

Abstract
Selective breeding for performance has resulted in distinct breeds of horse, such as the Quarter Horse (bred for acceleration) and the Arab (bred for endurance). Rapid acceleration, seen during Quarter Horse racing, requires fast powerful muscular contraction and the generation of large joint torques, particularly by the hind limb muscles. This study compared hind limb moment arm lengths in the Quarter Horse and Arab. We hypothesized that Quarter Horse hind limb extensor muscles would have longer moment arms when compared to the Arab, conferring a greater potential for torque generation at the hip, stifle and tarsus during limb extension. Six Quarter Horse and six Arab hind limbs were dissected to determine muscle moment arm lengths for the following muscles: gluteus medius, biceps femoris, semitendinosus, vastus lateralis, gastrocnemius (medialis and lateralis) and tibialis cranialis. The moment arms of biceps femoris (acting at the hip) and gastrocnemius lateralis (acting at the stifle) were significantly longer in the Quarter Horse, although the length of the remaining muscle moment arms were similar in both breeds of horse. All the Quarter Horse muscles were capable of generating greater muscle moments owing to their greater physiological cross-sectional area (PCSA) and therefore greater isometric force potential, which suggests that PCSA is a better determinant of muscle torque than moment arm length in these two breeds of horse. With the exception of gastrocnemius and tibialis cranialis, the observed muscle fascicle length to moment arm ratio (MFL : MA ratio) was greater for the Arab horse muscles. It appears that the Arab muscles have the potential to operate at slower velocities of contraction and hence generate greater force outputs when compared to the Quarter Horse muscles working over a similar range of joint motion; this would indicate that Arab hind limb muscles are optimized to function at maximum economy rather than maximum power output.
Keywords: Arab, biomechanics, equine, locomotion, moment arm, muscle, Quarter Horse
Introduction
The horse is an exceptional athlete, capable of rapid acceleration over short distances whilst maintaining energy-efficient locomotion over longer distances.
Selective breeding has continued to capitalize on these inherent performance traits, resulting in breeds of horse that excel at different equine athletic disciplines.
The Quarter Horse and the Arab are bred for their different racing performances. The Quarter Horse [named after its ability to rapidly accelerate over a quarter of a mile (402 m)], has been shown to be an even faster sprinter than both the racing Thoroughbred and the Arab (Nielsen et al., 2006). In comparison, the Arab (better suited to endurance events [Hoffman et al., 2002)], races at slower speeds over much longer distances and can cover 160 km in 9.5 h (Xandu Haji Buba, 2006). These two breeds of horse, situated at either end of the equine athletic spectrum, provide us with the opportunity to study how equine locomotor muscles might be adapted to operate at different contraction velocities and power outputs.
Previously, we reported that the Quarter Horse hind limb muscles were of a greater muscle volume, physiological cross-sectional area (PCSA) and had greater isometric force potential when compared to those of the Arab (Crook et al., 2008). Here, we compare moment arms of the same hind limb muscles to determine if there are geometric conformational differences that could influence joint angular velocity and joint torque production. Joint angular velocity and muscle force output are inherently linked such that a muscle with a greater length of moment arm is required to shorten at a higher contraction velocity for a given joint angular velocity whilst the muscles force output is simultaneously reduced due to the force–velocity relationship (Nagano & Komura, 2003).
Very few studies have looked at moment arm lengths in the equine limb (Brown et al., 2003a,b;) and few, if any, have attempted to relate differences in equine performance to moment arm length.
Here we chose to compare moment arm lengths in the Quarter Horse and Arab for joint angles corresponding to those measured for the hip, stifle and tarsus joints throughout the functional joint range (Back et al., 1995). We studied moment arm lengths of the hind limb muscles, as these are the muscles which are primarily responsible for powering equine locomotion (Merkens et al., 1993).
We hypothesized that, compared to the Arab, Quarter Horse hind limb extensor muscles would have longer moment arms at the hip, stifle and tarsus during limb extension at joint angles corresponding to those measured during walk, trot and canter over level ground.
Materials and methods
Horses
The study was conducted at the University of Queensland (UoQ) and approved by the UoQ Animal Ethics Committee. Six Quarter Horses [four geldings and two mares, age 13 ± 8 years (mean ± SD)], and six Arab horses [five geldings and one mare, age 13 ± 7 years (mean ± SD)] were euthanized for reasons unrelated to musculoskeletal disease (Table 1). All horses had previously been in light ridden work.
Table 1.
Morphometric details of subjects, including pelvic limb segment measurements used to scale moment arm measurements.
| Age (years) | Sex | Mass (kg) | Height (m) | Femur length (cm) | Tibia length (cm) | Breed | |
|---|---|---|---|---|---|---|---|
| Limb 1 | 20 | Gelding | * | 1.51 | 46.0 | 38.5 | Quarter Horse |
| Limb 2 | 12 | Mare | * | 1.49 | 45.0 | 40.0 | Quarter Horse |
| Limb 3 | 14 | Gelding | 504 | 1.51 | 46.0 | 41.0 | Quarter Horse |
| Limb 4 | 25 | Mare | 461 | 1.52 | 44.0 | 37.0 | Quarter Horse |
| Limb 5 | 3.5 | Gelding | 445 | 1.49 | 45.0 | 40.0 | Quarter Horse |
| Limb 6 | 4.5 | Gelding | 419 | 1.48 | 44.0 | 38.5 | Quarter Horse |
| Limb 7 | 4 | Gelding | * | 1.50 | 44.0 | 37.5 | Arab |
| Limb 8 | 8 | Gelding | * | 1.46 | 43.0 | 36.5 | Arab |
| Limb 9 | 20 | Mare | 388 | 1.41 | 45.0 | 37.0 | Arab |
| Limb 10 | 20 | Gelding | 370 | 1.44 | 41.5 | 33.5 | Arab |
| Limb 11 | 20 | Gelding | 327 | 1.48 | 44.0 | 37.0 | Arab |
| Limb 12 | 20 | Gelding | 448 | 1.51 | 44.5 | 37.5 | Arab |
| Mean QH | 13 | 457 | 1.50 | 45.0 | 39.0 | ||
| Mean Arab | 15 | 383 | 1.47 | 44.0 | 36.5 |
It was not possible to determine the mass of these animals because scales were not available prior to euthanasia.
A tape measure was used to determine the height (m) of each animal, measuring from the ground up to the highest point of the withers, whilst the horse was standing square and sedated in stocks. All of the subjects were unshod. The mass (kg) of eight of the 12 subjects was determined by standing the animal on an electronic weigh scale (Tru Test Easy Weigh II, Sunbeam, Sydney, Australia). The scales were not available to weigh the remaining four subjects.
Tendon travel method
Moment arm lengths for gluteus medius, biceps femoris, semitendinosus, vastus lateralis, gastrocnemius and tibialis cranialis were determined using the tendon travel method (An et al., 1981; Spoor & van Leeuwen, 1992). These muscles were selected as they are a representative sample of the proximal and distal hind limb flexor and extensor muscles. In addition, their superficial positioning enables the use of surface electromyography to determine their activation patterns during different locomotor tasks, which is the subject of a complimentary study.
The tendon travel method is based on the fact that when the radius of a circle moves through an angle of one radian, any point on that radius will have moved through a distance equal in length to the distance between that point and the centre of the circle. If it is assumed that the joint axis is the centre of the circle and the tendon path is the curve of the radius, the distance that the tendon moves whilst the joint moves through an angle of one radian equals the perpendicular distance between the tendon and the joint axis or moment arm. This relationship is expressed by the following equation:
where r is the moment arm length, Lm is the distance moved by the tendon in metres and θ is the joint angle in radians.
Measurement procedure
Moment arms of six muscles were measured, at multiple joints where appropriate, with the pelvic limb placed on a dissection bench.
Measurements were taken from superficial hip then stifle muscles (with progressively deeper muscles exposed for measurement as required), before the distal muscles crossing the tarsus were assessed. The order in which muscles were measured was chosen to avoid disturbing the line of action of muscles, as the measurement technique involves removal of the muscle belly.
Gluteus medius was dissected away from the iliac crest approximately 5 cm away from its proximal insertion. Lengths of stiff fishing nylon (30 lb, 0.5 mm diameter; clear nylon Drennan Sea-Thru, Oxford, UK) were sutured into the remaining muscle stump at five equal distances, starting at the most cranial to the most caudal insertion on the iliac crest; this then allowed five equidistant lines of muscle pull on the greater trochanter to be approximated. In all other muscles, a single length of fishing nylon was sutured into the stump of the individual muscle belly at its bony insertion. In all muscles, the free end of the suture nylon was passed through a 0.5-mm brass eyelet screwed into the relevant long bone close to the muscle’s distal insertion to replicate the muscle’s line of action.
Millimetre scales were super-glued (Loctite, Düsseldorf, Germany) onto the longitudinal aspect of an exposed area of the long bone(s) in order to define the long axis of the bone and to act as a scale for digital image calibration.
A small marker was placed on the nylon adjacent to the nearest calibration scale, between the eyelet and the muscle stump; this enabled tracking of the motion of the suture nylon as the joint was moved through its range of motion. A 30-kg weight was tied to the free end of the suture nylon and allowed to hang over the side of the dissection table, thus ensuring that tension was maintained on the suture nylon during the measurement process. Tendon travel was determined by calibrating the displacement of the marker relative to the brass eyelet (Fig. 1).
Fig. 1.
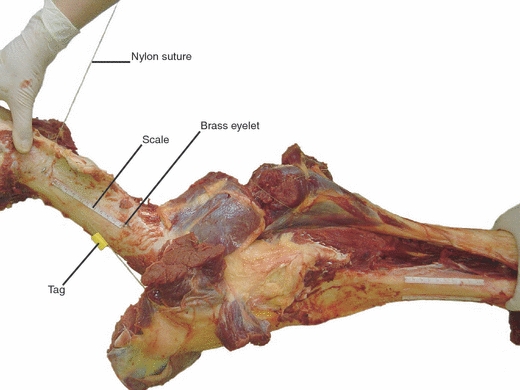
Image of the experimental set-up for the determination of tendon travel.
Joint angles were defined at the cranial aspect of the hip and tarsal joint and at the caudal aspect of the stifle joint, so that an extended limb angle would result in a joint angle of 180° and a flexed limb angle would be expressed as a lesser angle.
Recording the moment arm length changes
A digital camera (Nikon D70S) was positioned parallel to the plane of motion (ensuring that marker, eyelet and millimetre scale were visible) and pictures taken at approximately every 5–10º of joint motion whilst the joint was flexed, extended and flexed again through its movement range in the sagittal plane.
When muscles acted over more than one joint, the joint not of primary interest was held in a fixed, neutral position at mid-range whilst data were collected.
The measurements were made from the digital images using image analysis software (Image J; National Institute of Health, Bethesda, MD, USA). For each image a calibration was measured on the scale, the displacement of the marker relative to the brass eyelet was measured and the angle between the axes of the long bones was also calculated. This was repeated for each image at each joint angle. The data were exported into a spreadsheet (excel, Microsoft Office 2003) and the actual relative position of the marker was then determined from the calibration measurement.
Analysis of the moment arm data
Graphs of tendon excursion against joint angle (in radians) were plotted for each muscle. The muscle moment arm was defined as the derivative of the change in distance with respect to the change in angle and was therefore the gradient of the line of best fit. Linear and polynomial regression was used to model the relationship between tendon travel and joint angle. The Pearson product correlation coefficient (R2) was determined for the line of best fit in each case, such that some muscles showed a constant moment arm length which did not alter with joint angle (linear regression) whilst others were joint angle-dependent (second-order polynomial regression). When linear and polynomial regressions resulted in a similar goodness of fit and had similar R2 values, the linear fit was used if the moment arms calculated from the regression at minimum and maximum joint angles varied by < 10 mm. The resulting polynomial regression equations were subsequently differentiated to determine moment arm values (Hughes et al., 1998). Moment arms for maximum and minimum joint angles were determined from the derived regression equations along with the mean moment arm values for each breed of horse. Mean mid-stance moment arm values were also determined for joint angles that have been measured for the trailing hind limb during canter (hip 110°, stifle 145° and tarsus 152°; Back et al., 1997). Moment arm lengths were scaled by segment length (those acting at the hip and stifle to femur length and those at the tarsus to tibia length; Table 1) (Payne et al., 2006).
Muscle fascicle length to moment arm ratio
The ratio between the raw data obtained for muscle fascicle length and moment arm length was determined using breed fascicle length data previously reported for these two cohorts of horses (Crook et al., 2008). The ratio was scaled by segment length (those acting at the hip and stifle to femur length and those at the tarsus to tibia length; Table 1) to allow statistical comparison (Payne et al., 2006).
Calculation of mean moments of force
Maximum mean moments of force (M) for each muscle at mid-stance joint angles were determined using previously determined muscle physiological cross-sectional area data (Crook et al., 2008) and the following equation:
where σ is the maximum isometric stress of muscle (estimated at 0.3 MPa for vertebrate muscle (Wells, 1965), A is the muscle cross-sectional area and rmax is the maximum muscle moment arm.
Statistical analysis
Statistical analysis was used to compare the scaled moment arms of the two breeds between the functional ranges of joint angles of 80–115º at the hip, 120–180º at the stifle and 115–180º at the tarsus (Back et al., 1995). A two-sample t-test, at a significance level of P < 0.05, compared muscles with fixed moment arms, as moment arm lengths remained at a constant length throughout joint motion. A one-way anova, at a significance level of P < 0.05, compared moment arms of muscles that were joint angle-dependent and therefore changed with joint motion. A post-hoc Bonferroni test analysed moment arm length change and joint angle. Statistical analyses of the scaled ratio of muscle fascicle length to moment arm length and the maximum moments of force were undertaken using a two-sample t-test, at a significance level of P < 0.05.
Data were tabulated using Microsoft Office excel 2003 (Microsoft Corporation, Redmond, WA, USA). Statistical analysis was performed using spss, version 16 (SPSS Inc., Chicago, IL, USA).
Results
Moment arms were measured and scaled to limb segment length for eight hind limb muscles in each breed of horse. In effect, scaled measurements for 16 moment arm lengths were compared: five different lines of action were measured for the large, fan-shaped gluteus medius; individual moment arm lengths were obtained for each of the different heads of biceps femoris (three) and gastrocnemius (two) acting at the stifle; and moment arm lengths were also measured at two separate joints for the bi-articular muscles (biceps femoris, semitendinosus and gastrocnemius).
Tendon travel measured during joint motion varied between subjects. However, for most muscles the shape of the curve was similar in both breeds of horse (Fig. 2). Occasionally, it was not possible to determine moment arm data for each muscle in every horse (n = 5, gluteus medius (L3, L4 & L5 QH; L1, L2 & L3 Arab), semitendinosus at the Arab hip, biceps femoris (vertebral head) at the QH stifle, biceps femoris (intermediate and caudal head) at the Arab stifle. There were four biceps femoris (vertebral head) at the Arab stifle), owing to movement of the marker on the wire or poor images which failed to capture all the relevant markers and calibration scales. These errors only became apparent when analysing the images, by which time the limbs had already been disposed of, meaning it was not possible to repeat measurements.
Fig. 2.
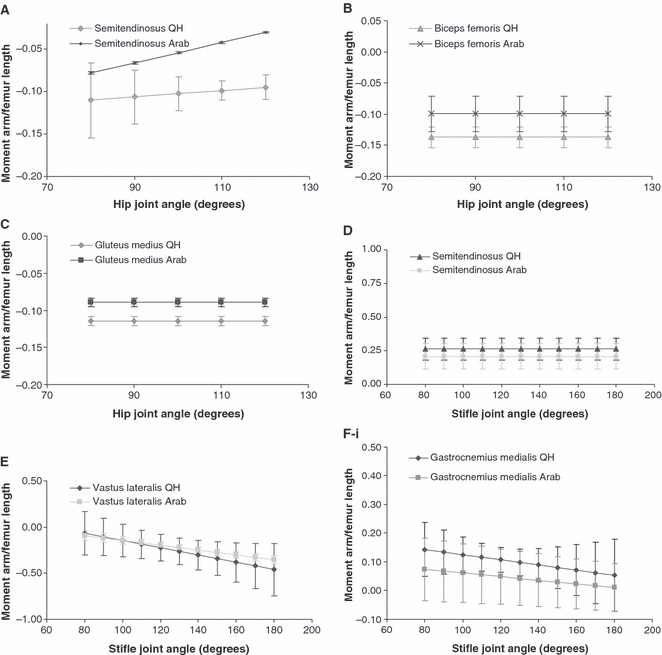
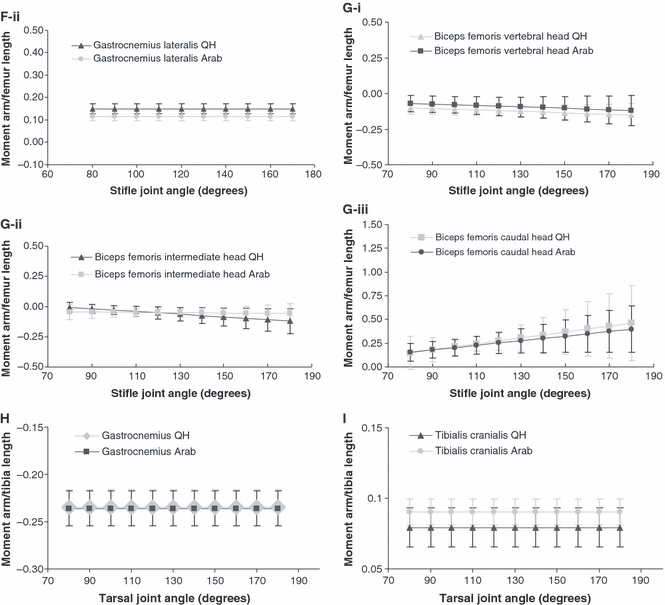
Mean moment arm curves (± SD) (scaled to femur or tibia length, see Methods described in text; mean femur length, 450 mm QH: 440 mm Arab, mean tibia length, 390 mm QH: 365 mm Arab) for muscles acting at the hip in the Quarter Horse and Arab: (A) mean moment arms curves (± SD) (scaled) for semitendinosus and (B) biceps femoris at the hip in the Quarter Horse (QH) and Arab. (C) Mean moment arms curves (± SD) (scaled) for the five lines of action of gluteus medius, lines one to five (L1–5) at the hip in the Quarter Horse (QH) and Arab. At the stifle in the Quarter Horse and Arab: (D) mean moment arms curves (± SD) (scaled) for semitendinosus, (E) vastus lateralis, (F-i) gastrocnemius medialis and (F-ii) gastrocnemius lateralis at the stifle in the Quarter Horse (QH) and Arab. Mean moment arms curves (± SD) (scaled) for biceps femoris: vertebral head (G-i), intermediate head (G-ii), and caudal head (G-iii) at the stifle in the Quarter Horse (QH) and Arab. At the tarsus in the Quarter Horse and Arab: (H) mean moment arms curves (scaled) for gastrocnemius and (i) tibialis cranialis at the tarsus in the Quarter Horse (QH) and Arab.
Regression equations, describing mean tendon travel with joint angle for Quarter Horse and Arab hind limb muscles, and the Pearson product-moment correlation coefficient (R2 value) determined for the line of best fit are shown in Table 2.
Table 2.
Mean regression equations for the plots of tendon travel (cm) against joint angle (radians) for named Quarter Horse and Arab hind limb muscles.
| Muscle | Joint | Quarter Horse | R2 | Arab | R2 |
|---|---|---|---|---|---|
| Gluteus medius L1 | Hip | y = −5.19 × + 16.44 | 0.85 | *y = −4.14 × + 17.39 | 0.76 |
| Gluteus medius L2 | Hip | y = −5.51 × + 17.47 | 0.89 | *y = −3.58 × + 14.91 | 0.82 |
| Gluteus medius L3 | Hip | *y = −4.74 × + 15.79 | 0.73 | *y = −3.98 × + 17.06 | 0.75 |
| Gluteus medius L4 | Hip | *y = −5.17 × + 17.30 | 0.82 | y = −4.16 × + 17.77 | 0.83 |
| Gluteus medius L5 | Hip | *y = −5.20 × + 20.22 | 0.76 | y = −3.81 × + 17.30 | 0.78 |
| Biceps femoris | Hip | y = −6.16 × + 21.03 | 0.89 | y = −4.38 × + 17.57 | 0.77 |
| Semitendinosus | Hip | y = 0.50 x2 − 6.36 × + 20.82 | 0.84 | *y = 1.50 x2 − 7.63 × + 19.67 | 0.81 |
| Biceps femoris (vertebral head) | Stifle | *y = −0.59 x2 − 2.99 × + 21.98 | 0.95 | †y = −0.61 x2 − 1.35 × + 20.37 | 0.89 |
| Biceps femoris (intermediate head) | Stifle | y = −1.48 x2 + 3.86 × + 8.83 | 0.79 | *y = −0.17 x2 − 1.44 × + 17.68 | 0.80 |
| Biceps femoris (caudal head) | Stifle | y = 4.03 x2 − 4.54 × + 8.57 | 0.95 | *y = 3.06 x2 − 1.76 × + 2.31 | 0.93 |
| Semitendinosus | Stifle | y = 11.82 × − 13.80 | 0.87 | y = 9.18 × − 6.95 | 0.89 |
| Vastus lateralis | Stifle | *y = −5.08 x2 + 11.15 × + 1.77 | 0.97 | y = −3.31 x2 + 5.20 × + 12.72 | 0.97 |
| Gastrocnemius lateralis | Stifle | y = 6.77 × − 3.88 | 0.97 | y = 5.06 × − 0.75 | 0.96 |
| Gastrocnemius medialis | Stifle | y = −0.43 x2 + 6.77 × − 1.81 | 0.88 | y = −0.78 x2 + 5.41 × − 0.23 | 0.92 |
| Gastrocnemius | Tarsus | y = −9.17 × + 32.85 | 0.99 | y = −8.61 × + 29.81 | 0.99 |
| Tibialis cranialis | Tarsus | y = 3.10 × + 1.33 | 0.94 | y = 3.30 × − 0.57 | 0.95 |
Equations and R2 values are means. n= 6 for all muscles in each breed except where indicated.
n = 5
n = 4: Gluteus medius L1–5 indicates the five lines of muscle pull determined for each muscle, L1 being most cranial, L5 most caudal.
Moment arms and joint torques of muscles acting at the hip joint
The mean moment arm values for biceps femoris at the hip joint were significantly greater (P = 0.041) in the Quarter Horse (62 ± 7 mm) than in the Arab (44 ± 12 mm) (Tables 3 and 5).
Table 3.
Minimum, maximum and mid-stance (± SD) moment arm values (mm) for hind limb muscles of the Quarter Horse and Arab for joint angles corresponding to those measured during the canter.
| Quarter Horse |
Arab |
||||||
|---|---|---|---|---|---|---|---|
| Muscle | Joint | Min (± SD) | Max (± SD) | Mid-stance (± SD) | Min (± SD) | Max (± SD) | Mid-stance (± SD) |
| Gluteus medius L1 | Hip | 52 ± 11 | 52 ± 11 | 52 ± 11 | 41 ± 20 | 41 ± 20 | 41 ± 20 |
| Gluteus medius L2 | Hip | 55 ± 14 | 55 ± 14 | 55 ± 14 | 36 ± 23 | 36 ± 23 | 36 ± 23 |
| Gluteus medius L3 | Hip | 47 ± 19 | 47 ± 19 | 47 ± 19 | 40 ± 15 | 40 ± 15 | 40 ± 15 |
| Gluteus medius L4 | Hip | 52 ± 20 | 52 ± 20 | 52 ± 20 | 42 ± 16 | 42 ± 16 | 42 ± 16 |
| Gluteus medius L5 | Hip | 52 ± 26 | 52 ± 26 | 52 ± 26 | 38 ± 12 | 38 ± 12 | 38 ± 12 |
| Biceps femoris | Hip | 62 ± 7 | 62 ± 7 | 62 ± 7 | 44 ± 12 | 44 ± 12 | 44 ± 12 |
| Semitendinosus | Hip | 44 ± 5 | 49 ± 18 | 44 ± 5 | 17 ± 25 | 32 ± 31 | 19 ± 25 |
| Biceps femoris (vertebral head) | Stifle | 55 ± 18 | 66 ± 33 | 60 ± 23 | 39 ± 28 | 51 ± 45 | 44 ± 34 |
| Biceps femoris (intermediate head) | Stifle | 24 ± 22 | 53 ± 45 | 36 ± 31 | 22 ± 12 | 25 ± 35 | 23 ± 18 |
| Biceps femoris (caudal head) | Stifle | 126 ± 47 | 204 ± 170 | 159 ± 95 | 113 ± 50 | 171 ± 105 | 137 ± 71 |
| Semitendinosus | Stifle | 118 ± 36 | 118 ± 36 | 118 ± 36 | 92 ± 42 | 92 ± 42 | 92 ± 42 |
| Vastus lateralis | Stifle | 105 ± 64 | 202 ± 122 | 146 ± 77 | 89 ± 14 | 153 ± 19 | 115 ± 13 |
| Gastrocnemius lateralis | Stifle | 68 ± 10 | 68 ± 10 | 68 ± 10 | 51 ± 7 | 51 ± 7 | 51 ± 7 |
| Gastrocnemius medialis | Stifle | 25 ± 53 | 47 ± 20 | 46 ± 29 | 6 ± 40 | 21 ± 42 | 15 ± 40 |
| Gastrocnemius | Tarsus | 92 ± 7 | 92 ± 7 | 92 ± 8 | 86 ± 7 | 86 ± 7 | 86 ± 7 |
| Tibialis cranialis | Tarsus | 31 ± 5 | 31 ± 5 | 31 ± 6 | 33 ± 3 | 33 ± 3 | 33 ± 3 |
The greater moment arm of biceps femoris at the hip in the Quarter Horse coupled with the greater PCSA of this muscle almost doubled the potential muscle moment of biceps femoris at the hip in the Quarter Horse (540 Nm) when compared to the Arab (280 Nm) (Table 4).
Table 4.
Mean muscle moment arm lengths (mm), maximum isometric force (kN) muscle moment (Nm) and MFL : MA ratios for named hind limb muscles in the Quarter Horse and Arab at joint angles corresponding to those measured for the mid-stance phase of the canter.
| Mean moment arm (mm) |
Isometric force (kN) |
Maximum muscle moment (Nm) |
MFL : MA ratio |
||||||
|---|---|---|---|---|---|---|---|---|---|
| Muscle | Joint | QH | Arab | QH | Arab | QH | Arab | QH | Arab |
| Gluteus medius L1 | Hip | 52 | 41 | 13.20 | 9.50 | 680 | 400 | 2.99 | 3.38 |
| Gluteus medius L2 | Hip | 55 | 36 | 13.20 | 9.50 | 730 | 340 | 2.81 | 3.91 |
| Gluteus medius L3 | Hip | 47 | 40 | 13.20 | 9.50 | 620 | 380 | 3.27 | 3.52 |
| Gluteus medius L4 | Hip | 52 | 42 | 13.20 | 9.50 | 680 | 400 | 3.00 | 3.37 |
| Gluteus medius L5 | Hip | 52 | 38 | 13.20 | 9.50 | 680 | 360 | 2.98 | 3.67 |
| Biceps femoris | Hip | 62 | 44 | 8.80 | 6.40 | 540 | 280 | 3.07 | 4.06 |
| Semitendinosus | Hip | 44 | 19 | 8.00 | 4.60 | 350 | 90 | 3.29 | 7.80 |
| Biceps femoris (vertebral head) | Stifle | 60 | 44 | 6.00 | 4.50 | 360 | 200 | 3.20 | 4.12 |
| Biceps femoris (intermediate head) | Stifle | 36 | 23 | 1.10 | 1.10 | 40 | 20 | 5.18 | 6.61 |
| Biceps femoris (caudal head) | Stifle | 159 | 137 | 1.80 | 0.90 | 280 | 120 | 1.19 | 1.45 |
| Semitendinosus | Stifle | 118 | 92 | 8.00 | 4.60 | 940 | 430 | 1.24 | 1.58 |
| Vastus lateralis | Stifle | 146 | 115 | 3.30 | 3.20 | 470 | 370 | 0.80 | 0.90 |
| Gastrocnemius lateralis | Stifle | 68 | 51 | 5.00 | 4.30 | 340 | 220 | 0.53 | 0.65 |
| Gastrocnemius medialis | Stifle | 46 | 15 | 3.00 | 2.20 | 140 | 30 | 1.02 | 3.01 |
| Gastrocnemius | Tarsus | 92 | 86 | 8.00 | 6.50 | 730 | 560 | 0.46 | 0.44 |
| Tibialis cranialis | Tarsus | 31 | 33 | 1.40 | 1.00 | 40 | 30 | 2.19 | 2.12 |
There was no significant difference in any of the gluteus medius scaled moment arms between the Quarter Horse and Arab (P = 0.476 L1, P = 0.154 L2, P = 0.595 L3, P = 0.442 L4, P = 0.360 L5); 52 ± 18 mm QH; 40 ± 18 mm Arab, mean for all lines of action). The Quarter Horses’ greater mean muscle PCSA resulted in them having the potential to generate a greater mean muscle moment at the hip joint (680 Nm) when compared to the Arab (380 Nm) (Table 4). The mean moment arm lengths of gluteus medius and biceps femoris remained at a constant length with changes in joint angle, which meant that their hip joint torques did not alter with variations in hip joint angle (Table 4).
The mean moment arm lengths of semitendinosus at the hip were similar in the Quarter Horse and Arab (P = 0.389), and increased linearly with hip joint flexion (Table 3), which meant the muscle was able to exert its maximum extensor muscle moment when the joint was in full flexion. Semitendinosus in both breeds of horse had the smallest calculated mean hip joint muscle moment of those muscles measured, at joint angles which correspond to the mid-stance phase of the canter (350 Nm QH; 90 Nm Arab) (Table 4).
Moment arms and joint torques of muscles acting at the stifle joint
Moment arm lengths were determined for each of the three heads of biceps femoris acting at the stifle and were similar in the Quarter Horse and Arab (P = 0.445 vertebral head, P = 0.194 intermediate head, P = 0.711 caudal head) (Table 5).
Table 5.
Statistical comparison of scaled hind limb muscle moment arm lengths, muscle fascicle length to moment arm ratios and maximum moments of force in the Quarter Horse and Arab.
| MA lengths |
MFL : MA |
MMF |
||
|---|---|---|---|---|
| Muscle | Joint | P value | P value | P value |
| Gluteus medius L1 | Hip | 0.476 | 0.363 | 0.728 |
| Gluteus medius L2 | Hip | 0.154 | 0.293 | 0.410 |
| Gluteus medius L3 | Hip | 0.595 | 0.925 | 0.618 |
| Gluteus medius L4 | Hip | 0.442 | 0.742 | 0.799 |
| Gluteus medius L5 | Hip | 0.360 | 0.698 | 0.445 |
| Biceps femoris | Hip | **0.041 | **0.047 | 0.592 |
| Semitendinosus | Hip | 0.389 | 0.261 | 0.196 |
| Biceps femoris (vertebral head) | Stifle | 0.445 | 0.357 | 0.375 |
| Biceps femoris (intermediate head) | Stifle | 0.194 | 0.521 | 0.427 |
| Biceps femoris (caudal head) | Stifle | 0.711 | 0.195 | 0.343 |
| Semitendinosus | Stifle | 0.343 | 0.272 | 0.865 |
| Vastus lateralis | Stifle | 0.532 | 0.713 | 0.945 |
| Gastrocnemius lateralis | Stifle | **0.003 | 0.549 | 0.751 |
| Gastrocnemius medialis | Stifle | 0.228 | 0.157 | 0.349 |
| Gastrocnemius | Tarsus | 1.00 | 1.00 | **0.041 |
| Tibialis cranialis | Tarsus | 0.073 | 0.804 | 0.090 |
MA lengths, muscle moment arm lengths; MFL : MA, muscle fascicle length to moment arm ratios; MMF, maximum moments of force.
Significance level set at P = 0.05.
Indicates a significant difference.
The caudal head of biceps femoris acting at the stifle had the greatest moment arm length of all the muscles measured (159 ± 95 mm QH; 137 ± 71 mm Arab). The mid-stance moment arm length was approximately two and a half times greater than that of the vertebral head and three and a half times that of the intermediate head; this led to a substantially increased muscle moment-generating capacity at the stifle, to 280 Nm in the Quarter Horse and 120 Nm in the Arab. All the Quarter Horse muscles had a greater mean muscle moment potential than the Arab, by virtue of their muscles greater mean muscle PCSA (Table 4).
The mid-stance moment arm lengths of the vertebral and intermediate heads of biceps femoris were of a similar length in both breeds of horse (vertebral head P = 0.445, 60 ± 23 mm QH and 44 ± 34 mm Arab; intermediate head P = 0.194, 36 ± 31 mm QH and 23 ± 18 mm Arab, Table 3). The larger mean PCSA of the vertebral head in both breeds enabled this head to generate the greatest mid-stance muscle moment of 360 Nm (QH) and 200 Nm (Arab) at the stifle joint. The intermediate head had the smallest mean mid-stance muscle moment potential (40 Nm in the Quarter Horse and 20 Nm in the Arab) (Table 4).
The moment arm lengths of the vertebral and intermediate heads increased with stifle flexion (vertebral head maximum value 66 ± 33 mm QH and 51 ± 45 mm Arab; intermediate head 53 ± 45 mm QH and 25 ± 35 mm Arab) and thus their moment-generating potential increased as the stifle flexed. The caudal head moment arm length increased with stifle extension to a maximum of 204 ± 170 mm (QH) and 171 ± 105 mm (Arab) (Table 3).
Vastus lateralis mid-stance moment arm lengths were similar in the Quarter Horse (146 ± 77 mm) and Arab (115 ± 13 mm) (P = 0.532) and were the second longest of the mid-stance moment arms measured at the stifle. The Quarter Horse mean mid-stance muscle moment was greater (470 Nm) than that calculated for the Arab (370 Nm). In both breeds the muscles moment-generating capacity increased with stifle flexion, owing to lengthening of their respective moment arms (maximum value 202 ± 122 mm in the QH and 153 ± 19 mm in the Arab) (Table 3).
The moment arm lengths of semitendinosus at the stifle joint were similar (118 ± 36 mm QH; 92 ± 42 mm Arab; P = 0.343) and remained at a constant length regardless of changes in joint angle. The relatively long moment arm length substantially increased the muscle’s moment-generating potential at the stifle. However, the Quarter Horses’ greater mean muscle PCSA provided them with almost double the mean muscle moment potential of that of the Arabs at all stifle joint angles (940 Nm QH; 430 Nm Arab; Table 4).
The moment arms values for the two heads of gastrocnemius acting at the stifle were determined separately. The mid-stance moment arm of gastrocnemius medialis was a similar length in the Quarter Horse (38 ± 29 mm) and Arab (15 ± 40 mm) (P = 0.228) and was the shortest of the muscles that were measured acting at the stifle. However, its moment arm length increased in both breeds as the stifle flexed, to a maximum of 47 ± 20 mm in the Quarter Horse and 21 ± 42 mm in the Arab. Its mid-stance mean muscle moment-generating values were greater in the Quarter Horse (at 140 Nm) than in the Arab (30 Nm), owing to the greater muscle PCSA (Crook et al., 2008).
The mean mid-stance moment arm length of gastrocnemius lateralis at the stifle was significantly longer in the Quarter Horse, at 68 ± 10 mm, than in the Arab, at 51 ± 7 mm (P = 0.003, Table 5). The moment arm lengths in both breeds remained at the same length during stifle flexion and extension. The mid-stance muscle moment potential of gastrocnemius lateralis in both breeds was almost double that of the medial head, at 340 Nm in the Quarter Horse and 220 Nm in the Arab (Table 4).
Moment arms and joint torques of muscles acting at the tarsus
The mid-stance moment arm of the combined heads of gastrocnemius acting at the tarsus was similar in the Quarter Horse and Arab (92 ± 8 mm QH; 86 + 7 mm Arab; P = 1.00). It was approximately double the length of the measured hip muscle moment arms and almost three times greater than that measured in tibialis cranialis.
The greater PCSA of gastrocnemius in the Quarter Horse resulted in a greater mean mid-stance torque-generating potential (730 Nm) when compared to that of the Arab (560 Nm); this value remained unchanged as the tarsus flexed and extended because moment arm length remained the same.
The moment arm of tibialis cranialis remained at a constant length and was similar in both breeds (31 ± 6 mm QH; 33 ± 3 mm Arab). This muscle had the smallest mean torque-generating potential of all the muscles measured (40 Nm in the Quarter Horse and 30 Nm in the Arab).
Maximum moments of force
All the Quarter Horses’ muscles by virtue of their greater PCSA (Crook et al., 2008) were able to generate greater mean maximum moments of force. However, this was only statistically significant for gastrocnemius acting at the tarsus (P = 0.041), but for none of the other muscles: gluteus medius P = 0.728 L1, P = 0.410 L2, P = 0.618 L3, P = 0.799 L4, P = 0.445 L5; biceps femoris and semitendinosus acting at the hip P = 0.592 and 0.196, respectively; biceps femoris acting at the stifle P = 0.375 (vertebral head), P = 0.427 (intermediate head), P = 0.343 (caudal head); semitendinosus acting at the stifle P = 0.865; vastus lateralis P = 0.945; gastrocnemius lateralis and medialis P = 0.751 and 0.349, respectively; tibialis cranialis P = 0.090 (Table 5).
Muscle fascicle length to moment arm ratio (MFL : MA Ratio)
With the exception of gastrocnemius and tibialis cranialis at the tarsus, it was observed that all the Arab muscles had a greater mean MFL : MA ratio when compared to those of the Quarter Horse (Crook et al., 2008) (Table 4). However, this observed difference was only significant for biceps femoris acting at the hip (P = 0.047), but for none of the remaining muscles: gluteus medius P = 0.363 L1, P = 0.293 L2, P = 0.925 L3, P = 0.742 L4, P = 0.698 L5; semitendinosus acting at the hip P = 0.261; biceps femoris acting at the stifle P = 0.357 (vertebral head), P = 0.521 (intermediate head), P = 0.195 (caudal head); semitendinosus acting at the stifle P = 0.272; vastus lateralis P = 0.713; gastrocnemius lateralis and medialis P = 0.549 and 0.157, respectively; and tibialis cranialis P = 0.0804 (Table 5).
Discussion
Selective breeding for athletic performance has resulted in distinct breeds of horse, such as the Quarter Horse (bred for acceleration) and the Arab (bred for endurance). We compared hind limb muscle moment arms in these two breeds to gain important insights into how equine limb muscle anatomy might be adapted for different athletic requirements. We hypothesized that the Quarter Horse hind limb extensor muscles would have a greater length of moment arm when compared to those of the Arab, conferring on them greater muscle moment-generating capacity at the hip, stifle and tarsus during limb extension and providing their muscles with the potential to generate superior power outputs required during rapid acceleration.
Although there was a trend towards the Quarter Horse muscles having a greater length of moment arm, our findings fail to support our hypothesis. The only two muscles to be of a significantly greater length in the Quarter Horse were biceps femoris (P = 0.041) acting at the hip, and gastrocnemius lateralis (P = 0.003) acting at the stifle. All the remaining moment arms were of a similar length throughout the functional joint range of motion (Tables 3 and 5). It appears that biceps femoris and gastrocnemius lateralis in the Quarter Horse may have adapted to play an important role during rapid acceleration, and it is recommended that further studies of their electromyographic activation patterns during rapid acceleration be undertaken to help clarify their function.
The greater length of moment arm combined with the greater PCSA (and therefore greater isometric force potential) for both of the Quarter Horse muscles mean that they have the potential to generate almost double the muscle moments at their respective joints when compared to those of the Arab (Table 4). Large joint torques, particularly in large-bodied animals, are a prerequisite for high power outputs and hence rapid acceleration.
The estimated mean maximum muscle moment potential of all the Quarter Horse muscles were larger than those of the Arab (Table 4), although this was only significant for gastrocnemius at the tarsus (P = 0.041) (Table 5), which is possibly a reflection of the relatively small sample size used in the study. The Quarter Horses’ tendency for greater maximum moment potential resulted from their greater muscle volume and PCSA (both breeds having similar fascicle length; Crook et al., 2008), rather than greater moment arm length (Table 3). Our findings are in agreement with other researchers who have found muscle volume and PCSA to be the major determinants of joint torque in humans (Fukunaga et al., 2001), and suggest that muscle volume rather than moment arm length is more readily influenced through selective breeding and strength training regimes.
Gastrocnemius acting at the tarsus had the greatest torque potential of all the hind limb muscles (730 Nm QH; 560 Nm Arab; Table 4), in common with other cursorial animals such as the ostrich (Smith et al., 2007). The muscles’ relatively long moment arm length (92 mm QH; 86 mm Arab; Table 4) and small MFL : MA ratio (0.46 QH; 0.44 Arab; Table 4) were almost identical in both breeds of horse, although the Quarter Horse muscles were able to generate significantly greater moments of force when compared to those of the Arab (P = 0.041, Table 5). As such, the muscle tendon unit is less suited to moving the tarsus through large joint excursions when working in isolation (perhaps ground reaction force and tendon elongation are enough to do this (Williams et al., 2007), but confirms its function as an elastic storage mechanism (Biewener, 1998). By storing and returning elastic strain energy throughout the gait cycle, the muscle tendon unit reduces the energetic cost of locomotion in both breeds of horse (Dimery et al., 1986). In addition, being bi-articular, it links stifle and tarsus movement, and may modulate power flow between these two joints.
The majority of the hind limb muscles had moment arm lengths that varied with joint position (Table 3), illustrating the importance of using the tendon travel method, which enabled us to track these length changes. In both breeds, the moment arm of vastus lateralis was capable of undergoing the greatest length change, its torque potential at maximum stifle flexion being almost twice that of when the stifle is near full extension (Table 3). All of the bi-articular muscles (biceps femoris, semitendinosus and gastrocnemius) had either a proximal or distal moment arm that varied with joint position (Tables 2 and 3). By altering moment arm length, these muscles are capable of altering their effective mechanical advantage (EMA) and function throughout the gait cycle (Biewener, 1989), either increasing or decreasing joint torques as required. This ability to alter moment arm length, and therefore muscle EMA, has been observed in a wide variety of mammalian species and has been termed ‘dynamic gearing’ (Carrier et al., 1998; Williams et al., 2007). It is thought that dynamic gearing maximizes the efficiency of muscle work, allowing the muscle to operate at the optimum range of its length–tension relationship (Carrier et al., 1998).
Synergistic muscles had differing moment arm lengths and different gear ratios at their joint of action in both the Quarter Horse and the Arab. Electromyography suggests that synergistic muscles with differing gear ratios, such as gluteus medius, biceps femoris and semitendinosus (Robert et al., 1999), may be involved in a ‘power sharing mechanism’ (Dutto et al., 2004), where each muscle is active at different stages in the gait cycle. Similar to dynamic gearing, power sharing allows each muscle within the group to work in an optimum length–tension relationship, thus optimizing the efficiency of the muscles’ individual function within its synergistic group (Dutto et al., 2004). It is likely that a similar mechanism occurs at other joints (for example, the stifle), allowing power sharing between the different heads of biceps femoris, semitendinosus and medial and lateral gastrocnemius during stifle flexion.
It is interesting that different heads of the same muscle often had differing moment arm lengths and torque potential at same joint of action (e.g. biceps femoris and lateral gastrocnemius, Table 4). This may indicate that different parts of these muscles have a different function during equine locomotion and may be able to operate at the optimum moment of the length–tension relationship to fulfil this function; further research is required to explore if this is the case.
With the exception of lateral gastrocnemius and tibialis cranialis, the observed MFL : MA ratios were greater for the Arab muscles than for the Quarter Horse muscles (even though the difference was only significant for biceps femoris acting at the hip, P = 0.047). This resulted from the fact that, even though both groups of horses had similar muscle fascicle lengths (Crook et al., 2008), the Quarter Horse muscles tended to have a greater length of moment arm (despite this being only statistically significant for biceps femoris and lateral gastrocnemius). The tendency for the Arabs’ muscles to have a higher MFL : MA ratio when compared to those of the Quarter Horse, suggests that, for a similar change in joint angle, the Arab muscle fibres undergo a proportionally smaller length change. This means that at the same angular velocity the Arabs’ muscle sarcomeres operate at a slower velocity of contraction and have a greater force potential (operating earlier in the force-velocity curve).
The tendency for shorter muscle moment arms in the Arab coupled with their tendency for a greater MFL : MA ratio, suggest that the Arabs muscles are optimized to function closer to maximum economy than to maximum power output.
Observed differences in muscle fibre type between the two breeds also support this assumption. The Arab muscles possess a greater percentage of type I muscle fibres when compared to those of the Quarter Horse (Snow & Guy, 1980). This allows their muscles to operate at lower velocity of contraction (Rome et al., 1990) and optimizes their muscles for endurance events (Rivero et al., 1993). In addition, recent research has demonstrated that shorter muscle moment arms in humans are linked to greater running economy (Scholz et al., 2008).
Experimental assumptions
The joint angles used in this study were taken from studies of the kinematics of Dutch Warm Blood horses on a treadmill (Back et al., 1995, 1997), as to date very few data exist regarding joint angle kinematics for the Quarter Horse or Arab. It is appreciated that horses adopt a differing gait pattern on a treadmill when compared to locomotion over ground (Barrey et al., 1993) and that limb kinematics (Cano et al., 2001; Galisteo et al., 2001) and kinetics (Back et al., 2007) are breed-specific. However, once kinetic and kinematic data have been determined for the galloping Quarter Horse and Arab, our moment arm data will allow the specific determination of muscle moments and EMAs in these two breeds.
Moment arm lengths were determined using the tendon travel method. The advantage of this method is that it is dynamic and does not assume a fixed centre of joint rotation (Brown et al., 2003a), and enables the estimation of changes of moment arm length with differences in joint position (Brown et al., 2003a; Payne et al., 2006; Williams et al., 2007). We found that several of the equine hind limb moment arms were joint angle-dependent, similar to those of the equine distal forelimb (Brown et al., 2003a) and those of other species (Payne et al., 2006; Smith et al., 2007; Williams et al., 2007). This changing moment arm length allows the muscles to function differently with different limb positions and may reflect the muscles’ ability to generate a large moment if its moment arm is longer at that point in the gait cycle.
The potential disadvantage of the tendon travel method assumes joint motion occurs in a sagittal plane only. Whilst motion at the equine hip is restricted primarily to flexion and extension by the presence of the accessory ligament (peculiar to the equine and the donkey), movements at the stifle and tarsus in domestic species are accompanied by a degree of abduction, adduction and rotation (Badoux, 1984; Lanovaz et al., 2002). However, these movements are relatively small in comparison, with flexion and extension being the predominant movement.
Moment arm length changes in feline bi-articular muscles are dependent on the relative positions of the two joints that they act upon (Macfadden & Brown, 2007). The methodology employed in this study meant that moment arms were determined individually at each joint, with the proximal and distal joints stabilized at mid-range. It may have useful to investigate the interaction of the two joints of action for bi-articular muscles; however, time constraints meant that this was beyond the scope of the present study.
The relatively small sample size used in this study together with some of the reported standard deviations in the dataset possibly explain why statistical differences in moment arm length, muscle fascicle length : moment arm ratio and maximum muscle moment potential were only significant for a few of the muscles. This study should be viewed as a preliminary insight into equine breed differences in muscle moment arms and it is hoped that future studies will be undertaken to substantiate our findings.
Conclusion
Although there was a trend towards the Quarter Horse muscles having a greater length of moment arm, the majority of the muscles studied were of a similar length. The only two muscles to have significantly longer moment arms in the Quarter Horse were those of biceps femoris (P = 0.041) acting at the hip, and gastrocnemius lateralis (P = 0.003) acting at the stifle. Quarter Horse hind limb muscles are better suited to generating the greater joint torques associated with rapid acceleration by virtue of their greater muscle PCSA and thus greater isometric force potential, rather than by having greater length of muscle moment arm. Arab muscles at the hip and stifle tend to have a greater MFL : MA ratio, which indicates that their muscles have the potential to operate at slower velocities of contraction and therefore generate greater force outputs, optimizing their muscles for economical locomotion during endurance racing.
References
- An KN, Hui FC, Morrey BF, et al. Muscles across the elbow joint: a biomechanical analysis. J Biomech. 1981;14:659–669. doi: 10.1016/0021-9290(81)90048-8. [DOI] [PubMed] [Google Scholar]
- Back W, Schamhardt HC, Savelberg HH, et al. How the horse moves: 2. Significance of graphical representations of equine hind limb kinematics. Equine Vet J. 1995;27:39–45. doi: 10.1111/j.2042-3306.1995.tb03030.x. [DOI] [PubMed] [Google Scholar]
- Back W, Schamhardt HC, Barneveld A. Kinematic comparison of the leading and trailing fore- and hindlimbs at the canter. Equine Vet J. 1997;(Suppl 23):80–83. doi: 10.1111/j.2042-3306.1997.tb05060.x. [DOI] [PubMed] [Google Scholar]
- Back W, Macallister CG, Heel MCVV, et al. Vertical frontlimb ground reaction forces of sound and lame warmbloods differ from those in quarter horses. J Equine Vet Sci. 2007;27:123–129. [Google Scholar]
- Badoux DM. The geometry of the cruciate ligaments in the canine and equine knee joint, a Tchebychev mechanism. Acta Anat (Basel) 1984;119:60–64. doi: 10.1159/000145862. [DOI] [PubMed] [Google Scholar]
- Barrey E, Galloux P, Valette JP, et al. Stride characteristics of overground versus treadmill locomotion in the saddle horse. Acta Anat (Basel) 1993;146:90–94. doi: 10.1159/000147427. [DOI] [PubMed] [Google Scholar]
- Biewener AA. Scaling body support in mammals: limb posture and muscle mechanics. Science. 1989;245:45–48. doi: 10.1126/science.2740914. [DOI] [PubMed] [Google Scholar]
- Biewener AA. Muscle-tendon stresses and elastic energy storage during locomotion in the horse. Comp Biochem Physiol B Biochem Mol Biol. 1998;120:73–87. doi: 10.1016/s0305-0491(98)00024-8. [DOI] [PubMed] [Google Scholar]
- Brown NA, Pandy MG, Buford WL, et al. Moment arms about the carpal and metacarpophalangeal joints for flexor and extensor muscles in equine forelimbs. Am J Vet Res. 2003a;64:351–357. doi: 10.2460/ajvr.2003.64.351. [DOI] [PubMed] [Google Scholar]
- Brown NA, Pandy MG, Kawcak CE, et al. Force- and moment-generating capacities of muscles in the distal forelimb of the horse. J Anat. 2003b;203:101–113. doi: 10.1046/j.1469-7580.2003.00206.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cano MR, Vivo J, Miro F, et al. Kinematic characteristics of Andalusian, Arabian and Anglo-Arabian horses: a comparative study. Res Vet Sci. 2001;71:147–153. doi: 10.1053/rvsc.2001.0504. [DOI] [PubMed] [Google Scholar]
- Carrier DR, Gregersen CS, Silverton NA. Dynamic gearing in running dogs. J Exp Biol. 1998;201:3185–3195. doi: 10.1242/jeb.201.23.3185. [DOI] [PubMed] [Google Scholar]
- Crook TC, Cruickshank SE, Mcgowan CM, et al. Comparative anatomy and muscle architecture of selected hind limb muscles in the Quarter Horse and Arab. J Anat. 2008;212:144–152. doi: 10.1111/j.1469-7580.2007.00848.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dimery NJ, Alexander RM, Ker RF. Elastic extension of leg tendons in the locomotion of horses (Equus caballus) J Zool. 1986;210:415–425. [Google Scholar]
- Dutto DJ, Hoyt DF, Clayton HM, et al. Moments and power generated by the horse (Equus caballus) hind limb during jumping. J Exp Biol. 2004;207:667–674. doi: 10.1242/jeb.00808. [DOI] [PubMed] [Google Scholar]
- Fukunaga T, Miyatani M, Tachi M, et al. Muscle volume is a major determinant of joint torque in humans. Acta Physiol Scand. 2001;172:249–255. doi: 10.1046/j.1365-201x.2001.00867.x. [DOI] [PubMed] [Google Scholar]
- Galisteo AM, Morales JL, Cano MR, et al. Inter-breed differences in equine forelimb kinematics at the walk. J Vet Med A Physiol Pathol Clin Med. 2001;48:277–285. doi: 10.1046/j.1439-0442.2001.00344.x. [DOI] [PubMed] [Google Scholar]
- Hoffman RM, Hess TM, Williams CA, et al. Speed associated with plasma pH, oxygen content, total protein and urea in an 80 km race. Equine Vet J. 2002;(Suppl 34):39–43. doi: 10.1111/j.2042-3306.2002.tb05389.x. [DOI] [PubMed] [Google Scholar]
- Hughes RE, Niebur G, Liu J, et al. Comparison of two methods for computing abduction moment arms of the rotator cuff. J Biomech. 1998;31:157–160. doi: 10.1016/s0021-9290(97)00113-9. [DOI] [PubMed] [Google Scholar]
- Lanovaz JL, Khumsap S, Clayton HM, et al. Three-dimensional kinematics of the tarsal joint at the trot. Equine Vet J. 2002;(Suppl 34):308–313. doi: 10.1111/j.2042-3306.2002.tb05438.x. [DOI] [PubMed] [Google Scholar]
- Macfadden LN, Brown NA. Biarticular hip extensor and knee flexor muscle moment arms of the feline hindlimb. J Biomech. 2007;40:3448–3457. doi: 10.1016/j.jbiomech.2007.05.021. [DOI] [PubMed] [Google Scholar]
- Merkens HW, Schamhardt HC, Osch Van GJ, et al. Ground reaction force patterns of Dutch Warmbloods at the canter. Am J Vet Res. 1993;54:670–674. [PubMed] [Google Scholar]
- Nagano A, Komura T. Longer moment arm results in smaller joint moment development, power and work outputs in fast motions. J Biomech. 2003;36:1675–1681. doi: 10.1016/s0021-9290(03)00171-4. [DOI] [PubMed] [Google Scholar]
- Nielsen BD, Turner KK, Ventura BA, et al. Racing speeds of quarter horses, thoroughbreds and Arabians. Equine Vet J Suppl. 2006;(Suppl 36):128–132. doi: 10.1111/j.2042-3306.2006.tb05528.x. [DOI] [PubMed] [Google Scholar]
- Payne RC, Crompton RH, Isler K, et al. Morphological analysis of the hindlimb in apes and humans. II. Moment arms. J Anat. 2006;208:725–742. doi: 10.1111/j.1469-7580.2006.00564.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivero JL, Serrano AL, Henckel P, et al. Muscle fiber type composition and fiber size in successfully and unsuccessfully endurance-raced horses. J Appl Physiol. 1993;75:1758–1766. doi: 10.1152/jappl.1993.75.4.1758. [DOI] [PubMed] [Google Scholar]
- Robert C, Valette JP, Degueurce C, et al. Correlation between surface electromyography and kinematics of the hindlimb of horses at trot on a treadmill. Cells Tissues Organs. 1999;165:113–122. doi: 10.1159/000016681. [DOI] [PubMed] [Google Scholar]
- Rome LC, Sosnicki AA, Goble DO. Maximum velocity of shortening of three fibre types from horse soleus muscle: implications for scaling with body size. J Physiol. 1990;431:173–185. doi: 10.1113/jphysiol.1990.sp018325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scholz MN, Bobbert MF, Soest Van AJ, et al. Running biomechanics: shorter heels, better economy. J Exp Biol. 2008;211:3266–3271. doi: 10.1242/jeb.018812. [DOI] [PubMed] [Google Scholar]
- Smith NC, Payne RC, Jespers KJ, et al. Muscle moment arms of pelvic limb muscles of the ostrich (Struthio camelus) J Anat. 2007;211:313–324. doi: 10.1111/j.1469-7580.2007.00762.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snow DH, Guy PS. Muscle fibre type composition of a number of limb muscles in different types of horse. Res Vet Sci. 1980;28:137–144. [PubMed] [Google Scholar]
- Spoor CW, Leeuwen Van JL. Knee muscle moment arms from MRI and from tendon travel. J Biomech. 1992;25:201–206. doi: 10.1016/0021-9290(92)90276-7. [DOI] [PubMed] [Google Scholar]
- UAE Equestrian & Racing Federation Xandu Haji Buba. 2006. CEI* 160 Km, Euston Park, UK. F\UAE EQUESTRIAN & RACING FEDERATION.htm.
- Wells JB. Comparison of mechanical properties between slow and fast mammalian muscles. J Physiol. 1965;178:252–269. doi: 10.1113/jphysiol.1965.sp007626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams SB, Payne RC, Wilson AM. Functional specialisation of the pelvic limb of the hare (Lepus europeus) J Anat. 2007;210:472–490. doi: 10.1111/j.1469-7580.2007.00704.x. [DOI] [PMC free article] [PubMed] [Google Scholar]