

Abstract
Background
Vaccinia virus gene B1R encodes a serine/threonine protein kinase. In vitro this protein kinase phosphorylates ribosomal proteins Sa and S2 and vaccinia virus protein H5R, proteins that become phosphorylated during infection. Nothing is known about the sites phosphorylated on these proteins or the general substrate specificity of the kinase. The work described is the first to address these questions.
Results
Vaccinia virus protein H5R was phosphorylated by the B1R protein kinase in vitro, digested with V8 protease, and phosphopeptides separated by HPLC. The N-terminal sequence of one radioactively labelled phosphopeptide was determined and found to correspond to residues 81-87 of the protein, with Thr-84 and Thr-85 being phosphorylated. A synthetic peptide based on this region of the protein was shown to be a substrate for the B1R protein kinase, and the extent of phosphorylation was substantially decreased if either Thr residue was replaced by an Ala.
Conclusions
We have identified the first phosphorylation site for the vaccinia virus B1R protein kinase. This gives important information about the substrate-specificity of the enzyme, which differs from that of other known protein kinases. It remains to be seen whether the same site is phosphorylated in vivo.
Background
Vaccinia virus is a large DNA virus that replicates in the host cell cytoplasm in granular sites called virosomes [1]. It encodes at least two protein kinases belonging to the cellular family of serine/threonine protein kinases, the products of the B1R [2,3] and F10L genes [4]. The F10L kinase is encapsidated in the virion and plays an essential role in virion morphogenesis [5,6]. The B1R protein kinase is expressed early in infection, is found in the virosomes, and is also packaged into virions [7]. It appears to be an essential viral protein, and temperature-sensitive mutations that map to the B1R gene produce virus that cannot replicate its DNA at the restrictive temperature [2,8]. The B1R kinase does not appear to have a broad substrate specificity, and, although it has some activity against the acidic protein, casein, this is a poor substrate compared with the enzyme's known physiological substrates. Three proteins which become phosphorylated during infection of cells with vaccinia virus have been shown to be substrates of the B1R protein kinase in vitro. Two of these are the ribosomal proteins Sa and S2 [9,10] and the third is the product of the H5R open reading frame of the vaccinia virus genome [10,11].
The fact that the B1R kinase phosphorylates these proteins in vitro does not, of course, prove that it is responsible for their phosphorylation during infection by virus in vivo. However, one piece of evidence consistent with this possibility is that all these proteins have multiple phosphorylation sites predominantly involving threonine (rather than the more usual serine) residues, and it is threonine residues on the proteins that the B1R kinase phosphorylates in vitro. In the case of protein H5R - the subject of the current work - there is further reason to believe that the B1R kinase contributes to the phosphorylation in vivo. In a mutant strain of virus, temperature-sensitive for B1R, the proportion of underphosophorylated H5R decreases at the restrictive temperature (G. Beaud and R. Beaud, unpublished).
In cells infected with a temperature-sensitive mutant of the B1R gene, the proportion of underphosphorylated H5R protein decreases at the restrictive temperature, showing that the B1R protein kinase controls the phosphorylation state of the H5R protein synthesized at the early stage of vaccinia virus infection [12]. It has recently been shown that most of the H5R protein is found in virosomes, although some of the more highly phosphorylated forms of the protein appear to be cytoplasmic, suggesting multiple roles in vaccinia virus development [13]. Kovacs and Moss [14] have demonstrated that the H5R protein is, in fact, equivalent to the late stage-specific transcription factor VLTF-4, and Black et al. [15] have showed that it associates with protein G2R, a putative late transcription elongation factor. In contrast, studies of a dominant temperature-sensitive mutant of H5R by DeMasi and Traktman [16] suggest a role in virion morphogenesis. A knowledge of the phosphorylation sites on the H5R protein is needed to test whether phosphorylation has a role in either of these processes, and we have made the first steps in this direction by identifying two threonine residues in the protein that are substrates for the B1R protein kinase.
Results
Vaccinia virus protein H5R isolated from infected HeLa cells was labelled in vitro with [γ-32P]ATP by recombinant B1R protein kinase, all as described in Experimental. The products of the reaction were digested with V8 protease and then passed through a SEP-PAK cartridge to remove the majority of the unreacted [γ-32P]ATP. The radioactive V8 peptides recovered from the cartridge were then applied to a reverse-phase HPLC column and eluted with a gradient of acetonitrile. This removed the residual [γ-32P]ATP, and the majority of the radioactivity associated with peptides eluted as a doublet at 20-30% acetonitrile (designated 3 in Fig. 1a), which was collected, concentrated by centrifugal evaporation, redissolved and applied to a microbore HPLC column to resolve the peptides further. A first run using a 15-25% gradient of acetonitrile in 0.1% trifluoroacetic acid revealed that this preparation of the radioactive doublet contained perhaps a dozen peptides (Fig. 1b). As these were incompletely resolved, two regions of the gradient containing the most radioactive peaks (designated in the figure as I and II, containing gradient fractions 30-33 and 35-36, respectively) were collected and reapplied in turn to the microbore column, but this time the gradient for elution comprised 0-50% acetonitrile in 20 mM NaCl without trifluoroacetic acid. Insufficient radioactivity was recovered from the peaks resolved in fraction I for sequence analysis. Fraction II yielded more radioactive peptides, separated from adjacent peptides and partly resolved from each other (peaks 28 and 29 in Fig 1c).
Figure 1.
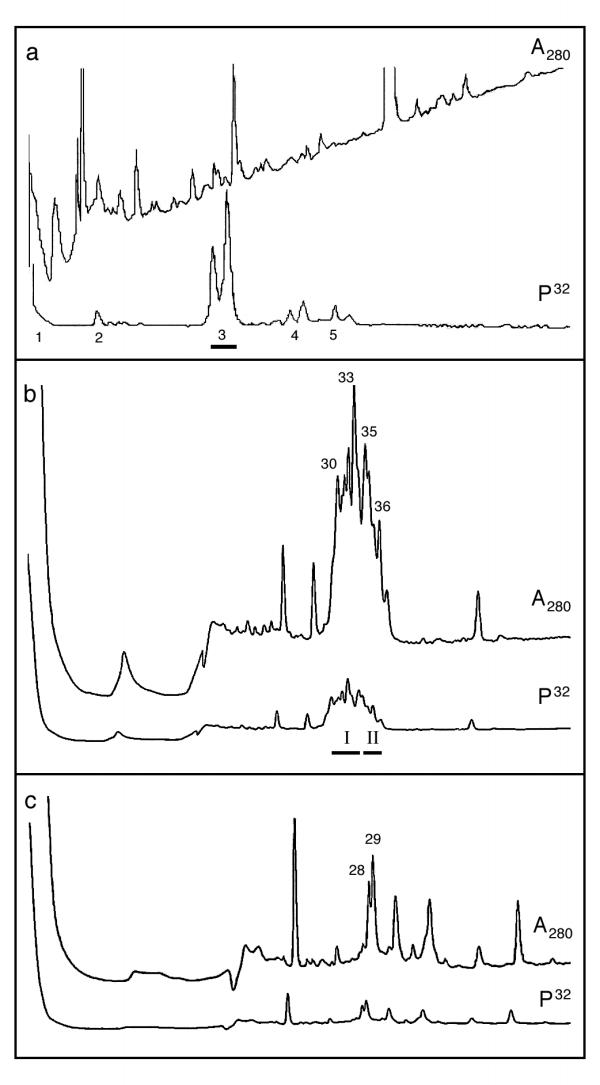
HPLC fractionation of V8 peptides from vaccinia virus protein H5R. (a) HPLC fractionation of hydrolysate after passage through a SEP-PAK column using a gradient of 0-50% acetonitrile. The fractions encompassed by the horizontal bar (region 3) were collected for further analysis. (b) Microbore HPLC fractionation of region 3 from (a). A gradient of 15-25% acetonitrile in 0.1% trifluoroacetic acid was used and the fractions encompassed by the horizontal bars (regions I and II) collected for further analysis). (c) Microbore HPLC fractionation of region II from (b). A gradient of 0-50% acetonitrile in 20 mM NaCl (in the absence of trifluoroacetic acid - i.e. at neutral pH) was used and the fractions 28 and 29 collected for sequence analysis). The direction of increasing gradient is from left to right in all three frames; the ultraviolet absorbance trace is labelled as A280, and the radioactivity trace as 32P.
The two peaks, 28 and 29, from the microbore HPLC separation of fraction II (Fig. 1c) were separately subjected to sequence analysis using an Applied Biosystems gas phase sequencer, and the radioactivity of the PTH derivative at each cycle of Edman degradation was determined by scintillation spectrometry. The results of thirteen cycles of automatic Edman degradation on peak 29 are shown in Fig 2. The first seven residues of the peptide were identified as YHQTTEK, which correspond exactly to residues 81-87 of H5R and do not occur elsewhere in the protein [17]. (The preceding residue 80 in the protein is glutamic acid, consistent with the substrate specificity of V8 protease.) The 32P was released from the peptide predominantly at cycles 4, 5 and 6. As there is always carry-over from one cycle to the next, we can conclude that both residues Thr-84 and Thr-85 of H5R are phosphorylated by the vaccinia virus B1R protein kinase. Analysis of fraction 28 gave similar results to those for fraction 29 (not shown).
Figure 2.

Sequence analysis of H5R phosphopeptide. Fraction 29 from Fig 1c was subjected to automatic Edman degradation, and the 32P radioactivity released at each stage measured. The identity of the phenylthiohydantoin derivative from the first seven cycles was unequivocal and is indicated.
In many cases it has been found that peptides containing the phosphorylation sites of proteins are also substrates for the protein kinase catalysing the phosphorylation of the protein. We therefore synthesized the peptide RRIEEYHQTTEKN, which represents amino acid residues 78-88 of H5R preceded by two arginine residues (not present at positions 76-77 of the protein) to make the peptide easier to handle. As preliminary studies indicated that this served as a substrate for the B1R kinase we had two additional peptides constructed in one of which Ala replaced Thr-84 and in the other of which it replaced Thr-85. After incubation with B1R kinase and [γ-32P]ATP the peptides were subjected to high-voltage thin layer electrophoresis to resolve the peptides from the unreacted [γ-32P]ATP. Fig 3 shows that although there was some labelling of the two Ala-replacement peptides, this was much less than with the parent peptide. (Measurement indicated that this was about 10% of incorporation into the parent peptide). It would therefore appear that the recognition specificity for phosphorylation at either Thr residue includes the other Thr, despite the fact that in one case it is on the N-terminal side of the phosphorylated residue, and in the other case on the C-terminal side.
Figure 3.

Phosphorylation of synthetic peptides. Peptides were phosphorylated as described in the Experimental section and subjected to electrophoresis on thin-layer cellulose at pH 3.5. The position of the phosphopeptides is indicated by the arrow, the identification being based on the fact that the ninhydrin-stained material (predominantly unphosphorylated and hence with a greater positive charge) migrated just ahead of this. Lane 1 contained RRIEEYHQTTEKN, lane 2: RRIEEYHQATEKN, lane 3: RRIEEYHQTAEKN, and lane 4; no peptide.
Discussion
We have identified two phosphorylation sites for vaccinia virus protein kinase B1R in viral protein H5R, the first phosphorylation sites to be determined for this protein kinase. Although we have not established whether these same sites are also phosphorylated during viral infection, our work provides important biochemical information on the substrate specificity of the protein kinase. The sequence containing the two phosphorylated threonine residues - EEYHQTTEKNSP - is neither particularly basic or acidic, consistent with the failure of basic or acidic model peptides to serve as substrates for the B1R protein kinase. It is interesting that this does not resemble sequences reported to be phosphorylated by other protein kinases, including those phosphorylated by CK1, the known cellular protein kinase most closely related to the B1R kinase. This latter generally requires a phosphoserine three amino acids to the N-terminal side of the phosphate-acceptor site, although in some artificial substrates a highly acidic N-terminal region can substitute for this [18]. One interesting biochemical question is what residues in the target sequence we have identified are the determinants for the substrate-specificity for the B1R protein kinase. In one approach to this we have examined the sequences of other known substrates for the B1R kinase (ribosomal proteins Sa and S2) but have been unable to identify potential similarities to the H5R phosphopeptide. In this regard it should be mentioned that although the synthetic peptide RRIEEYHQTTEKN did serve as a substrate for the enzyme (Fig. 3), we found its phosphorylation to be weak compared with that produced by other protein kinases on their model peptides (Approximately 0.1 mol phosphate per mol of peptide, compared with 1.8-2.0 mol phosphate per mol H5R protein). Although the additional arginine residues in the peptide complicate the interpretation, it may well be that there are three-dimensional determinants - absent from the synthetic peptide - that contribute to the substrate specificity of the enzyme. There are precedents for this in some other protein kinases [18].
The vaccinia protein H5R has been shown by isoelectric focusing to exist in at least four differently charged forms [11,13], suggesting that there are at least least five different phosphorylation sites on the protein. Consistent with this are the results of experiments (not shown) in which we employed recombinant H5R in which Thr-84 and Thr-85 had been replaced by Ala residues, and found that this still served as a substrate for the B1R protein kinase. It is possible that one or more of these additional sites is present in the HPLC fraction I of Fig 1b, the peptides resolved from which were insufficiently radioactive for sequence analysis.
Although this work represents the first identification of phosphorylation sites for the vaccinia B1R kinase, it remains to be established whether these same sites are phosphorylated in vivo, and, if so, with what physiological significance. However it should now be possible to address this question by constructing recombinant virus with amino acid substitutions at the positions of phosphorylation.
Conclusions
Vaccinia virus protein kinase B1R phosphorylates the virus protein H5R in vitro at the threonine residues Thr-84 and Thr-85 within the region:
EEYHQTTEKNSP
A synthetic peptide based on this sequence also acted as a substrate. We conclude that this sequence determines, at least in part, the substrate specificity of the vaccinia B1R protein kinase, although it is unclear which amino acid residues are the key determinants within this sequence. There are other phosphorylation sites for the kinase on protein H5R, but these remain to be determined.
Materials and Methods
Preparation and labelling of H5R protein
The preparation of authentic vaccinia H5R protein and recombinant B1R protein kinase were as previously described [11]. In some experiments a trpE-H5R fusion protein (pATH11-Ag35) was used [19]. For phosphorylation, three identical reactions contained H5R protein (70 pmol), B1R protein kinase (90 μl), Tris-HCl, pH 7.4 (20 mM), magnesium chloride (5 mM), ATP (50 μM), [γ-32P]ATP (50 μCi) and dithiothreitol (2 mM) in a total volume of 500 μl. Incubation was for 30 min at 30° C.
Proteolytic digestion of H5R protein
The 500 μl reaction mixtures, above, were adjusted to 50 mM Tris-HCl, pH 7.4, and 0.01% reduced Triton X-100 at a final volume of 600 μl. To this was added 0.4 μg V8 protease (Boehringer Mannheim) and incubation carried out at 30° C for 18 h. To prepare the peptides for HPLC analysis the reaction mixtures were pooled and applied to a SEP-PAK cartridge (prewashed with successive 10 ml portions of 50% acetonitrile and water) and eluted with water (40 ml) followed by 50% acetonitrile (40 ml) and finally 100% acetonitrile (30 ml). A large peak of radioactivity (unreacted ATP) eluted with the water and was discarded, and a smaller broad peak of radioactivity that eluted with 50% acetonitrile was retained and concentrated to 200 μl by rotary evaporation.
Purification of H5R peptides and sequence analysis
Initial purification was with a Vydac protein and Peptide C18 column (25 × 04 cm) on a Gilson HPLC system, and this was followed by further purification on a Vydac C18 2.1 × 180 mm microbore column. Details of the gradients used are given in the text. Peptides were sequenced on an Applied Biosystems 476A protein sequencer and phosphorylation sites were analysed using solid phase Edmann sequencing [20].
Labelling and analysis of synthetic peptides
Synthetic peptides were purchased from Thistle Research, Glasgow, UK. Each peptide (3 mM) was incubated with [γ-32P]ATP (6.3 μCi) B1R protein kinase (4 μl), Tris-HCl, pH 7.4 (20 mM), magnesium chloride (5 mM), ATP (50 μM), and dithiothreitol (2 mM) in a total volume of 20 μl. Incubation was for 30 min at 30° C. The reaction mixtures were applied in 1 cm strips to thin layer cellulose plates and subjected to electrophoresis for 4 h at 200 V in a solution of pyridine : acetic acid : water (20:200:1780) at pH 3.5. The plates were dried, stained with ninhydrin to locate the unphosphorylated peptides, and subjected to autoradiography.
Acknowledgments
Acknowledgements
We thank the Wellcome Trust for support.
Contributor Information
Neil G Brown, Email: n.g.brown@bio.gla.ac.uk.
D Nick Morrice, Email: dnmorrice@dux.dundee.ac.uk.
Georges Beaud, Email: beaud@ijm.jussieu.fr.
Grahame Hardie, Email: dghardie@dux.dundee.ac.uk.
David P Leader, Email: d.leader@bio.gla.ac.uk.
References
- Moss B. Poxviridae: the viruses and their replication. In Fields Virology, 3rd edn (Edited by Field BN, Knipe DM, Howley, PM) Philadelphia: Lippincott-Raven, 1996. pp. 2637–2671.
- Rempel RE, Traktman P. Vaccinia virus B1 kinase: phenotypic analysis of temperature-sensitive mutants and enzymatic characterization of recombinant proteins. J Virol. 1992;66:4413–4426. doi: 10.1128/jvi.66.7.4413-4426.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin SQ, Wen C, Broyles SS. The vaccinia virus B1R gene product is a serine/threonine protein kinase. J Virol. 1992;66:2717–2723. doi: 10.1128/jvi.66.5.2717-2723.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin S, Broyles SS. Vaccinia protein kinase 2: a second essential serine/threonine protein kinase encoded by vaccinia virus. Proc Natl Acad Sci U S A. 1994;91:7653–7657. doi: 10.1073/pnas.91.16.7653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Traktman P, Caligiuri A, Jesty SA, Liu K, Sankar U. Temperature-sensitive mutants with lesions in the vaccinia virus F10 kinase undergo arrest at the earliest stage of virion morphogenesis. J Virol. 1995;69:6581–6587. doi: 10.1128/jvi.69.10.6581-6587.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang S, Shuman S. Vaccinia virus morphogenesis is blocked by temperature-sensitive mutations in the F10 gene, which encodes protein kinase 2. J Virol. 1995;69:6376–88. doi: 10.1128/jvi.69.10.6376-6388.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banham AH, Smith GL. Vaccinia virus gene B1R encodes a 34-kDa serine/threonine protein kinase that localizes in cytoplasmic factories and is packaged into virions. Virology. 1992;191:803–812. doi: 10.1016/0042-6822(92)90256-o. [DOI] [PubMed] [Google Scholar]
- Rempel RE, Anderson MK, Evans E, Traktman P. Temperature-sensitive vaccinia virus mutants identify a gene with an essential role in viral replication. J Virol. 1990;64:574–583. doi: 10.1128/jvi.64.2.574-583.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banham AH, Leader DP, Smith GL. Phosphorylation of ribosomal proteins by the vaccinia virus B1R protein kinase. FEBS Lett. 1993;321:27–31. doi: 10.1016/0014-5793(93)80614-Z. [DOI] [PubMed] [Google Scholar]
- Beaud G, Sharif A, Topa-Masse A, Leader DP. Ribosomal protein S2/Sa kinase purified from HeLa cells infected with vaccinia virus corresponds to the B1R protein kinase and phosphorylates in vitro the viral ssDNA-binding protein. J Gen Virol. 1994;75:283–293. doi: 10.1099/0022-1317-75-2-283. [DOI] [PubMed] [Google Scholar]
- Beaud G, Beaud R, Leader DP. Vaccinia virus gene H5R encodes a protein that is phosphorylated by the multisubstrate vaccinia virus B1R protein kinase. J Virol. 1995;69:1819–1826. doi: 10.1128/jvi.69.3.1819-1826.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beaud G, Beaud R. Temperature-dependent phosphorylation state of the H5R protein synthesised at the early stage of infection in cells infected with vaccinia virus ts mutants of the B1R and F10L protein kinases. Intervirology. 2000;43:67–70. doi: 10.1159/000025025. [DOI] [PubMed] [Google Scholar]
- Beaud G, Beaud R. Preferential virosomal location of underphosphorylated H5R protein synthesized in vaccinia virus-infected cells. J Gen Virol. 1997;78:3297–3302. doi: 10.1099/0022-1317-78-12-3297. [DOI] [PubMed] [Google Scholar]
- Kovacs GR, Moss B. The vaccinia virus H5R gene encodes late gene transcription factor 4: purification, cloning, and overexpression. J Virol. 1996;70:6796–6802. doi: 10.1128/jvi.70.10.6796-6802.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Black EP, Moussache N, Condit RC. Characterization of the interactions among vaccinia virus transcription factors G2R, A18R, and H5R. . Virology. 1998;245:313–322. doi: 10.1006/viro.1998.9166. [DOI] [PubMed] [Google Scholar]
- DeMasi J, Traktman P. Clustered charge-to-alanine mutagenesis of the vaccina virus H5 gene: isolation of a dominant temperature-sensitive mutant with a profound defect in morphogenesis. J Virol. 2000;74:2393–2405. doi: 10.1128/JVI.74.5.2393-2405.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goebel SJ, Johnson GP, Perkus ME, Davis SW, Winslow JP, Paoletti E. The complete sequence of vaccinia virus. Virology. 1990;179:247–266. doi: 10.1016/0042-6822(90)90294-2. [DOI] [PubMed] [Google Scholar]
- Pinna LA, Ruzzene M. How do protein kinases recognize their substrates? Biochim Biophys Acta. 1996;1314:191–225. doi: 10.1016/S0167-4889(96)00083-3. [DOI] [PubMed] [Google Scholar]
- Mohandas AR, Dekaban GA, Dales S. Vaccinia virion surface polypeptide Ag35 expressed from a baculovirus vector is targeted to analogous poxvirus and insect virus components. Virology. 1994;200:207–219. doi: 10.1006/viro.1994.1179. [DOI] [PubMed] [Google Scholar]
- Ben-Levy R, Leishton IA, Doza YN, Attwood P, Morrice N, Marshall CJ, Cohen P. Identification of novel phosphorylation sites required for activation of MAPKAP kinase-2. EMBO J. 1995;23:5920–5930. doi: 10.1002/j.1460-2075.1995.tb00280.x. [DOI] [PMC free article] [PubMed] [Google Scholar]