

Abstract
In vertebrate retina, melatonin regulates various physiological functions. In this work we investigated the mechanisms underlying melatonin-induced potentiation of glycine currents in rat retinal ganglion cells (RGCs). Immunofluorescence double labelling showed that rat RGCs were solely immunoreactive to melatonin MT2 receptors. Melatonin potentiated glycine currents of RGCs, which was reversed by the MT2 receptor antagonist 4-P-PDOT. The melatonin effect was blocked by intracellular dialysis of GDP-β-S. Either preincubation with pertussis toxin or application of the phosphatidylcholine (PC)-specific phospholipase C (PLC) inhibitor D609, but not the phosphatidylinositol (PI)-PLC inhibitor U73122, blocked the melatonin effect. The protein kinase C (PKC) activator PMA potentiated the glycine currents and in the presence of PMA melatonin failed to cause further potentiation of the currents, whereas application of the PKC inhibitor bisindolylmaleimide IV abolished the melatonin-induced potentiation. The melatonin effect persisted when [Ca2+]i was chelated by BAPTA, and melatonin induced no increase in [Ca2+]i. Neither cAMP-PKA nor cGMP-PKG signalling pathways seemed to be involved because 8-Br-cAMP or 8-Br-cGMP failed to cause potentiation of the glycine currents and both the PKA inhibitor H-89 and the PKG inhibitor KT5823 did not block the melatonin-induced potentiation. In consequence, a distinct PC-PLC/PKC signalling pathway, following the activation of Gi/o-coupled MT2 receptors, is most likely responsible for the melatonin-induced potentiation of glycine currents of rat RGCs. Furthermore, in rat retinal slices melatonin potentiated light-evoked glycine receptor-mediated inhibitory postsynaptic currents in RGCs. These results suggest that melatonin, being at higher levels at night, may help animals to detect positive or negative contrast in night vision by modulating inhibitory signals largely mediated by glycinergic amacrine cells in the inner retina.
Introduction
Melatonin is known to regulate various physiological functions by activating specific receptors, namely MT1, MT2 and MT3 subtypes (Vanecek, 1998; Dubocovich et al. 2003; Barrenetxe et al. 2004). In the vertebrate retina melatonin is synthesized and released by photoreceptors and may be implicated in retinomotor responses, rod disc shedding, phagocytosis, etc. (Cahill & Besharse, 1992; Vanecek, 1998; Tosini & Fukuhara, 2003). It has been recently shown that melatonin plays neuromodulatory roles in the retina, like those observed in suprachiasmatic nucleus (SCN) (McArthur et al. 1997; Gerdin et al. 2004). In the outer retina, melatonin modulates the function of cone-driven horizontal cells in teleost fish directly by altering AMPA receptor-mediated currents of these cells through the activation of MT1 receptors (Huang et al. 2005) and/or influencing dopamine release from interplexiform and amacrine cells (Ribelayga et al. 2004). Most recently, it is reported that melatonin potentiates rod signals to ON-type bipolar cells in carp by modulating the activity of the metabotropic glutamate receptor expressed on these cells through the activation of MT2 receptors (Ping et al. 2008). In the inner retina, retinal ganglion cells (RGCs), which are output neurons in the retina, receive inhibitory GABAergic and glycinergic inputs from amacrine cells, which shape their responses driven by excitatory input from bipolar cells (Protti et al. 1997; Slaughter, 2004). All three subtypes of melatonin receptors are found in RGCs in a variety of species (Fujieda et al. 1999; Wiechmann, 2003; Wiechmann et al. 2004; Alarma-Estrany & Pintor, 2007), suggesting that melatonin may modulate the activity of these cells. Indeed, previous work in this laboratory demonstrated that melatonin potentiates glycine receptor-mediated currents in rat RGCs by activating MT2 receptors (Zhang et al. 2007). However, the mechanisms underlying this melatonin effect are still unclear.
It has been generally thought that activation of MT2 receptors may regulate several signalling pathways, including the cAMP–PKA pathway (Vanecek, 1998; Alarma-Estrany & Pintor, 2007) and cGMP–PKG pathway (Petit et al. 1999). Accumulating evidence also suggests that, following activation of MT2 receptors, phospholipase C (PLC)-dependent pathways could come into play (McArthur et al. 1997; Lai et al. 2002; MacKenzie et al. 2002). In the present work, we first show that MT2 receptors are expressed in isolated rat RGCs by immunocytochemistry, and demonstrate that melatonin potentiates glycine currents of RGCs by activating pertussis toxin (PTX)-sensitive Gi/o coupled MT2 receptors. Downstream of the activation of G-proteins there is stimulation of a distinct intracellular phosphatidylcholine (PC)-specific PLC, followed by a Ca2+-independent increase of the activity of protein kinase C (PKC). Consistent with this, we show that melatonin potentiates light-evoked glycine receptor-mediated inhibitory postsynaptic currents (IPSCs) of RGCs in rat retinal slices, which is reversed by 4-P-PDOT.
Methods
All experimental procedures described in the present work were in accordance with the National Institutes of Health (NIH) Guidelines for the Care and Use of Laboratory Animals, the guidelines of Fudan University on the ethical use of animals, The Journal of Physiology's policy and UK regulations on animal experimentation (Drummond, 2009). During this study all efforts were made to minimize the number of animals used and their suffering. In the present work, a total of 78 rats were used, among which were 52 for dissociated cell electrophysiological recordings; 8 for electrophysiological recordings in retinal slices; 6 for calcium imaging; and 12 for immunocytochemistry and Western blots. A detailed description of anaesthetic regime follows.
Retrograde labelling of RGCs
Retrograde labelling of RGCs was previously described in detail (Chen et al. 2004). Briefly, after male albino rats (Sprague–Dawley, 100–150 g) were deeply anaesthetized with 40 mg ml−1 sodium pentobarbital (0.1 ml (100 g)−1), 2% aminostilbamidine methanesulphonate (fluorogold, FG; Molecular Probes, Eugene, OR, USA) was injected into the superior colliculus bilaterally (6.0 mm posterior and 2.0 mm lateral to the bregma and 4–5 mm deep from the cortical surface). After a survival period of 5–7 days, RGCs were clearly labelled for immunocytochemical analysis. For electrophysiology, rhodamine-B-isothiocyanate (RITC) was instead used to label RGCs under the same protocol as FG, because of the less toxic effects of this dye.
Preparation of isolated RGCs
RGCs were acutely dissociated from retinas retrogradely labelled with RITC or FG by enzymatic and mechanical methods as previously described (Chen et al. 2004) with minor modifications. Animals were deeply anaesthetized with 4% sodium pentobarbital and killed by decapitation. Retinas were removed quickly and incubated in oxygenated Hanks’ solution containing the following (in mm): NaCl 137, NaHCO3 0.5, NaH2PO4 1, KCl 3, CaCl2 2, MgSO4 1, Hepes 20, sodium pyruvate 1 and glucose 16 adjusted to pH 7.4 with NaOH. The retinas were then digested in 1.6 U ml−1 papain (Worthington Biochemical, Freehold, NJ, USA) containing Hanks’ solution, supplemented with 0.2 mg ml−1l-cysteine and 0.2 mg ml−1 bovine serum albumin for 30 min at 35.5–36.5°C. The solution was bubbled continuously with 100% O2 and adjusted pH to 7.4 with NaOH. After several rinses in Hanks’ solution, the retinas were mechanically dissociated with fire-polished Pasteur pipettes and cell suspension was plated onto a culture dish mounted on an inverted microscope (IX 70; Olympus Optical, Tokyo, Japan). RITC-labelled RGCs, showing red fluorescence, were chosen for whole-cell patch-clamp recording within 2–3 h after dissociation.
Western blot analysis and immunocytochemistry
Rat retinal extracts were prepared following the procedure described in detail previously (Chen et al. 2004) with minor modifications. The extract samples (2.0 mg ml−1, 20 μl) were loaded, subjected to 12% SDS-PAGE and electro-blotted onto polyvinylidene fluoride (PVDF) membranes using the Mini-PROTEAN 3 Electrophoresis System and the Mini Trans-Blot Electrophoretic Transfer System (Bio-Rad, Hercules, CA, USA). The membranes were blocked with skimmed milk at room temperature for 2 h, and then incubated with the goat polyclonal antibody against MT1 or MT2, respectively (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at working dilution of 1:500 (for MT1) or 1:200 (for MT2) overnight at 4°C. The blots were washed and incubated with horseradish peroxidase-conjugated donkey anti-goat IgG (1:5000; Santa Cruz Biotechnology) for 2 h at 4°C, and visualized with enhanced chemiluminescence (Amersham Biosciences, Arlington Heights, IL, USA).
Immunocytochemistry of isolated retinal cells refers to Yu et al. (2006) for detailed description. Isolated RGCs, labelled by FG, were placed on a slide in phosphate-buffered saline (PBS) for 30–60 min at room temperature and fixed with 4% paraformaldehyde in 0.1 m fresh phosphate buffer (PB) for 30 min, rinsed with PBS three times. They were then blocked for 1 h in PBS with 6% donkey serum plus 0.2% Triton X-100, followed by incubating with the anti-MT1 or -MT2 primary antibody at working dilution of 1:500 or 1:200, respectively, for 2 h and further incubated with the secondary antibody rhodamine red X (RRX)-conjugated donkey anti-goat IgG (Jackson ImmunoResearch Laboratories, West Grove, PA, USA) for 30 min at room temperature. The cells were visualized with two-photon laser scanning microscope (Zeiss, Oberkochen, Germany) using a 100× oil-immersion objective lens. To avoid any possible reconstruction stacking artifact, double labelling was precisely evaluated by sequential scanning on single-layer optical sections at intervals of 1.0 μm.
Whole-cell patch-clamp recordings
The dissociated cells were bathed in Ringer solution containing (mm): NaCl 145, KCl 5, CaCl2 2, MgCl2 1, Hepes 10, and glucose 16; pH adjusted to 7.4 with NaOH. Patch pipettes were made by pulling BF150-86-10 glass (Sutter Instrument Co., Novato, CA, USA) on a P-97 Flaming/Brown micropipette puller (Sutter) and fire polished (model MF-830, Narishige, Tokyo, Japan) before recording. The pipette resistance was typically 3–5 MΩ after filling with the internal solution consisting of the following (in mm): CsCl 128, CaCl2 1, MgCl2 2, EGTA 10, Hepes 10, ATP-Mg 2, GTP-Na 0.4, and phosphocreatine 10; adjusted to pH 7.2 with CsOH and to 290–300 mosmol l−1. Whole-cell membrane currents induced by 100 μm glycine, which was applied for 5 s every 2 min, were picked up from RGCs, voltage-clamped at −60 mV, by a patch amplifier (EPC 9/2; HEKA Elektronik, Lambrecht/Pfalz, Germany). Fast capacitance was fully cancelled and cell capacitance was partially cancelled by the circuits of the amplifier as much as possible. Seventy per cent of the series resistance of the recording electrode was compensated. Commonly, data were acquired by using Pulsefit 8.62 (HEKA Elektronik) at a sample rate of 5 kHz, filtered at 2 kHz, and then stored for further analysis. All recordings were made at room temperature (20–25°C). The data analysis was performed using Clampfit 8.0 (Molecular Devices, Sunnyvale, CA, USA) and Igor 4.0 (WaveMetrics, Lake Oswego, OR, USA). Data are presented as means ± s.e.m. Student's t test or one-way ANOVA was used for statistical analysis.
For electrophysiological recordings in retinal slices, dark-adapted (overnight) rats were deeply anaesthetized with 4% sodium pentobarbital and killed by decapitation. Eyes were removed quickly and immersed in artificial cerebral spinal fluid (ACSF; in mm): NaCl 125, KCl 2.5, CaCl2 2, MgCl2 1, NaH2PO4 1.25, NaHCO3 25, glucose 15, pH 7.4) bubbled with 95% O2–5% CO2. Retinae were then isolated and sliced vertically at a thickness of 250 μm on a Narishige ST-21 slicer. Slices were transferred to a recording chamber and superfused continuously with oxygenated ACSF at 28–33°C. Whole cell light-evoked glycine receptor-mediated IPSCs of RGCs were recorded with an amplifier (EPC 10; HEKA Elektronik). The cell was held at +20 mV and the currents were filtered at 1 kHz. CNQX at 20 μm, 50 μm d-APV and 10 μm bicuculline were added to the perfusion solution to block AMPA, NMDA and GABAA receptor-mediated responses, respectively. The pipette solution consisted of (in mm): CsCH3SO3 120, TEA-Cl 10, Hepes 10, CaCl2 0.1, EGTA 1, phosphocreatine 12, ATP-Mg 3, GTP-Na 0.5; pH 7.2 adjusted with CsOH, 290 mosmol l−1. All procedures were carried out under dim red light. Diffuse light stimulation with duration of 1.5 s provided by a LED was triggered by the Pulse program (HEKA Elektronik) and delivered to the recording chamber through the microscope condenser. Light density on the surface of the slices was 1.88 μw cm−2.
Calcium imaging
Changes in intracellular calcium concentration ([Ca2+]i) were assessed using the ratiometric dye fura-2 AM (Dojindo, Kumamoto, Japan), which was first dissolved in 20% Pluronic F-127 (w/v, DMSO) and then added to a chamber containing Ringer solution to give a final concentration of 2 μm fura-2 AM. Isolated RITC-labelled RGCs were incubated in fura-2 AM for 30 min at room temperature and then perfused for at least 15 min with dye-free Ringer solution. Digital fluorescence images were recorded with a digital CCD camera (ORCA-ER; Hamamatsu Photonics, Shizuoka, Japan). A high-speed continuously scanning monochromatic light source (Polychrome V; Till Photonics, Gräfeling, Germany) was used for the excitations at wavelengths of 340 nm and 380 nm. Images showing RITC labelling were obtained at 535 nm. Fluorescence intensities at both wavelengths (F340 and F380) were measured every 3–10 s, and images were obtained using PC-based software (C-imaging systems; Hamamatsu Photonic). The ratio (R) between the two images was proportional to [Ca2+]i of the cell under study. Prior to an experiment, a bath ground level of fluorescence (due to autofluorescence and camera noise) was determined and subtracted from all the data obtained.
Drugs and solutions
U73122, d-APV, CNQX, bicuculline, 4-P-PDOT and PTX were from Tocris Bioscience (Ellisville, MO, USA). All others were purchased from Sigma-Aldrich, Inc. (St Louis, MO, USA). Melatonin, U73122, CNQX, PMA and Bis IV were first dissolved in DMSO and then diluted in solutions with a final DMSO concentration less than 0.1%, which has no significant effects on glycine-induced currents of isolated RGCs. All other drug solutions were prepared in distilled water, stored at −20°C and freshly diluted to the final concentration using normal solutions. Solution delivery and changes were performed using a stepper motor-based rapid solution changer RSC-100 (Bio-Logic, Claix, France), as described in detail previously (Chen et al. 2004), with the drug delivery capillaries being placed near the cell. For Ca2+-free internal solution, Ca2+ was removed and 10 mm BAPTA was added. Impermeant drugs (GDP-β-S, KT5823 and BAPTA) were dialysed into neurons after membrane rupture by filling them in the recording pipettes.
Results
MT2 receptors are expressed on rat RGCs
Expression of melatonin receptors on isolated RGCs was examined by double immunofluorescence labelling with the antibody against MT1 or MT2 receptors. The specificity of the antibodies was first tested using Western blot analysis. As shown in Fig. 1A and C, respectively, these two antibodies both revealed a single band at ∼45 kDa, corresponding to the molecular weights of native MT1 and MT2 receptors (Reppert et al. 1994, 1995). Figure 1Ba and b shows an isolated RGC, retrogradely labelled by FG, which had two processes extending from the soma. No labelling for MT1 was found in the cell at all (Fig. 1Bc and d), and all 62 RGCs tested were MT1-negative, without exception. In our previous work, immunohistochemical experiments with this antibody showed that MT1 receptors were expressed in horizontal cells in the carp retina (Huang et al. 2005), demonstrating that the antibody works well with the retina in both Western blot and immunohistochemistry. In contrast, labelling for MT2 was seen in 49 out of 55 RGCs tested. An example of the MT2-positive RGCs is shown in Fig. 1Da–d, in which intense labelling for MT2 was clearly seen on the plasma membrane. It was noteworthy that some punctate immunoreactivity was also found in the cell dendrites (Fig. 1Dc and d). We further evaluated the specificity of the immnuoflorescence labelling of the MT2 receptor antibody. No immunoflorescence labelling for MT2 receptors could be found (Fig. 1Ea and b) when the antibody was pre-absorbed with the corresponding immunizing antigen (Santa Cruz Biotechnology). Similar results were observed in all RGCs tested (n = 20).
Figure 1. Expression of melatonin MT2 receptors in rat retinal ganglion cells (RGCs).
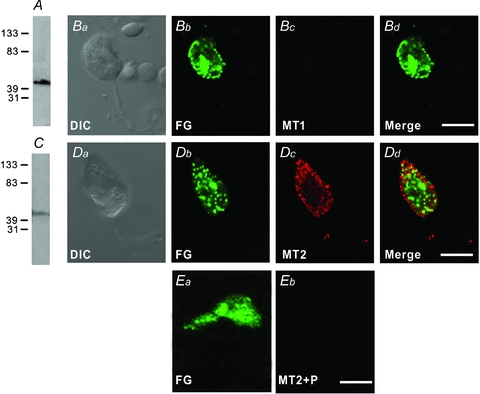
Double immunofluorescence labelling was performed using the antibody against MT1 or MT2 receptors on acutely dissociated RGCs labelled retrogradely by injecting fluorogold (FG) into the superior colliculus bilaterally. A, Western blot of whole rat retinal extract using the antibody against MT1, revealing a single band at the corresponding molecular weight of around 45 kDa. Ba–Bd, a RGC, double labelled by FG and MT1. Ba is the DIC image of the cell. The cell was labelled by FG (Bb), but not by MT1 (Bc). Bd is the merged image of Bb and Bc. C, Western blot of whole rat retinal extract using the antibody against MT2, also revealing a single band at around 45 kDa, corresponding to the molecular weight of the native MT2 receptor. Da–d, another RGC labelled by FG and MT2. Da is the DIC image of the cell. Dd is the merged image of Db, showing labelling for FG, and Dc, showing labelling for MT2. Note that the labelling for MT2 is primarily located on the membrane of the soma, and some punctate labelling is also observed in the dendrites. Ea, a RGC labelled by FG. Eb, no immunoflorescence labelling for MT2 receptors could be found when the MT2 antibody was pre-absorbed with the immunizing antigen. Scale bars, 10 μm.
Potentiation by melatonin of glycine currents of RGCs involves activation of G-proteins
In our previous work (Zhang et al. 2007), glycine-induced currents from rat RGCs commonly showed significant rundown with time, which made a precise analysis of the underlying mechanisms impossible. In the present study we protected the currents from rundown by adding ATP, GTP and phosphocreatine to the internal solution. As shown in Supplemental Fig. 1A, 100 μm glycine-induced currents were kept unchanged during a 10 min recording period, and at 10 min the peak current amplitudes were 100.7 ± 1.5% of the values at 0 min (n = 6, P > 0.05) (Supplemental Fig. 1C). Stable recordings could be commonly obtained for a period of 40 min. As reported before (Zhang et al. 2007), perfusion with melatonin of 10 nm reversibly increased the glycine-induced currents (Supplemental Fig. 1B and C; also see Fig. 2A). The peak current amplitudes were significantly increased to 138.8 ± 10.2% of control during melatonin application (n = 12). Such potentiation by melatonin of glycine currents were observed in 12 out of 19 cells tested. In the remaining seven cells no potentiation was observed. We also confirmed the observation reported in the previous work (Zhang et al. 2007) that the potentiation was completely reversed by co-application of 100 nm 4-P-PDOT (n = 7, Supplemental Fig. 1D and E), suggesting the involvement of MT2 receptors.
Figure 2. G-proteins mediating potentiation by melatonin of glycine-induced currents.
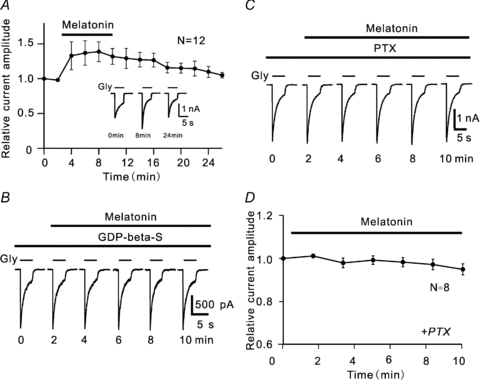
A, average peak amplitudes of glycine currents of RGCs (n = 12) are plotted as a function of time, showing the time course of the potentiation of glycine currents following the application of 10 nm melatonin. Glycine (Gly) of 100 μm was applied for 5 s every 2 min (this protocol is also applicable to Figs 3–7 and Supplemental Fig. 1). Inset, recordings from a RGC obtained at different times. B, glycine current traces taken from a RGC, showing no potentiation of the current by 10 nm melatonin in the presence of 3 mm GDP-β-S. C, representative recordings, showing that no potentiation of glycine currents by 10 nm melatonin was observed in a pertussis toxin (PTX) 1 μg ml−1 pretreated RGC. D, average peak amplitudes of glycine currents of RGCs as a function of time, showing no change in amplitudes by 10 nm melatonin in PTX-treated RGCs (n = 8). The data obtained in each cell were normalized to the control current of that cell (at 0 min), and the data collected from these cells in these experiments were then averaged, a procedure that was used to process all relevant data shown in Figs 2–7 and Supplemental Fig. 1. The data are presented as means ± s.e.m.
Since MT2 receptors are G-protein coupled receptors, inhibition of the G-protein activity should eliminate the melatonin-induced potentiation of glycine currents. This was confirmed by internal dialysis of GDP-β-S, a non-hydrolysable inhibitor of G-proteins. Figure 2B shows that, during internal dialysis of 3 mm GDP-β-S, application of 10 nm melatonin for 10 min failed to potentiate the glycine current of a RGC. In all seven cells tested no melatonin-induced potentiation was observed. Compared to the amplitudes recorded before melatonin application (control), those obtained at 10 min in all these cells were 96.3 ± 5.5% of control (P > 0.05).
Activation of melatonin MT2 receptors may be coupled to PTX-sensitive Gi/o and/or PTX-insensitive Gq proteins (Roka et al. 1999; Lai et al. 2001; Masana & Dubocovich, 2001; MacKenzie et al. 2002). We further examined which subtype(s) of G-proteins mediated the melatonin effect. For this purpose we pre-incubated RGCs in PTX (1 μg ml−1)-containing solution for at least 30 min, and then recorded glycine currents. As shown in Fig. 2C, in this PTX-treated cell melatonin (10 nm) did not potentiate the glycine current and the current was almost unchanged. This result was obtained in all eight cells tested and the peak amplitudes of currents recorded at 10 min after melatonin application were 95.0 ± 2.6% of control (n = 8, P > 0.05) (Fig. 2D), suggesting the involvement of PTX-sensitive Gi/o, but not PTX-insensitive Gq.
cAMP-PKA and cGMP-PKG signalling pathways are not involved in the potentiation by melatonin of glycine currents
Activation of MT2 receptors may modulate forskolin-stimulated cAMP production and cGMP formation through the soluble guanylyl cyclase pathway in mammalian cell lines expressing the recombinant MT1 receptors (Masana & Dubocovich, 2001; Alarma-Estrany & Pintor, 2007). We first examined whether membrane permeant 8-Br-cAMP or 8-Br-cGMP affects glycine currents of isolated RGCs. Figure 3A shows that extracellular application of 1 mm 8-Br-cAMP hardly changed the glycine current. The peak amplitudes of glycine currents recorded at 10 min after the application of 8-Br-cAMP were 96.8 ± 3.1% of control (n = 7, P > 0.05) (Fig. 3C). Similar results were obtained with application of 1 mm 8-Br-cGMP (Fig. 3B). The average peak amplitude obtained after a 10 min application of 8-Br-cGMP was 103.3 ± 13.6% of control (n = 10, P > 0.05) (Fig. 3C). Furthermore, melatonin-induced potentiation of glycine currents remained unchanged when PKA and PKG were inhibited. The result obtained in a RGC, which was pre-incubated in the external solution containing the PKA inhibitor H-89 (2 μm) for 10 min, is shown in Fig. 3D. In the presence of H-89, application of 10 nm melatonin enhanced the glycine current from 1.15 nA to 1.66 nA in 10 min (Fig. 3D). Similar results were obtained in 5 out of 11 cells tested and the peak current amplitudes recorded in the 5 cells were 140.3 ± 8.0% (n = 5) of control at 10 min after melatonin application (P < 0.01, Fig. 3F). It is likely that, as shown in Fig. 3E, application of 10 nm melatonin potentiated the glycine current of the RGC, which was first dialysed intracellularly with the PKG inhibitor KT5823 at 20 μm for 10 min, from 1.18 nA to 1.84 nA (Fig. 3E). Such potentiation was observed in 5 out of 8 cells tested and the peak amplitude of glycine currents recorded in these five cells was 141.9 ± 13.1% (n = 5) of control at 10 min after melatonin application (P < 0.01, Fig. 3F).
Figure 3. No involvement of cAMP-PKA and cGMP-PKG signalling pathways in the potentiation by melatonin of glycine currents.
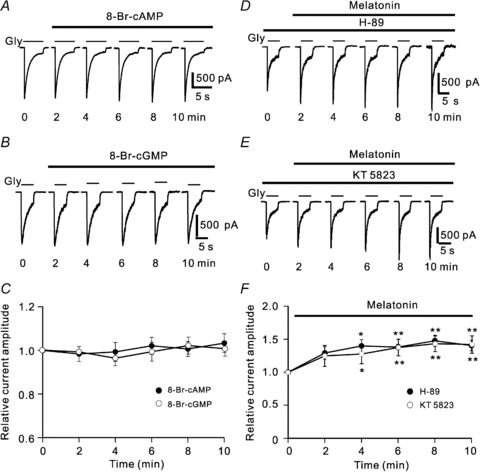
A and B, representative recordings made in two different cells, showing that the glycine currents were almost unchanged by application of 1 mm 8-Br-cAMP (A) or 1 mm 8-Br-cGMP (B). C, plot of average peak current amplitudes as a function of time, showing that either 8-Br-cAMP or 8-Br-cGMP hardly affected the glycine currents of RGCs (n = 7 for 8-Br-cAMP and n = 10 for 8-Br-cGMP). D and E, recordings made in two different RGCs showing that application of 2 μm H-89 (D) or 20 μm KT5823 (E) did not block the potentiation by melatonin of the glycine currents. F, plot of average peak current amplitudes as a function of time, showing the time course of the effects of 10 nm melatonin on glycine currents in the presence of H-89 (n = 5) or KT5823 (n = 5). *P < 0.05 and **P < 0.01, as compared to the current amplitudes before melatonin application (control).
PC-PLC/PKC signalling pathway mediates melatonin-induced potentiation of glycine currents
As well as the PKA and PKG pathways, the PLC-PKC signalling pathway is another target when MT1 or MT2 receptors are activated (McArthur et al. 1997; Lai et al. 2002; MacKenzie et al. 2002; Alarma-Estrany & Pintor, 2007). Thus, we explored if PLC may be involved in the melatonin-induced potentiation of glycine currents by extracellularly applying D609, a phosphatidylcholine (PC)-specific phospholipase (PLC) inhibitor (Girón-Calle et al. 2002). Figure 4A shows a representative result. Application of 60 μm D609 gradually reduced the glycine current, and the current reached a steady level at 4 min, which was of about 75% of control. Co-application of 10 nm melatonin for 8 min did not influence the decreased current amplitude. Once D609 was withdrawn from the external solution with melatonin remaining, the peak amplitude of the glycine current was considerably increased. Similar observations were obtained in 4 out of 6 cells tested. The average changes in current amplitudes obtained in these four cells during such experiments are shown in Fig. 4B as a function of time. The peak amplitudes of glycine currents reduced to 73.8 ± 9.1% of control during a first 6 min D609 application. Subsequent co-application of 10 nm melatonin for 8 min did not influence the reduced peak amplitudes of currents (69.7 ± 10.2% of control). When D609 was withdrawn, however, the peak currents were gradually increased in size and reached 141.2 ± 7.3% of control at 24 min.
Figure 4. Involvement of PC-PLC pathways in melatonin-induced potentiation of glycine currents.
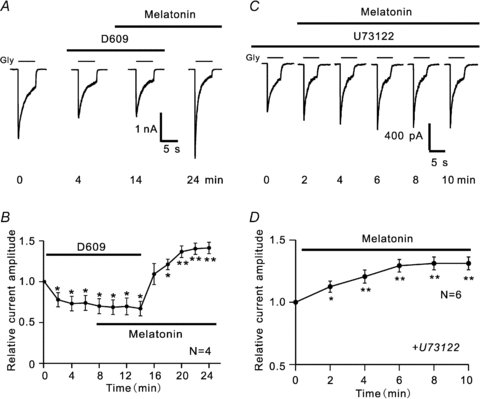
A, representative recordings from a RGC, showing the effect of melatonin on glycine currents in the presence of 60 μm D609. Note that D609 itself reduced the current amplitude (4 min). Addition of 10 nm melatonin in the presence of D609 did not increase the current (14 min). When D609 was removed from the external solution, the peak current amplitude was significantly increased (24 min). B, plot of average peak amplitudes of glycine currents (n = 4) as a function of time, showing the change in amplitudes. *P < 0.05 and **P < 0.01 as compared to that recorded in Ringer solution (at 0 min). C, current traces, showing the effect of melatonin on glycine currents in a RGC dialysed with 5 μm U73122. In the presence of U73122, melatonin still caused a potentiation of the glycine current. D, average peak glycine current amplitudes (n = 6) changed as a function of time in the presence of 5 μm U73122, showing that melatonin persisted to cause a current potentiation. *P < 0.05 and **P < 0.01 (vs. control).
To determine whether or not activation of MT2 receptors may potentiate the glycine current through the phosphatidylinositol (PI)-PLC pathway (McArthur et al. 1997; Ross et al. 1998), we dialysed U73122 (5 μm), a PI-PLC inhibitor, into RGCs through recording pipettes for at least 10 min and then observed the effects of 10 nm melatonin on glycine currents. In 6 out of 9 cells tested melatonin persisted to potentiate the glycine currents (Fig. 4C) and at 10 min after melatonin application, the peak current amplitudes were 131.1 ± 5.2% of control (n = 6, P < 0.01) (Fig. 4D), which is comparable to the potentiation extent observed in normal Ringer solution (versus 138.8 ± 10.2% in Ringer solution).
Activated PC-PLC could hydrolyse membrane phosphatidylcholine (PC) into diacylglycerol (DAG) and phosphocholine, and DAG is known to enhance the PKC activity (Way et al. 2000). We therefore tested if activation of PKC would affect glycine currents. As shown in Fig. 5A, extracellular application of 1 μm phorbol 12-myristate 13-acetate (PMA), a PKC activator, just like melatonin, gradually increased the glycine current of the cell. In the presence of PMA, melatonin did not cause a further increase of the peak amplitude. The PMA-induced potentiation of glycine currents was observed in 9 out of 17 cells tested. In these nine cells, on average, the peak currents were increased to 131.4 ± 8.5% of control (P < 0.05, Fig. 5B). Co-application of 10 nm melatonin did not change the potentiated peak currents, being 129.7 ± 13.4% of control (P < 0.05 vs. control and P > 0.05 vs. PMA) (Fig. 5B). We also examined the effects of bisindolylmaleimide IV (Bis IV), a PKC inhibitor, on glycine currents, and a representative result obtained in such experiments is shown in Fig. 5C. Application of 5 μm Bis IV alone caused a decrease of the current, and the addition of 10 nm melatonin during the Bis IV application did not significantly change the current amplitude. When Bis IV was removed from the solution but melatonin remained, the glycine current was increased in size and became even larger than that recorded before the application of Bis IV. Similar observation was obtained in 6 out of 8 cells tested and the data collected from these six cells are summarized in Fig. 5D. The peak currents were reduced to 69.6 ± 5.7% of control (P < 0.01) during Bis IV application, which was followed by a significant increase in current amplitudes (to 132.3 ± 8.6% of control, P < 0.01) once Bis IV was removed but melatonin remained.
Figure 5. PKC is involved in melatonin-caused potentiation of glycine currents.
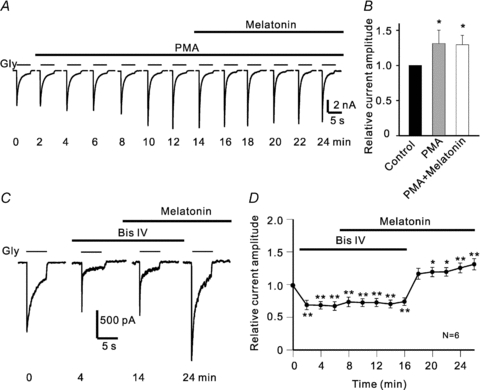
A, current traces from a RGC, showing that extracellular application of 1 μm PMA gradually increased the current amplitude, and in the presence of PMA 10 nm melatonin did not cause a further increase. B, cumulative data from nine cells showing the effects of PMA and melatonin on glycine currents. The data were taken at different time points as shown in A (0 min for control; 12 min for PMA; 24 min for PMA+melatonin). *P < 0.05 vs. control. C, current traces recorded at different times, showing that application of 5 μm Bis IV reduced the glycine current of a RGC, and in the presence of Bis IV 10 nm melatonin failed to potentiate the current, but potentiation of the glycine current was clearly observed when Bis IV was withdrawn but melatonin remained. D, average peak amplitudes of glycine currents (n = 6) are plotted as a function of time during such experiments. *P < 0.05 and **P < 0.01, as compared to that in Ringer solution (0 min).
Intracellular Ca2+ is not involved in melatonin-induced potentiation of glycine currents
Whereas PKC activation may be either calcium dependent or independent, we then tried to determine the PKC type(s) mediating the melatonin-induced potentiation. We first examined whether melatonin could change [Ca2+]i in RGCs by calcium imaging. Figure 6A shows a representative result obtained from an isolated RGC retrogradely labelled by RITC. Application of 20 nm melatonin did not induce significant change in [Ca2+]i of the cell, represented as the ratio (340/380) (also see the CCD pictures shown in the inset). Following washout with Ringer solution for 4 min, however, application of 60 mm KCl induced a dramatic increase in [Ca2+]i in the cell. In 13 RGCs tested, the average ratio of fura-2 (340/380) after the application of 20 nm melatonin was 1.26 ± 0.16 (n = 13), which was comparable to that obtained in Ringer solution (1.27 ± 0.16) (P > 0.05) (Fig. 6B). In contrast, 60 mm KCl increased the ratio from the basal value of 1.30 ± 0.14 to 2.62 ± 0.13 (P < 0.01, Fig. 6B).
Figure 6. Intracellular Ca2+ is not involved in melatonin-induced potentiation of glycine currents.
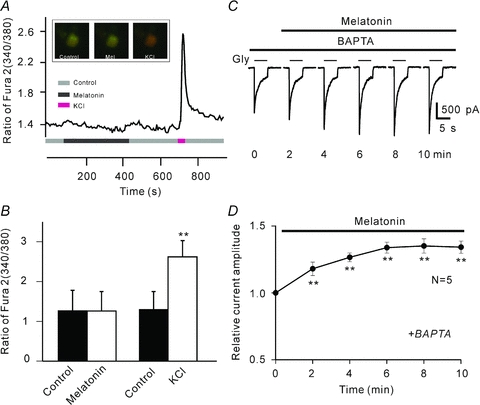
A, a continuous recording of [Ca2+]i in a RGC, represented by the ratio of fura-2 AM fluorescence at 340 nm and 380 nm (340/380). Application of 20 nm melatonin hardly changed [Ca2+]i. After washout with Ringer solution for 4 min, application of 60 mm KCl induced a dramatic increase of [Ca2+]i. Inset, three CCD images of a RITC-labelled RGC loaded with fura-2 AM, taken before (left) and after (middle) 6 min perfusion of 20 nm melatonin, and in the presence of 60 mm KCl (right). B, bar chart summarizing the changes of [Ca2+]i induced by melatonin (20 nm) and KCl (60 mm) in RGCs (n = 13, **P < 0.01 vs. control). C, current traces recorded at different times from a RGC, showing that elimination of [Ca2+]i by 10 mm BAPTA did not change the potentiation by melatonin of the glycine current. D, average peak amplitudes of glycine currents are plotted as a function of time, showing the effect of melatonin on glycine currents when 10 mm BAPTA-filled recording pipettes were used (n = 5, **P < 0.01, as compared to the currents recorded at 0 min).
We also eliminated [Ca2+]i to determine if the melatonin-induced potentiation might be changed. For this experiment, the recording pipettes were filled with the Ca2+-free internal solution containing 10 mm BAPTA, a fast Ca2+ chelator. A calculation using MaxChelator version 6.81 (Bers et al. 1994) indicated that [Ca2+]i may be reduced to 1 pm under this condition. In 5 out of 7 cells tested, application of 10 nm melatonin still caused a potentiation of glycine currents (Fig. 6C). In these 5 cells the average peak amplitudes were increased to 131.0 ± 3.4% of control (P < 0.01, Fig. 6D), which was not different from that obtained in normal internal solution (138.8 ± 10.2% of control).
Melatonin potentiates light-evoked glycine receptor-mediated IPSCs in RGCs in rat retinal slice preparations
To further confirm the potentiation of glycine currents by melatonin and explore physiological implication of the potentiation, we tested the effects of melatonin on light-evoked glycine receptor-mediated IPSCs in retinal slice preparations. In total, whole cell light responses were recorded from 24 RGCs. In 13 of these cells, light-induced responses were completely blocked by co-application of CNQX, d-APV and bicuculline, and no glycine receptor-mediated IPSCs could be recorded. In the remaining 11 cells, light-induced glycine receptor-mediated IPSCs were recorded, and the current recordings obtained from a representative one are shown in Fig. 7A. Application of 50 nm melatonin significantly increased the peak current amplitude, and the melatonin effect was eliminated by co-application of 100 nm 4-P-PDOT. All these responses were abolished by 5 μm strychnine. Similar results were obtained from 8 out of the 11 cells. In these eight cells the average peak amplitudes of IPSCs after application of melatonin were increased to 124.8 ± 3.3% of control (P < 0.001), and they returned to 101.0 ± 3.4% of control (P > 0.05) after co-application of 4-P-PDOT (Fig. 7B). The potentiation was not observed in the remaining three cells.
Figure 7. Melatonin potentiates light-evoked glycine receptor-mediated IPSCs in RGCs of rat retinal slices.
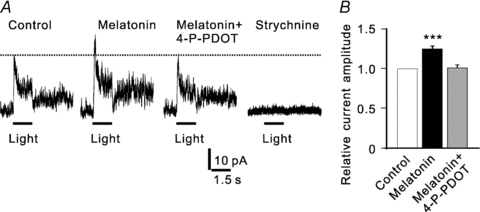
A, light evoked current traces from a RGC, recorded in the presence of d-APV, CNQX, and bicuculline. Application of 50 nm melatonin increased the peak amplitude, and the effect was reversed by co-application of 4-P-PDOT. The current was eliminated by application of 5 μm strychnine. The cell was held at +20 mV. B, bar chart summarizing the changes of peak amplitudes of light-evoked glycine currents induced by melatonin and co-application with 4-P-PDOT. n = 8, ***P < 0.001 vs. control.
Discussion
MT2 receptors mediate melatonin-induced potentiation of glycine currents through activation of PTX-sensitive G-proteins
Whilst MT1 receptors are rather widely distributed in the retina (Fujieda et al. 1999; Natesan & Cassone, 2002; Savaskan et al. 2002; Huang et al. 2005; Alarma-Estrany & Pintor, 2007), the knowledge about the distribution of MT2 receptors in the retina is quite limited (Natesan & Cassone, 2002; Wiechmann, 2003; Wiechmann et al. 2004; Alarma-Estrany & Pintor, 2007). Specifically, in the inner retina both MT1 and MT2 receptors were found to be present on RGCs in Xenopus laevis (Wiechmann, 2003; Wiechmann et al. 2004). Using in situ hybridization, Fujieda et al. (1999) demonstrated the localization of MT1 mRNA in the IPL of rat retina, but the weak labelling did not allow identification of the cell types that contributed to the staining. In the present work we demonstrated that MT2, but not MT1, receptors are expressed in the majority, but not all, of rat RGCs. It was of particular interest that intense labelling for MT2 was on the plasma membrane and punctate immunoreactivity was found in the dendrites. This result is reminiscent of the observation made in domestic chick that MT2 mRNA was found in RGCs (Natesan & Cassone, 2002). Consistent with the immunocytochemical result, melatonin induced a potentiation of glycine currents in the majority, but not all, of rat RGCs and the potentiation was completely blocked by 4-P-PDOT. It seems unlikely that the failure to observe the melatonin effect in some RGCs was due to destruction of MT2 receptors during the cell dissociation. Rather, we tend to speculate that expression of MT2 receptors may be cell subtype dependent, an issue that remains to be addressed. As MT2 receptors are G-protein coupled, it could be expected that the melatonin effect was eliminated by intracellular dialysis of GDP-β-S (Fig. 2B). Furthermore, melatonin-induced potentiation was abolished when the RGCs were treated by PTX incubation, suggesting the involvement of Gi/o, but not Gq.
The PC-PLC/PKC signalling pathway is responsible for melatonin-induced potentiation of glycine currents
Like MT1 receptors, activation of MT2 receptors may inhibit forskolin-stimulated cAMP formation by inhibiting adenylate cyclase activity, which is followed by a decrease in activated PKA, as shown in mammalian cell lines expressing the recombinant receptors (Vanecek, 1998; Masana & Dubocovich, 2001). Challenging MT2 receptors could also lead to changes in cGMP-PKG activity (Petit et al. 1999; Huang et al. 2005; Ping et al. 2008). In carp rod-dominant ON type bipolar cells, for instance, activation of MT2 receptors increases the cGMP level by inhibiting phosphodiesterase, which results in a potentiation of rod signals to these cells (Ping et al. 2008). PKA- and PKG-mediated modulation of glycine currents has been also found in various types of neurons (Tapia et al. 1997; Ren et al. 1998). In the present study, however, application of either 8-Br-cAMP or 8-Br-cGMP did not alter the glycine currents (Fig. 3A–C) and melatonin persisted to potentiate the glycine currents of RGCs when PKA and PKG were inhibited by H-89 and KT5823, respectively (Fig. 3D–F). All these results suggest no involvement of both cAMP-PKA and cGMP-PKG pathways in the melatonin-induced potentiation of glycine currents of rat RGCs.
It has been reported that activation of MT2 receptors may involve modulation of PLC activity. In rat SCN slices melatonin produces a phase shift that is mediated by MT2 receptors through an increase in PLC-linked PKC activity (Gerdin et al. 2004; Dubocovich et al. 2005), but this signalling pathway has not been elaborated. The pharmacological dissections conducted in this study provide an insight into the PLC/PKC signalling pathway that may be responsible for the melatonin-induced potentiation of glycine currents. Application of the specific PC-PLC inhibitor D609 caused a decrease in glycine current amplitudes and in the presence of D609 melatonin failed to cause the potentiation in the cells for which melatonin induced a potentiation of glycine currents (Fig. 4B). Even though the mammalian enzyme PC-PLC has not yet been isolated or cloned (Weernink et al. 2007), there is a lot of indirect evidence to suggest that PC-PLC is indeed expressed in a variety of cells (Ramoni et al. 2001; Mateas et al. 2006; Spadaro et al. 2006; Cecchetti et al. 2007; Spadaro et al. 2008; Tzeng et al. 2010). While D609 is recently reported to inhibit group IV cytosolic phospholipase A2 (cPLA2) and reduce the release of arachidonic acid non-competitively and uncompetitively, but with a Ki value of 86.25 μm and an IC50 value of 375 μm, respectively (Kang et al. 2008), accumulating evidence suggests that D609 is a rather specific PC-PLC inhibitor (Spadaro et al. 2008; Tzeng et al. 2010). In addition, the D609 concentration we used in the present work was 60 μm. We therefore suppose that the effect of D609 may be due to an inhibition of the activity of PC-PLC, but not cPLA2 or others. Of course, a possible involvement of cPLA2 could not be excluded. Although activation of MT2 receptors may also stimulate the production of inositol 1,4,5-trisphosphate (IP3) through the phosphatidylinositol (PI)-PLC pathway, thus leading to an increased calcium release from intracellular Ca2+ stores, which in turn activates Ca2+-dependent PKC (McArthur et al. 1997; Ross et al. 1998), it seems not to be the case for the melatonin effect reported in this work. That is simply because the melatonin-induced potentiation persisted when PI-PLC was inhibited by U73122. Furthermore, as shown in Fig. 5A, application of the PKC activator PMA mimicked the melatonin effect and in the presence of PMA melatonin did not further boost the enhanced glycine currents, whereas the PKC inhibitor Bis IV eliminated the melatonin effect. These results are consistent with the evidence that PC-PLC enhances the activity of PKC through activation of DAG (Way et al. 2000). The involvement of PC-PLC was further supported by the calcium imaging results, showing no increase in [Ca2+]i by melatonin application and that the melatonin effect was calcium independent (Fig. 6A–D). There are several PKC phosphorylation sites in glycine receptor subunits (Ruiz-Gómez et al. 1991; Vaello et al. 1994), and activation of PKC is found to modulate glycine receptor-mediated currents positively or negatively (Vaello et al. 1994; Nishizaki & Ikeuchi, 1995; Gu & Huang, 1998; Han & Slaughter, 1998). Phosphorylation of the glycine receptor on rat RGCs caused by Ca2+-independent activation of PKC through a distinct Gi/o-PC-PLC signalling pathway may be responsible for the melatonin-induced potentiation of the glycine currents. The intracellular signalling pathway is illustrated in the schematic diagram Fig. 8.
Figure 8. A schematic diagram showing the possible signalling pathway mediating the potentiation by melatonin of glycine currents in rat RGCs.
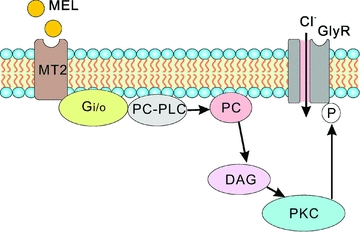
By challenging PTX-sensitive Gi/o-coupled MT2 receptors on rat RGCs, melatonin potentiates glycine currents via a distinct intracellular PC-PLC/PKC pathway. Mel: melatonin; GlyR: glycine receptor; PC-PLC: phosphatidylcholine-specific phospholipase C; PC: phosphatidylcholine; DAG: diacylglycerol; PKC: protein kinase C; ℗, phosphorylation.
In the mammalian inner retina glycine released from amacrine cells modulates visual information processing (Wässle et al. 2009). AII amacrine cells are probably the most numerous type of glycinergic amacrine cells in mammalian retina (Menger et al. 1998; Wässle et al. 2009). Like other types of glycinergic amacrine cells, AII cells have vertically oriented dendritic trees, thus largely being narrow field ones (Menger et al. 1998; Wässle, 2004; Hsueh et al. 2008; Molnar et al. 2009). These cells are thought to be primarily involved in the so-called crossover inhibition between ON and OFF pathways in the inner retina (Zaghloul et al. 2003; Wässle, 2004; Roska et al. 2006; Hsueh et al. 2008; Manookin et al. 2008; Molnar et al. 2009), which are used to encode positive and negative contrast respectively. In several species there is accumulating evidence suggesting the presence of a substantial inhibitory input from AII cells to OFF RGC dendrites mediated by glycine receptors (Wässle, 2004; Manookin et al. 2008; Murphy & Rieke, 2008; Van Wyk et al. 2009). Specifically, patch-clamp recordings demonstrated that light (positive contrast)-driven responses of OFF-A type RGCs in the mouse retina were mediated by direct glycinergic inhibitory inputs (Van Wyk et al. 2009), which is consistent with an early observation showing a complete blockade of extracellularly recorded light responses of OFF RGCs by application of strychnine (Müller et al. 1988). Light stimulation induced glycine receptor-mediated responses from a subpopulation of RGCs in this work, suggesting that these cells may receive direct inhibitory inputs from glycinergic amacrine cells, including AII cells. Although we did not try to identify the RGCs whose light induced glycine currents were recorded in our experiments, based on the location of their somata and our expertise, most of them might be OFF cells. On the other hand, AII cells would also excite OFF RGCs by reducing glycine release due to a hyperpolarization of these cells driven by electrical synapses with ON cone bipolar cells (disinhibition) when light decrement (negative contrast) appears (Manookin et al. 2008). In either case potentiation of the response of OFF RGCs to glycine implies that the direct inhibition (for positive contrast) or the disinhibition (for negative contrast) would be enhanced in the presence of melatonin. In other words, melatonin may drive the membrane potential of OFF RGCs to a more hyperpolarized level, which would be beneficial for these cells to detect small change in brightness at low light levels (Manookin et al. 2008). Furthermore, the crossover inhibition is known to take place among RGCs and even at higher levels of the visual pathway (Hirsch, 2003; Molnar et al. 2009). Another physiological implication of the melatonin-increased suppression of OFF RGCs therefore could be that it would enhance the light sensitivity of the ON pathway in the lateral geniculate nucleus and the visual cortex. It has recently been shown that crossover to OFF RGCs indeed enhances the light sensitivity of these cells (Manookin et al. 2008; Molnar et al. 2009). Taken together, the observation reported in this work suggests that melatonin, being at higher levels at night, might help animals detect targets in night vision by enhancing the direct inhibition and/or disinhibition mediated by glycinergic amacrine cells.
Acknowledgments
We thank Jie Chen and Xin-Jun Zhang for their technical help. This work was supported by grants from the National Program of Basic Research sponsored by the Ministry of Science and Technology of China (2006CB500805, 2007CB512205), the Natural Science Foundation of China (30870803, 30930034, 30900427), Pujiang Talent Project of the Shanghai Science and Technology Committee (08PJ14016), and the 211 Project sponsored by the Ministry of Education of China.
Glossary
Abbreviations
- Bis IV
bisindolylmaleimide IV
- DAG
diacylglycerol
- FG
fluorogold
- PC
phosphatidylcholine
- PI
phosphatidylinositol
- PKA
protein kinase A
- PKC
protein kinase C
- PKG
protein kinase G
- PLC
phospholipase C
- PMA
phorbol 12-myristate 13-acetate
- PTX
pertussis toxin
- RGC
retinal ganglion cell
- RITC
rhodamine-B-isothiocyanate
- SCN
suprachiasmatic nucleus
Author contributions
Z.W. and X.L.Y. designed and supervised the experiments. W.J.Z., M.Z. and Y.M. performed the experiments, and collected and analysed data. Z.W. and X.L.Y. analysed data and prepared the manuscript. All authors contributed to experimental design, discussed the results and commented on the report. All experiments were done in Fudan University.
Supplemental material
Supplemental Figure 1
As a service to our authors and readers, this journal provides supporting information supplied by the authors. Such materials are peer-reviewed and may be re-organized for online delivery, but are not copy-edited or typeset. Technical support issues arising from supporting information (other than missing files) should be addressed to the authors
References
- Alarma-Estrany P, Pintor J. Melatonin receptors in the eye: location, second messengers and role in ocular physiology. Pharmacol Ther. 2007;113:507–522. doi: 10.1016/j.pharmthera.2006.11.003. [DOI] [PubMed] [Google Scholar]
- Barrenetxe J, Delagrange P, Martinez JA. Physiological and metabolic functions of melatonin. J Physiol Biochem. 2004;60:61–72. doi: 10.1007/BF03168221. [DOI] [PubMed] [Google Scholar]
- Bers DM, Patton CW, Nuccitelli R. A practical guide to the preparation of Ca2+ buffers. Methods Cell Biol. 1994;40:3–29. doi: 10.1016/s0091-679x(08)61108-5. [DOI] [PubMed] [Google Scholar]
- Cahill GM, Besharse JC. Light-sensitive melatonin synthesis by Xenopus photoreceptors after destruction of the inner retina. Vis Neurosci. 1992;8:487–490. doi: 10.1017/s0952523800005009. [DOI] [PubMed] [Google Scholar]
- Cecchetti S, Spadaro F, Lugini L, Podo F, Ramoni C. Functional role of phosphatidycholine-specific phospholipases C in regulating CD16 membrane expression in natural killer cells. Eur J Immunol. 2007;37:2912–2922. doi: 10.1002/eji.200737266. [DOI] [PubMed] [Google Scholar]
- Chen L, Yu YC, Zhao JW, Yang XL. Inwardly rectifying potassium channels in rat retinal ganglion cells. Eur J Neurosci. 2004;20:956–964. doi: 10.1111/j.1460-9568.2004.03553.x. [DOI] [PubMed] [Google Scholar]
- Drummond GB. Reporting ethical matters in The Journal of Physiology: standards and advice. J Physiol. 2009;587:713–719. doi: 10.1113/jphysiol.2008.167387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dubocovich ML, Hudson RL, Sumaya IC, Masana MI, Manna E. Effect of MT1 melatonin receptor deletion on melatonin-mediated phase shift of circadian rhythms in the C57BL/6 mouse. J Pineal Res. 2005;39:113–120. doi: 10.1111/j.1600-079X.2005.00230.x. [DOI] [PubMed] [Google Scholar]
- Dubocovich ML, Rivera-Bermúdez MA, Gerdin MJ, Masana MI. Molecular pharmacology, regulation and function of mammalian melatonin receptors. Front Biosci. 2003;8:d1093–1108. doi: 10.2741/1089. [DOI] [PubMed] [Google Scholar]
- Fujieda H, Hamadanizadeh SA, Wankiewicz E, Pang SF, Brown GM. Expression of mt1 melatonin receptor in rat retina: evidence for multiple cell targets for melatonin. Neuroscience. 1999;93:793–799. doi: 10.1016/s0306-4522(99)00111-6. [DOI] [PubMed] [Google Scholar]
- Gerdin MJ, Masana MI, Rivera-Bermúdez MA, Hudson RL, Earnest DJ, Gillette MU, Dubocovich ML. Melatonin desensitizes endogenous MT2 melatonin receptors in the rat suprachiasmatic nucleus: relevance for defining the periods of sensitivity of the mammalian circadian clock to melatonin. FASEB J. 2004;18:1646–1656. doi: 10.1096/fj.03-1339com. [DOI] [PubMed] [Google Scholar]
- Girón-Calle J, Srivatsa K, Forman HJ. Priming of alveolar macrophage respiratory burst by H2O2 is prevented by phosphatidylcholine-specific phospholipase C inhibitor tricyclodecan-9-yl-xanthate (D609) J Pharmacol Exp Ther. 2002;301:87–94. doi: 10.1124/jpet.301.1.87. [DOI] [PubMed] [Google Scholar]
- Gu Y, Huang LY. Cross-modulation of glycine-activated Cl− channels by protein kinase C and cAMP-dependent protein kinase in the rat. J Physiol. 1998;506:331–339. doi: 10.1111/j.1469-7793.1998.331bw.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han Y, Slaughter MM. Protein kinases modulate two glycine currents in salamander retinal ganglion cells. J Physiol. 1998;508:681–690. doi: 10.1111/j.1469-7793.1998.681bp.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirsch JA. Synaptic physiology and receptive field structure in the early visual pathway of the cat. Cerebral Cortex. 2003;13:63–69. doi: 10.1093/cercor/13.1.63. [DOI] [PubMed] [Google Scholar]
- Hsueh HA, Molnar A, Werblin FS. Amacrine-to-amacrine cell inhibition in the rabbit retina. J Neurophysiol. 2008;100:2077–2088. doi: 10.1152/jn.90417.2008. [DOI] [PubMed] [Google Scholar]
- Huang H, Lee SC, Yang XL. Modulation by melatonin of glutamatergic synaptic transmission in the carp retina. J Physiol. 2005;569:857–871. doi: 10.1113/jphysiol.2005.098798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang MS, Jung SY, Jung KM, Kim SK, Ahn KH, Kim DK. D609, an inhibitor of phosphatidylcholine-specific phospholipase C, inhibits group IV cytosolic phospholipase A2. Mol Cells. 2008;30:481–485. [PubMed] [Google Scholar]
- Lai FP, Mody SM, Yung LY, Kam JY, Pang CS, Pang SF, Wong YH. Molecular determinants for the differential coupling of Gα16 to the melatonin MT1, MT2 and Xenopus Mel1c receptors. J Neurochem. 2002;80:736–745. doi: 10.1046/j.0022-3042.2002.00767.x. [DOI] [PubMed] [Google Scholar]
- Lai FP, Mody SM, Yung LY, Pang CS, Pang SF, Wong YH. Chimeric Gαq subunits can distinguish the long form of the Xenopus Mel1c melatonin receptor from the mammalian mt1 and MT2 melatonin receptors. J Pineal Res. 2001;30:171–179. doi: 10.1034/j.1600-079x.2001.300306.x. [DOI] [PubMed] [Google Scholar]
- MacKenzie RS, Melan MA, Passey DK, Witt-Enderby PA. Dual coupling of MT1 and MT2 melatonin receptors to cyclic AMP and phosphoinositide signal transduction cascades and their regulation following melatonin exposure. Biochem Pharmacol. 2002;63:587–595. doi: 10.1016/s0006-2952(01)00881-4. [DOI] [PubMed] [Google Scholar]
- Manookin MB, Beaudoin DL, Ernst ZR, Flagel LJ, Demb JB. Disinhibition combines with excitation to extend the operating range of the OFF visual pathway in daylight. J Neurosci. 2008;28:4136–4150. doi: 10.1523/JNEUROSCI.4274-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masana MI, Dubocovich ML. Melatonin receptors signalling: finding the path through the dark. Sci STKE. 2001;107:pe39. doi: 10.1126/stke.2001.107.pe39. [DOI] [PubMed] [Google Scholar]
- Mateas MV, Uranga RM, Salvador GA, Giusto NM. Coexistence of phosphatidycholine-specific phospholipases C and phospholipase D activities in rat cerebral cortex synaptosomes. Lipids. 2006;41:273–280. doi: 10.1007/s11745-006-5097-3. [DOI] [PubMed] [Google Scholar]
- McArthur AJ, Hunt AE, Gillette MU. Melatonin action and signal transduction in the rat suprachiasmatic circadian clock: activation of protein kinase C at dusk and dawn. Endocrinology. 1997;138:627–634. doi: 10.1210/endo.138.2.4925. [DOI] [PubMed] [Google Scholar]
- Menger N, Pow DV, Wässle H. Glycinergic amacrine cells of the rat retina. J Comp Neurol. 1998;401:34–46. doi: 10.1002/(sici)1096-9861(19981109)401:1<34::aid-cne3>3.0.co;2-p. [DOI] [PubMed] [Google Scholar]
- Molnar A, Hsueh HA, Roska B, Werbin FS. Crossover inhibition in the retina: circuitry that compensates for nonlinear rectifying synaptic transmission. J Comput Neurosci. 2009;27:569–590. doi: 10.1007/s10827-009-0170-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müller F, Wässle H, Voigt T. Pharmacological modulation of the rod pathway in the cat retina. J Neurophysiol. 1988;59:1657–1672. doi: 10.1152/jn.1988.59.6.1657. [DOI] [PubMed] [Google Scholar]
- Murphy GJ, Rieke F. Signals and noise in an inhibitory interneuron diverge to control activity in nearby retinal ganglion cells. Nat Neurosci. 2008;11:318–326. doi: 10.1038/nn2045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Natesan AK, Cassone VM. Melatonin receptor mRNA localization and rhythmicity in the retina of the domestic chick, Gallus domesticus. Vis Neurosci. 2002;19:265–274. doi: 10.1017/s0952523802192042. [DOI] [PubMed] [Google Scholar]
- Nishizaki T, Ikeuchi Y. Activation of endogenous protein kinase C enhances currents through α1 and α2 glycine receptor channels. Brain Res. 1995;687:214–216. doi: 10.1016/0006-8993(95)00543-y. [DOI] [PubMed] [Google Scholar]
- Petit L, Lacroix I, de Coppet P, Strosberg AD, Jockers R. Differential signalling of human Mel1a and Mel1b melatonin receptors through the cyclic guanosine 3′-5′-monophosphate pathway. Biochem Pharmacol. 1999;58:633–639. doi: 10.1016/s0006-2952(99)00134-3. [DOI] [PubMed] [Google Scholar]
- Ping Y, Huang H, Zhang XJ, Yang XL. Melatonin potentiates rod signals to ON type bipolar cell in fish retina. J Physiol. 2008;586:2683–2694. doi: 10.1113/jphysiol.2008.152959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Protti DA, Gerschenfeld HM, Llano I. GABAergic and glycinergic IPSCs in ganglion cells of rat retinal slices. J Neurosci. 1997;17:6075–6085. doi: 10.1523/JNEUROSCI.17-16-06075.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramoni C, Spadaro F, Menegon M, Podo F. Cellular localization and functional role of phosphatidycholine-specific phospholipases C in NK cells. J Immunol. 2001;167:2642–2650. doi: 10.4049/jimmunol.167.5.2642. [DOI] [PubMed] [Google Scholar]
- Ren J, Ye JH, McArdle JJ. cAMP-dependent protein kinase modulation of glycine-activated chloride current in neurons freshly isolated from rat ventral tegmental area. Brain Res. 1998;811:71–78. doi: 10.1016/s0006-8993(98)00959-7. [DOI] [PubMed] [Google Scholar]
- Reppert SM, Godson C, Mahle CD, Weaver DR, Slaugenhaupt SA, Gusella JF. Molecular charaterization of a second melatonin receptor expressed in human retina and brain: the Mel1b melatonin receptor. Proc Natl Acad Sci U S A. 1995;92:8734–8738. doi: 10.1073/pnas.92.19.8734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reppert SM, Weaver DR, Ebisawa T. Cloning and characterization of a mammalian melatonin receptor that mediates reproductive and circadian responses. Neuron. 1994;13:1177–1185. doi: 10.1016/0896-6273(94)90055-8. [DOI] [PubMed] [Google Scholar]
- Ribelayga C, Wang Y, Mangel SC. A circadian clock in the fish retina regulates dopamine release via activation of melatonin receptors. J Physiol. 2004;554:467–482. doi: 10.1113/jphysiol.2003.053710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roka F, Brydon L, Waldhoer M, Strosberg AD, Freissmuth M, Jockers R, Nanoff C. Tight association of the human Mel1a-melatonin receptor and Gi: precoupling and constitutive activity. Mol Pharmacol. 1999;56:1014–1124. doi: 10.1124/mol.56.5.1014. [DOI] [PubMed] [Google Scholar]
- Roska B, Molnar A, Werblin FS. Parallel processing in retinal ganglion cells: how integration of space-time patterns of excitation and inhibition form the spiking output. J Neurophysiol. 2006;95:3810–3822. doi: 10.1152/jn.00113.2006. [DOI] [PubMed] [Google Scholar]
- Ross AW, Webster CA, Thompson M, Barrett P, Morgan PJ. A novel interaction between inhibitory melatonin receptors and protein kinase C-dependent signal transduction in ovine pars tuberalis cells. Endocrinology. 1998;139:1723–1730. doi: 10.1210/endo.139.4.5869. [DOI] [PubMed] [Google Scholar]
- Ruiz-Gómez A, Vaello ML, Valdivieso F, Mayor F. Phosphorylation of the 48-kDa subunit of the glycine receptor by protein kinase C. J Biol Chem. 1991;266:559–566. [PubMed] [Google Scholar]
- Savaskan E, Wirz-Justice A, Olivieri G, Pache M, Krauchi K, Brydon L, Jockers R, Müller-Spahn F, Meyer P. Distribution of melatonin MT1 receptor immunoreactivity in human retina. J Histochem Cytochem. 2002;50:519–526. doi: 10.1177/002215540205000408. [DOI] [PubMed] [Google Scholar]
- Slaughter MM. Inhibition in the retina. In: Chalupa LM, Werner JS, editors. The Visual Neurosciences. Vol. 1. Cambridge, MA, USA: MIT Press; 2004. pp. 355–368. [Google Scholar]
- Spadaro F, Cecchetti S, Sanchez M, Ausiello CM, Podo F, Ramoni C. Expression and role of phosphatidycholine-specific phospholipases C in human NK and T-lymphocyte subsets. Eur J Immunol. 2006;36:3277–3287. doi: 10.1002/eji.200635927. [DOI] [PubMed] [Google Scholar]
- Spadaro F, Ramoni C, Mezzanzanica D, Miotti S, Alberti P, Cecchetti S, Iorio E, Dolo V, Canevari S, Podo F. Phosphatidycholine-specific phospholipases C activity in epithelial ovarian cancer cells. Cancer Res. 2008;68:6541–6549. doi: 10.1158/0008-5472.CAN-07-6763. [DOI] [PubMed] [Google Scholar]
- Tapia JC, Espinoza F, Aguayo LG. Differential intracellular regulation of cortical GABAA and spinal glycine receptors in cultured neurons. Brain Res. 1997;769:203–210. doi: 10.1016/s0006-8993(97)00672-0. [DOI] [PubMed] [Google Scholar]
- Tosini G, Fukuhara C. Photic and circadian regulation of retinal melatonin in mammals. J Neuroendocrinol. 2003;15:364–369. doi: 10.1046/j.1365-2826.2003.00973.x. [DOI] [PubMed] [Google Scholar]
- Tzeng JI, Chen BC, Chang HM, Wang JJ, Sureshbabu M, Chien MH, Hsu MJ, Bien MY, Chiu WT, Hong CY, Lin CH. Involvement of phosphatidycholine-phospholipase C and protein kinase C in peptidolycan-induced nuclear factor-κB activation and cyclooxygenase-2 expression in RAW 264.7 macrophages. Pharmacol Res. 2010;61:162–166. doi: 10.1016/j.phrs.2009.09.005. [DOI] [PubMed] [Google Scholar]
- Vaello ML, Ruiz-Gómez A, Lerma J, Mayor F. Modulation of inhibitory glycine receptors by phosphorylation by protein kinase C and cAMP-dependent protein kinase. J Biol Chem. 1994;269:2002–2008. [PubMed] [Google Scholar]
- Vanecek J. Cellular mechanisms of melatonin action. Physiol Rev. 1998;78:687–721. doi: 10.1152/physrev.1998.78.3.687. [DOI] [PubMed] [Google Scholar]
- Van Wyk M, Wässle H, Taylor WR. Receptive field properties of ON- and OFF-ganglion cells in the mouse retina. Vis Neurosci. 2009;26:297–308. doi: 10.1017/S0952523809990137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wässle H. Parallel processing in the mammalian retina. Nat Rev Neurosci. 2004;5:1–11. doi: 10.1038/nrn1497. [DOI] [PubMed] [Google Scholar]
- Wässle H, Heinze L, Ivanova E, Majumdar S, Weiss J, Harvey RJ, Haverkamp S. Glycinergic transmission in the mammalian retina. Front Mol Neurosci. 2009;2:1–12. doi: 10.3389/neuro.02.006.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Way KJ, Chou E, King GL. Identification of PKC-isoform-specific biological actions using pharmacological approaches. Trends Pharmacol Sci. 2000;21:181–187. doi: 10.1016/s0165-6147(00)01468-1. [DOI] [PubMed] [Google Scholar]
- Weernink PAO, Han L, Jakobs KH, Schmidt M. Dynamic phospholipids signalling by G protein-coupled receptors. Biochim Biophys Acta. 2007;1768:888–900. doi: 10.1016/j.bbamem.2006.09.012. [DOI] [PubMed] [Google Scholar]
- Wiechmann AF. Differential distribution of Mel1a and Mel1c melatonin receptors in Xenopus laevis retina. Exp Eye Res. 2003;76:99–106. doi: 10.1016/s0014-4835(02)00230-0. [DOI] [PubMed] [Google Scholar]
- Wiechmann AF, Udin SB, Summers Rada JA. Localization of Mel1b melatonin receptor-like immunoreactivity in ocular tissues of Xenopus laevis. Exp Eye Res. 2004;79:585–594. doi: 10.1016/j.exer.2004.07.004. [DOI] [PubMed] [Google Scholar]
- Yu YC, Cao LH, Yang XL. Modulation by brain natriuretic peptide of GABA receptors on rat retinal ON-type bipolar cells. J Neurosci. 2006;26:696–707. doi: 10.1523/JNEUROSCI.3653-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang M, Cao LH, Yang XL. Melatonin modulates glycine currents of retinal ganglion cells in rat. Neuroreport. 2007;18:1675–1678. doi: 10.1097/WNR.0b013e3282f0b5a2. [DOI] [PubMed] [Google Scholar]
- Zaghloul KA, Boahen K, Demb JB. Different circuits for ON and OFF retinal ganglion cells cause different contrast sensitivities. J Neurosci. 2003;23:2645–2654. doi: 10.1523/JNEUROSCI.23-07-02645.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.