

Abstract
We have demonstrated that the α-chemokine stromal-derived factor (SDF)-1-CXCR4 axis plays an important role in rhabdomyosarcoma (RMS) metastasis. With the recent descriptionof CXCR7, a new receptor for SDF-1 that also binds the interferon-inducible T-cell alpha chemoattractant (ITAC) chemokine, we became interested in the role of the CXCR7-SDF-1/ITAC axis in RMS progression. To address this issue, we evaluated 6 highly metastatic alveolar (A)RMS and 3 less metastatic embryonal (E)RMS cell lines and found that all these cell lines express CXCR7. While CXCR4 was expressed at a much higher level by highly metastatic ARMS lines, CXCR7 was present at a high level on ERMS lines. We also noticed that CXCR7 expression on RMS cells was downregulated in hypoxic conditions. More importantly, the CXCR7 receptor on RMS cell lines was functional after stimulation with ITAC and SDF-1 as evidenced by mitogen-activated protein kinase (MAPK)p42/44 and AKT phosphorylation as well as CXCR7 internalization, chemotaxis, cell motility, and adhesion assays. Similarly to CXCR4, signaling from activated CXCR7 was not associated with increased RMS proliferation or cell survival. Moreover, CXCR7+ RMS cells responded to SDF-1 and I-TAC in the presence of CXCR4 antagonists (T140, AMD3100). Furthermore, while intravenous injection of RMS cells with overexpressed CXCR7 resulted in increased seeding efficiency of tumor cells to bone marrow, CXCR7 downregulation showed the opposite effect. In conclusion, the CXCR7-SDF-1/ITAC axis is involved in the progression of RMS; targeting of the CXCR4-SDF-1 axis alone without simultaneous blockage of CXCR7 will be an inefficient strategy for inhibiting SDF-1-mediated pro-metastatic responses of RMS cells.
Keywords: Rhabdomyosarcoma, SDF-1, I-TAC, CXCR4, CXCR7
Introduction
Rhabdomyosarcoma (RMS) is the most common soft-tissue sarcoma of adolescence and childhood and accounts for 5% of all malignant tumors in patients under 15 years of age. Most tumors originate in the head and neck region, the urogenital tract, and the extremities. It is well known that RMS cells, particularly alveolar (A)RMS, can infiltrate the bone marrow (BM) and, because they can resemble hematologic blasts, may sometimes be misdiagnosed as acute leukemia cells. The “contamination” of BM by these cells may compromise its use for autologous transplantation.
There are two major histologic subtypes of RMS, i.e., the aforementioned ARMS and embryonal (E)RMS. Clinical evidence indicates that ARMS is more aggressive and has a significantly worse outcome than ERMS. Genetic characterization of RMS has identified markers that show excellent correlation with histologic subtype. Specifically, ARMS is characterized by the translocation t(2;13)(q35;q14) in 70% of cases or the variant t(1;13)(p36;q14) in a smaller percentage of cases. These translocations disrupt the paired box (PAX)3 and PAX7 genes on chromosome 2 and 1, respectively, and the forkhead in RMS (FKHR) gene on chromosome 13. As such, they generate PAX3-FKHR and PAX7-FKHR fusion genes. These fusion genes encode the fusion proteins PAX3-FKHR and PAX7-FKHR, which are believed to act in cell survival and dysregulation of the cell cycle in ARMS cells 1–3.
In our previous work, we demonstrated a pivotal role of α-chemokine stromal-derived factor-1 (SDF-1) – seven transmembrane span, G protein-coupled receptor CXCR4 axis in metastasis of RMS to various organs including BM 4–5. For many years, it was postulated that CXCR4 was the only receptor for SDF-1 6–8. However, the concept of an exclusive interaction of SDF-1 with CXCR4 was questioned recently after observing murine fetal liver cells from CXCR4−/− mice still bind SDF-1 and that there were some inconsistencies between CXCR4 expression and SDF-1 binding on tumor-established cell lines 9. In addition, another chemokine called interferon-inducible T-cell alpha chemoattractant (I-TAC) was shown to partially block SDF-1 binding without interacting directly with the CXCR4 receptor. All of this suggested a presence of another SDF-1-binding receptor on the cell surface and the search for such a receptor began. This receptor was recently identified and named CXCR7 9.
After our preliminary studies revealed that human RMS cells express CXCR7, we became interested in a potential role of the SDF-1-CXCR7 axis in RMS growth and metastasis. Thus, we focused on the biological responses of CXCR7-positive ARMS and ERMS cell lines to stimulation by exogenous SDF-1 and I-TAC, such as phosphorylation of signaling proteins, proliferation, survival, adhesion, expression of matrix metalloproteinases (MMPs), chemotaxis, and chemoinvasion. We also overexpressed CXCR7 or downregulated its expression on selected RMS cell lines. Finally, by employing a xenotransplant model in vivo, we evaluated a role for CXCR7 in expanding human RMS cells inoculated into immunodeficient mice.
Our findings imply that human RMS expresses the functional CXCR7 receptor. We also identified overlapping and distinct effects of CXCR4-SDF-1 and CXCR7-SDF-1/ITAC axes in regulating metastatic behavior of RMS cells.
Material and Methods
Cell lines
We used human RMS cell lines (gift of Dr. Peter Houghton, St. Jude Children’s Research Hospital, Memphis, TN) comprising ARMS lines (RH2, RH5, RH28, RH30, and CW9019)and ERMS lines (RH18, RD, and SMS-CTR). RMS cells used for experiments were cultured in Roswell Park Memorial Institute medium (RPMI) 1640 (Sigma, St. Louis, MO), supplemented with 100 IU/ml penicillin, 10 μg/ml streptomycin, and 50 μg/ml neomycin (Life Technologies, Inc., Grand Island, NY) in the presence of 10% heat-inactivated fetal bovine serum (FBS; Life Technologies). The ERMS cell line, RD, transfected with the PAX3-FKHR gene (kind gift from Dr. Frederic G. Barr, Univ. of Pennsylvania, Philadelphia, PA), was cultured in the presence of the selective agent geneticin (G-418) as described 1, 10. The cells were cultured in a humidified atmosphere at 37°C in 5% CO2 at an initial cell density of 2.5 × 104 cells/flask (Corning, Cambridge, MA) and the media were changed every 48 hours.
Fluorescence-activated cell sorting (FACS) analysis
The expression of CXCR4 and CXCR7 on RMS cell lines was evaluated by FACS analysis as previously described 4, 11. The CXCR4 antigen was detected with allophycocyanin (APC)-anti-CXCR4 monoclonal antibody (BD Pharmingen, San Diego, CA), clone no. 12.G5. CXCR7 antigen was detected with phycoerythrin (PE)-anti-CXCR7 monoclonal antibody (MoAb; R&D Systems, Minneapolis, MN) clone # 11.G8. Isotype-matched APC and PE-conjugated immunoglobulin (Ig) served as controls (BD Biosciences). Briefly, the cells were stained in phosphate-buffered saline (PBS; Ca- and Mg-free) supplemented with 2% bovine calf serum (BCS; Hyclone, Logan, UT). After the final wash, cells were re-suspended in PBS and analyzed by FACS using the LSRII (Becton Dickinson, San Jose, CA).
Real-time quantitative reverse transcription PCR (RQ-PCR)
Total RNA was isolated from cells treated with hypoxia and controls with RNeasy Kit (Qiagen, Valencia, CA). The RNA was reverse-transcribed with MultiScribe reverse transcriptase and oligo dt primers (Applied Biosystems, Foster City, CA). Quantitative assessment of mRNA levels was performed by real-time RT-PCR on ABI 7500 instrument and Power SyBR Green PCR Master Mix reagent. Real-time conditions were as follows: 95°C (15 sec), 40 cycles − 95°C (15 sec) 60°C (1 min). According to a melting point analysis only one PCR product was amplified under these conditions. The relative quantization value of a target, normalized to the endogenous control β-2 microglobulin gene and relative to a calibrator, is expressed as 2-ΔΔCt (-fold difference), where ΔCt = (Ct of target genes) − (Ct of endogenous control gene, β-2 microglobulin), and ΔΔCt = (ΔCt of samples for target gene) − (ΔCt of calibrator for the target gene). The following primer pairs were used:
CXCR4 forward 5- GGTTCCTTCATGGAGTCATAGTC -3,
CXCR4 reverse 5- CGGTTACCATGGAGGGGATC -3,
CXCR7 forward 5- GGCTATGACACGCACTGCTACA -3,
CXCR7 reverse 5- TGGTTGTGCTGCACGAGACT -3,
CXCR3 forward 5 – TGCCAATACAACTTCCCACA -3,
CXCR3 reverse 5 – CGGAACTTGACCCCTACAAA – 3.
Phosphorylation of intracellular pathway proteins
Western blots were performed on extracts prepared from RMS cell lines (2 × 106 cells) that were kept in RPMI medium containing low levels of bovine serum albumin (BSA; 0.5%) to render the cells quiescent. The cells were divided and stimulated with optimal doses of SDF-1 (300 ng/mL) or I-TAC (100 ng/mL) for 5 minutes at 37°C and then lysed (for 10 minutes) on ice in M-Per lysing buffer (Pierce, Rockford, IL) containing protease and phosphatase inhibitors (Sigma). Subsequently, the extracted proteins were separated on either a 12% or 15% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and the fractionated proteins were transferred to a nitrocellulose membrane (Schleicher & Schuell, Keene, NH) as previously described 12. Phosphorylation of the intracellular kinases, 44/42 mitogen-activated protein kinase (MAPK) (Thr 202/Tyr 204) and AKT, was detected using commercial mouse phosphospecific MoAb (p44/42) or rabbit phosphospecific polyclonal antibodies for each of the remainder (all from New England Biolabs, Beverly, MA) with horseradish peroxidase (HRP)-conjugated goat antimouse immunoglobulin (Ig)G or goat antirabbit IgG as a secondary antibody (Santa Cruz Biotech, Santa Cruz, CA) as described (4). Equal loading in the lanes was evaluated by stripping the blots and reprobing with appropriate MoAbs: p42/44 anti-MAPK antibody clone no. 9102 and anti-AKT antibody clone no. 9272 (Santa Cruz Biotech). The membranes were developed with an enhanced chemiluminescence (ECL) reagent (Amersham Life Sciences, Little Chalfont, United Kingdom), dried, and subsequently exposed to film (HyperFilm; Amersham LifeSciences).
Chemotaxis assay
The 8-μm pore polycarbonate membranes were covered with 50 μL of 0.5% gelatin. Cells were detached [with 0.5 mmol/L ethylendiaminetetraacetic acid (EDTA)], washed in RPMI 1640, resuspended in RPMI 1640 with 0.5% BSA, and seeded at a density of 3 × 104 in 120 μL into the upper chambers of Transwell inserts (Costar Transwell; Corning Costar, Corning, NY). The lower chambers were filled with SDF (300 ng/mL), I-TAC (100 ng/mL), or 0.5% BSA RPMI 1640 (control). After 24 hours, the inserts were removed from the Transwells. Cells remaining in the upper chambers were scraped off with cotton wool and cells that had transmigrated were stained by HEMA3 (Protocol, Fisher Scientific, Pittsburgh, PA) and counted either on the lower side of the membranes or on the bottom of the Transwells. In come of the experiments cells were exposed to T140 (1 μm/ml) or AMD3100 (50 μm/ml).
Time-lapse monitoring of the locomotion of individual cells
The images of human RMS migrating on plastic at 37°C were evaluated with an inverted microscope using phase contrast optics. Analysis of cell migration began 18 hours after cell seeding. The locomotion images were recorded with a charge-coupled device (CCD) camera. RD, RH18, RH28, and RH30 cells were plated to Corning flasks at a density of 104 cells/cm2 and were mock-treated or prestimulated by I-TAC (100 ng/mL) 30 minutes before recording. The cell trajectories were constructed from 60 subsequent cell centroid positions recorded for 240 minutes at 5-minute time intervals. The cell trajectories were presented in circular diagrams 13 and the length of cell tracks was calculated in addition to the final displacement. Cell tracks were recorded under the conditions described earlier for each cell line.
The following parameters characterizing cell locomotion were computed for each cell using procedures written in the Mathematica language, including: (1) total length of cell trajectory (in micrometers); (2) the trajectory as a sequence of n straight-line segments, each corresponding to cell centroid translocation within one time interval between two successive images; (3) total length of the final displacement of the cell from the starting point to the final position, i.e., distance between the first and last points of the cell track (in micrometers); (4) average speed of cell locomotion defined as total length of cell trajectory/time of recording; and (5) the ratio of cell displacement length to cell trajectory length, called the coefficient of movement efficiency (CME) 14, 15.
Adhesion of RMS cells to fibronectin
Cells were made quiescent for 24 hours with 0.5% BSA in RPMI before incubation with SDF-1 (300 ng/mL) or I-TAC (100 ng/mL) for 5 minutes. Cells were added directly onto the protein-coated wells (5 × 104/well) for 5 minutes. The wells were coated with fibronectin (10 μg/mL) overnight at 4°C and blocked with BSA for 2 hours before the experiment. Following incubation at 37°C, the plates were vigorously washed 3 times and adherent cells were stained by HEMA 3 (Protocol, Fisher Scientific) and counted under microscope. In some of the experiments cells were exposed to T140 (1 μm/ml) or AMD3100 (50 μm/ml).
Adhesion to HUVECs
RMS cells were labeled before assay with the fluorescent dye calcein-AM and were subsequently stimulated with SDF-1 (300 ng/mL) or I-TAC (100 ng/mL) for 5 minutes and added (for 5 minutes) to the 96-well plates covered by HUVECs. After the non-adherent cells had been discarded, cells that adhered to the HUVECs were counted under fluorescent microscope 16.
Cell proliferation
Cells were plated in culture flasks at an initial density of 104 cells/cm2 in the presence or absence of SDF-1 (300 ng/mL) or I-TAC (100 ng/mL). The cell number was calculated at 12, 24, 36, 48, 60, and 72 hours after culture initiation. In some experiments, the cultures were prolonged upto 7 days. At the indicated time points, cells were harvested from the culture flasks by trypsinization and the number of cells was determined using a Bürker hemocytometer (Buffalo, NY) as described elsewhere 17.
Analysis of MMPs and tissue inhibitor of matrix metalloproteinases (TIMPs)
RMS cell lines were incubated in RPMI with 0.5% of BSA (2–4×106 cells/mL) at 37°C in 5% CO2 for 24 hours with SDF-1 (300 ng/mL) or I-TAC (100 ng/mL) or without reagents. After incubation, cell-conditioned media were collected for zymographic analysis of MMP (-2, -3, -9, and -14) and TIMP (-1 and -2) activities. Cell pellets were used for total RNA isolation and reverse transcriptase-polymerase chain reaction (RT-PCR) analysis as described (18). Zymography was carried out using 12% polyacrylamide gel copolymerized with 1.5 mg/mL gelatin (Sigma; http://www.sigma-aldrich.com) and clear bands at 92kDa and 72kDa against a Coomassie blue background indicated the presence of MMP-9 and -2 and TIMP-1 and -2, respectively. Total cellular RNA was extracted and the conversion of mRNA to cDNA was carried out using RT Reagents (Applied Biosystems, Foster City, CA). Sequences for human MMP-1, -2, -3, -9, and -14 were obtained from GenBank (Los Alamos, NM) and used to design primer pairs. Detection of human (h)MMP-2, -3, -9, and -14 levels was performed using real-time PCR and an ABI Prism 7500 Sequence Detection System. The primers used were as follows:
| hMMP-1 | forward 5′TGCTCATGCTTTTCAACCAGG 3′ |
| reverse 5′TGAGCCGCAACACGATGTAA 3′ | |
| hMMP-2 | forward 5′TGGGACAAGAACCAGATCACATA3′ |
| reverse 5′CGAGCAAAGGCATCATCCA 3′ | |
| hMMP-3 | forward 5′TGTTTTGGCCCATGCCTATG 3′ |
| reverse 5′CCAATTTCATGAGCAGCAACG 3′ | |
| hMMP-9 | forward 5′AATCTCACCGACAGGCAGCT 3′ |
| reverse 3′TTTCGACTCTCCACGCATCTC 3′ | |
| hMMP-14 | forward 5′GCAGAAGTTTTACGGCTTGCAA 3′ |
| reverse 5′CGAACATTGGCCTTGATCTCAG 3′ | |
| TIMP-1 | forward 5′ TCGTGGCTCCCTGGAACA 3′ |
| reverse 5′CCAACAGTGTAGGTCTTGGTGAAG 3′ | |
| TIMP-2 | forward 5′CACCCAGAAGAAGAGCCTGAA 3′ |
| reverse 5′GGCAGCGCGTGATCTTG 3′ |
Actin polymerization
For the visualization of the actin cytoskeleton, cells were cultured for 12 hours on glass coverslips in RPMI 1640 supplemented with 0.5% BSA in the presence (100 ng/ml I-TAC or 300 ng/ml SDF-1) or absence of chemokines (control) Subsequently, cells were fixed in 3.7% paraformaldehyde/Ca- and Mg-free PBS for 15 minutes, permeabilized by 0.1% triton X-100 in PBS for 1 minute at room temperature (RT) and stained with Alexa Fluor488 - phalloidin at a concentration of 500 ng/ml for 1 hour 4. Cells were examined using a BX51 fluorescence microscope (Olympus America, Melville, NY) equipped with a CCD camera (Olympus America). Each staining was repeated three times for each cell line.
Internalization of CXCR4 and CXCR7
For determination of chemokine-induced receptor internalization, cells were incubated at 37°C in culture medium for 20 minutes in the presence or absence of SDF-1 or I-TAC (500 ng/mL). Prior to staining, cell-bound chemokine was removed by an ice-cold acidic glycine wash (50 mM glycine/HCl, pH 3, 100 mM NaCl) for 90 seconds 19. Cells were labeled with with APC-anti-CXCR4 monoclonal antibody (BD Pharmingen), clone no. 12.G5. CXCR7 antigen was detected with PE-anti-CXCR7 monoclonal antibody, clone no. 11G8 (MoAb; R&D Systems). Expression was measured by flow cytometry. Receptor internalization was evaluated by the mean channel fluorescence values.
Knockdown of CXCR7 with short hairpin (sh)RNA and over expression of CXCR7
In RNAi experiments, shRNA generating plasmid pRNA-U6.1/Neo (GenScript) was used. The oligonucleotide targeting base sequence for human CXCR7 was: 5′-GACCACAGGCTATGACACG -3′. RMS cells were plated at 80% confluency and transfected with shRNA vector using Lipofectamine (Invitrogen, Carlsbad, CA) according to manufacturer’s protocol. Commercially available negative scrambled control plasmid was used (Dharmacon, Lafayette, CO). The CXCR7 overexpressing cell line was made by transfecting RH30 cells with pcDNA3.1-CXCR7 plasmid (kind gift from Dr. James R. Lillard) and empty vector as a control. For stable transfection of shRNA-producing vector or CXCR7 overexpression, single cell dilutions were prepared and the cells were further expanded in the presence of G418 (Cellgro, Manassas, VA).
Transplants of RMS cells into immunodeficient mice
To evaluate the in vivo metastatic behavior of three populations of RH30 cell lines (RH30, RH30 EpV, and RH30 with over expression of CXCR7) and three populations of RH18 (RH18, RH18 scrambled, and RH18 with knockdown of CXCR7), cells were injected intravenously (i.v.; 8 × 106 per mouse) into severe combined immunodeficient (SCID)-Beige inbred mice. Marrows, livers, and lungs were removed 48 hours after injection of these cells and the presence of RMS cells (i.e., murine-human chimerism) was evaluated as the difference in the level of human alpha-satellite expression. DNA was amplified in the extracts isolated from BM-, liver- and lung-derived cells using real-time PCR. Briefly, DNA was isolated using the QIAamp DNA Mini kit (Qiagen, Valencia, CA). Detection of human satellite and murine β-actin DNA levels was conducted using real-time PCR and an ABI Prism 7500 Sequence Detection System. A 25-μL reaction mixture containing 12.5 μL SYBR Green PCR Master Mix, 300 ng DNA template, 5′-ACC ACT CTG TGT CCT TCG TTG G-3′ forward, 5′-ATC GCG CTC TCA AAA GGA GTG T -3′, and 5′-AAA CGT CCA CTT GCA GAT TCT AG-3′ primers for the alpha-satellite, and 5′-GGA TGC AGA AGG AGA TCA CTG-3′ forward and 5′-CGA TCC ACA CGG AGT ACT TG-3′ reverse primers for the β-actin was used. Ct value was determined as before. The number of human cells present in the murine organs (degree of chimerism) was calculated from the standard curve obtained by mixing different numbers of human cells with a constant number of murine cells.
In some of the experiments, cells (5 × 106 per mouse) were inoculated into the hind limb muscles of SCID-Beige inbred mice. Six weeks later, the mice were sacrificed for evaluation of RMS cells present in blood, BM, liver, and lungs. Detection of human cells was performed as described above.
Statistical analysis
All results are presented as mean ± standard error of mean (SEM). Statistical analysis of the data was performed using the nonparametric Mann-Whitney test, with P<0.05 considered significant.
Results
CXCR7 is highly expressed in human RMS cell lines
We found that CXCR7 was expressed by FACS on all nine human RMS cell lines tested in this study (Fig. 1 panels A and B). As the RMS cell lines we investigated included both ARMS and ERMS types, we attempted to correlate the expression of CXCR4 on these cell lines with their histological phenotype (ARMS vs. ERMS). We observed that all three ERMS (RD, SMS-CTR, and RH18) cell lines stained highly positive for CXCR7 (>50% of cells; Fig. 1 panel B). In contrast, CXCR7 was expressed in ARMS cell lines at lower levels (Fig. 1 panel A). This is in striking difference to CXCR4 expression, which was highly expressed on several ARMS cell lines (RH5, RH28, and RH30) as shown in Fig. 1 panel A and Supplementary Figure 1 and 2.
Figure 1.

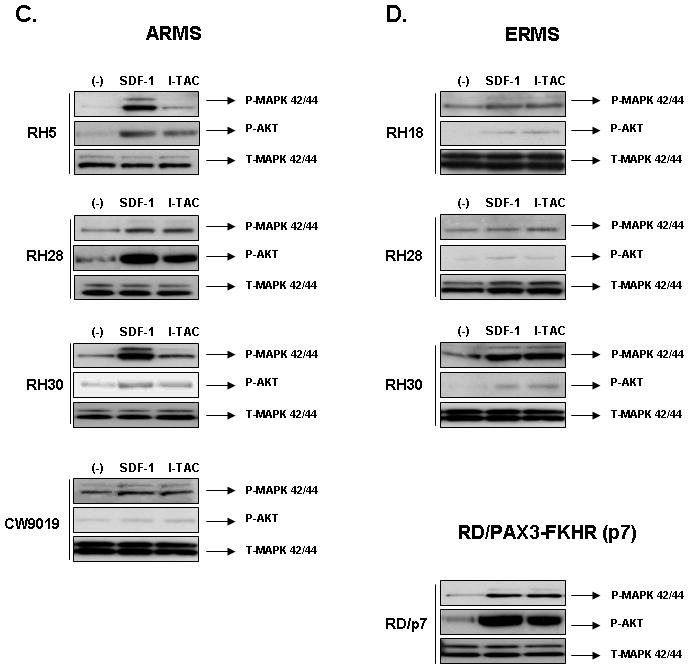
Panels A and B: Expression of CXCR4 and CXCR7 on human ARMS and ERMS cell lines. Flow cytometry was performed for detection of CXCR4 and CXCR7. The experiment was repeated three times with similar results. A representative study is shown. Panels C and D: SDF-1 and I-TAC activate intracellular signaling in human RMS cell lines. Phosphorylation of MAPK p42/44 and AKT in human RMS cell lines stimulated by SDF-1 (300 ng/mL for 5 min) and I-TAC (100 ng/mL for 5 min). The experiment was repeated three times with similar results. A representative study is shown.
To address if expression of CXCR7 receptor similarly as CXCR4 is hypoxia regulated, we evaluated CXCR7 expression in normoxic and hypoxic conditions. To our surprise, expression of CXCR7 was downregulated on RMS cells during exposure to hypoxia (Supplementary Figure 3). Accordingly, after 18 hours of culture in 1% O2, expression of CXCR7 decreased by ~ 40% on RH2 cells, ~50% on SMS-STR cells, and 55% on RD cells.
In our previous studies, we found that expression of CXCR4 correlates with t(2;13) and the presence of PAX3-FKHR fusion gene and ARMS phenotype (RH5, RH28, and RH30). Therefore, we sought to determine whether the PAX3-FKHR fusion gene regulates the expression of CXCR7. To address this issue, the ERMS cell line RD, which constitutively expresses a high level of CXCR7 and a low level of CXCR4, was stably-transfected with an expression vector containing the PAX3-FKHR fusion gene cDNA. We found that RD/PAX3-FKHR cells (RD-p7) transfected with this fusion gene (Fig. 1 panel B) downregulated expression of CXCR7 from ~ 55% to 10% and, at the same time, upregulated expression of CXCR4 to ~ 60% (Fig. 1 panel B).
Thus, this part of our experiments shows a reciprocal expression of CXCR7 and CXCR4 receptors on ERMS vs. ARMS cell lines and, in addition, suggests negative involvement of hypoxia and PAX3-FKHR on CXCR7 expression.
CXCR7 is signaling receptor in human RMS cell lines
Because RMS cells express both CXCR7 and CXCR4 receptors, to evaluate whether CXCR7 is functional on RMS cells, we evaluated responses of these cells to stimulation by I-TAC. First, by employing RT-PCR (data not shown) as well as FACS analysis, we excluded the possibility that CXCR7+ RMS cell lines express another I-TAC binding receptor CXCR3 (Supplementary Figure 4). Second, Figure 1 panels C and D show that I-TAC induces phosphorylation of MAPK p42/44 and AKT in human RMS cell lines similarly to SDF-1. Furthermore, we noticed that CXCR7 may be efficiently internalized after stimulation by both SDF-1 and I-TAC (Supplementary Fig. 5).
Thus, in contrast to some initial reports 20, our data support the concept 21, 22 that CXCR7 is not a decoy but a functional seven-transmembrane span chemokine receptor. To validate these observations, we excluded the potential expression of another I-TAC binding receptor, CXCR3, on our RMS cells (Supplementary Figure 4).
I-TAC, similarly to SDF-1, does not influence proliferation and survival of RMS cell lines
We selected two ERMS cell lines (RD and SMS-CTR) and two ARMS cell lines (RH28 and RH30) that responded to I-TAC and SDF-1 stimulation by phosphorylation of MAPK p42/44 and AKT to determine whether their proliferation is affected by these chemokines. We stimulated them with these chemokines or not (control) under serum-free conditions. We found that during the 72 hours of stimulation the RMS cell lines proliferated intensively in both types of media (Supplementary Fig. 6). However, the kinetics of their proliferation were similarly not affected by the presence of I-TAC or, as we previously demonstrated 4 by the presence of SDF-1 in the culture.
Because the biology of various tumors may be regulated by the autocrine/paracrine axes, we considered a possibility that RMS cells express CXCR4 and CXCR7 ligands. Since a potential involvement of SDF-1-mediated autocrine axis was excluded in our previous studies 23, herein, we asked whether RMS cells express I-TAC that potentiality could interact with CXCR7 on RMS cell lines in an autocrine manner. However, we did not find expression of I-TAC mRNA in any RMS cell lines employed in our studies (data not shown).
I-TAC may induce locomotion and migration of RMS cells similarly to SDF-1
Next, we studied whether I-TAC influences the locomotion on polystyrene dishes of two selected human ERMS cell lines (RD and RH18) and two ARMS cell lines (RH28, and RH30) using time-lapse monitoring of the movement of individual cells. Figure 2A shows the trajectories of RMS cell migration in the absence (left panel) or presence (right panel) of I-TAC in the culture media. Analyses of these trajectories and mean values as well as standard errors for the parameters of cell locomotion are summarized in Table 1.
Figure 2. Effect of I-TAC on the motility and the adhesiveness of RMS cell lines.
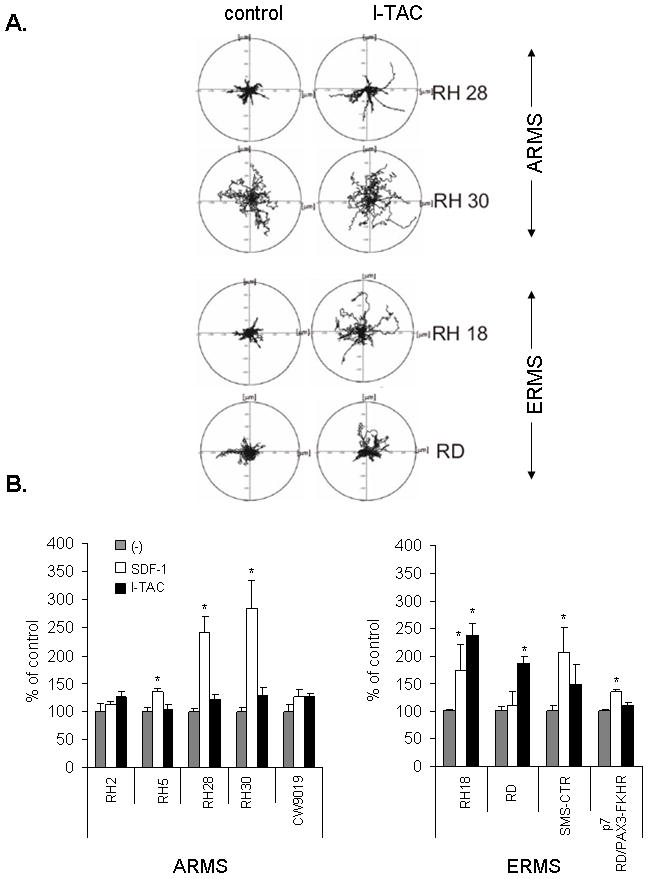
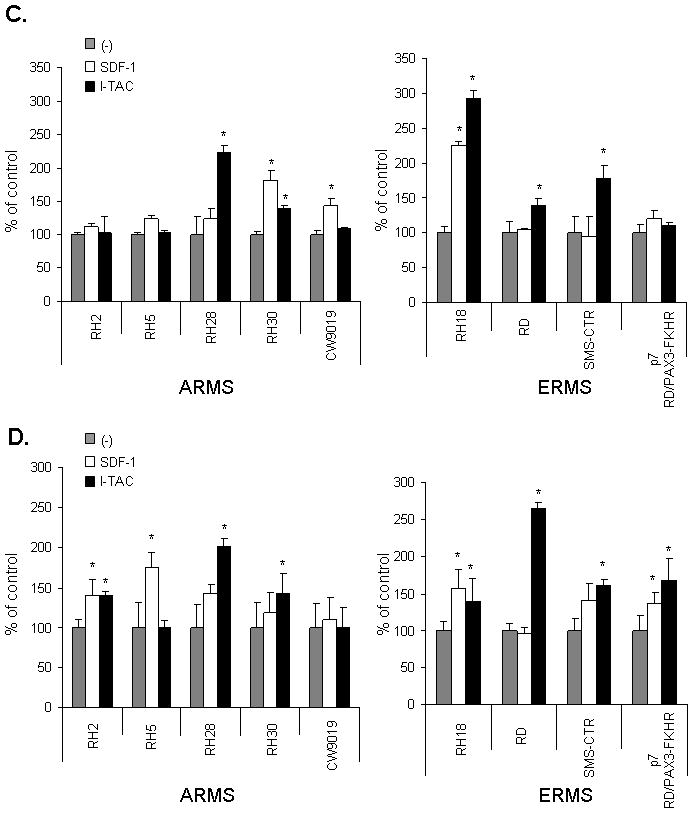
Panel A: The composite trajectories of RD, RH18, RH28, and RH30 cells migrating without (control) or with the addition of 100 ng/mL (I-TAC) are shown in circular diagrams drawn with the initial point of each trajectory at the origin of the plot. Panel B: Chemotaxis of RMS cells across Transwell membranes covered with gelatin to SDF-1 or I-TAC gradient. Gray bars show chemotaxis to control medium (no SDF-1 or I-TAC in upper and lower chambers), white bars show chemotaxis to SDF-1 (300 ng/mL) present in lower chamber, and black bars show chemotaxis to I-TAC (100 ng/mL) present in lower chamber. Data from 5 separate experiments are pooled together. * P<0,05. Panels C and D: Adhesion of human RMS cells to fibronectin(C) and to HUVECs (D). RMS were not stimulated (control; gray columns) or stimulated with SDF-1 (white columns) or I-TAC (black columns). Data from 4 separate experiments are pooled together. * p<0,05.
Table 1.
Parameters characterizing the movement of RMS RD, RH1, RH18, RH28, and RH30 cells moving in control medium in the presence of I-TAG (100 ng/ml).
| Cell line | TLcT | AScL | TLcD | AVcD | CME |
|---|---|---|---|---|---|
| RD | |||||
| control | 422.78 ± 102.62 | 1.76 ± 0.42 | 41.43 ± 26.34 | 0.17 ± 0.10 | 0.10 ± 0.06 |
| I-TAG | 383.43 ± 91.98 | 1.59 ± 0.38 | 39.06 ± 37.16 | 0.16 ± 0.15 | 0.12 ± 0.11 |
| RH18 | |||||
| control | 354.52 ± 59.51 | 1.47 ± 0.24 | 25.93 ±17.12 | 0.10 ± 0.07 | 0.07± 0.05 |
| I-TAG | 408.89 ± 97.94* | 1.71 ± 0.40* | 68.33 ± 20.34* | 0.28 ± 0.08* | 0.18 ± 0.14* |
| RH28 | |||||
| control | 177.45 ± 35.39 | 0.74 ± 0.14 | 33.76 ± 20.39 | 0.14 ± 0.08 | 0.19 ± 0.12 |
| I-TAG | 326.53 ± 81.51* | 1.36 ± 0.33* | 75.12 ± 40.83* | 0.31 ± 0.17* | 0.25 ± 0.16 |
| RH30 | |||||
| control | 370.06 ± 36.30 | 1.54 ± 0.15 | 85.78 ± 34.37 | 0.35 ± 0.14 | 0.23 ± 0.10 |
| I-TAG | 412.78 ± 83.87* | 1.72 ± 0.34* | 102.53 ± 40.07 | 0.43 ± 0.16 | 0.26 ± 0.12 |
TLcT - Total length of cell trajectory [μm].
AScM - Average speed of cell locomotion [μm/min] is defined as total length of cell trajectory/time of recording (240 min).
TLcD - Total length of cell displacement [μm].
AVcM- Average velocity of cell displacement [μm/min] is defined as length of final cell displacement/time of recording.
CME - Coefficient of movement efficiency is the ratio of cell displacement to cell trajectory length. CME would equal 1 for a cell moving persistently along one straight line in one direction and 0 for a random movement.
Note: Values are given as the mean ± SEM,
statistically significant at P<.05 (Mann-Whitney test).
The trajectories of RH18, RH28, and RH30 cells were significantly changed in the presence of I-TAC; analysis of individual cell trajectories showed that I-TAC stimulated motility of these cell lines (Fig. 2 panel A). Exposure of these cells to I-TAC caused a statistically significant increase of the average speed of locomotion (AScL) as well as the total length of cell displacement (TLcD) as compared with the control (Table 1). I-TAC most effectively stimulated motile activity of RH28 cells causing a 2-fold increase in the AScL and about a 2-fold increase in the TLcD. It is worth noting that because the migrating RMS cells made tortuous tracks (CME), their final displacement may be much smaller then the total length of their trajectories.
Next, we investigated directed migration of RMS cells to the I-TAC gradient by employing the Transwell system (Fig. 2 panel B). We observed that I-TAC chemoattracts several human RMS cell lines. We also observed difference in chemotactic responses to optimal doses of I-TAC and SDF-1. While RD cell line responded better to I-TAC, other lines (RH28, RH30, and SMS-CTR) responded better to SDF-1. Furthermore, with RH18, and SMS-CTR cells showing no or very low levels of CXCR4 expression, both I-TAC- and SDF-1-induced chemotaxis of these cells seems to be CXCR7-mediated.
Furthermore, RD cell line, for which locomotion activity on plastic dishes was not induced after the addition of I-TAC (Fig. 2 panel A), showed relatively high directional chemotaxis to this chemokine across Transwell membranes (Fig. 2 panel B).
Because both I-TAC and SDF-1 may increase chemotaxis, we decided to investigate whether both factors regulate metastatic behavior in an additive or a synergistic manner. To address this issue, we exposed the RH30 ARMS cell line, which showed strong chemotaxis to SDF-1 alone (Fig. 2 panel B), to increasing doses of I-TAC (0–100 ng/ml) in the presence of a constant suboptimal dose (100 ng/ml) of SDF-1. We observed that both factors increased the chemotactic activity of RH30 cells in an incremental manner (Supplementary Fig. 7).
Thus, simultaneous expression of SDF-1 and I-TAC increases the metastatic potential of RMS cells by engaging both CXCR7 and CXCR4 receptors.
Similarly to SDF-1, I-TAC increases adhesion to fibronectin and HUVECs
Subsequently, we decided to investigate whether CXCR7 receptor regulates expression/activation of integrins on human RMS. Using FACS analysis, we did not find any change in the level of expression of very late antigen (VLA)-4, VLA-5, or intercellular adhesion molecule (ICAM)-1 on RMS cells after incubation with I-TAC for 24 hours (data not shown). However, we observed that I-TAC affected the adhesion of several RMS cell lines to fibronectin and HUVECs (Fig. 2 panels C and D). As found using the Transwell system, with RH18, RD, and SMS-CTR cells showing very low levels of CXCR4 expression, both I-TAC- and SDF-1-induced chemotaxis of these cells seems to be CXCR7-mediated.
Furthermore, examination of the actin cytoskeleton organization by confocal microscopy revealed striking differences between the chemokine-treated and untreated cells. RMS cells growth in control medium displayed well-developed bundles of F-actin arranged in parallel to the long axis of the cell. Incubation of RD and RH18 cells with I-TAC for 12 hours induced a change in the organization of actin filaments and significantly increased both the number and thickness of F-actin bundles in the leading edge of migrating cells. (data not shown). In contrast to I-TAC, SDF-1 stimulation affected the actin cytoskeleton organization only in RH18 cells.
CXCR7 involvement in MMPs and TIMPs expression in RMS cell lines
Extensive experimental data show that upregulation of MMPs and downregulation of TIMPs actively contribute to cancer progression and metastasis and that relatively benign cells acquire malignant properties when MMP activity increases or TIMP activity diminishes (24). In this study, we evaluated expression of MMPs, which were reported to regulate RMS cell invasiveness 4 and observed whether SDF-1 and I-TAC stimulate their expression and/or have any effect on TIMP levels. We observed MMP-2, -3, -9, and -14 as well as TIMP-1 and TIMP-2 transcripts in all the RMS cell lines tested (Supplementary Table 1). Overall, both SDF-1 and I-TAC increased, depending on the cell line, to different degrees of expression of MMPs and, at the same time, downregulated expression of TIMPs.
Blockade of CXCR4 receptor by T140 or AMD3100 does not affect chemotaxis and adhesion to SDF-1 or ITAC of CXCR7+ cell lines
Finally, we employed T140 and AMD3100, which are specific blocking agents for CXCR4, in an experiment to determine whether these molecules affect the motility of CXCR7+ RMS cells. We employed two RMS cell lines, RH28, which expresses both CXCR7 and CXCR4, and RH18, which expresses CXCR7 only. As expected, we noticed that chemotaxis of RH28 cells to SDF-1, but not I-TAC, was inhibited after exposure to CXCR4 antagonists (Fig. 3 panel A). A lack of complete inhibition of RH28 cells to SDF-1 suggests that these cells, despite CXCR4 blockade, can still respond to SDF-1, which engages the CXCR7 receptor. To support this further, exposure of CXCR7+ CXCR4− RH18 cells to T140 and AMD3100 did not affect these cells’ chemotactic responses to SDF-1 and I-TAC (Fig. 3 panel B) as well as adhesion to fibronectin (Fig. 3 panel C and D).
Figure 3. Effect of T140 and AMD3100 on SDF-1 and I-TAC -dependent chemotaxis and adhesion of RMS cells.

Representative study of chemotaxis for ARMS (panel A) cells and ERMS (panel B) to medium alone (control) or medium with SDF-1 (300 ng/ml) and I-TAC (100 ng/ml). Before chemotaxis, cells were pre-incubated 1 hr with T140 or AMD3100. Data from 3 separate experiments are pooled together. * p<0.0001. Representative study of adhesion to fibronectin of ARMS (panel A) cells and ERMS (panel B) in presence of SDF-1 (300 ng/ml) and I-TAC (100 ng/ml). Before adhesion, cells were pre-incubated 5 min with T140 or AMD3100. Data from 3 separate experiments are pooled together. * p<0.0001.
Thus, we provide evidence that blockage of SDF-1-mediated chemotaxis by small molecular inhibitors of CXCR4 are ineffective if cells express CXCR7, which is insensitive to T140 and AMD3100 blockage (Fig. 3).
Effect of CXCR7 overexpression or downregulation on SDF-1- and I-TAC-mediated responses
To better assess a role of CXCR7 in regulating metastatic behavior of RMS cells, we upregulated its expression from ~10% to 70% in RH30 cells (Fig. 4 panel A) or downregulated its level from ~ 55% to 12% on RH18 cells (Fig. 4 panel C). However, SDF-1 and I-TAC did not affect proliferation or survival of these cells (data not shown). We noticed that CXCR7-overexpressing RH30 cells better responded to I-TAC (Fig. 4 panel B) and, at the same time, the responsiveness of RH18 cells in which CXCR7 was diminished (Fig. 4 panel D) decreased their responsiveness to both I-TAC and SDF-1. The lack of increased response to SDF-1 in RH30 cells in which CXCR7 was upregulated could be explained by a very high basic expression level of CXCR4 (~100%) on these cells that probably mediates maximal responsiveness to the SDF-1 gradient. In contrast, in the case of RH18 cells that do not express CXCR4, downregulation of CXCR7 impacted on both SDF-1 and I-TAC responsiveness.
Figure 4. Effect of SDF-1 and I-TAC on RMS cells with over-expression and knockdown of CXCR7.
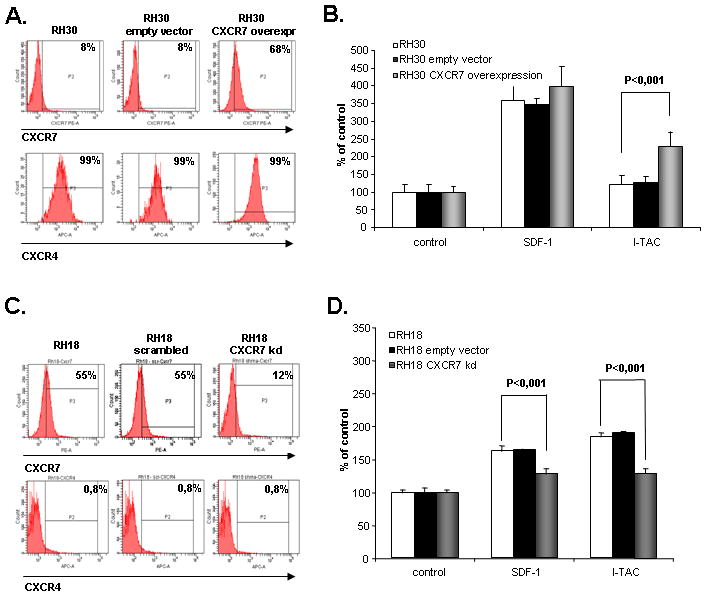
Panel A: shows expression of CXCR4 and CXCR7 on RH30 cells with over-expression of CXCR7. The experiment was repeated three times with similar results. A representative study is shown. Panel B shows chemotaxis of RH30, RH30 cells transfected with empty vector, and RH30 cells with over-expression of CXCR7 toward gradient of SDF-1 (300 ng/ml) and I-TAC (100 ng/ml). Data from 3 separate experiments are pooled together. Panel C shows expression of CXCR4 and CXCR7 on RH18 cells with knock-down of CXCR7. The experiment was repeated three times with similar results. A representative study is shown. Panel D shows chemotaxis of RH18, RH18 scrambled, and RH18 cells with knock-down of CXCR7 toward gradient of SDF-1 (300 ng/ml) and I-TAC (100 ng/ml). Data from 3 separate experiments are pooled together.
Next, because tumor growth depends on effective angiogenesis, we evaluated the influence of the SDF-1/I-TAC-CXCR7 axis on expression of selected pro-angiopoietic factors [interleukin (IL)-8, fibroblast growth factor (FGF)-2, vascular endothelial growth factor (VEGF), and macrophage inhibitory factor (MIF)]. We found that neither SDF-1 nor I-TAC was able to affect basic expression of these factors in wild type RH18 and RH30 RMS cells or cells in which expression of CXCR7 receptor was modulated (data not shown).
CXCR7 increases in vivo metastatic potential of RMS cells
To learn more about the role of the CXCR7 in the metastasis of RMS cells, we injected i.v. RMS cells in which expression of CXCR7 was upregulated or downregulated into SCID™-Beige inbred mice. After 48 hours, we evaluated the seeding efficiency of these human cells into murine BM, lungs, and liver (Supplementary Fig. 8 panel A). To estimate the number of human cells that seed to these organs in SCID™-Beige mice, we isolated DNA and amplified human α-satellite sequences and murine β-actin using real-time RT-PCR. The number of human cells present in murine organs was subsequently calculated from a standard curve derived from mixing varying numbers of human cells with a constant number of murine cells 25.
We found that RH30 cells in which CXCR7 was upregulated showed ~2 times higher seeding efficiency into the BM of SCID™-Beige mice compared to parental RH30 and RH30 empty vector transfected cells (Supplementary Fig. 8 panel A). At the same time, the seeding efficiency in lungs and liver was unaffected, which reflects a known BM-tropism of RMS cells. Next, the same cells (5×106/mouse) were injected intramuscularly into the hind limb. Six weeks later, we evaluated: i) primary tumor size and ii) the number of human RMS cells present in the BM, peripheral blood, lung, and liver of animals inoculated with RMS cells. To our surprise, we did not observe differences in primary tumor size growing after inoculation of control and CXCR7-overexpressing RH30 or control and CXCR7-downregulated RH18 cells (not shown). In contrast, six weeks after tumor inoculation, we noticed a significant increase of RMS cells in the BM of mice inoculated with CXCR7-overexpressing RH30 cells (Supplementary Fig. 8 panel B).
In a similar set of experiments, we noticed that RH18 cells in which CXCR7 was downregulated by shRNA showed significantly lower seeding efficiency into liver and lungs of SCID™-Beige mice compared to parental RH18 and RH18 empty vector transfected cells (Supplementary Figure 8 panel C). However, when we inoculated SCID™-Beige mice intramuscularly with the same cells (5×106/mouse) we did not observe differences in primary tumor size (not shown) but again, we noticed a significant increase of number of RMS cells in lungs and liver of these animals. Thus, downregulation of CXCR7 on RH18 cells, that constitutively do not express CXCR4 (Fig. 1 panel B) affects BM-tropism of these cells.
Effect of pertusis toxin (PTX) on CXCR7 signaling
Finally, to learn more about the controversial and potential involvement of Gαi in CXCR7 signaling 26 we exposed RMS cells to PTX, which is a known Gαi-specific inhibitor. In our experiments, we employed RH30 cells, which overexpress CXCR7 and thus respond robustly to SDF-1 and I-TAC (Fig. 4 panel A), and evaluated responsiveness of these cells to SDF-1 and I-TAC in a chemotaxis assay (Fig. 4 panel B), internalization of CXCR7 and CXCR4 receptors (Fig. 5 panel B), and phosphorylation of MAPK p42/44 and AKT (Fig. 5 panel C). We found that PTX inhibited SDF-1- and I-TAC-directed chemotaxis and phosphorylation of MAPK p42/44 and AKT (Fig. 5 panels A and C). However, it did not affect SDF-1- and I-TAC-induced internalization of CXCR7 (Fig. 5 panel B). Thus, some of the CXCR7 responses are Gαi-mediated and some are not.
Figure 5. Effect of PTX on CXCR7 signaling.
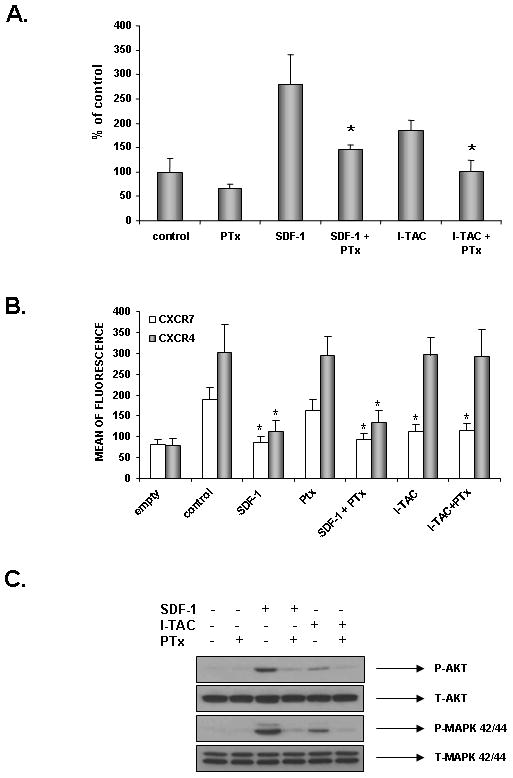
Panel A: Effect of PTX on chemotaxis of RH30 cells with over-expression of CXCR7 toward gradient of SDF-1 (300 ng/ml) and I-TAC (100 ng/ml). Data from 3 separate experiments are pooled together. * p<0.0001. Panel B: Effect of PTX on internalization of CXCR4 and CXCR7 induced by SDF-1 (1000 ng/ml) and I-TAC (500 ng/ml). Data from 3 separate experiments are pooled together. * p<0.0001. Panel C: Effect of PTX on phosphorylation of MAPK p42/44 and AKT induced in response to SDF-1 (300 ng/ml) and I-TAC (100 ng/ml). The experiment was repeated twice with similar results. A representative study is shown.
Discussion
RMS is the most common soft-tissue sarcoma of adolescents and children that frequently infiltrates the BM to the degree that often it mimics acute lymphoblastic leukemia 27. The prognosis is poor in particular for the more aggressive and metastatic ARMS 27. In addition to rapid tumor expansion, distant metastases and BM involvement are major problems for successful RMS therapy, making support of chemotherapy by new efficient anti-metastatic treatment strategies necessary. Our team is interested in molecular mechanisms regulating RMS metastasis. In one of our previous reports, we identified the SDF-1-CXCR4 4 and hepatocyte growth factor (HGF) – c-met receptor (23) axes to be involved in RMS spread. Our in vitro data were subsequently supported by clinical observations 28, 29.
Evidence has accumulated that the SDF-1-CXCR4 axis regulates metastasis of several tumors 30, 31. While tumor cells express CXCR4 receptor on the surface, SDF-1 is highly expressed in those tissue locations where CXCR4+ tumor cells frequently metastasize 32, 33. Based on this fact, several experimental strategies have been proposed to block CXCR4 receptor expression on tumor cells. Small CXCR4 locking agents such as bicyclam AMD3100 and T140 peptide have been proposed. However, with the identification of a new SDF-1 binding receptor, CXCR7, the potential targeting of CXCR4 receptor became more complicated. CXCR7 has already been shown to play an important role in metastasis of prostate 34, lung 35, 36, colon 37, breast 9, 36, 38, and brain tumor cells 39.
Herein, we describe that human RMS cells highly express CXCR7 and, in particular, this receptor was found to be highly expressed on the surface of ERMS cell lines. Our data also show that signaling through the CXCR7 receptor is not involved in RMS cell growth or cell survival. This is in agreement with our previous report showing that the SDF-1-CXCR4 axis, in contrast to other tumor types 40, does not involve RMS proliferation 4. Thus, in contrast to HGF-c-met receptor 23 and insulin growth factor (IGF)-1 and -2 IGF-1 receptor axes 41, 42 the chemokines – chemokine receptor axes are not directly involved in RMS cell growth.
However, based on our previously published 4, 5, 23 and new data, we postulate that chemokines mainly regulate pro-metastatic properties of RMS cells as evidenced herein by increasing RMS cell motility, directional chemotaxis, expression of MMPs, and cell adhesion. We also noticed some differences in CXCR4 and CXCR7 signaling. While CXCR4 activation is more important in increasing cell motility, CXCR7 seems to be more involved in increasing adhesiveness of RMS cells. This corresponds to our data that CXCR7 is highly expressed on less motile ERMS cells. Furthermore, we noticed that the SDF-1/I-TAC axis of CXCR7 is also important in expression of MMPs by RMS cells. However, in contrast to other tumors 43, CXCR4 and CXCR7 signaling are not involved in upregulation of pro-angiopoietic factors in these cells.
An intriguing observation demonstrated in this work is that CXCR7 expression becomes downregulated on RMS cells by hypoxia or overexpression of the ARMS-related fusion gene PAX3-FKHR. As we reported here, this explains lower expression of CXCR7 on ARMS cell lines and, in addition, has direct and important pro-metastatic implications. We envision that in hypoxic conditions, the downregulation of CXCR7 may enhance SDF-1 signaling through CXCR4 receptor and thus make these cells more motile. This fact may promote their egress from the primary tumor.
Our data presented from this work also add more information and resolve some controversies on the biological significance of CXCR7 receptor expression on tumor cells such as its ability to activate signal transduction pathways 26 or being internalized after stimulation 44. First, we clearly demonstrate that this receptor (at least in our RMS model) is functional and activates phosphorylation of MAPKp42/44 and serine-threonine kinase AKT. We also clearly show that activation of CXCR7 by SDF-1 or I-TAC increases internalization of the CXCR7 receptor.
In additional studies, we investigated whether some biological functions of CXCR7 (e.g., chemotaxis and activation of signaling pathways) are mediated by activation of Gai proteins. This latter issue is somewhat controversial in the literature 20, 45, 46. It is well known that chemokine receptors contain the so-called Asp-Arg-Tyr-Leu-Ala-Ile-Val (DRYLAIV) motif at the second intracellular loop, which is crucial for coupling a chemokine receptor to Gai-signaling proteins. However, in the case of CXCR7, this motif is slightly modified, which could potentially prevent its interaction with Gai-signaling proteins. We provide evidence here that Gai-signaling proteins are in fact involved in CXCR7 signaling as evidenced in our chemotaxis and signal transduction studies. However, at the same time, we observed a lack of Gai-signaling blockage by PTX on CXCR7 and CXCR4 internalization in response to stimulation by SDF-1 or I-TAC. This observation supports the idea that internalization of chemokine receptors may not always depend on Gai-signaling proteins 47. In our experiments the specificity of CXCR4 or CXCR7 activation on RMS cells was demonstrated by showing i) specific activation of CXCR7 receptor by I-TAC (RMS cells do not express another I-TAC binding receptor CXCR3 at mRNA level and protein level Supplementary Figure 4), ii) employing specific CXCR4 antagonists AMD3100 and T140 (Figure 3), iii) employing cell lines (RH2, RH18) that express CXCR7 but not CXCR7 and finally iv) by modulating level of expression of CXCR7 on RMS cells (Figure 4). Taking the latter into consideration it is important to mention that we did not observe that modulation of CXCR7 expression on our RMS cells affected, as reported for example in prostate cancer cells34, overall level of CXCR4.
Finally, we demonstrated that RMS cells that express the CXCR7 receptor on their surface may still respond to SDF-1 gradient after CXCR4 becomes blocked by CXCR4-specific blocking agents. We also show in an in vivo model that CXCR7 enhances metastasis of RMS cells and that these pro-metastatic properties correlate with the CXCR7 level on these cells. Based on this, we postulate that selective blockage of CXCR4 as a means to inhibit cancer metastasis may be ineffective for CXCR7+ tumors. To address this issue, we are planning to perform in vivo metastatic experiments where both receptors simultaneously will be blocked by CXCR7- or CXCR4-specific small molecular antagonists.
In conclusion, we demonstrated that CXCR7 is a new SDF-1-binding signaling receptor on human RMS cells. Furthermore, CXCR7 is more important in cell adhesion and becomes down-regulated during hypoxia, which may change the balance in SDF-1 signaling through both receptors (CXCR7 vs. CXCR4). As a result of this change, SDF-1 more robustly activates the CXCR4 receptor, which promotes chemotactic responses of RMS. As a biological consequence, this switch in SDF-1receptor signaling increases the metastatic potential of RMS by mobilizing tumor cells to egress from the tumor into the peripheral blood and lymph and migrate to new tissue locations. We also conclude that targeting of the CXCR4-SDF-1 axis alone without simultaneous blockage of CXCR7 will be an inefficient strategy to inhibit metastasis of RMS cells. This is currently being tested in our laboratories in immunodeficient mice inoculated with human RMS cells.
Supplementary Material
Binding assay was performed using labeled SDF-1 as CXCR4 and CXCR7 binding ligand. SDF-1 was labeled by employing Fluorokine Biotinylated Human SDF-1α kit (R&D Systems, Minneapolis, MN) according to the manufacturer protocol. Briefly, RMS cells were detached from culture dishes by employing non-enzymatic cell detachment solution - Cell Stripper (BD Becton Dickinson PharMingen, San Diego, CA) and subsequently, washed twice in PBS. Washed cells were re-suspended in PBS in final concentration of 4 × 106 cells/mL and stimulated with biotinylated SDF-1 (50 ng/ml) alone or with combination with SDF-1 (500 ng/ml) or I-TAC (500 ng/ml) for 60 minutes on ice. Receptor unbound ligands were washed out with cold PBS and bound biotynlabeled SDF-1 was detected with using avidin conjugated with FITC. Cells were analysed by flow cytometry. Panel A -Binding of biotinylated SDF-1 to RD cells that highly express CXCR7 was competed by using either unlabeled SDF-1 or unlabeled I-TAC. Panel B - Binding of biotinylated SDF-1 to wild type and scrambled transfected RH30 cells (that highly express CXCR4) was competed by SDF-1 but not I-TAC but the same binding in RH30 cells with over-expressed CXCR7 (RH30 CXCR7 OE) was effectively competed by both unlabeled SDF-1 or I-TAC. Data from four separate experiments are pooled together. * p<0.001
The mRNA expression was measured by Real-time PCR. Fold of difference was calculated on basis of 2ΔCt values, where CXCR4 expression in RD cells =1 and CXCR7 expression in CW9019=1.
Flow cytometry was performed for detection of CXCR7. The experiment was repeated three times with similar results. A representative study is shown
Flow cytometry was performed for detection of CXCR3. The experiment was repeated three times with similar results. A representative study is shown
RH30 cells with overexpression of CXCR7 were de-attached from plastic culture flasks with cell stripper, washed, and re-suspended in serum-free media. Cells were stimulated with SDF-1 (500 ng/mL), I-TAC (500 ng/mL), or a combination of both for 20 minutes in normal culture conditions. Prior to staining, cell-bound chemokine was removed by an ice-cold acidic glycine wash. Cells were labeled with APC-anti-CXCR4 and PE-anti-CXCR7 monoclonal antibodies. Expression was measured by flow cytometry. Receptor internalization was evaluated by the mean channel fluorescence values (shown as a bar graph).
ARMS cell lines (panel A): RH28, RH30 and ERMS (panel B): RD, SMS-CTR in the absence (control) or presence of SDF-1β (300 ng/ml) or I-TAC (300 ng/ml). Data from four independent experiments are demonstrated.
Chemotaxis of RH30 cells to increasing doses of SDF-1 (panel A), increasing I-TAC (panel B), and increasing doses of I-TAC alone or in the presence of a constant dose of SDF-1 (panel C). Data from three separate experiments are pooled together and means + SD are shown.
Panel A: RH30, RH30 empty vector, or RH30 cells with over-expression of CXCR7 were injected i.v. into SCID-Beige mice and organs were harvested after 48 hours for DNA isolation. The number of human cells in BM, lung, and liver was estimated by real-time RT-PCR per 106 murine cells. Real-time PCR data are pooled together from three independent experiments (five mice per experiment). * p< 0.001. Panel B: RH30, RH30 empty vector, or RH30 cells with over-expression of CXCR7 were inoculated into the hind limb muscles of SCID-Beige inbred mice. Four weeks later, mice were sacrificed and femora were harvested to evaluate the presence of metastatic cells in BM, peripheral blood, lung, and liver. The number of human cells was estimated by real-time RT-PCR per 106 murine cells. Real-time PCR data are pooled together from three independent experiments (five mice per experiment). * p< 0.001, ** p< 0.05. Panel C: RH18, RH18 scrambled and RH18 with downregulation of CXCR7 were injected i.v. into SCID-Beige mice and organs were harvested after 48 hours for DNA isolation. The number of human cells in BM, lung, and liver was estimated by real-time RT-PCR per 108 murine cells. Real-time PCR data are pooled together from three independent experiments (five mice per experiment). * p< 0.001.
* Data are pooled together from four independent experiments.
** RH30 cells with upregulated CXCR7
(−) = Lack of transcript
+ = Presence of transcript
++ = 3-fold increase and greater as compared to unstimulated cells
+++ = 10-fold increase and greater
d = decrease in expression
Acknowledgments
The authors are indebted to Dr. Peter Houghton, St. Jude Children’s Hospital, Memphis for critical comments. Supported by NIH grant R01 CA106281-01, NIH R01 DK074720, the Henry M. & Stella M. Hoenig Endowment and European Union structural funds, Innovative Economy Operational Program POIG 01.02-00-109/09 to MZR, R01 CA64202 and R01 CA104896 to FGB, and NIH Grant Number P20RR018733 from the National Center for Research Resources to MK.
References
- 1.Barr FG, Galili N, Holick J, Biegle JA, Rovera G, Emanuel BS. Rearrangement of the PAX3 paired box gene In the paediatric solid tumore alveolar rhabdomyosarcoma. Nat Genet. 1993;3:113–7. doi: 10.1038/ng0293-113. [DOI] [PubMed] [Google Scholar]
- 2.Davis RJ, Barr FG. Fusion genes resulting from alternative chromosomal translocations are overexpressed by gene –specific mechanisms in alveolar rhabdomyosarcoma. Proc Natl Acad Sci USA. 1997;94:8047–51. doi: 10.1073/pnas.94.15.8047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kelly KM, Womer RB, Barr FG. 3-FKHR and PAX7-FKHR fusions in rhabdomyosarcoma. J Pediatr Hematol Oncol. 1998;20:517–8. doi: 10.1097/00043426-199809000-00027. [DOI] [PubMed] [Google Scholar]
- 4.Labura J, Drukala J, Majka M, Tomescu O, Navenot JM, Kucia M, Marquez L, Peiper SC, Barr FG, Janowska-Wieczorek A, Ratajczak MZ. CXCR4-SDF-1 signaling is active in rhabdomyosarcoma cells and regulates locomotion, chemotaxis, and adhesion. Blood. 2002;100:2597–606. doi: 10.1182/blood-2002-01-0031. [DOI] [PubMed] [Google Scholar]
- 5.Kucia M, Reca R, Miekus K, Wanzeck J, Wojakowski W, Janowska-Wieczorek A, Ratajczak J, Ratajczak MZ. Trafficking of normal stem cells and metastasis of cancer stem cells involve similar mechanisms: pivotal role of the SDF-1-CXCR4 axis. Stem Cells. 2005;23:879–894. doi: 10.1634/stemcells.2004-0342. [DOI] [PubMed] [Google Scholar]
- 6.Nagasawa T, Hirota S, Tachibana K, Takakura N, Nishikawa S, Kitamura Y, Yoshida N, Kikutani H, Kishimoto T. Defects of B-cell lymphopoiesis and bone marrow myelopoiesis in mice lacking the CXC chemokine PBSF/SDF-1. Nature. 1996;382:635–638. doi: 10.1038/382635a0. [DOI] [PubMed] [Google Scholar]
- 7.Ma Q, Jones D, Springer TA. The chemokine receptor CXCR4 is required for the retention of B lineage and granulocytic precursors within the bone marrow microenvironment. Immunity. 1999;10:463–471. doi: 10.1016/s1074-7613(00)80046-1. [DOI] [PubMed] [Google Scholar]
- 8.Lazarini F, Tham TN, Casanowa P, Arenzana-Seisdedos F, Dubois-Dalcq M. Role of the alpha-chemokine stromal cell-derived factor (SDF-1) in the developing and mature central nervous system. Glia. 2003;42:139–148. doi: 10.1002/glia.10139. [DOI] [PubMed] [Google Scholar]
- 9.Burns JM, Summers BC, Wang Y, Melikian A, Berahovich R, Miao Z, Penfold ME, Sunshine MJ, Littman DR, Kuo CJ, Wei K, McMaster BE, Wright K, Howard MC, Schall TJ. A novel chemokine receptor for SDF-1 and I-TAC involved in cell survival, cell adhesion, and tumor development. J Exp Med. 2006;203:2201–2213. doi: 10.1084/jem.20052144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bennicelli JL, Advani S, Schafer BW, Barr FG. PAX3 and PAX7 exhibit conserved cis-acting transcription repression domains and utilize a common gain of function mechanism in alveolar rhabdomyosarcoma. Oncogene. 1999;18:4348–4356. doi: 10.1038/sj.onc.1202812. [DOI] [PubMed] [Google Scholar]
- 11.Majka M, Rozmyslowicz T, Honczrenko M, Ratajczak J, Wasik MA, Gaulton GN, Ratajczak MZ. Biological significance of the expression of HIV-related chemokine coreceptors (CCR5 and CXCR4) and their ligands by human hematopoietic cell lines. Leukemia. 2000;14:1821–1832. doi: 10.1038/sj.leu.2401891. [DOI] [PubMed] [Google Scholar]
- 12.Kijowski J, Baj M, Majka M, Reca R, Marquez LA, Christofidou-Solomidou M, Janowska-Wieczorek A, Ratajczak MZ. The SDF-1-CXCR4 axis stimulates VEGF secretion and activates integrines but does not affect proliferation and survival in lymphohematopoietic cells. Stem Cells. 2001;19:453–466. doi: 10.1634/stemcells.19-5-453. [DOI] [PubMed] [Google Scholar]
- 13.Gruler H, Nuccitelli R. Neural crest cell galvanotaxis: new data and a novel approach to the analysis of both galvanotaxis and chemotaxis. Cell Motil Cytoskeleton. 1991;19:121–33. doi: 10.1002/cm.970190207. [DOI] [PubMed] [Google Scholar]
- 14.Wójciak-Stothard B, Madeja Z, Korohoda W, Curtis A, Wilkinson C. Activation of macrophage-like cells by multiple grooved substrata. Topographical control of cell behaviour. Cell Biol Int. 1995;19:485–90. doi: 10.1006/cbir.1995.1092. [DOI] [PubMed] [Google Scholar]
- 15.Korohoda W, Madeja Z. Contact of sarcoma cells with aligned fibroblasts accelerates their displacement: computer-assisted analysis of tumor cell locomotion in co-culture. Biochem Cell Biol. 1997;75:263–276. [PubMed] [Google Scholar]
- 16.Clerck LS, Bridts CH, Mertens A, Moens MM, Stevens WJ. Use of fluorescent dyes in the determination of adherence of human leucocytes to endothelial cells and the effect of fluorochromes on cellular function. J Immunol Methods. 1994;172:115–124. doi: 10.1016/0022-1759(94)90384-0. [DOI] [PubMed] [Google Scholar]
- 17.Majka M, Rozmyslowicz T, Lee B, Murphy SL, Pietrzkowski Z, Gaulton GN, Silberstein L, Ratajczak MZ. Bone marrow CD34+ cells and megakaryoblasts secret β-chemokines: implications for infectability by M-tropic human immunodeficiency virus (R5 HIV) J Clin Invest. 1999;104:1739–1749. doi: 10.1172/JCI7779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Janowska-Wieczorek A, Marquez LA, Matsuzaki A, Hashmi HR, Larratt LM, Boshkov LM, Turner AR, Zhang MC, Edwards DR, Kossakowska AE. Expression of metalloproteinasees (MMP-2 and 9) and tissu inhibitors of metalloproteinases (TIMP1 and 2) in acute myelogenous leukaemia blasts:comparison with normal bone marrow cells. Br J Haematol. 1999;105:402–411. [PubMed] [Google Scholar]
- 19.Amara A, Gall SL, Schwartz O, Salamero J, Montes M, Loetscher P, Baggiolini M, Virelizier JL, Arenzana-Seisdedos F. HIV coreceptor downregulation as antiviral principle: SDF-1alpha-dependent internalization of the chemokine receptor CXCR4 contributes to inhibition of HIV replication. J Exp Med. 1997;186:139–46. doi: 10.1084/jem.186.1.139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Haraldsen G, Rot A. Coy decoy with a new ploy: intereceptor controls the levels of homeostatic chemokines. Eur J Immunol. 2006;36:1659–1661. doi: 10.1002/eji.200636327. [DOI] [PubMed] [Google Scholar]
- 21.Yoshida D, Nomura R, Teramoto A. Signaling Pathway Mediated by CXCR7, an Alternative Chemokine Receptor for Stromal-Cell Derived Factor-1alpha, in AtT20 mouse ACTH-secreting pituitary adenoma cells. J Neuroendocrinol. 2009 doi: 10.1111/j.1365-2826.2009.01867.x. [DOI] [PubMed] [Google Scholar]
- 22.Levoye A, Balabanian K, Baleux F, Bachelerie F, Lagane B. CXCR7 heterodimerizes with CXCR4 and regulates CXCL12-mediated G protein signalling. Blood. 2009;113:6085–93. doi: 10.1182/blood-2008-12-196618. [DOI] [PubMed] [Google Scholar]
- 23.Jankowski K, Kucia M, Wysoczynski M, Reca R, Zhao D, Trzyna E, Trent J, Peiper S, Zembala M, Ratajczak J, Houghton P, Janowska-Wieczorek A, Ratajczak MZ. Both hepatocyte growth factor (HGF) and stromal-derived factor-1 regulate the metastatic behavior of human rhabdomyosarcoma cells, but only HGF enhances their resistance to radiochemotherapy. Cancer Res. 2003;63:7926–35. [PubMed] [Google Scholar]
- 24.Stetler-Stevenson WG, Hewitt R, Corcoran M. Matrix metalloproteinases and tumor invasion: from correlation and causality to the clinic. Semin Cancer Biol. 1996;7:147–54. doi: 10.1006/scbi.1996.0020. [DOI] [PubMed] [Google Scholar]
- 25.Wysoczynski M, Miekus K, Jankowski K, Wanzeck J, Bertolone S, Janowska-Wieczorek A, Ratajczak J, Ratajczak MZ. Leukemia inhibitory factor: a newly identified metastatic factor in rhabdomyosarcomas. Cancer Res. 2007;67:2131–40. doi: 10.1158/0008-5472.CAN-06-1021. [DOI] [PubMed] [Google Scholar]
- 26.Thelen M, Thelen S. CXCR7, CXCR4 and CXCL12: an eccentric trio? J Neuroimmunol. 2008;198:9–13. doi: 10.1016/j.jneuroim.2008.04.020. [DOI] [PubMed] [Google Scholar]
- 27.Sandberg AA, Stone JF, Czarnecki L, Cohen JD. Hematologic masquerade of rhabdomyosarcoma. Am J Hematol. 2001;68:51–7. doi: 10.1002/ajh.1148. [DOI] [PubMed] [Google Scholar]
- 28.Scala S, Ottaiano A, Ascierto PA, Cavalli M, Simeone E, Giuliano P, Napolitano M, Franco R, Botti G, Castello G. Expression of CXCR4 predicts poor prognosis in patients with malignant melanoma. Clin Cancer Res. 2005;11:1835–41. doi: 10.1158/1078-0432.CCR-04-1887. [DOI] [PubMed] [Google Scholar]
- 29.Russell HV, Hicks J, Okcu MF, Nuchtern JG. CXCR4 expression in neuroblastoma primary tumors is associated with clinical presentation of bone and bone marrow metastases. J Pediatr Surg. 2004;39:1506–11. doi: 10.1016/j.jpedsurg.2004.06.019. [DOI] [PubMed] [Google Scholar]
- 30.Matsuo Y, Ochi N, Sawai H, Yasuda A, Sakamoto M, Takahashi H, Funahashi H, Takeyama H, Guha S. CXCL8/IL-8 and CXCL12/SDF-1alpha co-operatively promote invasiveness and angiogenesis in pancreatic cancer. Int J Cancer. 2009;124:853–61. doi: 10.1002/ijc.24040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gelmini S, Mangoni M, Serio M, Romagnani P, Lazzeri E. The critical role of SDF-1/CXCR4 axis in cancer and cancer stem cells metastasis. J Endocrinol Invest. 2008;31:809–19. doi: 10.1007/BF03349262. [DOI] [PubMed] [Google Scholar]
- 32.Yasuoka H, Tsujimoto M, Yoshidome K, Nakahara M, Kodama R, Sanke T, Nakamura Y. Cytoplasmic CXCR4 expression in breast cancer: induction by nitric oxide and correlation with lymph node metastasis and poor prognosis. BMC Cancer. 2008;8:340. doi: 10.1186/1471-2407-8-340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ratajczak MZ, Zuba-Surma E, Kucia M, Reca R, Wojakowski W, Ratajczak J. The pleiotropic effects of the SDF-1-CXCR4 axis in organogenesis, regeneration and tumorigenesis. Leukemia. 2006;20:1915–24. doi: 10.1038/sj.leu.2404357. [DOI] [PubMed] [Google Scholar]
- 34.Wang J, Shiozawa Y, Wang J, Wang Y, Jung Y, Pienta KJ, Mehra R, Loberg R, Taichman RS. The role of CXCR7/RDC1 as a chemokine receptor for CXCL12/SDF-1 in prostate cancer. J Biol Chem. 2008;15:4283–94. doi: 10.1074/jbc.M707465200. [DOI] [PubMed] [Google Scholar]
- 35.Goldmann T, Drömann D, Radtke J, Marwitz S, Lang DS, Schultz H, Vollmer E. CXCR7 transcription in human non-small cell lung cancer and tumor-free lung tissues; possible regulation upon chemotherapy. Virchows Arch. 2008;452:347–8. doi: 10.1007/s00428-008-0579-8. [DOI] [PubMed] [Google Scholar]
- 36.Miao Z, Luker KE, Summers BC, Berahovich R, Bhojani MS, Rehemtulla A, Kleer CG, Essner JJ, Nasevicius A, Luker GD, Howard MC, Schall TJ. CXCR7 (RDC1) promotes breast and lung tumor growth in vivo and is expressed on tumor-associated vasculature. Proc Natl Acad Sci U S A. 2007;104:15735–40. doi: 10.1073/pnas.0610444104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Meijer J, Ogink J, Roos E. Effect of the chemokine receptor CXCR7 on proliferation of carcinoma cells in vitro and in vivo. Br J Cancer. 99:1493–501. doi: 10.1038/sj.bjc.6604727. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 38.Müller A, Homey B, Soto H, Ge N, Catron D, Buchanan ME, McClanahan T, Murphy E, Yuan W, Wagner SN, Barrera JL, Mohar A, Verástegui E, Zlotnik A. Involvement of chemokine receptors in breast cancer metastasis. Nature. 2001;410:50–6. doi: 10.1038/35065016. [DOI] [PubMed] [Google Scholar]
- 39.Schönemeier B, Kolodziej A, Schulz S, Jacobs S, Hoellt V, Stumm R. Regional and cellular localization of the CXCl12/SDF-1 chemokine receptor CXCR7 in the developing and adult rat brain. J Comp Neurol. 2008;510:207–20. doi: 10.1002/cne.21780. [DOI] [PubMed] [Google Scholar]
- 40.Ehtesham M, Mapara KY, Stevenson CB, Thompson RC. CXCR4 mediates the proliferation of glioblastoma progenitor cells. Cancer Lett. 2008;274:305–12. doi: 10.1016/j.canlet.2008.09.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tsuchiya K, Hosoi H, Misawa-Furihata A, Houghton PJ, Sugimoto T. Insulin-like growth factor-I has different effects on myogenin induction and cell cycle progression in human alveolar and embryonal rhabdomyosarcoma cells. Int J Oncol. 2007;31:41–7. [PubMed] [Google Scholar]
- 42.Rikhof B, de Jong S, Suurmeijer AJ, Meijer C, van der Graaf WT. The insulin-like growth factor system and sarcomas. J Pathol. 2009;217:469–82. doi: 10.1002/path.2499. [DOI] [PubMed] [Google Scholar]
- 43.Otsuka S, Bebb G. The CXCR4/SDF-1 chemokine receptor axis: a new target therapeutic for non-small cell lung cancer. J Thorac Oncol. 2008;3:1379–83. doi: 10.1097/JTO.0b013e31818dda9d. [DOI] [PubMed] [Google Scholar]
- 44.Balabanian K, Lagane B, Infantino S, Chow KY, Harriague J, Moepps B, Arenzana-Seisdedos F, Thelen M, Bachelerie F. The chemokine SDF-1/CXCL12 binds to and signals through the orphan receptor RDC1 in T lymphocytes. J Biol Chem. 2005;280:35760–6. doi: 10.1074/jbc.M508234200. [DOI] [PubMed] [Google Scholar]
- 45.Boldajipour B, Mahabaleshwar H, Kardash E, Reichman-Fried M, Blaser H, Minina S, Wilson D, Xu Q, Raz E. Control of chemokine-guided cell migration by ligand sequestration. Cell. 2008;132:463–73. doi: 10.1016/j.cell.2007.12.034. [DOI] [PubMed] [Google Scholar]
- 46.Mellado M, Vila-Coro AJ, Martínez C, Rodríguez-Frade JM. Receptor dimerization: a key step in chemokine signaling. Cell Mol Biol. 2001;47:575–82. [PubMed] [Google Scholar]
- 47.Hartmann TN, Grabovsky V, Pasvolsky R, Shulman Z, Buss EC, Spiegel A, Nagler A, Lapidot T, Thelen M, Alon R. A crosstalk between intracellular CXCR7 and CXCR4 involved in rapid CXCL12-triggered integrin activation but not in chemokine-triggered motility of human T lymphocytes and CD34+ cells. J Leukoc Biol. 2008;84:1130–40. doi: 10.1189/jlb.0208088. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Binding assay was performed using labeled SDF-1 as CXCR4 and CXCR7 binding ligand. SDF-1 was labeled by employing Fluorokine Biotinylated Human SDF-1α kit (R&D Systems, Minneapolis, MN) according to the manufacturer protocol. Briefly, RMS cells were detached from culture dishes by employing non-enzymatic cell detachment solution - Cell Stripper (BD Becton Dickinson PharMingen, San Diego, CA) and subsequently, washed twice in PBS. Washed cells were re-suspended in PBS in final concentration of 4 × 106 cells/mL and stimulated with biotinylated SDF-1 (50 ng/ml) alone or with combination with SDF-1 (500 ng/ml) or I-TAC (500 ng/ml) for 60 minutes on ice. Receptor unbound ligands were washed out with cold PBS and bound biotynlabeled SDF-1 was detected with using avidin conjugated with FITC. Cells were analysed by flow cytometry. Panel A -Binding of biotinylated SDF-1 to RD cells that highly express CXCR7 was competed by using either unlabeled SDF-1 or unlabeled I-TAC. Panel B - Binding of biotinylated SDF-1 to wild type and scrambled transfected RH30 cells (that highly express CXCR4) was competed by SDF-1 but not I-TAC but the same binding in RH30 cells with over-expressed CXCR7 (RH30 CXCR7 OE) was effectively competed by both unlabeled SDF-1 or I-TAC. Data from four separate experiments are pooled together. * p<0.001
The mRNA expression was measured by Real-time PCR. Fold of difference was calculated on basis of 2ΔCt values, where CXCR4 expression in RD cells =1 and CXCR7 expression in CW9019=1.
Flow cytometry was performed for detection of CXCR7. The experiment was repeated three times with similar results. A representative study is shown
Flow cytometry was performed for detection of CXCR3. The experiment was repeated three times with similar results. A representative study is shown
RH30 cells with overexpression of CXCR7 were de-attached from plastic culture flasks with cell stripper, washed, and re-suspended in serum-free media. Cells were stimulated with SDF-1 (500 ng/mL), I-TAC (500 ng/mL), or a combination of both for 20 minutes in normal culture conditions. Prior to staining, cell-bound chemokine was removed by an ice-cold acidic glycine wash. Cells were labeled with APC-anti-CXCR4 and PE-anti-CXCR7 monoclonal antibodies. Expression was measured by flow cytometry. Receptor internalization was evaluated by the mean channel fluorescence values (shown as a bar graph).
ARMS cell lines (panel A): RH28, RH30 and ERMS (panel B): RD, SMS-CTR in the absence (control) or presence of SDF-1β (300 ng/ml) or I-TAC (300 ng/ml). Data from four independent experiments are demonstrated.
Chemotaxis of RH30 cells to increasing doses of SDF-1 (panel A), increasing I-TAC (panel B), and increasing doses of I-TAC alone or in the presence of a constant dose of SDF-1 (panel C). Data from three separate experiments are pooled together and means + SD are shown.
Panel A: RH30, RH30 empty vector, or RH30 cells with over-expression of CXCR7 were injected i.v. into SCID-Beige mice and organs were harvested after 48 hours for DNA isolation. The number of human cells in BM, lung, and liver was estimated by real-time RT-PCR per 106 murine cells. Real-time PCR data are pooled together from three independent experiments (five mice per experiment). * p< 0.001. Panel B: RH30, RH30 empty vector, or RH30 cells with over-expression of CXCR7 were inoculated into the hind limb muscles of SCID-Beige inbred mice. Four weeks later, mice were sacrificed and femora were harvested to evaluate the presence of metastatic cells in BM, peripheral blood, lung, and liver. The number of human cells was estimated by real-time RT-PCR per 106 murine cells. Real-time PCR data are pooled together from three independent experiments (five mice per experiment). * p< 0.001, ** p< 0.05. Panel C: RH18, RH18 scrambled and RH18 with downregulation of CXCR7 were injected i.v. into SCID-Beige mice and organs were harvested after 48 hours for DNA isolation. The number of human cells in BM, lung, and liver was estimated by real-time RT-PCR per 108 murine cells. Real-time PCR data are pooled together from three independent experiments (five mice per experiment). * p< 0.001.
* Data are pooled together from four independent experiments.
** RH30 cells with upregulated CXCR7
(−) = Lack of transcript
+ = Presence of transcript
++ = 3-fold increase and greater as compared to unstimulated cells
+++ = 10-fold increase and greater
d = decrease in expression