

Summary
The contractile activity of striated muscle depends on myofibrils that are highly ordered macromolecular complexes. The protein components of myofibrils are well characterized, but it remains largely unclear how signaling at the molecular level within the sarcomere and the control of assembly are coordinated. We show that the Rho GTPase TC10 appears during differentiation of human primary skeletal myoblasts and it is active in differentiated myotubes. We identify obscurin, a sarcomere-associated protein, as a specific activator of TC10. Indeed, TC10 binds directly to obscurin via its predicted RhoGEF motif. Importantly, we demonstrate that obscurin is a specific activator of TC10 but not the Rho GTPases Rac and Cdc42. Finally, we show that inhibition of TC10 activity by expression of a dominant-negative mutant or its knockdown by expression of specific shRNA block myofibril assembly. Our findings reveal a novel signaling pathway in human skeletal muscle that involves obscurin and the Rho GTPase TC10 and implicate this pathway in new sarcomere formation.
Keywords: Cell Differentiation; Cells, Cultured; Enzyme Activation; Guanine Nucleotide Exchange Factors; chemistry; metabolism; Humans; Muscle Fibers, Skeletal; cytology; enzymology; Muscle Proteins; chemistry; metabolism; Myofibrils; enzymology; Organogenesis; Phosphorylation; Protein Binding; Protein Structure, Tertiary; RNA, Small Interfering; metabolism; Sarcomeres; enzymology; metabolism; p21-Activated Kinases; metabolism; rho GTP-Binding Proteins; antagonists & inhibitors; metabolism
Keywords: Myofibrillogenesis, Rho GTPase, Obscurin
Introduction
The myoplasm of adult striated muscle fibers contains highly ordered macromolecular complexes, named myofibrils, that are necessary for contraction. The myofibril assembly takes place during the embryonic differentiation of heart and skeletal muscle, but also on a continuous basis during the physiological turnover of muscle. The thousands of protein subunits that compose sarcomeres, the smallest contractile units of myofibrils, are well characterized. On the other hand, it remains largely unclear how signaling at the molecular level within the sarcomere and the control of assembly are coordinated. Therefore, identifying and characterizing key elements of sarcomeric signal transduction and their roles in the control of myofibrillogenesis will be essential to understand the basic mechanisms of the biology of muscle under normal and pathological conditions.
The process of myofibril assembly requires both spatial and temporal coordination of protein interactions with high precision (Gautel et al., 1999). In vertebrate muscle, this is mediated by three giant modular proteins: titin (Maruyama, 1976; Wang et al., 1979), nebulin (Wang et al., 1996) and obscurin (Young et al., 2001), which act as molecular scaffolds or blueprints. Titin and nebulin regulate myofibril assembly, providing specific attachment sites for other proteins and thus specifying their sarcomeric positions (Maruyama, 1976; Trinick, 1996; Trinick and Tskhovrebova, 1999; Wang et al., 1979). In cardiac myocytes and muscle cell lines, the suppression of titin expression was associated with a significant structural disturbance of the contractile apparatus and a failure to incorporate myosin filaments in the nascent sarcomeres (Person et al., 2000; van der Ven et al., 2000). Obscurin, the third and last identified sarcomeric giant protein (Young et al., 2001), interacts with titin and myomesin (Fukuzawa et al., 2008) and localizes mainly to the M-band in mature myofibrils (Borisov et al., 2004; Kontrogianni-Konstantopoulos et al., 2004; Young et al., 2001). Obscurin is also implicated in myofibril assembly in various organisms, but its mechanism of action remains unidentified. Defective myofibrillogenesis has been reported in cardiac myocytes following the disruption of obscurin expression by small interfering RNA-mediated gene silencing (Borisov et al., 2006; Kontrogianni-Konstantopoulos et al., 2006). In C. elegans, the ortholog of obscurin, Unc-89, is localized to the M-band in obliquely striated muscle and a mutation in this gene leads to loss of M-line assembly and regular A-bands (Benian et al., 1996). In Zebrafish, inhibition of obscurin expression with morpholino antisense oligos results in diminished numbers of skeletal myofibrils and impaired lateral alignment of adjacent myofibrils (Raeker et al., 2006).
These giant proteins contain potential signaling domains, suggesting that they receive and propagate signals from various pathways that remain to be characterized. Titin contains an active protein kinase domain implicated in myofibril assembly (Mayans et al., 1998), whereas nebulin and obscurin possess a src homology 3 (SH3) domain (Labeit and Kolmerer, 1995; Young et al., 2001). In addition, obscurin contains a predicted GDP/GTP exchange factor (Rho GEF) domain (Young et al., 2001), also known as dbl-homology (DH) domain, immediately followed by a pleckstrin homology (PH) domain (Hart et al., 1991). This tandem organization is found in RhoGEF proteins, the activators of the Rho GTPases (Rossman et al., 2005). Rho GTPases are key regulators of gene expression and actin cytosketal reorganization that control cell shape dynamics during diverse physiological processes, such as cell polarity, motility, adhesion, differentiation and apoptosis (Etienne-Manneville and Hall, 2002). Rho GTPases adopt either an active GTP-bound or an inactive GDP-bound conformation. Their activity is controlled positively by guanine nucleotide exchange factors (RhoGEF) and negatively by GTPase activating proteins (RhoGAPs). Moreover, guanine nucleotide dissociation inhibitors (GDIs) recycle Rho proteins before their reactivation (Van Aelst and D’Souza-Schorey, 1997). Once loaded with GTP, the GTPase binds to and activates a set of downstream effector proteins. Previous studies have implicated Rho GTPase-dependent signaling pathways in muscle differentiation. Rac1 signaling is required for myoblast fusion (Luo et al., 1994; Nolan et al., 1998) while RhoA positively regulates sarcomeric protein expression by modulating the transcriptional activity of tissue-specific transcription factors (Carnac et al., 1998; Wei et al., 1998). Recently, we have established a role of TC10 in myofibrillogenesis in an ascidian model (Coisy-Quivy et al., 2006). Inhibition of TC10 activity by expression of a dominant negative mutant in tail muscle cells of Ciona intestinalis larva impairs myofibril assembly. Consistent with our results in ascidia, TC10 mRNA is easily detected in striated muscle tissues from mouse and human (Abe et al., 2003; Neudauer et al., 1998). In the mouse C2C12 cell line, TC10 mRNA is detected at a moderate level in undifferentiated myoblasts. It is then induced in myotubes and reaches high levels in terminally differentiated myotubes (Abe et al., 2003). Although Jebailey et al. observed TC10 transcripts in differentiated rat L6 skeletal muscle cells and 3T3-L1 adipocytes, the protein was only detected in adipocytes (JeBailey et al., 2004). In the latter, TC10 regulated insulin-induced translocation of the GLUT4 glucose transporter protein from intracellular storage sites to the plasma membrane. Since this function appears to be performed by Rac in rat L6 skeletal muscle cells, the functional role of TC10 remains unknown in vertebrate striated muscles (JeBailey et al., 2004; Khayat et al., 2000).
In this study, we show that TC10 is present and active in human differentiated skeletal muscle cells. Moreover, we find that obscurin binds directly to TC10 and promotes its activation in human primary myotubes. Our data provide the first evidence that obscurin is a functional RhoGEF. Finally, we demonstrate that TC10 expression and activity are essential for human myofibrillogenesis, indicating that this function of TC10 is conserved from ascidia to humans.
Results
TC10 is expressed and active in human differentiated myotubes
We recently demonstrated the role of TC10 in myofibrillogenesis in an ascidian model (Coisy-Quivy et al., 2006). To explore its role in human myofibrillogenesis, we first examined TC10 expression during differentiation of cultured human primary myoblasts. Immunoblot analysis showed that TC10 is not expressed, or at very low levels, in proliferating undifferentiated myoblasts (Fig. 1A). TC10 appeared after myoblast fusion, which is marked by a strong expression of the myogenic marker myogenin and by the appearance of differentiation-specific proteins such as obscurin, myosin heavy chain (MHC) and sarcomeric α-actinin (Fig. 1A). During differentiation, TC10 expression was maintained (Fig. 1A). This suggests that TC10 is probably not necessary for myoblast fusion, but rather for later processes of differentiation. We next examined whether TC10 was active in differentiated myotubes, incubating lysates from proliferating myoblasts and differentiated myotubes with the p21-binding domain (PBD) of PAK1 fused to glutathione S-transferase (GST) (Fig. 1B). The PBD of PAK1 binds specifically to GTP-bound active forms of TC10, Cdc42 and Rac (Benard et al., 1999; Neudauer et al., 1998). Endogenous TC10 was detected neither in lysates nor in GST-PBD precipitates from undifferentiated proliferating myoblasts (Fig. 1B). In contrast, endogenous TC10 associated with GST-PBD and was therefore active in extracts from differentiated myotubes (Fig. 1B). These data present the first evidence that TC10 is present and active in differentiated myotubes.
Figure 1. Expression and activity of TC10 during differentiation of human primary myoblasts.

(A) Lysates from human primary myoblasts, collected at different time of differentiation, were separated by SDS PAGE and western blots incubated with the indicated antibodies. The results shown are typical of four independent experiments.
(B) The level of active TC10 in lysates from proliferating (“proliferation”) and differentiated (“differentiation”) myoblasts was measured by binding to GST-PBD (IP). Proteins were separated by SDS PAGE and visualized by immunoblotting (IB) with the indicated antibodies. The results shown are typical of three independent experiments.
TC10 binds to the DH domain of obscurin in cells and in vitro
The activation of Rho GTPase results from an exchange of a GDP by a GTP that is mediated by a dbl homology (DH) domain, immediately followed by a pleckstrin homology (PH) domain in Rho guanine nucleotide exchange factor (Rho GEF). A similar DH-PH module in obscurin (Young et al., 2001) interested us because this sarcomeric protein is essential for de novo myofibril assembly (Borisov et al., 2006). To explore whether obscurin could activate TC10 during myofibrillogenesis, we first tested the interaction between these two proteins in the human cell line 293T. Two myc-tagged expression vectors for obscurin were constructed. The first encodes the C-terminal sequences in obscurin (ObsCT) that contain the DH-PH domains and two immunoglobulin-like repeats (Fig. 2A). The second uniquely contains the DH domain (ObsDH) (Fig. 2A), which is usually sufficient for interaction between Rho-GEF and Rho GTPase family proteins. Myc-ObsCT or myc-ObsDH were co-expressed in 293T cells with either HA-tagged TC10 or HA-tagged control protein (HA-Elmo). Cell lysates were incubated with anti-HA antibody and the immunoprecipitated proteins were detected by western blot analysis. Myc-ObsCT and myc-ObsDH coimmunoprecipitated with HA-TC10 but not with the control HA-Elmo (Fig. 2B). To confirm the interaction between obscurin and TC10, lysates from 293T cells overexpressing HA-TC10 and myc-ObsCT were incubated with either control rabbit immunoglobulin (IgG) or an anti-obscurin antibody that recognizes the C-terminus region of obscurin. HA-TC10 specifically coimmunoprecipitated with myc-ObsCT (Fig. 2C, lane 6). As obscurin is not expressed in 293T cells, no HA-TC10 was immunoprecipitated by the obscurin-specific antibody without myc-ObsCT expression (Fig. 2C, lane 4). Next, we analyzed the interaction of obscurin with constitutively activated mutant (HA-TC10CA) and the dominant negative form of TC10 (HA-TC10DN). The constitutively-activated mutants of Rho GTPase, which cannot hydrolyse GTP, bind to and activate their downstream effector. In contrast, dominant-negative mutants of Rho GTPases are unable to adopt the GTP-bound conformation and titrate out GEFs, leading to the inhibition of a Rho GTPase-dependent pathway. Lysates from 293T cells overexpressing either HA-TC10CA or HA-TC10DN and either myc-ObsCT or its DH-PH deleted version (Myc-ObsCT(ΔDH-PH)) (Fig. 2A) were incubated with the anti-obscurin antibody. As expected, HA-TC10DN coimmunoprecipitated with myc-ObsCT and very slightly with Myc-ObsCT(ΔDH-PH) (Fig. 2D). This result confirms that TC10 interacts with the DH-PH domain of obscurin, although we cannot exclude that an additional domain of obscurin contributes to this interaction. In contrast, HA-TC10CA did not coimmunoprecipitate with myc-ObsCT or Myc-ObsCT(ΔDH-PH) (Fig. 2D), indicating clearly that obscurin is not an effector of TC10.
Figure 2. Interaction between Obscurin and TC10 proteins overexpressed in 293T cells.
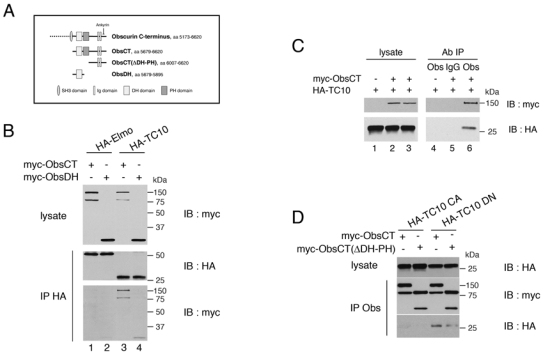
(A) Schematic representation of the obscurin C-terminus and the various expression vectors used.
(B) 293T cells were transfected with expression vectors for myc-tagged DH domain (myc-ObsDH) or carboxyterminal region (myc-ObsCT) of Obscurin, along with HA-TC10 or HA-Elmo as indicated. The latter were immunoprecipitated (IP) from total cell lysates with an anti-HA monoclonal antibody (HA), and the binding of obscurin was detected by immunoblotting (IB) with a polyclonal antibody to the Myc epitope. The results shown are typical of four independent experiments.
(C) The same experiment as in (B) except that the immunoprecipitation (IP) was performed with anti-obscurin polyclonal antiserum (Obs) or non-specific rabbit IgG (IgG). Binding of TC10 to the immunoprecipitated protein was detected by immunoblotting (IB) with an anti-HA monoclonal antibody. The lanes marked Lysate were loaded with 10% of the total cell lysates. The results shown are typical of four independent experiments.
(D) 293T cells were transfected with expression vectors for either myc-tagged carboxyterminal region (myc-ObsCT) of Obscurin or same vector lacking the DH-PH domain (myc-ObsCT(ΔDH-PH)), along with either HA-TC10CA (TC10/Q75L) (Chiang et al., 2001) or HA-TC10DN (TC10/T31N) (Chiang et al., 2001) as indicated. The immunoprecipitation was performed with anti-obscurin polyclonal antiserum (IP Obs). The binding of TC10 mutants to the immunoprecipitated protein was detected by immunoblotting (IB) with an anti-HA monoclonal antibody. The lanes marked Lysate were loaded with 10% of the total cell lysates. The results shown are typical of three independent experiments.
To determine whether the endogenous obscurin also interacts with TC10, lysates from human primary myotubes overexpressing HA-TC10 were incubated with the anti-obscurin antibody. We found that HA-TC10 was specifically coimmunoprecipitated with endogenous obscurin (Fig. 3A). To establish whether obscurin and TC10 interacted directly, we tested their association in vitro. Full-length TC10 was produced in bacteria as a MBP fusion protein and bound to amylose resin. The DH domain of obscurin was produced in bacteria as a GST fusion protein, purified on glutathione-agarose beads and eluted with glutathione. GST protein alone or GST-ObsDH were incubated with immobilized MBP-TC10. GST-ObsDH, but not GST, interacted with MBP-TC10, indicating that obscurin binds directly to TC10 via its DH domain (Fig. 3B).
Figure 3. Obscurin interacts with TC10 in myotubes.
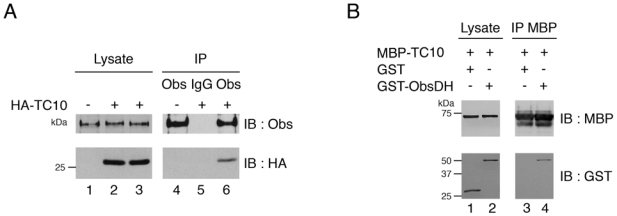
(A) Differentiated human myotubes were transduced with empty lentiviral vector or vector expressing HA-TC10 as indicated. Endogenous obscurin was immunoprecipitated (IP) from total cell lysates with anti-obscurin polyclonal antiserum (Obs) or non-specific rabbit IgG (IgG). Binding of TC10 to the obscurin immunoprecipitates was detected by immunoblotting (IB) with an anti-HA monoclonal antibody (HA). Lysate represents 5% of the total cell lysates. The results shown are typical of three independent experiments.
(B) Purified recombinant GST-obscurin DH domain (GST-ObsDH) or GST alone were incubated with immobilized MBP-TC10, and the washed complexes were separated by SDS-PAGE. Binding of the GST fusion was detected by immunoblotting (IB) with an anti-GST polyclonal antiserum (GST). 5% of the recombinant protein input was loaded in the lane marked Lysate. The results shown are typical of three independent experiments.
Obscurin specifically activates TC10 in myotubes
We next tested whether obscurin could activate TC10 in cells. The GTP-bound, active form of TC10 was assayed via its interaction with the PBD of PAK1, as described in Figure 1. We cotransfected 293T cells with expression vectors for obscurin (Myc-ObsCT), wild-type TC10 (HA-TC10wt) or its constitutively active mutant (HA-TC10CA), and the PBD of PAK1 linked to GFP (GFP-PBD). As expected, the constitutively active mutant of TC10 (HA-TC10 CA) was strongly immunoprecipitated by the PBD (Fig. 4A, lane 3). While a low level of active HA-TC10wt bound PBD in the absence of obscurin (Fig. 4A, lane 1), this was strongly increased when obscurin (Myc-ObsCT) was coexpressed (Fig. 4A, lane 2). Quantification revealed a four-fold increase in active TC10 in the presence of obscurin, and a six-fold increase with overexpressed HA-TC10CA (Fig. 4A). These results suggest that obscurin can mediate GTP exchange on TC10 in cells. To verify whether obscurin can activate endogenous TC10 in human primary myotubes and to evaluate its specificity, we performed the same assays for endogenous TC10, Cdc42 and Rac. All are expressed in myotubes and interact with PBD of PAK1 when activated. Overexpression of myc-ObsCT in differentiated myotubes induced an increase of endogenous active TC10 (Fig. 4B), but did not significantly alter the level of active Rac and Cdc42 (Fig. 4B). As expected, the TC10 activation is dependent on the DH-PH domain, since overexpression of Myc-ObsCT(ΔDH-PH) failed to activate TC10 (Fig. 4B). Thus, obscurin specifically activates TC10 in human primary myotubes. To further confirm this observation, we directly analyzed PAK activation induced by TC10 in human primary myotubes. The activation of PAK1 by Rho GTPases leads to its phosphorylation at Ser199 and/or Ser204 which can be detected with an antibody specific for this modification (Chong et al., 2001). Phosphorylated PAK was detected in myoblasts; however, its level increased after fusion and during differentiation of myotubes (Fig. 5A). We next verified that the phosphorylation level of PAK in human skeletal muscle cells depended on TC10 activity. For this, we transduced human primary myoblasts with lentivirus vectors expressing either a short hairpin RNA (shRNA) directed against human TC10 (TC10 shRNA) or lacZ control gene (C shRNA). Five days post-fusion, myotubes were lysed and proteins were evaluated by immunoblotting. TC10 expression was reduced by the shRNA (Fig. 5B). This did not affect the expression of PAK but led to a 50% reduction of its phosphorylation on Ser199 and 204. This result suggests that PAK is an effector of TC10 in human skeletal cells, and thus can serve as a measure of the level of active TC10 in cells. To test this, we overexpressed myc-ObsCT in human primary myotubes and examined the level of phosphorylated PAK. Myc-ObsCT or HA-TC10wt expression slightly increased the phosphorylation of endogenous PAK (Fig. 5C, lanes 2 and 3 respectively). Interestingly, myotubes ectopically expressing both proteins showed a higher level of phosphorylated PAK (Fig. 5C, lane 4), matching that induced by the constitutively active mutant of TC10 (Fig. 5C, lane 5). Altogether, these data provide the first demonstration that obscurin is a functional RhoGEF and it displays specific activity toward TC10 in human differentiated myotubes.
Figure 4. Obscurin activates TC10.
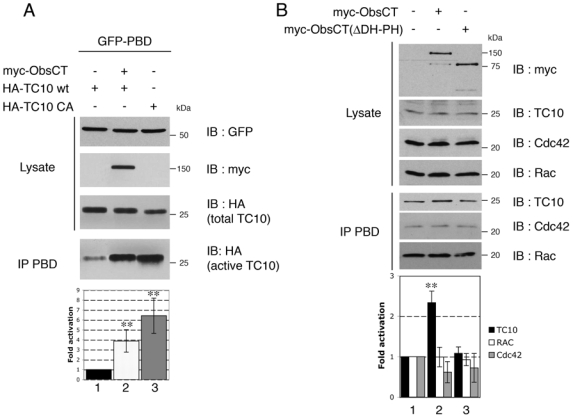
(A) 293T cells were transfected with expression vectors for myc-tagged carboxyterminal region of Obscurin (myc-ObsCT), HA-tagged wild-type (HA-TC10 wt) or constitutively active TC10 (HA-TC10 CA), and the PBD of PAK1 fused to GFP (GFP-PBD), as indicated. Expression levels of the different proteins was confirmed by immunoblotting SDS-PAGE loaded with 10% of each cell lysate (IB). Active TC10 was isolated by immunoprecipitating its complexes with GFP-PBD with an anti-GFP polyclonal antiserum (IP PBD). It was visualized by immunoblotting with the HA monoclonal antibody. The histogram represents the level of active HA-TC10 normalized for the amount of total HA-TC10 expressed in cells. Data are the mean ± S.E.M of four independent experiments (n=4) (**p < 0.001).
(B) Human primary myotubes were transfected with myc-tagged carboxyterminal region of Obscurin (myc-ObsCT) or the same vector lacking the DH-PH domain (myc-ObsCT(ΔDH-PH)) as indicated (+). The endogenous Rho GTPases TC10, Cdc42 and Rac in their active state were isolated via their binding to GST-PBD (IP PBD). Cell lysates were separated by SDS PAGE and proteins visualized by immunoblotting (IB) with the indicated antibodies. Lysate represents 5% of the total cell lysates. The histogram represents the level of active Rho GTPase normalized for the amount of total Rho GTPase expressed in cells as indicated. Data are the mean ± S.E.M of three independent experiments (n=3) (**p < 0.001).
Figure 5. Activation of TC10 by expression of carboxyterminal region of Obscurin induces PAK autophosphorylation in human primary myotubes.

(A) Lysates from human primary myoblasts, collected at different times of differentiation, were separated by SDS PAGE and western blots incubated with the indicated antibodies. The histogram represents the endogenous level of phosphorylated PAK normalized for the amount of total PAK expressed in cells. Data are the mean ± S.E.M of three independent experiments (n=3).
(B) Human myoblasts were transduced with lentiviral vectors expressing either a control (C shRNA, lane 1) or a TC10-specific shRNA (TC10 shRNA, lane 2) and maintained in culture until terminally differentiated. Cell lysates were separated by SDS PAGE and proteins visualized by immunoblotting (IB) with the indicated antibodies. The histogram shows the quantification of phosphorylated PAK normalized to the amount of PAK in cells. Data are the mean ± S.E.M of five independent experiments (n=5) (***p < 0.0001).
(C) Human primary myotubes were transfected with expression vectors for myc-tagged carboxyterminal region of Obscurin (myc-ObsCT) and/or HA-tagged wild-type (HA-TC10 wt) or constitutively active TC10 (HA-TC10 CA), as indicated. Cell lysates were separated by SDS-PAGE and proteins visualized by immunoblotting (IB) with the indicated antibodies. The histogram shows the quantification of phosphorylated PAK normalized for the amount of PAK in cells. Data are the mean ± S.E.M of four independent experiments (n=4) (*p<0.05).
TC10 activity is required for myofibrillogenesis
As obscurin is essential for de novo myofibril assembly (Borisov et al., 2006; Kontrogianni-Konstantopoulos et al., 2006) and for TC10 activation, we explored whether TC10-dependent signaling was implicated in myofibrillogenesis of human skeletal cells. Previous studies have shown that the Rho-family GTPase RhoA can regulate sarcomeric protein expression by modulating the activity of the myogenic transcription factor SRF (Carnac et al., 1998; Wei et al., 1998). We investigated whether the deregulation of TC10 activity affected sarcomeric protein expression, using two types of lentiviral vectors. One reduced TC10 levels and thereby TC10 activity via a shRNA whereas the second increased TC10 activity by expressing either HA-tagged wild-type or constitutively active TC10. Since sarcomeric proteins are induced and begin to assemble rapidly during differentiation, we infected undifferentiated human skeletal cells with lentivirus at the myoblast stage. Five days post-fusion, infected myotubes were lysed and protein expression was analyzed by immunoblot. The level of TC10 was dramatically downregulated in myotubes expressing the TC10 shRNA compared to the control lacZ shRNA (Fig. 6A and 6C). TC10 downregulation did not significantly affect the expression of obscurin, α-actinin, myogenin and α-tubulin (Fig. 6A and 6C), whereas it led to reduced levels of myosin heavy chain (34%) and troponin T (33%) (Fig. 6A and 6C). In contrast, the level of these proteins was not significantly modified in myotubes infected with lentiviral vectors expressing HA-tagged wild-type (HA-TC10wt) or constitutively active TC10 (HA-TC10CA) (Fig. 6B, lanes 2 and 3 and 6C). This result suggests that the reduction obtained in myotubes containing TC10 shRNA is primarily due to inhibition of myofibril assembly. Indeed, the stability of myosin and myomesin is reduced when they are not assembled in myofibrils (Kontrogianni-Konstantopoulos et al., 2006). These results suggest that TC10 activity is crucial for myofibril assembly.
Figure 6. TC10 effect on the sarcomeric protein expression.
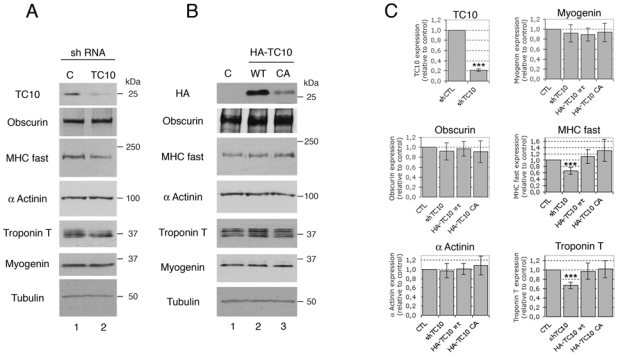
(A) Human myoblasts were transduced with lentiviral vectors expressing either a control (C shRNA, lane 1) or a TC10-specific shRNA (TC10 shRNA, lane 2) and maintained in culture until terminally differentiated. Cell lysates were separated by SDS-PAGE and proteins analyzed by immunoblotting with the indicated antibodies. The results shown are typical of six independent experiments.
(B) Human myoblasts were transduced with empty lentiviral vector (C, lane 1) or vectors expressing either HA-tagged wild-type (HA-TC10 wt, lane 2) or constitutively active TC10 (HA-TC10 CA, lane 3), as indicated. As above, cells were maintained in culture until terminally differentiated. Cell lysates were separated by SDS PAGE and proteins visualized by immunoblotting with the indicated antibodies. The results shown are typical of six independent experiments.
(C) The histograms show the quantification of the expression level of the proteins obtained in experiments (A) and (B). Data are the mean ± S.E.M of six independent experiments (n=6) (***p < 0.0001).
To confirm this idea, we analyzed myofibril organization in human primary myotubes expressing HA-tagged wild-type TC10, a dominant negative mutant or GFP as a control. At 5 days post-fusion, cultures were immunostained for α-actinin and HA-tagged protein. Mature myofibrils, revealed by the cross striated organization of α-actinin, were observed in the majority of myotubes expressing GFP or wtTC10 (Fig. 7A). In contrast, most myotubes expressing the dominant negative form of TC10 (HA-TC10 DN) contained no mature myofibrils but only nascent myofibrils (Fig. 7A). These results show that TC10 signaling is critical for normal myofibril assembly during human skeletal differentiation.
Figure 7. Inhibition of TC10 expression or activity blocks myofibrillogenesis.

(A) Human myotubes were transfected with expression vectors for GFP or either HA-tagged wild-type (HA-TC10 wt) or a dominant negative TC10 (HA-TC10 DN, TC10/T31N). Cells were fixed and incubated with antibodies to the HA epitope and sarcomeric α-actinin as indicated, followed by fluorescently-labeled secondary antibodies. DNA was stained with DAPI. Proteins and DNA were detected by fluorescence microscopy: GFP or HA-tagged TC10 proteins (green), sarcomeric α-actinin (red) and nuclei (blue). The upper panels show the merged images and the lower panels show α-actinin staining. The results shown are typical of three independent experiments. Scale bars: 10 μm.
(B) Human myoblasts were transduced with lentiviral vectors expressing GFP and either a control shRNA (shCTL) or TC10 shRNA (shTC10). The cells were maintained in culture until terminally differentiated. Myotubes were fixed and immunostained for sarcomeric α-actinin (red), along with DAPI to visualize the nuclei (blue). The upper panels show the merge of GFP, α-actinin and DAPI fluorescence. The lower panels are a higher magnification of the area delimited by the square in the upper panel and show α-actinin staining. The results shown are typical of three independent experiments. Scale bars: 10 μm.
(C) Human myotubes described in (B) were analyzed for their myofibril content by α-actinin immunostaining. This revealed punctate filaments (premyofibrils), filaments that have begun to align (nascent myofibrils) and cross-striated myofibrils (mature myofibrils). Myotubes containing cross-striated myofibrils were counted as mature myofibrils. Myotubes containing aligned but not cross-striated myofibrils were counted as nascent myofibrils and other myotubes, containing only punctuated stain, were counted as pre-myofibrils. The percentage of myotubes displaying myofibrils at different stages of maturation (pre-, nascent or mature) was determined by immunostaining for sarcomeric α-actinin in three independent experiments.
Inhibition of TC10 expression by shRNA results in impaired myofibril maturation
To further confirm the importance of TC10 in myofibrillogenesis, we used a RNA interference to reduce TC10 levels and activity. Myoblasts were infected with lentiviral vectors containing TC10 or control shRNAs (shTC10 and shCtl). Five days post-fusion, myotubes were fixed and immunostained for α-actinin to evaluate myofibril organization. The majority of myotubes expressing the control shRNA showed mature myofibrils (Fig. 7B), while those expressing the shRNA directed against TC10 contained immature myofibrils (Fig. 7B). Quantitative analysis showed more than 72% of control myotubes contained mature myofibrils, compared to only 9% of myotubes where TC10 expression is disrupted (Fig. 7C). The latter did not change even after seven days in differentiating conditions (data not shown), indicating that myofibrillogenesis is efficiently blocked and not simply delayed. Taken together, our results demonstrate that TC10 signaling is required for normal myofibril organization in human skeletal muscle cells.
Discussion
Our study presents the first evidence for the function of TC10 in vertebrate skeletal muscle and identifies obscurin as a specific activator for this Rho GTPase. First, we show that TC10 expression and activity are required for myofibril assembly. Second, we provide evidence that TC10 binds directly to a predicted RhoGEF motif of the sarcomeric protein obscurin. Finally, we demonstrate that this RhoGEF domain is functional and specifically activates TC10 in human primary myotubes.
The implication of TC10 in myofibrillogenesis
TC10 exhibits a tissue-specific expression pattern with the highest level of TC10 mRNA found in the heart and skeletal muscle (Abe et al., 2003; Neudauer et al., 1998). These data suggest that TC10 plays an important and specific role in these tissues. In the mouse muscle cell line C2C12, TC10 mRNA is moderately expressed in undifferentiated myoblasts and is strongly induced in myotubes, remaining at a high level in terminally differentiated myotubes (Abe et al., 2003). In agreement with these results, we find that TC10 is not detectable in proliferating human primary myoblasts. It appears after myoblast fusion and is maintained during differentiation. This expression profile during differentiation suggests that TC10 does not play a role in myoblast fusion, as described for Rac (Luo et al., 1994) but rather is required for a later process. Consistent with this notion, TC10 is highly expressed in the heart where muscle cells do not fuse and are mononucleated. Surprisingly, while TC10 expression is stronger in striated muscle, its function remains unknown in this tissue. In adipocytes, TC10 participates in glucose transport, regulating insulin-induced translocation of the GLUT4 glucose transporter protein from intracellular compartments to the plama membrane (Chiang et al., 2001; Watson et al., 2001). However, adipocytes and muscle cells differ in their reliance on TC10 for insulin-induced actin remodeling (JeBailey et al., 2004). Jebailey et al. have shown that in L6 skeletal muscle cells, the dominant-negative mutant of TC10 does not prevent insulin-induced actin remodeling in either myoblasts or myotubes and does not interfere with insulin-mediated recruitment of GLUT4 to the cell surface (JeBailey et al., 2004). In the same cells, Klip proposed that Rac is the Rho GTPase necessary for insulin-induced actin remodeling and translocation (JeBailey et al., 2004; Khayat et al., 2000), leaving TC10 without any known function in skeletal muscle.
Our studies initially highlighted the importance of TC10 for myofibrillogenesis in the prochordate Ciona intestinalis (Coisy-Quivy et al., 2006). We expressed the dominant-negative mutants and constitutively active forms of each Rho family GTPase in striated muscle cells of the larva. Two Rho GTPases, RhoA and TC10 were required for myofibril assembly (Coisy-Quivy et al., 2006). The mechanisms underlying these phenotypes have not been determined in ascidia. In the muscle cell line C2C12, RhoA is required for serum response factor (SRF)-mediated activation of several muscle-specific gene promoters (Wei et al., 1998). Consequently, RhoA positively regulates MyoD expression and skeletal muscle cell differentiation. Here, we suggest that TC10 plays a different role than RhoA in muscle differentiation. Disruption of TC10 expression by shRNA in human primary myotube does not affect expression of the differentiation-specific proteins myogenin, obscurin and sarcomeric α-actinin. It leads to reduced levels of myosin heavy chain and troponin T, but overexpression of constitutively active TC10 does not induce the expression of these genes. Thus, our data implicate TC10 in sarcomere assembly and therefore in myofibrillogenesis in human muscle cells as in ascidia (Coisy-Quivy et al., 2006). This strongly suggests an important conserved function for this Rho GTPase in muscle differentiation.
Identification of Obscurin as an RhoGEF for TC10
Although Rho GTPases may influence different stages of muscle differentiation, the regulation of their activity in mature myotubes is largely uncharacterized. In particular, the mechanisms underlying the cross-talk between myofibril assembly and the modulation of Rho GTPases activity are still unclear and no regulator (RhoGEFs or RhoGAPs) of Rho GTPases has been identified. In searching for a RhoGEF that might be involved in TC10 activation during myofibrillogenesis, we focused on obscurin, a sarcomere-associated protein containing a putative GDP-GTP exchange domain (RhoGEF) (Young et al., 2001). This signaling domain is found in all orthologs of obscurin, namely in mammals, zebrafish, ascidia and nematode, suggesting an important conserved function in muscle differentiation (Raeker et al., 2006; Sutter et al., 2004; Young et al., 2001). Obscurin wraps around myofibrils at the level of the M-line and Z-disk in striated muscle (Kontrogianni-Konstantopoulos et al., 2003). Disruption of its expression in vertebrates (Borisov et al., 2006; Raeker et al., 2006) like invertebrates (Benian et al., 1996), leads to misorganization of myofibrillar structure. In consequence, Fukuzawa at al. suggest that obscurin signalling functions are necessary for sarcomere assembly (Fukuzawa et al., 2008). These results, together with our observations that implicate TC10 in myofibril organization, led us to test whether obscurin is a functional RhoGEF for TC10 during myofibrillogenesis. In human myotubes, TC10 binds directly to the RhoGEF domain of obscurin and, with a weaker affinity, a downstream region containing two immunoglobulin-like repeats that could stabilize the obscurin/TC10 interaction. Using Rho GTPase activation assays, we demonstrate that obscurin is a specific activator of TC10 but not Rac and Cdc42 in human primary myotubes. This important role of obscurin is confirmed by the observation that the expression of obscurin with TC10 increases the active phosphorylated form of PAK1 in human myotubes. These results show that obscurin is a functional and specific RhoGEF for TC10 in human skeletal muscle.
In embryonic hearts, Z-disk association of obscurin is mainly observed in cells where the actin cytoskeleton is not yet regularly aligned in parallel myofibrils, and M-line localization is observed when myofibrils start to be arranged in parallel bundles (Young et al., 2001). In mature cardiomyocytes, obscurin is found at the M-band of mature myofibrils (Young et al., 2001). This coincidence of the redistribution of obscurin with remodelling of the actin cytoskeleton suggests that obscurin may be involved in this structural transition. The identification of a functional Rho GEF domain in obscurin supports this idea, because Rho GTPases, including TC10, are known to be key regulators of actin cytoskeletal reorganization (Ridley, 2006). The precise molecular role of obscurin in the morphogenesis of the Z-disks, A-bands and M-lines is not known and is probably complex. Recent results suggest that the RhoGEF domain of obscurin interacts with RanBP9, a scaffolding protein, and that both can interact with the N-terminal region of titin to influence the formation of the Z-disk and A/I junction (Bowman et al., 2008). Given the moderate binding affinity of RanBP9 to obscurin, their interaction could play a transient role in myotube development (Bowman et al., 2008). In addition, RanBP9 might help stabilize the TC10/obscurin interaction. Since our results suggest that obscurin contains a functional RhoGEF motif, the functions of this protein likely involve signal transduction. Several mechanisms might link the obscurin/TC10-dependent signalling pathway to myofibril formation and organization. In terminally differentiated cardiac myocytes, desmin helps align myofibrils into striations and localizes at the Z-disk like obscurin. Desmin serves as an excellent substrate for PAK, which is an effector of TC10, and its phosphorylation by PAK dramatically inhibits its filament-forming ability (Ohtakara et al., 2000). Therefore, obscurin/TC10 pathways could control myofibril alignement via PAK and desmin. Obscurin, together with other interacting proteins like myomesin and titin (Fukuzawa et al., 2008), is also involved in the organization of A-bands and the incorporation of myosin filaments into myofibrils. These processes could be mediated by the regulatory myosin light chain (R-MLC) and PAK1. Indeed, PAK1 modulates R-MLC function via direct phosphorylation and inhibition of myosin light chain kinase (MLCK) (Bokoch, 2003). Since inhibition of MLCK disrupts thick filament assembly in embryonic myocytes in culture (Li et al., 2004), the phosphorylation of R-MLC seems essential for myofibril organization. The identification of targets of the obscurin/TC10-dependent signalling pathway will be essential to understand the mechanisms controling myofibril assembly.
In this study, we attribute a functional role to TC10 in human skeletal muscle and identify this RhoGTPase as an important signalling protein for sarcomere assembly. Our identification of a functional RhoGEF domain in obscurin indicates that direct communication between the sarcomere and TC10 pathway leads to new sarcomere formation. Since mutations in RhoGEFs are already associated with pathologies, such as cancer, and, given the phenotype of obscurin and TC10 disruption in striated muscle, we expect that some human skeletal myopathies arise from defects in obscurin/TC10 pathway.
Materials and Methods
DNA constructs
cDNAs encoding differents fragments of the carboxyterminal region of human Obscurin were generated by PCR from cDNA DKFZp451F056 (from clone DKFZp451F056) which encodes the C-terminus of Obscurin between aa 5173-6620. PCR products were subcloned into pDONR221 by Gateway technology (Invitrogen) to generate pDONR221 ObsCT(ΔDH-PH) (aa 6007-6620), pDONR221ObsCT (aa 5679-6620), pDONR221ObsDH (aa 5679-5895). Expression vectors were obtained by recombination of entry vector with destination vector pGEX 2T-RfB or pCDNA3 myc-RfC. cDNAs encoding HA-TC10 wt and HA-TC10 CA (TC10/Q75L) were inserted into pDONR221 by Gateway technology and expression vectors were obtained by recombination of entry vector with lentiviral destination vector. ds oligonucleotides encoding a TC10-specific shRNA, as well as a negative control ds oligonucleotides were cloned into pENTR/U6 vector. Lentiviral vector expressing shRNA were obtained by recombinaison of entry vector with destination vector. All constructs was verified by sequence analysis by using an ABI Prism Sequencer.
Antibodies
Antibodies were obtained as follow: anti-sarcomeric α-actinin mAb, anti-MHC fast mAb, anti-TC10 pAb, anti-Troponin T mAb and anti-Tubulin mAb from Sigma; anti-Caveolin 3 mAb, anti-Rac mAb, anti-CD56 mAb and anti-Myogenin mAb from BD Transduction Laboratories; anti Cdc42 pAb and anti-HA mAb from Santa cruz Biotechnology, anti-HA pAb from Zymed Laboratories, anti-GFP mAb from Roche, anti-GFP pAb from Torrey Pines Biolabs, anti MBP mAb from New England Biolabs; anti-myc pAb from Medical and Biological Laboratories; anti PAK pAb and anti P PAK pAb from Cell Signaling Technology. 9E10 anti-myc mAb was a gift from D. Matthieu and anti GST antiserum was a gift from A. Debant. The Alexa Fluor secondary antibodies were purchased from Molecular Probes, Invitrogen.
Cell cultures, transfection and lentiviral infection
Human primary myoblasts were isolated as described previously (Kitzmann et al., 2006) from quadriceps muscle obtained from the “AFM-BTR Banque de tissus pour la recherche” (Hôpital de la Pitié’-Salpêtrière, 75013 Paris). 30 000 myoblasts/ml were grown in GM (DMEM supplemented with 10% fetal calf serum (FCS) and 1% UltroserG (Biosepra)) during five days. At the fifth day of culture, myoblasts fuse and cultures were switched to differentiation medium (DM) composed of DMEM supplemented with 10% FCS, 1μM human Insulin and Penicillin/Streptomycin. Thus, the sixth day of culture is the first “day post fusion”. 293T cell lines were maintained in DMEM supplemented with 10% FCS and Penicillin/Streptomycin. 293T cells and human primary myotubes were transfected using jet PEI (Qbiogen) according to the manufacturer’s instructions. 72 hours after plating, Myoblasts were infected with lentivirus in GM supplemented with 8μg/ml Hexadimethrine bromide (Sigma). The medium was replaced after 24 hours and cells were left 72 hours in GM before switching to DM.
Immunoprecipitation and immunoblotting
293T cells and human myotubes were washed with ice-cold PBS and lysed with lysis buffer containing 50mM Tris-HCl pH 7.5, 150mM NaCl, 5mM MgCl2, 1% TritonX-100 and protease inhibitors (Sigma). Lysates were clarified by centrifugation and the supernatants were incubated with the indicated antibodies and Protein G–Sepharose. Immunocomplexes were washed with lysis buffer and bound proteins were analyzed by SDS/PAGE and immunoblotting. Analysis and quantification were performed with Kodak 1D3.6 software.
Recombinant protein purification and antibody production
GST and MBP fusions were expressed in E. coli strain BL21 using pGEX and pMAL vectors respectively, according to standard procedures. GST fusion proteins were eluted in buffer containing 10 mM Tris-HCl pH 7.5, 50mM reduced glutathione (Sigma) supplemented with 2mM MgCl2 and 10mM GDP for MBP-TC10 purification. Recombinant GST-ObsDH was further used for biochemical studies and GST-ObsCT(ΔDH-PH) for raising antibodies. Rabbits were submitted to three rounds of injection of 0.1μg of GST-ObsCT(ΔDH-PH). Immunosera were depleted on GST-bound Sepharose and affinity-purified on GST-Obs-bound Sepharose.
In vitro interaction
For in vitro interaction studies, 5μg of MBP-TC10 bound to amylose resin was incubated with 5μg of GST or GST-ObsDH in binding buffer (10mM Tris-Hcl pH 7.5, 150mM NaCl, 0.1% casamino acids, 0.1% Tween 20). Amylose resin were washed three times in ice-cold binding buffer. Total and amylose-bound proteins were denatured in Laemmli buffer and analyzed by Western blotting using anti-MBP and anti-GST antibodies.
GST-PBD pull down assay
Myotubes were transfected with myc-ObsCT or myc-ObsCT(ΔDH-PH). 72h after transfection, cells were rapidly washed in ice-cold PBS and lysed with lysis buffer. Lysates were centrifuged for 5min at 14,000g at 4°C, and samples were taken from the supernatant to estimate total protein concentration. 20mg of GST-PBD fusion bound to Sepharose beads were added to cell lysate and incubated for 1h at 4°C. Beads were washed four times in lysis buffer, and bound proteins were eluted in Laemmli buffer. Total proteins and PBD affinity-purified proteins were analyzed by Western blotting and the specific bands were quantified with Kodak 1D3.6 software.
Immunofluorescence and microscopy
Cells were fixed with 4% paraformaldehyde in PBS and permeabilized with 0.1% Triton X-100 in PBS. Cells were incubated with primary antibodies for 2h, followed by Alexa Fluor secondary antibodies and DAPI staining. Cell imaging was performed at the “Centre Regional d’imagerie Cellulaire de Montpellier”. Fluorescence was viewed with a Leica Microscope (Leica DM6000, Wetzlar, Germany) using a 63x/1.40-0.60 HCX PL APO grade oil objectives. Images were captured as 16 TIFF files with MicroMax 1300 CCD cameras (RS-Princeton Instruments Inc.) driven by the MetaMorph (version 7; Universal Imaging Corp.) software. Images were then restored with Huygens deconvolution software (Scientific Volume Imaging) and processed using ImageJ sofwtare as indicated.
Statistical analysis
All quantitative data are presented as means ± SEM. Statistical analysis to determine significance was performed using Student’s t tests. Differences were considered to be statistically significant at the p < 0.05 level.
Supplementary Material
Acknowledgments
We are grateful to A. Saltiel, A. Blangy, E. Bertrand, C. Mettling and P. Vaudin for the gifts of the TC10 vectors, MBP-TC10, pCDNA3myc-RfC and lentiviral destination vectors, respectively. We thank D. Mathieu and A. Debant for the gifts of anti-myc mAb and GST antiserum. We would like to thank the tissue bank for research (MYOBANK-AFM) of the Association Française contre les Myopathies (AFM) for generously providing human muscle biopsies. This work was supported by grants to A.P. from the AFM. M.C-Q. and O.T. were supported by the AFM and the Agence Nationale de Recherche contre le Sida (ANRS) respectively.
References
- Abe T, Kato M, Miki H, Takenawa T, Endo T. Small GTPase Tc10 and its homologue RhoT induce N-WASP-mediated long process formation and neurite outgrowth. J Cell Sci. 2003;116:155–68. doi: 10.1242/jcs.00208. [DOI] [PubMed] [Google Scholar]
- Bang ML, Centner T, Fornoff F, Geach AJ, Gotthardt M, McNabb M, Witt CC, Labeit D, Gregorio CC, Granzier H, et al. The complete gene sequence of titin, expression of an unusual approximately 700-kDa titin isoform, and its interaction with obscurin identify a novel Z-line to I-band linking system. Circ Res. 2001;89:1065–72. doi: 10.1161/hh2301.100981. [DOI] [PubMed] [Google Scholar]
- Benard V, Bohl BP, Bokoch GM. Characterization of rac and cdc42 activation in chemoattractant-stimulated human neutrophils using a novel assay for active GTPases. J Biol Chem. 1999;274:13198–204. doi: 10.1074/jbc.274.19.13198. [DOI] [PubMed] [Google Scholar]
- Benian GM, Tinley TL, Tang X, Borodovsky M. The Caenorhabditis elegans gene unc-89, required fpr muscle M-line assembly, encodes a giant modular protein composed of Ig and signal transduction domains. J Cell Biol. 1996;132:835–48. doi: 10.1083/jcb.132.5.835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bokoch GM. Biology of the p21-activated kinases. Annu Rev Biochem. 2003;72:743–81. doi: 10.1146/annurev.biochem.72.121801.161742. [DOI] [PubMed] [Google Scholar]
- Borisov AB, Kontrogianni-Konstantopoulos A, Bloch RJ, Westfall MV, Russell MW. Dynamics of obscurin localization during differentiation and remodeling of cardiac myocytes: obscurin as an integrator of myofibrillar structure. J Histochem Cytochem. 2004;52:1117–27. doi: 10.1369/jhc.3A6183.2004. [DOI] [PubMed] [Google Scholar]
- Borisov AB, Sutter SB, Kontrogianni-Konstantopoulos A, Bloch RJ, Westfall MV, Russell MW. Essential role of obscurin in cardiac myofibrillogenesis and hypertrophic response: evidence from small interfering RNA-mediated gene silencing. Histochem Cell Biol. 2006;125:227–38. doi: 10.1007/s00418-005-0069-x. [DOI] [PubMed] [Google Scholar]
- Bowman AL, Catino DH, Strong JC, Randall WR, Kontrogianni-Konstantopoulos A, Bloch RJ. The rho-guanine nucleotide exchange factor domain of obscurin regulates assembly of titin at the Z-disk through interactions with Ran binding protein 9. Mol Biol Cell. 2008;19:3782–92. doi: 10.1091/mbc.E08-03-0237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carnac G, Primig M, Kitzmann M, Chafey P, Tuil D, Lamb N, Fernandez A. RhoA GTPase and serum response factor control selectively the expression of MyoD without affecting Myf5 in mouse myoblasts. Mol Biol Cell. 1998;9:1891–902. doi: 10.1091/mbc.9.7.1891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiang SH, Baumann CA, Kanzaki M, Thurmond DC, Watson RT, Neudauer CL, Macara IG, Pessin JE, Saltiel AR. Insulin-stimulated GLUT4 translocation requires the CAP-dependent activation of TC10. Nature. 2001;410:944–8. doi: 10.1038/35073608. [DOI] [PubMed] [Google Scholar]
- Chong C, Tan L, Lim L, Manser E. The mechanism of PAK activation. Autophosphorylation events in both regulatory and kinase domains control activity. J Biol Chem. 2001;276:17347–53. doi: 10.1074/jbc.M009316200. [DOI] [PubMed] [Google Scholar]
- Coisy-Quivy M, Sanguesa-Ferrer J, Weill M, Johnson DS, Donnay JM, Hipskind R, Fort P, Philips A. Identification of Rho GTPases implicated in terminal differentiation of muscle cells in ascidia. Biol Cell. 2006;98:577–88. doi: 10.1042/BC20060032. [DOI] [PubMed] [Google Scholar]
- Etienne-Manneville S, Hall A. Rho GTPases in cell biology. Nature. 2002;420:629–35. doi: 10.1038/nature01148. [DOI] [PubMed] [Google Scholar]
- Fukuzawa A, Lange S, Holt M, Vihola A, Carmignac V, Ferreiro A, Udd B, Gautel M. Interactions with titin and myomesin target obscurin and obscurin-like 1 to the M-band - implications for hereditary myopathies. J Cell Sci. 2008;121:1841–51. doi: 10.1242/jcs.028019. [DOI] [PubMed] [Google Scholar]
- Gautel M, Mues A, Young P. Control of sarcomeric assembly: the flow of information on titin. Rev Physiol Biochem Pharmacol. 1999;138:97–137. doi: 10.1007/BFb0119625. [DOI] [PubMed] [Google Scholar]
- Gupte A, Mora S. Activation of the Cbl insulin signaling pathway in cardiac muscle; dysregulation in obesity and diabetes. Biochem Biophys Res Commun. 2006;342:751–7. doi: 10.1016/j.bbrc.2006.02.023. [DOI] [PubMed] [Google Scholar]
- Hart MJ, Eva A, Evans T, Aaronson SA, Cerione RA. Catalysis of guanine nucleotide exchange on the CDC42Hs protein by the dbl oncogene product. Nature. 1991;354:311–4. doi: 10.1038/354311a0. [DOI] [PubMed] [Google Scholar]
- JeBailey L, Rudich A, Huang X, Di Ciano-Oliveira C, Kapus A, Klip A. Skeletal muscle cells and adipocytes differ in their reliance on TC10 and Rac for insulin-induced actin remodeling. Mol Endocrinol. 2004;18:359–72. doi: 10.1210/me.2003-0294. [DOI] [PubMed] [Google Scholar]
- Khayat ZA, Tong P, Yaworsky K, Bloch RJ, Klip A. Insulin-induced actin filament remodeling colocalizes actin with phosphatidylinositol 3-kinase and GLUT4 in L6 myotubes. J Cell Sci. 2000;113(Pt 2):279–90. doi: 10.1242/jcs.113.2.279. [DOI] [PubMed] [Google Scholar]
- Kitzmann M, Bonnieu A, Duret C, Vernus B, Barro M, Laoudj-Chenivesse D, Verdi JM, Carnac G. Inhibition of Notch signaling induces myotube hypertrophy by recruiting a subpopulation of reserve cells. J Cell Physiol. 2006;208:538–48. doi: 10.1002/jcp.20688. [DOI] [PubMed] [Google Scholar]
- Kontrogianni-Konstantopoulos A, Jones EM, Van Rossum DB, Bloch RJ. Obscurin is a ligand for small ankyrin 1 in skeletal muscle. Mol Biol Cell. 2003;14:1138–48. doi: 10.1091/mbc.E02-07-0411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kontrogianni-Konstantopoulos A, Catino DH, Strong JC, Randall WR, Bloch RJ. Obscurin regulates the organization of myosin into A bands. Am J Physiol Cell Physiol. 2004;287:C209–17. doi: 10.1152/ajpcell.00497.2003. [DOI] [PubMed] [Google Scholar]
- Kontrogianni-Konstantopoulos A, Catino DH, Strong JC, Sutter S, Borisov AB, Pumplin DW, Russell MW, Bloch RJ. Obscurin modulates the assembly and organization of sarcomeres and the sarcoplasmic reticulum. FASEB J. 2006;20:2102–11. doi: 10.1096/fj.06-5761com. [DOI] [PubMed] [Google Scholar]
- Labeit S, Kolmerer B. The complete primary structure of human nebulin and its correlation to muscle structure. J Mol Biol. 1995;248:308–15. doi: 10.1016/s0022-2836(95)80052-2. [DOI] [PubMed] [Google Scholar]
- Li H, Cook JD, Terry M, Spitzer NC, Ferrari MB. Calcium transients regulate patterned actin assembly during myofibrillogenesis. Dev Dyn. 2004;229:231–42. doi: 10.1002/dvdy.10428. [DOI] [PubMed] [Google Scholar]
- Luo L, Liao YJ, Jan LY, Jan YN. Distinct morphogenetic functions of similar small GTPases: Drosophila Drac1 is involved in axonal outgrowth and myoblast fusion. Genes Dev. 1994;8:1787–802. doi: 10.1101/gad.8.15.1787. [DOI] [PubMed] [Google Scholar]
- Maruyama K. Connectin, an elastic protein from myofibrils. J Biochem. 1976;80:405–7. doi: 10.1093/oxfordjournals.jbchem.a131291. [DOI] [PubMed] [Google Scholar]
- Mayans O, van der Ven PF, Wilm M, Mues A, Young P, Furst DO, Wilmanns M, Gautel M. Structural basis for activation of the titin kinase domain during myofibrillogenesis. Nature. 1998;395:863–9. doi: 10.1038/27603. [DOI] [PubMed] [Google Scholar]
- Neudauer CL, Joberty G, Tatsis N, Macara IG. Distinct cellular effects and interactions of the Rho-family GTPase TC10. Curr Biol. 1998;8:1151–60. doi: 10.1016/s0960-9822(07)00486-1. [DOI] [PubMed] [Google Scholar]
- Nolan KM, Barrett K, Lu Y, Hu KQ, Vincent S, Settleman J. Myoblast city, the Drosophila homolog of DOCK180/CED-5, is required in a Rac signaling pathway utilized for multiple developmental processes. Genes Dev. 1998;12:3337–42. doi: 10.1101/gad.12.21.3337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohtakara K, Inada H, Goto H, Taki W, Manser E, Lim L, Izawa I, Inagaki M. p21-activated kinase PAK phosphorylates desmin at sites different from those for Rho-associated kinase. Biochem Biophys Res Commun. 2000;272:712–6. doi: 10.1006/bbrc.2000.2854. [DOI] [PubMed] [Google Scholar]
- Person V, Kostin S, Suzuki K, Labeit S, Schaper J. Antisense oligonucleotide experiments elucidate the essential role of titin in sarcomerogenesis in adult rat cardiomyocytes in long-term culture. J Cell Sci. 2000;113(Pt 21):3851–9. doi: 10.1242/jcs.113.21.3851. [DOI] [PubMed] [Google Scholar]
- Raeker MO, Su F, Geisler SB, Borisov AB, Kontrogianni-Konstantopoulos A, Lyons SE, Russell MW. Obscurin is required for the lateral alignment of striated myofibrils in zebrafish. Dev Dyn. 2006;235:2018–29. doi: 10.1002/dvdy.20812. [DOI] [PubMed] [Google Scholar]
- Ridley AJ. Rho GTPases and actin dynamics in membrane protrusions and vesicle trafficking. Trends Cell Biol. 2006;16:522–9. doi: 10.1016/j.tcb.2006.08.006. [DOI] [PubMed] [Google Scholar]
- Rossman KL, Der CJ, Sondek J. GEF means go: turning on RHO GTPases with guanine nucleotide-exchange factors. Nat Rev Mol Cell Biol. 2005;6:167–80. doi: 10.1038/nrm1587. [DOI] [PubMed] [Google Scholar]
- Sutter SB, Raeker MO, Borisov AB, Russell MW. Orthologous relationship of obscurin and Unc-89: phylogeny of a novel family of tandem myosin light chain kinases. Dev Genes Evol. 2004;214:352–9. doi: 10.1007/s00427-004-0413-5. [DOI] [PubMed] [Google Scholar]
- Trinick J. Titin as a scaffold and spring. Cytoskeleton. Curr Biol. 1996;6:258–60. doi: 10.1016/s0960-9822(02)00472-4. [DOI] [PubMed] [Google Scholar]
- Trinick J, Tskhovrebova L. Titin: a molecular control freak. Trends Cell Biol. 1999;9:377–80. doi: 10.1016/s0962-8924(99)01641-4. [DOI] [PubMed] [Google Scholar]
- Van Aelst L, D’Souza-Schorey C. Rho GTPases and signaling networks. Genes Dev. 1997;11:2295–322. doi: 10.1101/gad.11.18.2295. [DOI] [PubMed] [Google Scholar]
- van der Ven PF, Bartsch JW, Gautel M, Jockusch H, Furst DO. A functional knock-out of titin results in defective myofibril assembly. J Cell Sci. 2000;113 (Pt 8):1405–14. doi: 10.1242/jcs.113.8.1405. [DOI] [PubMed] [Google Scholar]
- Wang K, Knipfer M, Huang QQ, van Heerden A, Hsu LC, Gutierrez G, Quian XL, Stedman H. Human skeletal muscle nebulin sequence encodes a blueprint for thin filament architecture. Sequence motifs and affinity profiles of tandem repeats and terminal SH3. J Biol Chem. 1996;271:4304–14. doi: 10.1074/jbc.271.8.4304. [DOI] [PubMed] [Google Scholar]
- Wang K, McClure J, Tu A. Titin: major myofibrillar components of striated muscle. Proc Natl Acad Sci U S A. 1979;76:3698–702. doi: 10.1073/pnas.76.8.3698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watson RT, Shigematsu S, Chiang SH, Mora S, Kanzaki M, Macara IG, Saltiel AR, Pessin JE. Lipid raft microdomain compartmentalization of TC10 is required for insulin signaling and GLUT4 translocation. J Cell Biol. 2001;154:829–40. doi: 10.1083/jcb.200102078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei L, Zhou W, Croissant JD, Johansen FE, Prywes R, Balasubramanyam A, Schwartz RJ. RhoA signaling via serum response factor plays an obligatory role in myogenic differentiation. J Biol Chem. 1998;273:30287–94. doi: 10.1074/jbc.273.46.30287. [DOI] [PubMed] [Google Scholar]
- Young P, Ehler E, Gautel M. Obscurin, a giant sarcomeric Rho guanine nucleotide exchange factor protein involved in sarcomere assembly. J Cell Biol. 2001;154:123–36. doi: 10.1083/jcb.200102110. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.