

It’s hard to imagine a class of neurons that has generated more excitement than mirror neurons (MN), cells discovered by Rizzolatti and colleagues in macaque area F5 that fire both during action execution and action observation[1]. We suggest, however, that the interpretation of MNs as supporting action understanding was a wrong turn at the start, and that a more appropriate interpretation was lying in wait with respect to sensorimotor learning. We argue: (i) Given their previous work, it would have been natural for Rizzolatti’s group to interpret MNs as involved in action selection rather than action understanding. (ii) They did not make this assumption because, at the time, the data suggested that monkey behavior did not support such an interpretation. (iii) Recent evidence shows that monkeys do, in fact, exhibit behaviors that support this alternative interpretation. (iv) Thus, the original basis for claiming that MNs mediate action understanding is no longer compelling. (v) There are independent arguments against the action understanding claim and in support of a sensorimotor learning origin for MNs. (vi) Therefore, the action understanding theory of MN function requires serious reconsideration if not abandonment.
MNs behave like non-mirroring sensorimotor cells
MNs were discovered in the context of research aimed at understanding how the visual properties of objects are integrated with motor codes for action. Cells in F5 were found to respond to visually presented objects as well as during grasping actions towards those objects. The interpretation of this circuit was that it coded a “vocabulary of motor acts and that this vocabulary can be accessed by … visual stimuli” (p. 491) [2] and that it was critical for “learning associations, including arbitrary associations between stimuli and [motor] schemas” (p. 317) [3]. This is a “’pragmatic’ mode of processing, the function of which is to extract parameters that are relevant to action, and to generate the corresponding motor commands” (p. 320), as opposed to “’semantic’ analysis [which is] performed in the temporal lobe” (p.314) [3]. Thus, the meaning of objects is not coded in F5, although clearly, “the semantic system can influence the pragmatic system” (p. 320) [3] (e.g., we want to reach for food not snakes).
MNs were discovered within this same circuit and found to have similar sensorimotor properties [1, 4]. It was even suggested that “the actions performed by other monkeys must be a very important factor in determining action selection” (p. 179) [4] and that “the [motor] vocabulary of F5 can be addressed in two ways: by objects and by events [actions]” (p. 317) [3]. Thus, the theoretical and empirical pieces were in place to interpret MNs as sensorimotor association cells relevant to action selection, just like object-oriented cells (Figure 1). But this interpretation was not considered. Why?
Figure 1.
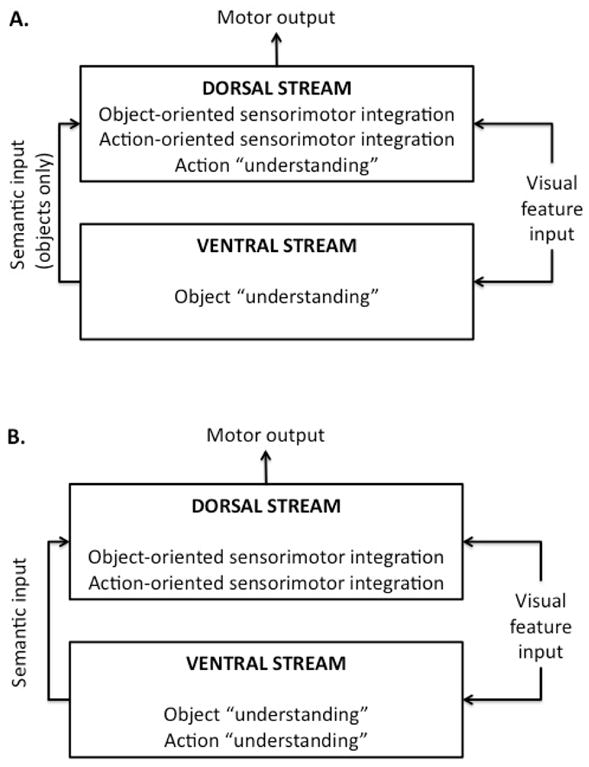
Schematic models of dorsal and ventral stream function. A. The current dominant model [1], which holds that object- and action-oriented processes for sensorimotor integration and “understanding” are organized differentially, with action understanding part of the dorsal sensorimotor stream and object “understanding” part of the ventral stream. B. A more conventional model in which object- and action-oriented processes for sensory-motor integration and understanding are organized similarly. Both models assume that semantic information from the ventral stream can modulate sensorimotor processes in the dorsal stream.
Do monkeys mirror?
It was the mirroring property of MNs that steered investigators away from a straightforward sensorimotor interpretation. The logic was, if mirror actions (e.g., imitation) are not in the species’ repertoire, then mirror neurons can have no motor selection function. Rizzolatti and Craighero used this argument, pitting “two main hypotheses” of MN function, imitation and action understanding; since macaques don’t imitate, they argued, MNs must support action understanding (p. 172) [1]. However, these authors, and the field generally, have failed to notice that other, goal-directed forms of mirror actions are in the macaque motor repertoire, e.g., in competitive reaching for food and in coalition attacks [5]. Further, experimental work has found that another’s grasping actions toward one of two food receptacles serves as a cue to goal-directed grasping toward that same receptacle [6] -- an experimental situation reminiscent of the MN studies. Even domesticated dogs mirror goal-directed actions of a model dog [7]; one would expect to find MNs in dogs given this behavioral evidence.
Observed actions can serve as important inputs to action selection, including but not necessarily limited to, mirror actions. Therefore, the motivating argument for the action understanding theory over a sensorimotor theory (e.g., [8]) does not hold.
Distinguishing sensorimotor and action understanding theories
Can we distinguish the sensorimotor and action understanding theories of MNs? Yes. Empirical findings favor the sensorimotor account by showing that action understanding and motor system function dissociate [9], that motor actions alone are insufficient to explain action understanding [10], that animals comprehend many actions that they cannot execute [10], and that sensorimotor learning can transform the mirror system [8].
In summary, a sensorimotor theory can explain the response properties of MNs, does so more straightforwardly, and doesn’t suffer the empirical roadblocks of the action understanding theory [9, 10]. It is time to reconsider MN function and the neural basis of action understanding.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Rizzolatti G, Craighero L. The mirror-neuron system. Annu Rev Neurosci. 2004;27:169–192. doi: 10.1146/annurev.neuro.27.070203.144230. [DOI] [PubMed] [Google Scholar]
- 2.Rizzolatti G, Camarda R, Fogassi L, Gentilucci M, Luppino G, Matelli M. Functional organization of inferior area 6 in the macaque monkey. II. Area F5 and the control of distal movements. Exp Brain Res. 1988;71:491–507. doi: 10.1007/BF00248742. [DOI] [PubMed] [Google Scholar]
- 3.Jeannerod M, Arbib MA, Rizzolatti G, Sakata H. Grasping objects: the cortical mechanisms of visuomotor transformation. Trends Neurosci. 1995;18:314–320. [PubMed] [Google Scholar]
- 4.di Pellegrino G, Fadiga L, Fogassi L, Gallese V, Rizzolatti G. Understanding motor events: a neurophysiological study. Exp Brain Res. 1992;91:176–180. doi: 10.1007/BF00230027. [DOI] [PubMed] [Google Scholar]
- 5.Hauser MD, Glynn D, Wood J. Rhesus monkeys correctly read the goal-relevant gestures of a human agent. Proc Biol Sci. 2007;274:1913–1918. doi: 10.1098/rspb.2007.0586. [DOI] [PMC free article] [PubMed] [Google Scholar] [Research Misconduct Found]
- 6.Wood JN, Glynn DD, Phillips BC, Hauser MD. The perception of rational, goal-directed action in nonhuman primates. Science. 2007;317:1402–1405. doi: 10.1126/science.1144663. [DOI] [PubMed] [Google Scholar]
- 7.Range F, Viranyi Z, Huber L. Selective imitation in domestic dogs. Curr Biol. 2007;17:868–872. doi: 10.1016/j.cub.2007.04.026. [DOI] [PubMed] [Google Scholar]
- 8.Heyes C. Where do mirror neurons come from? Neurosci Biobehav Rev. 2010;34:575–583. doi: 10.1016/j.neubiorev.2009.11.007. [DOI] [PubMed] [Google Scholar]
- 9.Hickok G. Eight problems for the mirror neuron theory of action understanding in monkeys and humans. J Cogn Neurosci. 2009;21:1229–1243. doi: 10.1162/jocn.2009.21189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hauser M, Wood J. Evolving the capacity to understand actions, intentions, and goals. Annu Rev Psychol. 2010;61:303–324. C301. doi: 10.1146/annurev.psych.093008.100434. [DOI] [PubMed] [Google Scholar]