

Abstract
Background
In vertebrates, two types of cholinesterases exist, acetylcholinesterase and butyrylcholinesterase. The function of acetylcholinesterase is to hydrolyse acetylcholine, thereby terminating the neurotransmission at cholinergic synapse, while the precise physiological function of butyrylcholinesterase has not been identified. The presence of cholinesterases in tissues that are not cholinergically innervated indicate that cholinesterases may have functions unrelated to neurotransmission. Furthermore, cholinesterases display a genuine aryl acylamidase activity apart from their predominant acylcholine hydrolase activity. The physiological significance of this aryl acylamidase activity is also not known. The study on the aryl acylamidase has been, in part hampered by the lack of a specific method to visualise this activity. We have developed a method to visualise the aryl acylamidase activity on cholinesterase in polyacrylamide gels.
Results
The o-nitroaniline liberated from o-nitroacetanilide by the action of aryl acylamidase activity on cholinesterases, in the presence of nitrous acid formed a diazonium compound. This compound gave an azo dye complex with N-(1-napthyl)-ethylenediamine, which appeared as purple bands in polyacrylamide gels. Treating the stained gels with trichloroacetic acid followed by Tris-HCl buffer helped in fixation of the stain in the gels. By using specific inhibitors for acetylcholinesterase and butyrylcholinesterase, respectively, differential staining for the aryl acylamidase activities on butyrylcholinesterase and acetylcholinesterase in a sample containing both these enzymes has been demonstrated. A linear relationship between the intensity of colour developed and activity of the enzyme was obtained.
Conclusions
A novel method to visualise the aryl acylamidase activity on cholinesterases in polyacrylamide gels has been developed.
Background
Cholinesterases (ChEs) are evolutionarily conserved type B carboxylesterase enzymes that share extensive sequence homology. In vertebrates two types of ChEs were identified based on their distinct substrate specificity and inhibitor sensitivity. The acetylcholinesterase (AChE; EC 3.1.1.7) specifically catalyses the hydrolysis of acetylcholine and is subjected to marked inhibition by its own natural substrate. In contrast, butyrylcholinesterase (BChE; EC 3.1.1.8) is capable of degrading a wider range of choline esters and is not inhibited by its substrate [1, 2]. AChE is selectively inhibited by BW 284c51, while BChE is specifically inhibited by tetraisopropylpyrophosphoramide (iso-OMPA) [3]. AChE is widely distributed in the nervous system and its role in rapidly terminating nerve impulse by hydrolysing acetylcholine in cholinergic synapses is well documented [1]. BChE is produced in the liver and enriched in the circulation. In addition, it is also present in adipose tissue, intestine, smooth muscle cells, white matter of the brain and many other tissues [4]. The exact physiological function of BChE is still elusive. It is generally viewed as a back up for the homologous AChE and to act as a scavenger for anticholinesterase compounds [5].
The presence of ChEs in tissues that are not cholinergically innervated provides the most compelling evidence for the view that AChE and BChE may have functions, other than the termination of cholinergic neurotransmission. There is considerable body of evidence to suggest that ChEs may be involved in embryonic neural development, including a role in cell proliferation, differentiation and cell adhesion [6, 7]. ChEs may also have a causative/permissive role in various pathological conditions as exemplified by the overexpression of ChE genes in various types of tumours and presence of abnormal levels of ChEs with altered properties in Alzheimer's disease [8, 9]. The histochemical staining of esterase activity on ChE developed by Koelle and Friedenwald [10] and modified by Karnovsky and Roots [11] has been extensively used to elucidate the functions of ChEs, examine their tissue specificity, developmental alterations and pathological changes from many species [2].
Other than the predominant choline esterase activity, ChEs also display a genuine aryl acylamidase (AAA) activity capable of hydrolysing the synthetic substrate o-nitroacetanilide into o-nitroaniline and acetate [12]. Apart from being strongly inhibited by choline esters and classical ChE inhibitors, this AAA activity is susceptible to selective inhibition by 5-hydroxytryptamine (serotonin) [12, 13, 14]. The characteristic feature of the AAA activity associated with human serum BChE is its several fold activation by tyramine [12]. The natural substrate or the precise physiological role of the AAA activity on ChEs is not known.
Studies on the tissue specific and developmental regulations/alterations of AAA on ChEs have not been attempted so far partly due to the lower specific activity of the AAA activity on ChEs and mainly due to the absence of a specific method to visualise this activity. In the present paper, a method to visualise the AAA activity on ChEs in polyacrylamide gels using human serum BChE and electric eel AChE as models is described.
Results and discussion
The principle of the staining procedure is depicted in Figure 1. The AAA on ChEs acts on ONAA to release acetate and o-nitroaniline. o-nitroaniline then reacts with nitrous acid (provided by the reaction of sodium nitrite with HCl) at about 0°C yielding the corresponding diazonium compound. The reaction mixture must be kept very cold during the process, otherwise the diazonium compound may be partially hydrolysed to the corresponding phenol. The diazonium compound then complexes with NED to form purple coloured azo dye complex. This coupling reaction is an electrophilic substitution reaction [15].
Figure 1.
Scheme representing the principle of staining of the AAA actitvity.
The concentrations of sodium nitrite and HCl in the staining solution were found to be critical, since the generated nitrous acid is the one that reacts with o-nitroaniline to form the diazonium compound. For optimal detection of the AAA activity, a concentration of 0.1 % (w/v) sodium nitrite and 1 N HCl were found to be required. Similarly, studies with varying concentration of NED indicated that the maximum intensity of colour developed was with 0.75 % (w/v) of the colouring reagent.
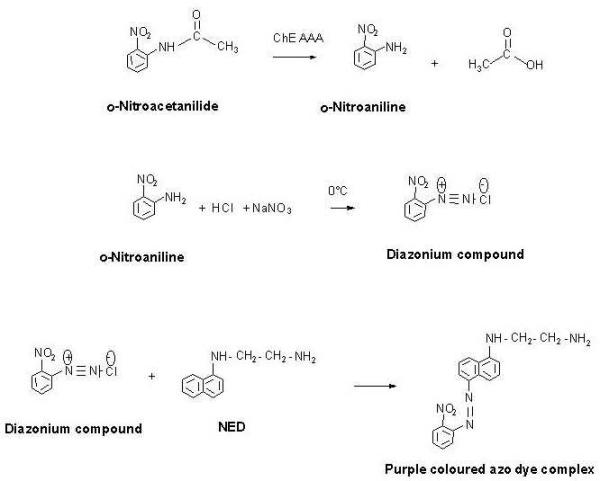
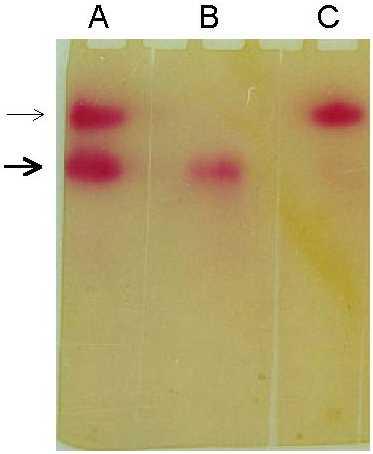
The stained gel pattern for AAA on AChE/BChE using the above protocol is shown in Figure 2. It has been shown that the AAA activities on both AChE and BChE can be visualised by this method. Further, selective staining for the AAA activities on either BChE or AChE in a sample containing both these enzymes has been demonstrated by the use of the specific inhibitors of BChE and AChE, namely, tetraisopropyl pyrophosphoramide (iso-OMPA) and 1,5-bis(4 Allyldimethylammoniumphenyl) pentan-3-one dibromide (BW 248c51), respectively. In addition, the intensity of staining increased linearly with the enzyme concentration (in terms of AAA activity) applied in the gel (Figure 3). From Figure 3, it is also clear that, to visualise a recognisable AAA activity band in the gel, at least 0.07 U of the enzyme is required.
Figure 2.
Differential staining for the AAA activities on ChEs from eel AChE and human serum BChE. Electrophoresis was performed on 3.5% native polyacrylamide gel slabs under non-denaturing conditions as detailed in 'Materials and methods'. All lanes were loaded with 1 U each of AAA on eel AChE and AAA on human serum BChE . After electrophoresis, 'A' was incubated in substrate solution without any inhibitor; 'B' was incubated in substrate solution containing 100 μM of iso-OMPA; and 'C' was incubated in substrate solution containing 100 μM of BW 284c51. After incubation for 40 min, the gels were stained for the AAA activity. The upper and lower arrows denote the AAA activity due to BChE and AChE, respectively.
Figure 3.
Effect of enzyme concentration (in terms of AAA activity) on the intensity of staining. Electrophoresis was performed on 3.5% native polyacrylamide gel as described under 'Materials and methods'. Lanes 1-5, respectively, corresponds to 0.07 U, 0.14 U, 0.21 U, 0.28 U and 0.35 U of AAA activity on BChE. After electrophoresis, the gel was stained for the AAA activity as given under 'Materials and Methods'.
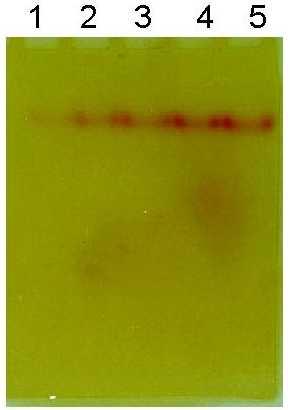
Prolonged incubation of the stained gels in the acid solution caused a rapid decrease in the intensity of the purple colour bands. Treating the gels with 0.3 M TCA for 30 min at 4°C prevents this loss in colour. The stain was further fixed in the gels by changing the pH to alkaline side with Tris-HCl buffer, pH 8.6. The colour of the bands changed from purple to brick red (Figure 4-A) upon changing the pH from acid to alkaline side. The gels can be stored for weeks in this alkaline buffer without loss in the intensity of the bands. Documentation of the stored gels, if necessary, can be done after treating the gels with 0.3 M TCA which brings back the original purple colour, but with a bluish tint (Figure 4-B).
Figure 4.
Effect of storage condition on the colour of the activity band developed using NED. After electrophoresis (of 1 U each of AAA on eel AChE and AAA on human serum BChE) under non-denaturing conditions, the gel was stained for AAA activity as described under 'Materials and methods'. The stained gel was then treated with 0.3 M TCA and stored in Tris-HCl Buffer, pH 8.6, where upon the colour of the band changes from purple to brick red (A). The characteristic purple colour, with a blue tint is regained by treating the gel once again with the TCA solution (B).
α-Naphthol and β-naphthol were also tried as colouring agents instead of NED. In alkaline conditions, α-naphthol produced a red coloured band, while β-naphthol produced an orange coloured band (Figure not given). When compared to the colour developed with NED, the colour developed with either of the naphthols was faint and diffused. Moreover, the fixing of the stain in the gels was also difficult.
The AAA activities, other than those associated with cholinesterases (like those found in human liver, rat serum, pineal gland) and amidases that utilise o-nitroacetanilide as substrate can be visualised by the method described above. This method would also allow detection of such activities in crude tissue extracts, however, a minimum of 0.07 U of the enzyme has to be loaded per well to clearly visualise the activity in gels, which is the limit of detectability of this method.
Conclusion
This study describes a novel method to visualise the AAA activity on ChEs in polyacrylamide gels. The method has been shown to be sensitive and also can selectively detect either of the ChE's AAA activity. Use of this method to visualise AAA activity in tissue sections, however, needs further refinement/modifications to enhance the sensitivity. This is because, any tissue, at a particular locus might not have AAA activity to the extent of 0.07 U. Nevertheless, this study is the first successful attempt to visualise the AAA activity on cholinesterase in vitro.
Materials and methods
Materials
N-(1-napthyl)ethylenediamine (NED) was procured from E.Merck, Darmstadt, Germany. iso-OMPA and BW 284c51 were from Sigma Chemical Co., St. Louis, USA. All other chemicals and reagents were of analytical grade and of the highest purity available. o-Nitroacetanilide was prepared as described in reference 16.
Enzymes
Human serum BChE was purified to apparent homogeneity as previously described [17]. Electric eel AChE (product No. C 3389) was procured from Sigma Chemical Co., St. Louis, USA.
The AAA activity on ChEs was assayed using o-nitroacetanilide (ONAA) as described in a previous report [16]. One unit of AAA activity is defined as the quantity of the enzyme required to liberate 1 μmole of o-nitroaniline in 1 hr under the standard assay conditions.
Staining procedure
Electrophoresis of ChEs were performed in 3.5% native polyacrylamide gel slabs under non-denaturing conditions according to Davis et al. [18]. After electrophoresis, the gels were stained for the AAA activity as follows. The gels were incubated in the substrate solution (10 mM ONAA in 0.1 M potassium phosphate buffer, pH 7.0) for 45 min at 40°C. The gels were transferred from the substrate solution to ice-cold staining solution of 1 N HCl containing 0.1% freshly prepared sodium nitrite and agitated briefly. Then 150 mg (final concentration 0.75% w/v) of N-(1-napthyl)ethylenediamine was added and the gels were gently agitated until sufficient intensity of the characteristic purple colour bands developed. The gels were then transferred to 0.3 M trichloroacetic acid (TCA) and kept for 30 min at 4°C. The gels were finally washed with 0.1 M Tris-HCl buffer, pH 8.6 and stored in the same buffer in a refrigerator. Differential staining for AChE AAA activity was performed by incubating the gels in the substrate solution containing 100 μM iso-OMPA to inhibit the AAA on BChE. Alternatively, in order to visualise BChE AAA activity, gels were incubated in substrate solution containing 100 μM BW 284c51 to inhibit the AAA on AChE.
Acknowledgements
The support by Defence Research & Development Establishment (DRDE), Gwalior, India (Project ID No. TC/05414/DRDE/Project-DRDE-149, dated 23 June, 1998) and the award of Senior Research Fellowship to L.J. by the Council of Scientific and Industrial Research, India, and DRDE are gratefully acknowledged.
Abbreviations
AAA, aryl acylamidase; AChE, acetylcholinesterase; BChE, butyrlycholinesterase; BW 284c51, 1,5-bis(4-Allyldimethylammoniumphenyl) pentan-3-one dibromide; ChE, cholinesterase; iso-OMPA, tetraisopropyl pyrophosphoramide; NED, N-(1-napthyl)ethylenediamine; ONAA, o-nitrooacetanilide; TCA, trichloroaceticacid.
Contributor Information
Lakshmanan Jaganathan, Email: jagscbe@yahoo.com.
Rathanam Boopathy, Email: boopathy@md4.vsnl.net.in.
References
- Quinn DM. Enzyme structure, reaction, dynamics, and virtual transition states. Chem Rev. 1987;87:955–979. [Google Scholar]
- Soreq H, Zakut H. Human Cholinesterases and Anticholinesterases. Academic Press. 1993.
- Austin L, Berry WK. Two selective inhibitors of cholinesterase. Biochem J. 1953;54:695–700. [PMC free article] [PubMed] [Google Scholar]
- Silver A. The Biology of Cholinesterases. Amsterdam: Elsevier. 1974.
- Schwarz M, Glick D, Loewensten Y, Soreq H. Engineering of human cholinesterases explains and predicts diverse consequences of administration of various drugs and poisons. Pharmacol Ther. 1995;67:283–322. doi: 10.1016/0163-7258(95)00019-D. [DOI] [PubMed] [Google Scholar]
- Layer PG, Willbold E. Cholinesterases in Avian Neurogenesis. Intel Rev Cytol. 1994;151:139–181. doi: 10.1016/s0074-7696(08)62632-7. [DOI] [PubMed] [Google Scholar]
- Layer PG, Ebert C, Treskatis S, Weikert T, Willbold E. Glycosylated inactive forms of chicken butyrylcholinesterasesand their possible functions. In Enzymes of the Cholinesterase Family, Edited by DM Quinn, AS Balasubramainan, BP Doctor and P Taylor New York: Plenum Press. 1995. pp. 427–434.
- Geula C, Mesulam MM. Cholinesterase and the Pathology of Alzheimer's Disease, Alzheimer's Disease & Related Diseases. 1995. pp. 23–28. [DOI] [PubMed]
- Soreq H, Lapidot-Lifson Y, Zakut H. A Role for Cholinesterases in Tumorigenesis? Cancer Cell. 1991;3:511–516. [PubMed] [Google Scholar]
- Koelle GB, Friedenwald JS. A histochemical method for localizing cholinesterase activity. Proc Soc Exp Biol Med. 1949;70:617–622. doi: 10.3181/00379727-70-17013. [DOI] [PubMed] [Google Scholar]
- Karnovsky MJ, Roots L. A "direct colouring" thiocholine method for cholinesterases. J Histochem Cytochem. 1964;12:219–221. doi: 10.1177/12.3.219. [DOI] [PubMed] [Google Scholar]
- Balasubramainan AS, Banumathy CD. Noncholinergic functions of cholinesterases. FASEB J. 1993;7:1354–1358. doi: 10.1096/fasebj.7.14.8224608. [DOI] [PubMed] [Google Scholar]
- Fujimoto D. Serotonin sensitive aryl acylamidase activity of acetylcholinesterase. FEBS Lett. 1976;71:121–123. doi: 10.1016/0014-5793(76)80912-X. [DOI] [PubMed] [Google Scholar]
- Weitnauer E, Robitizki A, Layer PG. Aryl acylamidase activity exhibited by butyrylcholinesterase is higher in chick than in horse, but much lower than in fetal calf serum. Neurosci Lett. 1998;254:153–156. doi: 10.1016/S0304-3940(98)00689-2. [DOI] [PubMed] [Google Scholar]
- Furniss BS, Hannaford AJ, Smith PWG, Tatchell AR. Vogel's Textbook of Practical Organic Chemistry. London: Addision Weseley Longman Limited. 1989.
- George ST, Balasubramanian AS. The Aryl Acylamidase and Their Relationship to Cholinesterases in Human Serum, Erythrocyte and Liver. Eur J Biochem. 1981;121:177–186. doi: 10.1111/j.1432-1033.1981.tb06447.x. [DOI] [PubMed] [Google Scholar]
- Boopathy R, Balasubramanian AS. Chemical modification of bifunctional human serum pseudocholinesterase: Effect on the pseudocholinesterase and aryl acylamidase activities. Eur J Biochem. 1985;151:351–360. doi: 10.1111/j.1432-1033.1985.tb09108.x. [DOI] [PubMed] [Google Scholar]
- Davis BJ. Disc electrophoresis. II. Method and application to human serum proteins. Ann NY Acad Sci. 1964;121:404–427. doi: 10.1111/j.1749-6632.1964.tb14213.x. [DOI] [PubMed] [Google Scholar]