

Abstract
Vitamin C (VC) is a crucial antioxidant in the brain. To assess whether different brain regions vary in their sensitivity to oxidative stress induced by VC depletion, we used the gulonolactone oxidase (gulo) knockout mouse. This mouse, like humans, cannot synthesize VC and thus its tissue VC levels can be varied by dietary VC intake. Gulo knockout mice were fed drinking water containing standard (0.33 g/L), low (0.033 g/L) or zero (0 g/L) VC supplementation levels. After 4 weeks, mice were sacrificed and different brain regions removed for assay of VC and malondialdehyde, a marker of lipid peroxidation. Compared to age-matched wild-type controls, the cerebellum, olfactory bulbs and frontal cortex had the highest VC content, whereas the pons and spinal chord had the lowest. However, in mice that did not receive VC, area differences were no longer significant as all values trended towards zero. Malondialdehyde increased in the cortex as VC supplementation was decreased. The same changes were not observed in the cerebellum or pons, suggesting that cortex is more susceptible to oxidative damage from low VC. These results suggest enhanced susceptibility of the cortex to oxidative stress induced by low VC compared to other brain regions.
Keywords: Vitamin C, oxidative stress, malondiadehyde, gulo, brain
1. Introduction
There has been recent resurgence of interest in the function of vitamin C (VC) in the brain. For hundreds of years the juice of citrus fruits has been known to ward off scurvy, and in the 1920s the active component responsible (ascorbic acid, VC), was isolated by Albert von Szent-Györgyi. VC is recognized as one of the most abundant antioxidants in the brain and works synergistically with other antioxidants, such as vitamin E and glutathione, to protect against oxidative stress damage caused by free radicals. VC may also have a neuromodulatory function, with proposed interactions with a number of neurotransmitter systems including acetylcholine, dopamine and glutamate (Harrison and May, 2009; Rice, 2000). VC is transported into neurons in a two-step process via the Sodium Vitamin C Transporter 2 (SVCT2) (Tsukaguchi et al., 1999); from blood to cerebrospinal fluid (CSF) and then from CSF to neurons (Harrison and May, 2009). This permits accumulation of VC in neural cells in much greater concentrations than nearly all other cell types except the adrenal gland. The importance of VC in the brain is suggested by the observation that mice lacking the SVCT2 die at birth, with respiratory failure, possibly related to intra-parenchymal haemorrhage (Sotiriou et al., 2002).
There are several reports of regional differences in VC levels in humans (Mefford et al., 1981; Oke et al., 1987; Schaus, 1957) and in rodents (Ferris et al., 1995; Harrison et al., 2008; Odumosu and Wilson, 1980; Stamford et al., 1984) and in more detail within specific brain areas (Basse-Tomusk and Rebec, 1991). There are many problems with examining human brain tissue, such as time between death and tissue removal, age at death, and reason for death. SVCT2 distribution varies across brain areas (Mun et al., 2006). It seems most likely that VC distribution will mirror SVCT2 distribution. Others have theorized that structures closest to ventricles that contain CSF will have the greatest VC levels, presumably because there is less distance for the VC to be transported (Oke et al., 1987). Still other studies have examined whether VC distribution correlates with catecholamine levels or other neurotransmitters (Mefford et al., 1981) and a further suggestion is that VC distribution follows a decreasing anterior-posterior pattern (Milby et al., 1982; Oke et al., 1987; Rice et al., 1995). Cell type and number also influence VC level. VC is found in high levels in neurons, but not glial cells and thus VC levels should be highest in areas with high concentrations of neuronal cell bodies rather than supporting cells and white matter (Rice, 2000; Rice and Russo-Menna, 1998).
Most mammals can synthesize their own VC in the liver from glucose. Humans, primates, guinea pigs and a few other species lack this ability due to the evolutionary loss of the gulo gene (L-gulonolactone oxidase), which is responsible for the final step in VC synthesis. Generation of the gulo(-/-) mouse line (Maeda et al., 2000) presented the ability to control VC level in mice via dietary manipulation, and thus artificially lower VC level in tissues. In guinea pigs dying of scurvy, brain VC levels are maintained at approximately 25 % of normal, even when VC is undetectable in other organs (Hughes et al., 1971). James Lind, the British naval surgeon notable for his research on scurvy in the 17th Century, also reported that the brains of sailors that had died of scurvy were found to be “sound and entire” (Lind, 1772). Thus, it seems that the brain will sequester VC at the expense of other organs in conditions of low VC intake. This supports the notion of a critical role for VC in the brain other than merely as an anti-scorbutic. Whether different areas of the brain have varying retention strengths is not known. Understanding the relationship between brain VC level and oxidative stress may have significance for a number of neurodegenerative diseases such as Alzheimer's, Parkinson's and Huntington's diseases in which oxidative stress and antioxidant imbalances are strongly implicated in their etiology (Halliwell, 2006). In fact, even in normal populations, a surprisingly large percent of the population has depleted or deficient plasma VC levels (Hampl et al., 2004; Johnston et al., 1998; Johnston and Thompson, 1998).
Despite the range of current research on VC in the brain, to our knowledge there has not been a comprehensive study of VC distribution in the brain in mice. Neither is the relationship between VC depletion and sensitivity to lipid peroxidation known. The goals of the present study were: first, to identify any differences in VC level in various brain areas under normal physiological conditions in wild-type mice; second, to discover whether any of these brain areas differed in VC retention in conditions of prolonged low VC intake; and third, to see whether VC level and retention led to differences in oxidative stress among the areas. This final aim is critical in determining the role of VC in the protection of the brain against oxidative damage. The study was carried out using gulo(-/-) mice, in which VC intake can be controlled, under standard (STD) and low (LOW) VC supplement conditions, or following 4 weeks of complete VC deprivation (WATER), with gulo(+/+) wild-type (WT) mice as controls.
2. Results
2.1 Weight in WATER mice
Although male mice weighed slightly more than female mice, there was no significant weight loss observed over the course of the 4-week VC-deprivation period in either sex (p> 0.29). Male WATER mice weighed on average 23.275 g (± S.E.M. 1.20) at the start of the VC-deprivation period and 25.25 g (± S.E.M. 1.23) at the end. Female mice weighed on average 19.01 g (± S.E.M. 0.65) at the start of the VC-deprivation period and 18.71 g (± S.E.M. 1.26) at the end.
2.2 Vitamin C distribution in WT and gulo(-/-) mice
When WT mice alone were analyzed, a significant omnibus ANOVA (p<0.05) indicated variability in VC level among the various brain regions assayed. Hippocampus and cerebellum had the highest VC contents whereas the lowest values were found in the pons and spinal chord (Table 1). Cerebellum had higher VC than almost all other areas (p<0.05), but was similar to parietal cortex and hippocampus. Hippocampus VC was higher than olfactory bulbs, striatum and thalamus (p<0.05). The pons had lower VC than both frontal and parietal cortex, cerebellum and striatum (p<0.05). Spinal cord was also significantly lower than cerebellum, although had slightly higher VC levels than pons.
Table 1. Vitamin C level in brain.
| Gulo(+/+) Control n=14 |
Gulo(-/-) STD n=7 |
Gulo(-/-) LOW n=12 |
Gulo(-/-) WATER n=12 |
|
|---|---|---|---|---|
| μmol/g tissue (wet weight) | ||||
| Olfactory bulbs | 2.16 ±0.31a | 2.50 ±0.15a | 1.08 ±0.10b | 0.25 ±0.04c |
| Frontal cortex | 2.17 ±0.19a | 2.05 ±0.13a | 0.77 ±0.12b | 0.21 ±0.09c |
| Parietal cortex | 2.29 ±0.22a | 2.12 ±0.19a | 0.60 ±0.04b | 0.15 ±0.02b |
| Striatum | 2.06 ±0.18a | 1.66 ±0.09a | 0.53 ±0.07b | 0.15 ±0.02b |
| Thalamic nucleii | 2.18 ±0.19a | 1.55 ±0.17b | 0.68 ±0.08c | 0.25 ±0.05c |
| Hippocampus | 2.57 ±0.33a | 1.95 ±0.15a | 0.87 ±0.11b | 0.22 ±0.04b |
| Cerebellum | 2.51 ±0.27a | 2.17 ±0.22a | 1.30 ±0.13b | 0.40 ±0.04c |
| Pons | 1.63 ±0.13a | 1.04 ±0.12b | 0.40 ±0.03c | 0.11 ±0.02c |
| Spinal chord | 1.92 ±0.16a | 0.96 ±0.23b | 0.38 ±0.08c | 0.07 ±0.01c |
| Liver | 0.92 ±0.09a | 0.59 ±0.10b | 0.13 ±0.11c | 0.14 ±0.02c |
VC level in each brain area measured for each group. Values in the same row (brain area) that do not share a superscript letter are significantly different, p<0.05.
When all groups were analyzed together there were significant main effects of brain region (p<0.001), group (p<0.001), and also a significant interaction between the two factors (p<0.001). The main effect of group, when all brain areas for a particular group are considered together, suggests that VC levels of STD mice did not differ from those of WT mice, and both groups had significantly higher VC levels than both LOW and WATER mice across all brain areas (p<0.001; Table 1). WATER mice also had lower VC levels than LOW mice (p<0.05). Comparisons among the different areas showed no difference between spinal chord and pons, but both of these areas were significantly lower than all other areas assayed (p<0.01). The highest VC values overall were found in the cerebellum, olfactory bulbs and frontal cortex. The Region × Group interaction demonstrated that differences among groups and regions were not perfectly linear. Group differences were seen in every brain area assayed (p<0.001; Table 1). Within groups, differences between areas were seen in WT, STD and LOW groups (ps<0.001) but not in the WATER group in which latter case all VC values were below 0.5 μmol/g tissue. VC levels in liver should provide a good indication of the overall VC status of an animal. There were significant differences among the groups (p<0.001; Table 1). VC liver levels differed among all the groups (p<.001) except LOW and WATER mice, which did not differ (p=0.83; Table 1).
2.3 Vitamin C retention in gulo(-/-) mice
Retention was calculated as a percentage of wild-type controls (Fig. 1). In general, VC retention reflected original values, thus, greatest retention in STD mice was seen in olfactory bulbs (>80% of WT), cerebellum and frontal and parietal cortex. In LOW mice, greatest retention (>40% of WT) was found only in cerebellum and olfactory bulbs. In WATER mice only cerebellum, olfactory bulbs and thalamus were >10% of WT controls. There were significant main effects of brain region, and group and a significant Region × Group interaction (p<0.001). Each group differed from each of the other two groups, with percent retention decreasing as expected with VC supplementation level. These group differences were evident in each area (p<0.01) except in the spinal chord, where LOW mice were not different from WATER mice (p=0.094).
Figure 1. Vitamin C retention calculated as percentage of Gulo(+/+).
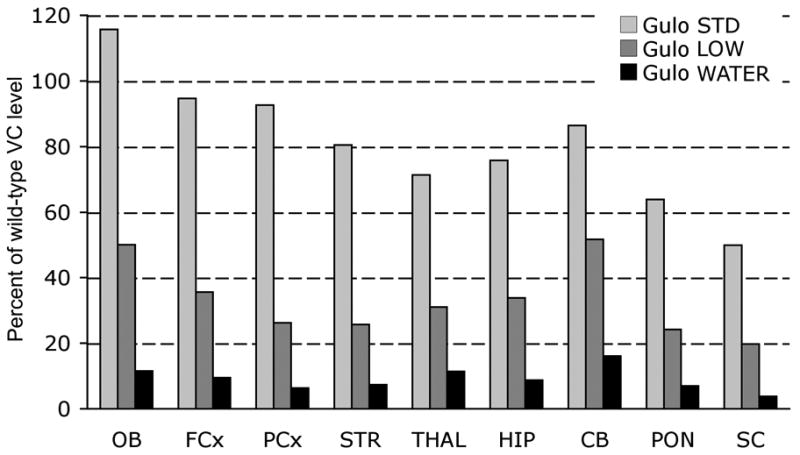
VC retention is reported in Olfactory bulbs (OB), Frontal cortex (FCx), Parietal cortex (PCx), Striatum (STR), Thalamus (THAL), Hippocampus (HIP), Cerebellum (CB), Pons (PON), and Spinal cord (SC). Values reported are group means.
In STD mice many group differences were preserved among the areas. Lower retention was observed in pons and spinal cord which did not differ from each other but had significantly lower retention than all other areas (p<0.05) except thalamus and hippocampus. In LOW mice, VC retention in the cerebellum (51.69 %) and olfactory bulbs (50.05 %) did not differ from each other but both had greater retention than each of the other areas (p<0.05). None of the lower values differed from one another. No specific differences were observed in WATER mice among areas as they all approached zero.
2.4 Oxidative stress in cortex, cerebellum and pons
Omnibus ANOVA presented significant main effects of brain region (p<0.001) and group (p<.001) with a significant interaction between the factors (p<0.01; Fig. 2). Bonferroni follow-up comparisons clarified the differences among the groups and brain regions. In wild-type mice MDA was similar in cortex, cerebellum and pons (p>0.05). In STD, LOW and WATER mice, there were higher levels of MDA in the cortex than cerebellum (p<0.01). In LOW mice MDA was also greater in cortex than pons (p<0.01). In WATER mice MDA was lower in cerebellum than pons (p<0.001). Comparisons were also made within each area. In the cortex, LOW and WATER groups had elevated MDA levels compared to wild-type mice (p<0.01), but STD mice did not differ from controls (p=0.295). MDA levels were equivalent in all groups in the cerebellum (p>0.126) and also in the pons (p>0.071).
Figure 2. Malondialdehyde in cortex, cerebellum and pons.
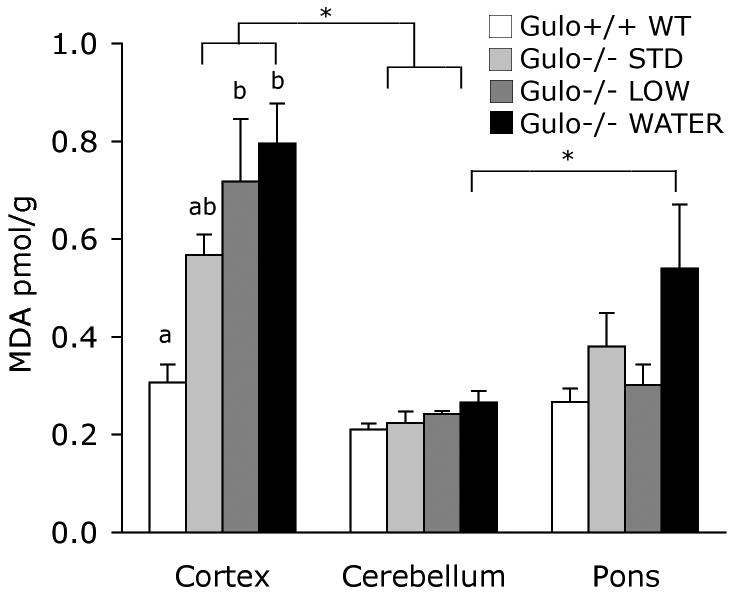
MDA was elevated in all gulo(-/-) mice in the cortex but did not differ among groups in cerebellum and pons. MDA was greater in cortex than cerebellum in all gulo(-/-) groups, and was lower in the cerebellum than pons in gulo(-/-) WATER mice. Values from the same area that have different letters are different p<0.01. Other differences are as marked, *p<0.01.
3. Discussion
3.1 Regional differences in vitamin C distribution
In gulo(+/+) WT mice VC levels were highest in the cerebellum, olfactory bulbs and frontal cortex. The pons and spinal chord consistently had the lowest VC levels. These data agree with reports from similar studies in other rodent species such as well-supplemented guinea pigs (Odumosu and Wilson, 1980) and rats (Milby et al., 1982). The distribution does not perfectly fit the anterior-posterior VC distribution theory noted previously and neither does it support the theory that VC level is based simply on proximity to ventricles. In fact, the simplest explanation for the difference depends on the type of cells found within each area. VC cannot cross the blood brain barrier and enters extracellular fluid of the brain via SVCT2 in the choroid plexus. SVCT2 is limited to the neurons, which is where the highest concentrations of VC are stored. Glial cells, including astrocytes also carry VC. These cells do not have the SVCT2 and VC enters the cells in its oxidized form, dehydroascorbic acid, via GLUT transporters where it is then recycled within the cell to VC. Thus, where there is the greatest concentration of neuronal cell bodies, cortex, hippocampus and cerebellum, there are higher VC levels. In pons and spinal chord, areas that contain more white matter as myelinated nerve fibers, less VC is found than in neuron-rich cortex and hippocampus. Mun and colleagues located SVCT2 within microglia but not astrocytes (Mun et al., 2006). Within the neuron it was found in cell bodies but not in the axonal or dendritic processes of cells. Thus the VC level depends significantly on the structure involved.
It was expected that VC level would reflect that of SVCT2 distribution. Indeed, Tsukaguchi and colleagues (Tsukaguchi et al., 1999) showed very high SVCT2 by in situ hybridization in cerebellum, hippocampus and olfactory bulb, which mirrors our findings. On the other hand, when SVCT2 distribution was assessed by immunohistochemical techniques in rat brain (Mun et al., 2006), SVCT2 was highest in frontal cortex but the SVCT2 was not seen at all in parietal cortex. Mun and colleagues ascribed each area examined a score on a 4-point scale, from 0-3, with 4 being the highest, and 0 being no SVCT2 reactivity. The hippocampus was rated between 1 and 3 depending on sub-region, the cerebellum was rated at 2, and the pons from 0-1. This quantification does not exactly agree with the values we found or with SVCT2 mRNA expression, so the steady-state tissue VC level may not always depend on the level of the SVCT2. Although these studies were performed in different species, it is unlikely that there is so significant a variation between rat and mouse as to negate this comparison. One area of clear agreement between this study and SVCT2 distribution is that the pons and spinal cord have lower SVCT2 expression and VC content than other brain areas. This may account for our previous observations that this was the area of damage in a guinea pig model of combined VC and vitamin E deficiency (Hill et al., 2003).
Distribution studies for Vitamin E, VE, performed in rats, consistently show that cortex has much higher VE than cerebellum. In more detailed studies hippocampus VE is high while striatum is even lower than cerebellum (Martin et al., 2000; Vatassery et al., 1984). These areas also have different retention and storage rates when VE is supplemented removed from the diet (Martin et al., 2000; Meydani et al., 1986). These data are all from rat studies and actual VE values vary among rodent species (Vatassery and Younoszai, 1978) but it is expected that the relative distribution across brain areas would be similar in mouse.
3.2 Group differences in vitamin C distribution and retention
Despite slight regional variations, VC levels in brains of STD mice were similar to those in the WT mice in all areas except thalamus, pons and spinal cord. Four weeks of LOW treatment significantly lowered VC levels in the gulo(-/-) mice and 4 weeks of VC deprivation led to even lower tissue VC in the WATER mice. The values obtained after 4 weeks on LOW were similar to those previously reported in mice exposed to this treatment for 12 weeks (Harrison et al., 2008) and thus probably reflect a steady-state low level of tissue VC had been reached. In each case VC distribution varied similarly among the areas except in the case of the WATER mice, in which all areas were tending toward zero and were no longer different. These data suggest the importance of the regional distribution pattern, which remains in place until stores are so severely depleted that such differences can no longer be preserved. We did not observe weight loss in the WATER mice following 4 weeks of VC deprivation, however, neither did we see the weight gain that would be expected if mice were VC-replete. Thus, the changes in brain occurred prior to onset of clinical symptoms of scurvy. Nevertheless, we selected the 4-week treatment time because in our experience this represents the limit of VC tolerance of gulo(-/-) mice at this age and we would expect extreme weight loss and sickness to occur in week 5.
We found greater relative decreases in liver VC levels as VC stores were, relative to losses in cortex and cerebellum. This pattern fits with the results of studies in guinea pigs, in which VC was retained in brain at 18-25% of normal even after extreme deprivation that caused very low levels in other organs (Hughes et al., 1971; Saner et al., 1975), which were less than 2 % VC remaining in spleen (Hughes et al., 1971), less than 5 % in liver (Hill et al., 2003), less than 2 % in leucocytes (Saner et al., 1975). However, after severe depletion in the WATER group in the present study, most brain areas were depleted of VC to 10% of control. It may be that the natural loss of gulo through evolutionary means in guinea pigs permitted the development of compensatory systems in order to protect brain levels that are lacking in mice but even given the methodological differences among the guinea pig studies, the variation among these results suggests that similar relationships exist in the gulo(-/-) mouse.
3.3 Vitamin C deprivation and Oxidative stress
In both conditions of restricted VC, but also in the presumed-to-be-sufficient STD mice, cortex had elevated levels of MDA that corresponded to the supplementation level. In contrast, MDA was not elevated in cerebellum even in WATER mice. Similarly, in our earlier study, F4-neuroprostanes, another measure of oxidative stress specific to neurons (Roberts and Fessel, 2004), were elevated in cortex in both STD and LOW supplemented gulo(-/-) mice, but in the cerebellum F4-neuroprostanes were only elevated in LOW supplemented mice (Harrison et al., 2008). These data suggest greater vulnerability to low VC in cortex and/or greater protection in the cerebellum. Although in general the cerebellar VC levels were higher than those in the cortex, the difference was not large enough to account for the variations in MDA. Excepting the WT mice, cerebellar MDA was lower than cortex MDA in all other groups, with the greater differences observed in the lower VC supplements. In the pons, there were no group differences, although there did seem to be a trend toward greater MDA in the gulo(-/-) mice. In combination these data reinforce the idea that all brain regions are not equally resilient to low VC and increases in oxidative stress. This is an area that merits detailed future investigation as it may provide valuable information into the course and treatment of neurodegenerative diseases. VC is a vital antioxidant in the brain. Decreasing levels of VC were clearly associated with increasing levels of lipid peroxidation products in the cortex. One could conjecture that in the cerebellum the VC was sufficient for antioxidant roles. However, in the cortex if VC has a different primary role, such as catecholamine synthesis, or some other neuromodulatory function, or if there was simply insufficient VC, then that area suffered. What is clear is that it is important part of the brain is used in studies and using whole brain may cloud relevant information. There are many changes associated with oxidative stress including apoptosis, and thus cognition and vital function may be affected by low VC. This might be especially relevant during development. Even in the absence of physical signs of scurvy these changes may damage human cognitive health, which could be even more important in neurodegenerative disorders.
3.4 Conclusions
When designing this study we had three goals in mind. The first was to identify differences among brain areas in the wild-type brain. We have clearly shown that there are large differences in VC level between areas and propose that these are likely related to cell type within each area. The second aim was to identify differences in VC retention under low VC conditions. VC retention was greater in olfactory bulbs and cerebellum than other areas in LOW mice, but as all VC stores became depleted in WATER mice, these variations were no longer evident. Finally we sought to investigate whether VC-deficiency-induced lipid peroxidation differed among the different areas and whether this was linked to VC retention. No change in MDA level was observed in cerebellum, which had the highest VC retention in LOW mice. The pons had very poor VC retention but LOW mice did not show increased MDA, whereas the cortex, which appeared to have better VC retention, showed the greatest changes in MDA level in LOW and water mice. Thus the relationship between VC level and oxidative stress varies depending on the area under investigation. These results are particularly important when making comparisons between studies that may use whole brain samples, or focus on different brain regions.
4. Experimental Procedure
4.1 Animals
C57BL/6J were obtained from Jackson laboratories (stock #000664) and a colony was maintained in-house. Gulo(-/-) mice were bred in-house from heterozygous gulo(+/-) mice obtained from Mutant Mouse Regional Resource Centers (http://www.mmrrc.org, stock #000015-UCD) and were maintained on a C57BL/6J background. gulo(-/-) male and female mice were mated to obtain litters of gulo(-/-) mice for the present study. Mice had free access to food and water. Mice were fed standard lab chow, (#5001; Purina Mills, St Louis, MO, USA) which contains no added VC. C57BL/6J mice received tap water. Gulo(-/-) mice received de-ionized water with 0.33 g/L VC (ascorbic acid, Sigma Aldrich) (Standard; STD), or 0.033 g/L VC (Low; LOW). 20 μl/L EDTA was added to treated water to help stabilize the VC in solution. Fresh supplemented water bottles were made up twice per week. Gulo(-/-) mice were all maintained on STD supplementation from weaning until just prior to the experiment when mice were randomly allocated to one of three groups; STD mice continued on the same supplement level as before, LOW mice were given the lower supplementation level for 4 weeks prior to sacrifice; WATER mice were given non-supplemented water for 4 weeks prior to sacrifice. WATER mice were weighed twice per week and observed daily to monitor for potential signs of scurvy (weight loss, hunched posture, poor grooming). Although we have not previously found sex differences in VC levels, (Harrison et al., 2008), sex differences have previously been observed in other studies (Ferris et al., 1995) and thus we used equal numbers of males and females and analyzed data to check for sex differences before collapsing data into a single group. Mice were sacrificed between 9 and 12 weeks of age. All procedures were approved by the Vanderbilt University Institutional Animal Care and Use Committee and were conducted in accordance with the NIH Guide for the Care and Use of Laboratory Animals. Final groups numbers were gulo(+/+) WT n = 14, gulo(-/-) STD n = 7, gulo(-/-) LOW n = 12, and gulo(-/-) WATER n = 12.
4.2 Tissue collection
Mice were anaesthetized using inhaled isoflurane and then sacrificed by decapitation. Brains were quickly removed from the skull and dissected on ice. Olfactory bulbs were removed first. Then cerebellum, brain stem (pons) and spinal cord were removed. The remaining tissue was hemisected and thalamus, hippocampus and striatum were dissected out. Two cortex samples were taken corresponding to frontal and parietal regions. Cortex, cerebellum and pons samples were split into two for oxidative stress measurements. Liver samples were also removed and stored at -80 °C. Samples were weighed prior to storage at -80 °C until assayed for VC or MDA.
4.3 Vitamin C measurements
Vitamin C concentrations were measured by ion pair HPLC (Pachla and Kissinger, 1979) and electrochemical detection as previously described (Harrison et al., 2008; May et al., 1998), except that tetrapentyl ammonium bromide was used as the ion pair reagent.
4.4 Malondialdehyde measurements
Weighed tissue samples were homogenized with 1 ml 5% trichloroacetic acid and centrifuged at 13,000 g for 5 min at 4 °C. 250 μl of the supernantant was removed and added to 250 μl of 0.02 M thiobarbituric acid. Samples were heated at 95 °C for 35 mins and cooled for 10 mins at 4 °C. A standard curve was calculated from known concentrations of MDA treated in the same way as samples. MDA was specifically measured on a Spectramax M5 microplate reader (Molecular Devices, USA) with an excitation wavelength of 515 and an emission wavelength of 553 as previously described (Harrison et al., 2009).
4.5 Statistics
Data were analysed using SPSS 18 for Mac. Liver was analyzed using univariate ANOVA. VC and MDA levels across brain areas were measured using repeated measures ANOVA with area as the within subjects factor and group as the between subjects factor. Follow-up analyses were conducted using Bonferroni post-hocs or Bonferroni corrected T-tests. In cases where assumptions of sphericity were violated Greenhouse-Geisser corrected p values are given. Percent change was calculated using group means for each gulo(-/-) group compared to the group mean for the WT group.
Acknowledgments
This work was supported by NIH grant NS057674-03 to J.M.M.
Abbreviations
- VC
Vitamin C
- MDA
Malondialdehyde
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Basse-Tomusk A, Rebec GV. Regional distribution of ascorbate and 3,4-dihydroxyphenylacetic acid (DOPAC) in rat striatum. Brain Res. 1991;538:29–35. doi: 10.1016/0006-8993(91)90372-3. [DOI] [PubMed] [Google Scholar]
- Ferris DC, et al. Gender differences in cerebral ascorbate levels and ascorbate loss in ischemia. Neuroreport. 1995;6:1485–9. doi: 10.1097/00001756-199507310-00005. [DOI] [PubMed] [Google Scholar]
- Halliwell B. Oxidative stress and neurodegeneration: where are we now? J Neurochem. 2006;97:1634–58. doi: 10.1111/j.1471-4159.2006.03907.x. [DOI] [PubMed] [Google Scholar]
- Hampl JS, et al. Vitamin C deficiency and depletion in the United States: the Third National Health and Nutrition Examination Survey, 1988 to 1994. Am J Public Health. 2004;94:870–5. doi: 10.2105/ajph.94.5.870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison FE, et al. Ascorbic acid attenuates scopolamine-induced spatial learning deficits in the water maze. Behav Brain Res. 2009;205:550–558. doi: 10.1016/j.bbr.2009.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison FE, May JM. Vitamin C function in the brain: vital role of the ascorbate transporter SVCT2. Free Radic Biol Med. 2009 doi: 10.1016/j.freeradbiomed.2008.12.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison FE, et al. Elevated oxidative stress and sensorimotor deficits but normal cognition in mice that cannot synthesize ascorbic acid. J Neurochem. 2008;106:1198–1208. doi: 10.1111/j.1471-4159.2008.05469.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill KE, et al. Combined deficiency of vitamins E and C causes paralysis and death in guinea pigs. Am J Clin Nutr. 2003;77:1484–8. doi: 10.1093/ajcn/77.6.1484. [DOI] [PubMed] [Google Scholar]
- Hughes RE, et al. The retention of ascorbic acid by guinea-pig tissues. Br J Nutr. 1971;26:433–8. doi: 10.1079/bjn19710048. [DOI] [PubMed] [Google Scholar]
- Johnston CS, et al. Vitamin C status of a campus population: college students get a C minus. J Am Coll Health. 1998;46:209–13. doi: 10.1080/07448489809600224. [DOI] [PubMed] [Google Scholar]
- Johnston CS, Thompson LL. Vitamin C status of an outpatient population. J Am Coll Nutr. 1998;17:366–70. doi: 10.1080/07315724.1998.10718777. [DOI] [PubMed] [Google Scholar]
- Lind J. A treatise on the scurvy. In three parts. Containing an inquiry into the nature, causes, and cure, or that disease. Together with a critical and chronological view of what has been published on the subject. Leslie B. Adams, Jr; London: 1772. [Google Scholar]
- Maeda N, et al. Aortic wall damage in mice unable to synthesize ascorbic acid. Proc Natl Acad Sci U S A. 2000;97:841–6. doi: 10.1073/pnas.97.2.841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin A, et al. Effect of fruits, vegetables, or vitamin E--rich diet on vitamins E and C distribution in peripheral and brain tissues: implications for brain function. J Gerontol A Biol Sci Med Sci. 2000;55:B144–51. doi: 10.1093/gerona/55.3.b144. [DOI] [PubMed] [Google Scholar]
- May JM, et al. Protection and recycling of alpha-tocopherol in human erythrocytes by intracellular ascorbic acid. Arch Biochem Biophys. 1998;349:281–9. doi: 10.1006/abbi.1997.0473. [DOI] [PubMed] [Google Scholar]
- Mefford IN, et al. Regional distribution of ascorbate in human brain. Brain Res. 1981;212:223–6. doi: 10.1016/0006-8993(81)90056-1. [DOI] [PubMed] [Google Scholar]
- Meydani M, et al. Influence of dietary vitamin E, selenium and age on regional distribution of alpha-tocopherol in the rat brain. Lipids. 1986;21:786–91. doi: 10.1007/BF02535413. [DOI] [PubMed] [Google Scholar]
- Milby K, et al. Detailed mapping of ascorbate distribution in rat brain. Neurosci Lett. 1982;28:169–74. doi: 10.1016/0304-3940(82)90147-1. [DOI] [PubMed] [Google Scholar]
- Mun GH, et al. Immunohistochemical study of the distribution of sodium-dependent vitamin C transporters in adult rat brain. J Neurosci Res. 2006;83:919–28. doi: 10.1002/jnr.20751. [DOI] [PubMed] [Google Scholar]
- Odumosu A, Wilson CW. Regional brain ascorbic acid distribution: its functional relationship to appetite and leptazol-induced convulsions in guinea-pigs. Int J Vitam Nutr Res. 1980;50:52–60. [PubMed] [Google Scholar]
- Oke AF, et al. Ascorbic acid distribution patterns in human brain. A comparison with nonhuman mammalian species. Ann N Y Acad Sci. 1987;498:1–12. doi: 10.1111/j.1749-6632.1987.tb23747.x. [DOI] [PubMed] [Google Scholar]
- Pachla LA, Kissinger PT. Analysis of ascorbic acid by liquid chromatography with amperometric detection. Methods Enzymol. 1979;62:15–24. doi: 10.1016/0076-6879(79)62183-3. [DOI] [PubMed] [Google Scholar]
- Rice ME. Ascorbate regulation and its neuroprotective role in the brain. Trends Neurosci. 2000;23:209–16. doi: 10.1016/s0166-2236(99)01543-x. [DOI] [PubMed] [Google Scholar]
- Rice ME, et al. High levels of ascorbic acid, not glutathione, in the CNS of anoxia-tolerant reptiles contrasted with levels in anoxia-intolerant species. J Neurochem. 1995;64:1790–9. doi: 10.1046/j.1471-4159.1995.64041790.x. [DOI] [PubMed] [Google Scholar]
- Rice ME, Russo-Menna I. Differential compartmentalization of brain ascorbate and glutathione between neurons and glia. Neuroscience. 1998;82:1213–23. doi: 10.1016/s0306-4522(97)00347-3. [DOI] [PubMed] [Google Scholar]
- Roberts LJ, 2nd, Fessel JP. The biochemistry of the isoprostane, neuroprostane, and isofuran pathways of lipid peroxidation. Chem Phys Lipids. 2004;128:173–86. doi: 10.1016/j.chemphyslip.2003.09.016. [DOI] [PubMed] [Google Scholar]
- Saner A, et al. Cerebral monoamine metabolism in guinea-pigs with ascorbic acid deficiency. J Pharm Pharmacol. 1975;27:896–902. doi: 10.1111/j.2042-7158.1975.tb10244.x. [DOI] [PubMed] [Google Scholar]
- Schaus R. The ascorbic acid content of human pituitary, cerebral cortex, heart, and skeletal muscle and its relation to age. Am J Clin Nutr. 1957;5:39–41. doi: 10.1093/ajcn/5.1.39. [DOI] [PubMed] [Google Scholar]
- Sotiriou S, et al. Ascorbic-acid transporter Slc23a1 is essential for vitamin C transport into the brain and for perinatal survival. Nat Med. 2002;8:514–7. doi: 10.1038/0502-514. [DOI] [PubMed] [Google Scholar]
- Stamford JA, et al. Regional differences in extracellular ascorbic acid levels in the rat brain determined by high speed cyclic voltammetry. Brain Res. 1984;299:289–95. doi: 10.1016/0006-8993(84)90710-8. [DOI] [PubMed] [Google Scholar]
- Tsukaguchi H, et al. A family of mammalian Na+-dependent L-ascorbic acid transporters. Nature. 1999;399:70–5. doi: 10.1038/19986. [DOI] [PubMed] [Google Scholar]
- Vatassery GT, et al. Concentrations of vitamin E in various neuroanatomical regions and subcellular fractions, and the uptake of vitamin E by specific areas, of rat brain. Biochim Biophys Acta. 1984;792:118–22. doi: 10.1016/0005-2760(84)90211-x. [DOI] [PubMed] [Google Scholar]
- Vatassery GT, Younoszai R. Alpha tocopherol levels in various regions of the central nervous systems of the rat and guinea pig. Lipids. 1978;13:828–31. doi: 10.1007/BF02533486. [DOI] [PubMed] [Google Scholar]