 Kamikitazawa, Setagaya-ku, Tokyo 156-8585, Japan.
Stephen Sikkink, Neurodegeneration and Mental Health Research Group, Faculty of Medical and Human Sciences, School of Community Based Medicine, A V Hill Building, University of Manchester, Oxford Rd, Manchester M13 9PL, UK.
Stuart Pickering-Brown, Neurodegeneration and Mental Health Research Group, Faculty of Medical and Human Sciences, School of Community Based Medicine, A V Hill Building, University of Manchester, Oxford Rd, Manchester M13 9PL, UK.
David M. A. Mann, Email: david.mann@manchester.ac.uk, Neurodegeneration and Mental Health Research Group, Faculty of Medical and Human Sciences, School of Community Based Medicine, Greater Manchester Neurosciences Centre, Hope Hospital, University of Manchester, Salford M6 8HD, UK
References
- 1.Ala TA, Beh GO, Frey WH., 2nd Pure hippocampal sclerosis: a rare cause of dementia mimicking Alzheimer’s disease. Neurology. 2000;54:843–848. doi: 10.1212/wnl.54.4.843. [DOI] [PubMed] [Google Scholar]
- 2.Amador-Ortiz C, Lin WL, Ahmed Z, et al. TDP-43 immunoreactivity in hippocampal sclerosis and Alzheimer’s disease. Ann Neurol. 2007;61:435–445. doi: 10.1002/ana.21154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Arai T, Hasegawa M, Akiyama H, et al. TDP-43 is a component of ubiquitin-positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Biochem Biophys Res Commun. 2006;351:602–611. doi: 10.1016/j.bbrc.2006.10.093. [DOI] [PubMed] [Google Scholar]
- 4.Arai T, Mackenzie IR, Hasegawa M, et al. Phosphorylated TDP-43 in Alzheimer’s disease and dementia with Lewy bodies. Acta Neuropathol. 2009;117:125–136. doi: 10.1007/s00401-008-0480-1. [DOI] [PubMed] [Google Scholar]
- 5.Beach TG, Sue L, Scott S, et al. Hippocampal sclerosis dementia with tauopathy. Brain Pathol. 2003;13:263–278. doi: 10.1111/j.1750-3639.2003.tb00027.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bigio EH, Brown DF, White CL., 3rd Progressive supra-nuclear palsy with dementia: cortical pathology. J Neuropathol Exp Neurol. 1999;58:359–364. doi: 10.1097/00005072-199904000-00006. [DOI] [PubMed] [Google Scholar]
- 7.Bigio EH, Mishra M, Hatanpaa KJ, et al. TDP-43 pathology in primary progressive aphasia and frontotemporal dementia with pathologic Alzheimer disease. Acta Neuropathol. 2010 doi: 10.1007/s00401-010-0681-2. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Braak H, Braak E. Neurofibrillary changes confined to the entorhinal region and an abundance of cortical amyloid in cases of presenile and senile dementia. Acta Neuropathol. 1990;80:479–486. doi: 10.1007/BF00294607. [DOI] [PubMed] [Google Scholar]
- 9.Braak H, Alafuzoff I, Arzberger T, Kretzschmar H, Del Tredici K. Staging of Alzheimer disease-associated neurofibrillary pathology using paraffin sections and immunocytochemistry. Acta Neuropathol. 2006;112:389–404. doi: 10.1007/s00401-006-0127-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cairns NJ, Neumann M, Bigio EH, et al. TDP-43 in familial and sporadic frontotemporal lobar degeneration with ubiquitin inclusions. Am J Pathol. 2007;171:227–240. doi: 10.2353/ajpath.2007.070182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cordato NJ, Duggins AJ, Halliday GM, Morris JG, Pantelis C. Clinical deficits correlate with regional cerebral atrophy in progressive supranuclear palsy. Brain. 2005;128:1259–1266. doi: 10.1093/brain/awh508. [DOI] [PubMed] [Google Scholar]
- 12.Davidson Y, Kelley T, Mackenzie IR, et al. Ubiquitinated pathological lesions in frontotemporal lobar degeneration contain the TAR DNA-binding protein, TDP-43. Acta Neuropathol. 2007;113:521–533. doi: 10.1007/s00401-006-0189-y. [DOI] [PubMed] [Google Scholar]
- 13.Dickson DW, Davies P, Bevona C, et al. Hippocampal sclerosis: a common pathological feature of dementia in very old (> or = 80 years of age) humans. Acta Neuropathol. 1994;88:212–221. doi: 10.1007/BF00293396. [DOI] [PubMed] [Google Scholar]
- 14.Fujishiro H, Uchikado H, Arai T, et al. Accumulation of phosphorylated TDP-43 in brains of patients with argyrophilic grain disease. Acta Neuropathol. 2009;117:151–158. doi: 10.1007/s00401-008-0463-2. [DOI] [PubMed] [Google Scholar]
- 15.Geser F, Winton MJ, Kwong LK, et al. Pathological TDP-43 in parkinsonism-dementia complex and amyotrophic lateral sclerosis of Guam. Acta Neuropathol. 2008;115:133–145. doi: 10.1007/s00401-007-0257-y. [DOI] [PubMed] [Google Scholar]
- 16.Hasegawa M, Arai T, Akiyama H, et al. TDP-43 is deposited in the Guam parkinsonism-dementia complex brains. Brain. 2007;130:1386–1394. doi: 10.1093/brain/awm065. [DOI] [PubMed] [Google Scholar]
- 17.Hasegawa M, Arai T, Nonaka T, et al. Phosphorylated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Ann Neurol. 2008;64:60–70. doi: 10.1002/ana.21425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Higashi S, Iseki E, Yamamoto R, et al. Concurrence of TDP-43, tau and alpha-synuclein pathology in brains of Alzheimer’s disease and dementia with Lewy bodies. Brain Res. 2007;1184:284–294. doi: 10.1016/j.brainres.2007.09.048. [DOI] [PubMed] [Google Scholar]
- 19.Hu WT, Josephs KA, Knopman DS, et al. Temporal lobar predominance of TDP-43 neuronal cytoplasmic inclusions in Alzheimer disease. Acta Neuropathol. 2008;116:215–220. doi: 10.1007/s00401-008-0400-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Inukai Y, Nonaka T, Arai T, et al. Abnormal phosphorylation of Ser409/410 of TDP-43 in FTLD-U and ALS. FEBS Lett. 2008;582:2899–2904. doi: 10.1016/j.febslet.2008.07.027. [DOI] [PubMed] [Google Scholar]
- 21.Jellinger KA. Pure hippocampal sclerosis: a rare cause of dementia mimicking Alzheimer’s disease. Neurology. 2000;55:739–740. doi: 10.1212/wnl.55.5.735-d. [DOI] [PubMed] [Google Scholar]
- 22.Jellinger KA. Different tau pathology pattern in two clinical phenotypes of progressive supranuclear palsy. Neurodegener Dis. 2008;5:339–346. doi: 10.1159/000121388. [DOI] [PubMed] [Google Scholar]
- 23.Josephs KA, Dickson DW. Hippocampal sclerosis in tau-negative frontotemporal lobar degeneration. Neurobiol Aging. 2007;28:1718–1722. doi: 10.1016/j.neurobiolaging.2006.07.010. [DOI] [PubMed] [Google Scholar]
- 24.Josephs KA, Whitwell JL, Knopman DS, et al. Abnormal TDP-43 immunoreactivity in AD modifies clinicopathologic and radiologic phenotype. Neurology. 2008;70:1850–1857. doi: 10.1212/01.wnl.0000304041.09418.b1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kabashi E, Valdmanis PN, Dion P, et al. TARDBP mutations in individuals with sporadic and familial amyotrophic lateral sclerosis. Nat Genet. 2008;40:572–574. doi: 10.1038/ng.132. [DOI] [PubMed] [Google Scholar]
- 26.McKeith IG, Dickson DW, Lowe J, et al. Diagnosis and management of dementia with Lewy bodies: third report of the DLB Consortium. Neurology. 2005;65:1863–1872. doi: 10.1212/01.wnl.0000187889.17253.b1. [DOI] [PubMed] [Google Scholar]
- 27.Miklossy J, Steele JC, Yu S, et al. Enduring involvement of tau, β-amyloid, α-synuclein, ubiquitin and TDP-43 pathology in the amyotrophic lateral sclerosis/parkinsonism–dementia complex of Guam (ALS/PDC) Acta Neuropathol. 2008;116:625–637. doi: 10.1007/s00401-008-0439-2. [DOI] [PubMed] [Google Scholar]
- 28.Mirra SS, Heyman A, McKeel D, et al. The Consortium to Establish a Registry for Alzheimer’s Disease (CERAD). Part II. Standardization of the neuropathologic assessment of Alzheimer’s disease. Neurology. 1991;41:479–486. doi: 10.1212/wnl.41.4.479. [DOI] [PubMed] [Google Scholar]
- 29.Nakano I. Temporal lobe lesions in amyotrophic lateral sclerosis with or without dementia—a neuropathological study. Neuropathology. 1993;13:215–227. [Google Scholar]
- 30.Nakashima-Yasuda H, Uryu K, et al. Comorbidity of TDP-43 proteinopathy in Lewy body related diseases. Acta Neuropathol. 2007;114:221–229. doi: 10.1007/s00401-007-0261-2. [DOI] [PubMed] [Google Scholar]
- 31.Neumann M, Sampathu DM, Kwong LK, et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science. 2006;314:130–133. doi: 10.1126/science.1134108. [DOI] [PubMed] [Google Scholar]
- 32.Neumann M, Mackenzie IR, Cairns NJ, et al. TDP-43 in the ubiquitin pathology of frontotemporal dementia with VCP gene mutations. J Neuropathol Exp Neurol. 2007;66:152–157. doi: 10.1097/nen.0b013e31803020b9. [DOI] [PubMed] [Google Scholar]
- 33.Obi K, Akiyama H, Kondo H, et al. Relationship of phosphorylated alpha-synuclein and tau accumulation to Abeta deposition in the cerebral cortex of dementia with Lewy bodies. Exp Neurol. 2008;210:409–420. doi: 10.1016/j.expneurol.2007.11.019. [DOI] [PubMed] [Google Scholar]
- 34.Probst A, Taylor KI, Tolnay M. Hippocampal sclerosis dementia: a reappraisal. Acta Neuropathol. 2007;114:335–345. doi: 10.1007/s00401-007-0262-1. [DOI] [PubMed] [Google Scholar]
- 35.Saito Y, Ruberu NN, Sawabe M, et al. Staging of argyrophilic grains: an age-associated tauopathy. J Neuropathol Exp Neurol. 2004;63:911–918. doi: 10.1093/jnen/63.9.911. [DOI] [PubMed] [Google Scholar]
- 36.Schneider JA, Watts RL, Gearing M, et al. Corticobasal degeneration: neuropathologic and clinical heterogeneity. Neurology. 1997;48:959–969. doi: 10.1212/wnl.48.4.959. [DOI] [PubMed] [Google Scholar]
- 37.Schwab C, Arai T, Hasegawa M, Yu S, McGeer PL. Colocalization of transactivation-responsive DNA-binding protein 43 and huntingtin in inclusions of Huntington disease. J Neuropathol Exp Neurol. 2008;67:1159–1165. doi: 10.1097/NEN.0b013e31818e8951. [DOI] [PubMed] [Google Scholar]
- 38.Sreedharan J, Blair IP, Tripathi VB, et al. TDP-43 mutations in familial and sporadic amyotrophic lateral sclerosis. Science. 2008;319:1668–1672. doi: 10.1126/science.1154584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.The National Institute on Aging, and Reagan Institute Working Group. Consensus recommendations for the postmortem diagnosis of Alzheimer disease. The National Institute on Aging, and Reagan Institute Working Group on diagnostic criteria for the neuropathological assessment of Alzheimer disease. Neurobiol Aging. 1997;18:S1–S2. [PubMed] [Google Scholar]
- 40.Uryu K, Nakashima-Yasuda H, Forman MS, et al. Concomitant TAR-DNA-binding protein 43 pathology is present in Alzheimer disease and corticobasal degeneration but not in other tauopathies. J Neuropathol Exp Neurol. 2008;67:555–564. doi: 10.1097/NEN.0b013e31817713b5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Van Deerlin VM, Leverenz JB, Bekris LM, et al. TARDBP mutations in amyotrophic lateral sclerosis with TDP-43 neuropathology: a genetic and histopathological analysis. Lancet Neurol. 2008;7:409–416. doi: 10.1016/S1474-4422(08)70071-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Williams DR, Lees AJ. Progressive supranuclear palsy: clinicopathological concepts and diagnostic challenges. Lancet Neurol. 2009;8:270–279. doi: 10.1016/S1474-4422(09)70042-0. [DOI] [PubMed] [Google Scholar]
- 43.Williams DR, Holton JL, Strand C, et al. Pathological tau burden and distribution distinguishes progressive supranuclear palsy-parkinsonism from Richardson’s syndrome. Brain. 2007;130:1566–1576. doi: 10.1093/brain/awm104. [DOI] [PubMed] [Google Scholar]
- 44.Yokoseki A, Shiga A, Tan CF, et al. TDP-43 mutation in familial amyotrophic lateral sclerosis. Ann Neurol. 2008;63:538–542. doi: 10.1002/ana.21392. [DOI] [PubMed] [Google Scholar]