

Abstract
According to the pre-recruitment hypothesis, Escherichia coli SoxS activates transcription of the genes of the SoxRS regulon by forming binary complexes with RNA polymerase that scan the chromosome for class I and class II SoxS-dependent promoters. Previously, we showed that the a subunit’s C-terminal domain plays a role in activating both classes of promoter by making protein-protein contacts with SoxS; some of these contacts are made in solution in the absence of promoter DNA, a critical prediction of the pre-recruitment hypothesis. Here, we identified seven single alanine substitutions of region 4 of the σ70 subunit (σ70 R4) of RNA polymerase that reduce SoxS activation of class II promoters. With genetic epistasis tests between these σ70 R4 mutants and positive control mutants of SoxS we identified ten pairs of amino acids that interact with each other in E. coli. Using the yeast two-hybrid system and affinity immobilization assays we showed that SoxS and σ70 R4 can interact in solution, i.e., “off-DNA”. The interaction requires amino acids of the class I/II but not the class II positive control surface of SoxS and five amino acids of σ70 R4 that reduce activation in E. coli also reduce the SoxS-σ70 R4 interaction in yeast. One of the epistatic interactions that occur in E. coli also occurs in the yeast two-hybrid system, i.e., off-DNA. Importantly, we infer that the five epistatic interactions occurring in E. coli that require an amino acid of the class II surface occur “on-DNA” at class II promoters. Finding that SoxS contacts σ70 R4 both off- DNA as well as on-DNA is consistent with the pre-recruitment hypothesis. Moreover, SoxS is now the first example of an E. coli transcriptional activator that uses a single positive control surface to make specific protein-protein contacts with two different subunits of RNA polymerase.
Keywords: yeast two-hybrid system, SoxRS regulon, genetic epistasis, transcription activation
Introduction
During normal growth of Escherichia coli, most genes are transcribed constitutively and encode the general “housekeeping” functions required for growth. Transcription of these genes is effected by a holo-RNA polymerase (RNAP), which is comprised of the “core” subunits, α2, β, β’ and ω 1; 2, and the predominant σ factor, σ70 3; 4. The function of the core RNAP is to carry out transcription elongation 5, whereas the function of σ70 is to promote transcription initiation by recognizing and binding to specific DNA sequence elements lying within the various promoters 6. The most common recognition elements in these promoters are two hexameric sequences optimally separated by 17 bp and centered at −10 and −35 bp with respect to the start site for transcription initiation 7. Promoters may also contain an extended −10 sequence, which compensates for a −35 element with little resemblance to typical hexamers 8, and an “UP” element, which provides a third recognition element that enhances RNAP binding to promoters like the rRNA P1 promoters 9.
In contrast to the housekeeping genes, a number of other genes are transcribed at high levels only under unique environmental conditions of carbon or nitrogen source, the absence of specific nutrients, or stress. The elevated transcription of these genes is controlled by small molecule effectors or environmental signals that regulate the activity of the transcriptional activators that turn-on their target genes or the transcriptional repressors that turn-off their target genes 10; 11.
Recruitment is by far the most prevalent mechanism of transcription activation. In recruitment, a transcriptional activator binds to a specific DNA sequence within the promoter region of its target gene(s) 12. In some cases, the activator, e.g., catabolite gene activator protein (CAP, also known as cyclic AMP receptor protein, CRP) only binds its target sequence(s) following activation by an inducing signal, e.g., cyclic AMP 13; 14. Then, the DNA-bound activator “recruits” RNAP by making adhesive protein-protein interactions with it 15. Once recruited to the promoter, RNAP proceeds to form an open complex, then the initiation complex, and then the form that escapes the promoter 7; 16. Activators functioning by recruitment can stimulate one or more of the steps that lead to transcription initiation and promoter escape 17. Recently, evidence has been presented for a new mechanism of transcriptional activation, called “pre-recruitment” 18; 19 or “DNA scanning” 20. In this mechanism, the transcriptional activator forms a binary complex with RNAP in solution or when the two molecules collide during diffusion along the DNA. The activator-RNAP binary complexes then scan the chromosome for activator-dependent promoters that contain both a properly positioned binding site for the activator and promoter elements recognized by σ70. So far, only SoxS 18; 19, MarA 20 and Rob (E. F. Keen, III and R. E. Wolf, Jr., unpublished results) appear to activate transcription by this mechanism.
These three proteins form a closely related subset of the AraC/XylS family of bacterial regulatory proteins 21; 22. The three paralogs function as monomers 23–25, bind to the same highly degenerate 20 bp DNA sequence 26–29 and activate the transcription of the same set of ∼50 target genes 30–32, the SoxS/MarA/Rob regulon. Expression of the regulon provides resistance to superoxide stress, antibiotics, heavy metals, bile salts, and medium-chain fatty acids, as well as tolerance to organic solvents 30; 33–42.
Despite their high degree of amino acid sequence identity, the three proteins differentially activate transcription of the regulon’s genes 30–32, whose promoters fall into two classes 27; 28; 43. For example, SoxS efficiently activates transcription from class I promoters, wherein the soxbox lies upstream of the −35 hexamer and in the backward orientation, and from class II promoters, wherein the soxbox overlaps the −35 hexamer and resides in the forward orientation 43. In contrast, MarA and Rob efficiently activate transcription from class II promoters (e.g., fumC) but activate class I promoters (e.g., fpr) less well 23; 33; 44; 45 (E. F. Keen III and R. E. Wolf, Jr., unpublished results). Note that despite this general distinction between the three paralogs, each activator activates transcription of the individual target genes to different degrees.
Besides appearing to activate transcription by pre-recruitment, the three paralogs differ in another important way from most other transcriptional activators, namely, the means by which they become active and able to activate transcription. Thus, SoxS and MarA activate transcription of the member genes of the regulon only upon their de novo synthesis in the second stage of a two-gene, two-stage system 46; 47. For example, in the first stage of the SoxR/SoxS system, the 2Fe-2S centers of constitutively expressed SoxR become oxidized by the stress imposed by superoxide-generating, redox-cycling compounds like paraquat; this oxidation alters the conformation of SoxR so that it becomes an active activator, which then activates transcription of only one gene, soxS 48–52. In the second stage, newly synthesized molecules of SoxS form binary complexes with RNAP, which then scan the chromosome for soxboxes that lie within SoxS-dependent promoters, thereby distinguishing them from the sequence-equivalent but non-functional soxboxes that do not lie within SoxS-dependent promoters 18; 19. Thus, in this two-component system, SoxR functions as the sensor-transmitter of oxidative stress, while SoxS functions as the response-regulator.
In contrast to SoxS and MarA, Rob is expressed constitutively 53; 54 and, like most activators, it is inactive until activated by an inducing signal 40; 45; 55. What is unique about Rob is that its activity is regulated by a new mechanism, “sequestration-dispersal”. Thus, most of Rob’s 5,000–10,000 molecules/cell are sequestered into 3–5 immunostainable clusters and the clustered molecules are inactive 56; 57. Then, upon exposure to inducing compounds 40; 55, Rob molecules become dispersed and form binary complexes with RNAP, which then scan the chromosome for activatable promoters 57 (E.F. Keen III and R.E. Wolf, Jr., unpublished results).
A critical function of activators that bind DNA is how they interact with RNAP, either “on-DNA”, in the case of activators that function by recruitment, or both “off-DNA and on-DNA, in the case of activators that function by pre-recruitment. Much is known about the protein-protein contacts with RNAP made by activators that activate transcription by recruitment: they interact with determinants on either the C-terminal or N-terminal domains of the α subunits (αCTD and αNTD, respectively) or with amino acids lying within σ70 region 4 (σ70 R4), with the specific contact sites usually depending on the location of the activator binding site within the target promoter 10; 58; 59.
Because the discovery of the pre-recruitment mechanism is relatively recent, less is known about the nature of the protein-protein interactions between these activators and RNAP. However, evidence has been presented that SoxS 60, MarA 61 and Rob (E. F. Keen III and R. E. Wolf, Jr., unpublished results) differ from other activators in yet another mechanistic property: transcription activation of both class I and class II promoters requires previously unknown protein-protein interactions between the activator and the 265 determinant of the αCTD, the set of amino acids known to bind specifically to the UP element within rRNA promoters 15; 62. As such, these interactions enhance the respective defense responses by diverting RNAP from strong, UP element-containing promoters to the activator-dependent promoters 60 (E.F. Keen III and R.E. Wolf, Jr., unpublished results). Moreover, with SoxS and Rob, the interactions with the 265 determinant have been shown to require amino acids of the class I/II surface, mutations of which interfere with activation of both classes of promoters, but not amino acids of the class II surface, mutations of which interfere only with activation of class II promoters 60; 63 (E.F. Keen III and R.E. Wolf, Jr., unpublished results).
Since almost all transcriptional activators that activate transcription of class II promoters do so by making protein-protein contacts between the activator and σ70 R4, we thought it highly likely that SoxS would do so too. Moreover, we expected that, at the least, amino acid residue D75 of the class II surface of SoxS would interact with an amino acid of σ70 R4, since the homologous amino acids of AraC/XylS family members 64 RhaS (D241), RhaR (D276) and MelR (D261) are all known to require protein-protein contact with R599 of σ70 R4 65–67. Accordingly, in this work, we used a library of single alanine substitutions of σ70 R4 encompassing positions 590 – 613 to identify amino acids in this segment of RNAP that are important to transcriptional activation of SoxS-dependent promoters. Finding several, we then determined which of the positive control mutants of SoxS are epistatic to the substitutions in σ70 R4 that confer a defect in activation. The epistatic interactions identified in E. coli could represent interactions between SoxS and RNAP that occur either off-DNA in binary complexes or on-DNA at SoxS-dependent promoters (or both). Then, as a step toward determining whether SoxS can interact with σ70 R4 in solution, i.e., not on promoter-specific DNA, we examined the SoxS-σ70 R4 interaction in a yeast two-hybrid (Y2H) system 68 and by affinity immobilization assays (AIA). Finding that SoxS does interact with σ70 R4 in solution in the Y2H system, we then identified the alanine substitutions within the class I/II and class II surfaces of SoxS that disrupt the interaction in yeast and we determined which alanine substitutions of σ70 R4 that reduce transcriptional activation by SoxS in E. coli also disrupt the interaction in yeast. Then, we conducted genetic epistasis tests with the Y2H system and determined that one interaction between SoxS and σ70 R4 that occurs in E. coli also occurs in yeast. Lastly, we conducted AIA with full-length his6-σ70 and SoxS and found that immobilized his6-σ70 can capture SoxS from a crude cell extract. Together, the results of these analyses provide evidence for the existence of two types of interactions between SoxS and σ70R4, with one being able to occur off-DNA, e.g., within a SoxS-RNAP binary complex that scans the chromosome for SoxS-dependent promoters, and the other occurring on-DNA e.g., within a SoxS-RNAP binary complex residing at a SoxS-dependent promoter.
Results and Discussion
Effects of single alanine substitutions in σ70 R4 on SoxS-dependent transcription activation at class I and class II promoters
We introduced a library of single alanine substitutions at sixteen positions between amino acid residues 590 to 613 of σ70 R4 69; 70 into four derivatives of strain N7840 Δ(araBAD)714 [pBAD33-his6-SoxS]; two carried single-copy transcriptional fusions of lac to class I promoters fpr and zwf and two carried lac fusions to class II fusions fumC and micF. Wild type σ70 and the single alanine substitutions of it are expressed constitutively from the pGEX-2T-σ70 plasmids carrying the respective σ70 alleles 69; 70. After induction of his6-SoxS expression with 0.02% arabinose, the effects of the substitutions on SoxS-activated transcription of the four promoters were determined by assay of β-galactosidase activity. Since the abundance of the mutant σ70 proteins expressed from the leaky tac promoter of the pGEX-2T-σ70 plasmids approximately equals that of the chromosomally encoded wild type protein 70, we accepted, as have others 66, activity values below 80% of that obtained with wild type σ70 as indicating that the substitution within σ70 R4 confers a significant defect in transcription activation.
Table 1 shows that single alanine substitutions at positions R593, R596, R599 and R603 of σ70 R4 each reduce SoxS-activation of the class II promoters fumC and micF by more than 30%. Thus, as expected from prior work on activation of class II promoters by other members of the AraC/XylS family 65–67, σ70 R4 is required for SoxS-dependent activation of its class II promoters.
Table 1.
The effect of σ70 mutants on SoxS-dependent transcription activation at two class I promoters (fpr, zwf) and two class II promoters (fumC, micF).
| σ70 Region 4 Substitutions |
SoxS Dependent Transcription Activation | |||
|---|---|---|---|---|
| Class I SoxS Promoters | Class II SoxS Promoters | |||
| fpr | zwf | fum C | micF | |
| I590A | 87% ±6 | 144% ±11 | 151% ±17 | 111% ±12 |
| E591A | 92% ±8 | 106% ±19 | 110% ±9 | 90% ±2 |
| K593A | 115% ±4 | 127% ±23 | 44% ±8 | 55% ±3 |
| L595A | 115% ±7 | 117% ±14 | 137% ±17 | 106% ±5 |
| R596A | 82% ±3 | 106% ±8 | 38% ±8 | 59% ±5 |
| K597A | 90% ±5 | 112% ±10 | 125% ±8 | 106% ±11 |
| L598A | 136% ±7 | 109% ±10 | 129% ±17 | 105% ±10 |
| R599A | 94% ±8 | 125% ±15 | 63% ±9 | 69% ±3 |
| H600A | 123% ±9 | 132% ±11 | 96% ±12 | 117% ±9 |
| S602A | 101% ±6 | 107% ±10 | 116% ±15 | 115% ±9 |
| R603A | 88% ±4 | 88% ±2 | 27% ±4 | 43% ±5 |
| S604A | 89% ±2 | 86% ±4 | 110% ±6 | 117% ±6 |
| E605A | 85% ±2 | 70% ±5 | 88% ±4 | 103% ±6 |
| R608A | 84% ±6 | 37% ±5 | 112% ±17 | 95% ±10 |
| D612A | 83% ±3 | 39% ±7 | 85% ±5 | 110% ± |
| D613A | 88% ±1 | 88% ± 2 | 103% ±8 | 115% ±6 |
The experiments were conducted with four Δ(araBAD) 714 derivatives of strain N7840 [pBAD33-his6-SoxS] each lysogenic for a single copy λ prophage carrying one of the above SoxS-dependent promoters fused to lacZ and expressing the wild type allele of rpoD from the chromosome. Each of the four fusion strains was transformed with a member of the pGEX-2T-σ70 library composed of wild type σ70 and single alanine substitutions of σ70 R4 mutants; the plasmid-encoded rpoD allele is expressed constitutively. As described in Materials and Methods, the cultures were grown in LB medium in polypropylene blocks to A600 = 0.1–0.2, treated with 0.02% arabinose to induce SoxS synthesis and β-galactosidase activity was determined when the A600 of the cultures reached 0.6–0.8. The average values and standard errors of the mean were determined. Activity values obtained with the σ70 R4 mutants were expressed as a percent of the value produced by wild type σ70. As described in the text, when a σ70 R4 mutant produced an average activity value that is below 80% of the value produced by pGEX-2T-σ70 carrying the wild type rpoD allele, the σ70 R4 mutant was considered to confer a defect in SoxS-dependent transcription activation of that promoter. Mutant values are highlighted. The activity values produced by the four fusion strains carrying pGEX-2T-σ70 and expressing the wild type rpoD allele were 1333 and 2720 units for the class I fpr and zwf promoters, respectively, and 586 and 2520 units for the class II promoters micF and fumC promoters, respectively.
Importantly, none of the substitutions of σ70 R4 reduce SoxS activation of fpr (Table 1), a conventional class I promoter whose binding site lies in the backward orientation and 15 bp upstream of the putative −35 promoter element, presumably because SoxS bound to its binding site is too far from σ70 R4 to interact with it even though SoxS bends DNA about 35° 23; 25. This absence of effects of substitutions of σ70 R4 on SoxS activation of this class I promoter contrasts with the arrangement of protein-protein contacts at class I CAP-dependent promoters, e.g., lac, where the αCTD is wedged between DNA-bound CAP and promoter-bound RNAP such that the 287 determinant of the αCTD interacts with Activation Region 1 of CAP’s promoter-proximal subunit while the αCTD’s 261 determinant interacts with amino acid residues within σ70 R4 71. Indeed, the results with fpr are consistent with the fact that SoxS interacts with the αCTD by directly contacting its 265, DNA-binding determinant, thereby placing the αCTD in a position where it cannot interact with σ70 R4 60.
Table 1 also shows that alanine substitutions at positions E605, R608 and E612 of σ70 R4 significantly reduce Sox-dependent activation of the zwf promoter, even though SoxS binds 7 bp upstream of zwf’s −35 promoter hexamer 26; 63. A similar situation pertains to transcription activation by Ada of the aidB, ada, and alkA promoters wherein different amino acids of σ70 R4 are required for activation by Ada, depending on whether the binding site overlaps the −35 region or whether the site is 5–7 bp upstream of the promoter hexamer 72. For SoxS, its binding at the zwf promoter is likely able to interact with σ 70 R4 because its binding to DNA induces a ∼35° bend in the DNA 23; 25 that would bring SoxS and RNAP close enough together to allow protein-protein contact.
Previously, zwf has been classified as having a class I promoter SoxS-dependent promoter because its SoxS binding site lies upstream of the −35 hexamer 43, even though the site lies in the forward orientation like those of canonical class II SoxS-dependent promoters. Now, with the zwf promoter behaving like a class II SoxS-dependent promoter because its activation by SoxS requires amino acid residues of σ70 R4, it would be appropriate to reclassify zwf as a non-conventional class II promoter, i.e. class II*.
All of the amino acid residues of σ70 R4 that cause a defect in transcription activation by SoxS are charged, with five being positively charged and two being negatively charged. As mentioned above, some of the residues that are defective in SoxS-dependent transcription activation are also important to transcription activation by other AraC/XylS family members at their respective promoters. Thus, residues K593 and R599 of σ70 R4 are important to transcription activation by RhaS and RhaR 65; 66 while these two residues and R596 are also important for MelR-dependent transcription activation 67. In addition, residues R599 and R603 of σ70 are important to Rob-dependent transcription activation (E.F. Keen III and R.E. Wolf, Jr., unpublished results). Interestingly, K593 not only makes protein-protein contact with transcription activators but it also interacts specifically with DNA 73, suggesting that certain amino acids of σ70 play a dual role of interacting with promoter DNA and interacting with activators. Importantly, several residues of σ70 R4 that are required for transcription activation of SoxS promoters are also necessary for transcription activation by CAP, FNR and λcI of their target promoters 70; 73–76. Thus, in agreement with the observation of Lonetto et al 70, the same amino acid residues of σ70 R4 are used to make specific protein-protein contacts with many different transcription activators
In addition to amino acid residues within the helix-turn-helix motif of region 4.2 of σ70, some of the residues important to transcription activation by SoxS reside in the C-terminal tail, viz., R603, E605, R608 and D612. Since most activators that interact with σ70 R4 do so by making protein-protein contacts with region 4.2 and do not contact the C-terminal tail, we infer that the mechanism of transcription activation by SoxS at certain promoters requires these additional contacts. For example, the position of the soxbox with respect to the −35 hexamer varies from promoter to promoter 43. Thus, the soxbox of the fumC promoter overlaps the entire −35 hexamer and extends 4 bp beyond it whereas the soxbox of the zwf promoter lies 7 bp upstream of the hexamer. Accordingly, SoxS bound at zwf might only be able to contact amino acid residues of the C-terminal tail while SoxS bound at fumC might only be able to contact a subset of amino acids in region 4.2. Alternatively, activation by pre-recruitment might require two types of protein-protein contacts between SoxS and RNAP, contacts that mediate binary complex formation and contacts necessary for transcription initiation that are made after the binary complex binds the promoter.
Identification of amino acids of σ70 R4 that make protein-protein contact(s) with amino acids of the class I/II and/or class II positive control surfaces of SoxS in the activation of transcription from class II and class II* promoters
To identify protein-protein interactions critical to SoxS activation of class II and class II* promoters, we carried out genetic epistasis tests, also called “loss of contact” 66; 77 experiments, using the single alanine substitutions of σ70 R4 that confer defects in transcription activation at these promoters (Table 1) and the single alanine substitutions of SoxS that display a positive control phenotype 63. Before beginning the epistasis tests, we needed to determine whether the presence of plasmid pGEX-2T-σ70 interfered in some way with the reduction in SoxS-dependent transcription conferred by the SoxS positive control substitutions 63. To do this, we transformed plasmid pGEX-2T-σ70 into the strains carrying lac fusions to the fumC, micF, and zwf promoters. We then introduced plasmid pBAD33-his6-SoxS and its mutant derivatives carrying single positive control mutations of the class I/II (K5A, D9A, K30A, S31A, and V45A) or class II (F74A, D75A, M78A, D79A and Q85A) surfaces 63. (The class I/II surface lies above the N- terminal helix-turn-helix DNA binding motif of SoxS while the class II surface lies above the C-terminal motif 63). We then assayed β-galactosidase activity in the three sets of 11 strains. All of the positive control mutants reduced transcription activation at the three promoters and did so by amounts similar to those obtained previously in fusion strains that express σ70 only from the chromosomal rpoD gene (data not shown) 63.
For the epistasis tests, we prepared three sets of strains carrying each of the σ70 R4 substitutions that reduce transcription activation of a given promoter in combination with each of the ten substitutions of SoxS that confer defects in positive control. The activity of β-galactosidase was determined for each combination of relevant σ70 R4 substitution and SoxS positive control mutant at each of the three promoters. For each combination, we also used β-galactosidase assays to determine the effect of the σ 70 R4 substitution on activation of the promoter by wild type SoxS and the effect on SoxS-dependent transcription of the SoxS positive control substitution in the presence of pGEX-2T-σ70 carrying wild type σ70.
The logic underlying the epistasis tests is that if the side group of a specific amino acid residue of one protein, e.g., σ70, makes a protein-protein interaction with the side group of a specific amino acid residue of a second protein, e.g., SoxS, then the effect of combining single alanine substitutions at those positions of the two proteins should be no greater than the more severe effect of the two individual substitutions; such a combination of mutations is referred to as being epistatic. On the other hand, if the side group of a specific amino acid residue of one protein does not make protein-protein contact with the side group of a specific amino acid residue of the second protein, then the effect of combining single alanine substitutions at those positions of the two proteins should be independent of each other, with the result that the effect of the combination of mutations is greater than that of either substitution alone; in this case, the combination is non-epistatic. Figure 1 shows a representative example of an epistatic and a non-epistatic interaction at each promoter.
Figure 1.

Representative epistatic and non-epistatic interactions between alanine substitutions of σ70 R4 and alanine substitutions of SoxS that individually confer a defect in transcription activation at a given SoxS-dependent promoter. The experiments were carried out as described in Materials and Methods. A. The left panel shows that at the fumC promoter amino acid substitution R599A of σ70 R4 is epistatic to amino acid substitution D9A of SoxS, while the right panel shows that R599A is not epistatic to D79A of SoxS. B. The left panel shows that at the micF promoter, amino acid substitution R603A of σ70 R4 is epistatic to amino acid substitution D75A of SoxS, while the right panel shows that R603A is not epistatic to M78A of SoxS. C. The left panel shows that at the zwf promoter amino acid substitution D612A of σ70 R4 is epistatic to amino acid substitution K30A, while the right panel shows that D612A is not epistatic to S31A of SoxS.
Table 2 summarizes the epistasis data obtained with the seven single alanine substitutions of σ 70 R4 that reduce SoxS-dependent transcription activation at the two class II promoters and the one class II* promoter examined. Epistasis at one or more promoters was obtained with five of the seven mutants of σ 70 R4. We found a total of ten epistatic interactions when we combined the ten positive control mutants of SoxS with σ 70 R4 substitutions K593A, R596A, R599A and R603A and examined their effects at the micF and fumC promoters (Table 2). (The zwf promoter was not included in these tests because, as shown in Table 1, these four σ 70 R4 substitutions have no effect on SoxS activation of this promoter.)
Table 2.
Summary of the epistatic interactions in E. coli between amino acid residues of σ70 R4 and amino acid residues of SoxS at specific SoxS-dependent promoters.
| Promoter (Class) | σ70 R4 Amino Acid Residue Conferring Activation Defect |
Interacting SoxS residue | |
|---|---|---|---|
| Class I/II Surface |
Class II Surface | ||
| fumC (Class II) | K593 | S31 | M78, D75 |
| micF (Class II) | S31 | M78, D79 | |
| fumC (Class II) | R596 | None | None |
| micF (Class II) | None | None | |
| fumC (Class II) | R599 | D9 | D75 |
| micF (Class II) | None | None | |
| fumC (Class II) | R603 | None | None |
| micF (Class II) | None | D75, D79 | |
| zwf (Class I) | E605 | None | None |
| zwf (Class I) | R608 | D9 | None |
| zwf (Class I) | D612 | K30 | None |
| fpr (Class I) | None | None | None |
All amino acids of σ70 R4 that confer a defect in SoxS-dependent transcription activation at a given promoter when substituted with alanine are listed even if no interacting partners were identified. Epistasis was determined as described in the text. The amino acid residues of the two SoxS positive control surfaces that are epistatic partners of the σ70 R4 substitutions are listed according to the promoter at which the epistasis was observed.
Several epistatic interactions between SoxS and σ70 R4 are particularly interesting. (1) Substitution S31A of the class I/II surface of SoxS and M78A of the class II surface are epistatic to σ 70 R4 K593A at both the fumC and micF promoters; since S31 and M78 lie in two different positive control surfaces of SoxS, their interactions with K593 probably occur in different SoxS-RNAP complexes. (2) K593A is also epistatic to D75A and D79A of the class II surface at the fumC and micF promoters, respectively. Using two different amino acids of SoxS for activation of the two class II promoters may reflect the difference in the position of the two soxboxes, with the 3’ end of the fumC soxbox extending 5 bp further downstream of the −35 region of its promoter than does the micF soxbox 43; moreover, this difference may indicate that these particular interactions occur on DNA. (3) D9A of the class I/II surface and D75A of the class II surface are epistatic to σ 70 R4 R599A at the fumC promoter but not at the micF promoter; as with the epistatic interactions of S31A and M78 of SoxS with K593A of σ70 R4, these interactions are also likely to occur in different SoxS-RNAP complexes. (4) Since D75A of SoxS is epistatic to both K593A and R599A of σ70 R4 at the fumC promoter, it is unclear whether these interactions occur simultaneously or in different complexes. The same is true of D79A of SoxS, which is epistatic to K593A and R603A at the micF promoter. (5) In addition to being epistatic to R599A of σ70 R4 at the fumC promoter, D9A of the class I/II surface of SoxS is also epistatic to R608A at the zwf promoter; thus a single amino acid of a positive control surface of SoxS interacts with different amino acids of σ70 R4, presumably because the fumC soxbox overlaps the promoter’s −35 hexamer by 4 bp while the zwf soxbox lies 7 bp upstream of its promoter’s −35 hexamer. Indeed, the position of the Ada binding site with respect to the −35 region of its class II-type also affects the specific amino acids of sigma with which it makes protein-protein interactions. (6) No epistatic partners were found for R596A and E605A of σ70 R4. However, with charge reversal R596E and the experimental design of Westblade et al. 78, we determined that the role of the side group of R596 is to maintain the tertiary structure of σ70 R4 so that other amino acid residues are optimally positioned to bind to SoxS (data not shown). The same approach did not indicate a similar role for D605 and thus the basis for its ability to reduce SoxS-dependent transcription at the zwf promoter remains unknown. Lastly, we note that just as the amino acid homologs of D75 of SoxS in RhaS (D241), RhaR (D276), and MelR (D261) contact R599 of σ70 R4 at their respective target promoters 65–67 so too does D75 of SoxS contact R599 at the fumC promoter.
Determination of whether SoxS and σ70 R4 can interact with each other in vitro
As described in Materials and Methods, we bound his6-σ70 R4 to Ni-NTA resin and then, after washing, the resin was incubated with the soluble fraction of a cell extract prepared from a culture expressing native SoxS. Figure 2 shows that elution of N-his6-σ70 R4 from the Ni-NTA agarose resin leads to the co-elution of native SoxS. Thus, SoxS must have been captured by resin-bound N-his6-σ70 R4 because SoxS was absent when the resin was initially incubated with soluble material from a control extract lacking N-his6-σ70 R4 (data not shown). Importantly, this experiment demonstrates that SoxS can indeed interact with N-his6-σ70 R4 in solution and in the absence of specific promoter DNA. Accordingly, we proceeded to determine whether SoxS and σ70 R4 can interact in the Y2H; finding that they do, we investigated the nature of and the requirements for the interactions.
Figure 2.
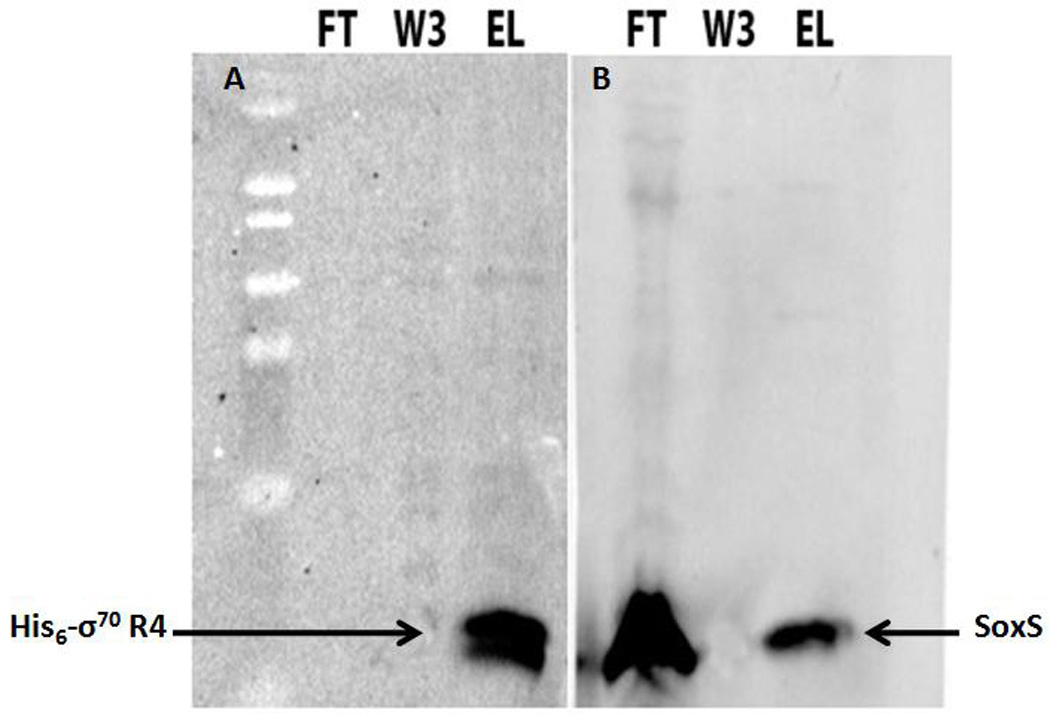
AIA of the protein–protein interactions between SoxS and amino acids 541–613 of σ70 R4. The immobilization of his6-σ70 R4 on Ni-NTA resin and its incubation with a cell-free extract containing wild type SoxS were carried out as described in Materials and Methods, as was the subsequent SDS-PAGE and western blotting of the respective fractions obtained after application of the SoxS-containing extract. FT, flow-through fraction; W3, third wash fraction; EL, eluate. Arrows indicate the location of his6-σ70 R4 and SoxS. A. The blot probed with antibody directed against the his6 tag. Samples FT and W3 show that all of the σ70 R4 bound to the column. Sample EL shows that all of the σ70 R4 was eluted from the Ni-NTA resin upon incubation with 0.5 M imidazole. B. The blot probed with anti-SoxS antibodies. The FT sample shows that a large portion of SoxS was not bound to the column while the W3 fraction reveals that all of the non-specifically bound SoxS was washed away. Sample EL shows that SoxS was eluted from the Ni-NTA resin column with 0.5 M imidazole. Taken together, the two blots suggest that σ70 R4 binds the column, that the bound protein retains SoxS, and that both proteins are co-eluted.
Analysis of protein-protein interactions in solution between SoxS and σ70 R4 with the Y2H system
In the Y2H system used here, three recombinant plasmids were introduced into strain EGY48, which carries nutritional markers complemented by genes carried on the plasmids. (1) The DNA binding domain (DBD) of the LexA repressor was fused in-frame to SoxS to form the “bait”-expressing plasmid pGILDA-SoxS. (2) An 88 amino acid peptide encoding an activation domain, B42AD, was fused in-frame to amino acids 531–613 of σ70 to form the “prey”-expressing plasmid pB42AD-σ70 R4. Expression of the bait and prey proteins is under control of a fully functional, galactose-inducible GAL1 promoter. (3) Plasmid p8oplacZ carries a lacZ gene whose transcription is controlled by a minimal GAL1 promoter comprised of a TATA box and eight upstream operators that bind LexA’s DBD. Thus, lacZ serves as the reporter of transcription activation from this promoter, since β-galactosidase will only be produced when the LexA-SoxS bait protein binds the operators and recruits the B42AD activation domain to the promoter through protein-protein interaction with the B42AD-σ70 R4 prey protein. Three negative control strains were also prepared as derivatives of strain EGY48[p8oplacZ] carrying the following plasmid pairs: pGILDA + pB42AD; pGILDA-SoxS + pB42AD; pGILDA + pB42AD-σ70 R4. Strain EGY48[p8oplacZ] carrying pGILDA-SoxS and pB42AD-αCTD served as the positive control, since the interaction between SoxS and the αCTD was first revealed in Y2H experiments using this strain 60. In this Y2H system, expression of β-galactosidase activity can be assessed qualitatively by the color of colonies formed by cells plated on triple drop-out medium containing raffinose (raf) as the carbon source, galactose (gal) as the inducer and X-gal as a chromogenic substrate; expression can also be assessed quantitatively by direct assay of β-galactosidase activity in cells growing under the same inducing conditions. The data presented in Table 3 show that all three negative control strains form white-colored colonies on the raf + gal + X-gal indicator plates and thus none of the negative controls produced false positives. In contrast, the test transformants co-expressing the LexA-SoxS bait protein and the B42AD-σ70 R4 prey protein formed pale blue-colored colonies on the indicator plate, indicating that transcription of the 8oplacZ reporter had been activated. Thus, the plate phenotype of the test transformants provides evidence for a protein-protein interaction between SoxS and σ70 R4. We note, however, that the strength of the SoxS-σ70 R4 interaction appears to be less than that of the interaction between SoxS and the αCTD, since cells co-expressing LexA-SoxS and B42AD-αCTD formed colonies on the indicator plate that were a darker blue color, as observed previously 60.
Table 3.
Demonstration of protein-protein interactions between SoxS and σ70 R4 in the Y2H system and the effects of positive control substitutions of SoxS on the interaction
| LexA Fusion (Bait) |
B42 Fusion (Prey) |
Colony Color | Induction Ratio |
Western Analysis |
|
|---|---|---|---|---|---|
| pGILDA (empty) | pB42AD (empty) | White | 2 | → β-Tubulin | |
| pGILDA-SoxS | pB42AD (empty) | White | 2 | N.T. | |
| pGILDA (empty) | pB42AD-σ70R4 | White | 2 | N.T. | |
| pGILDA-SoxS | pB42AD-αCTD | Blue | 25 | N.T. | |
| pGILDA-SoxS | pB42AD-σ70R4 | Pale Blue | 14 | → LexA-SoxS | |
| pGILDA-SoxS I/II− +II− (5, 9, 30, 31, 45, 74, 75 78, 79, 85) |
pB42AD-σ70R4 | White | 2 | ||
| pGILDA-SoxS I/II− (9, 30, 31, 45) |
pB42AD-σ70R4 | White | 2 | ||
| pGILDA-SoxS II− (74, 75, 78,79, 85) |
pB42AD-σ70R4 | Pale Blue | 14 |
Strain EGY48[p8oplacZ] carrying the indicated bait and prey fusions expressed from pGILDA and pB42AD plasmids was prepared as described in Materials and Methods. Colony color was determined after 48 hr of growth at 30 °C on SD/gal/raf/-his/-trp/-ura/X-gal plates. The β-galactosidase activities in the respective strains were determined during growth in liquid inducing medium, SD/gal/raf/-his/-trp/-ura, and liquid non-inducing medium, SD/glu/-his/-trp/-ura, as described in Materials and Methods. The induction ratios were computed by dividing the specific activity values obtained in inducing medium by the specific activity values obtained in non-inducing medium. Western blots were carried out to determine the relative expression levels of wild type and mutant SoxS proteins with antibodies directed against SoxS and β-tubulin. N.T., not tested.
Next, we quantified the strength of the interaction between SoxS and σ70 R4 by determining the effect of co-expressing the bait and prey fusion proteins on transcription of the lacZ reporter gene and comparing the results to the amounts of reporter gene transcription produced by the negative and positive control strains. Two exponential phase cultures of each strain were prepared, one growing under inducing conditions and the other growing under non-inducing Then, we determined the specific activity of β-galactosidase in each culture and calculated the induction ratio for each strain (the ratio of the amount of β-galactosidase activity produced under inducing conditions to the amount of activity produced under non-inducing conditions) The induction ratio for the test cultures co-expressing the LexA-SoxS and B42AD-σ70 R4 fusion proteins was 14, which is about sevenfold higher than the ratio obtained with the three negative control strains and about half of that obtained with cultures of the positive control strain that co-expressed the SoxS and αCTD fusion proteins (Table 3). Thus, the extent of activated transcription of the reporter gene by the co-expression of the LexA-SoxS and B42AD-σ70R4 fusion proteins relative to that of the negative and positive control strains correlates well with the respective plate phenotypes (Table 3). More importantly, both the qualitative and the quantitative tests of protein-protein interactions indicate that SoxS and σ70 R4 can interact in the absence of complete, holo-RNAP and without the specific binding of SoxS to soxbox DNA. Accordingly, SoxS and σ70 R4 appear to be able to interact in solution, i.e., off-DNA. In addition, the results indicate that the interaction between SoxS and σ70 R4 is only half as strong as the SoxS-αCTD interaction.
Effects of positive control mutations of SoxS on the interaction between SoxS and σ70 R4 in the Y2H system
As mentioned above, the characterization of our library of single alanine substitutions of SoxS enabled us to identify two classes of mutants that bind DNA normally but are defective in transcription activation, i.e., positive control mutants 63. The five class I/II mutants reduce transcription activation of both class I and class II SoxS-dependent promoters while the five class II mutants are defective only in activation of class II promoters. All of the positive control mutants bind and bend target DNA normally but they are leaky with respect to their effects on transcriptional activation of SoxS promoters: substitution K30A reduces transcriptional activation by threefold while the remaining positive control mutants are even more leaky, reducing activation by only about twofold 63.
The availability of the positive control mutants provided several important benefits to our analysis of the interaction of SoxS and σ70 R4 in the Y2H system. First, if mutants can be found that disrupt the interaction, then the interaction occurring in yeast is likely to be specific. Second, finding substitutions that prevent the interaction will identify amino acids important to complex formation in yeast. Third, since positive control mutants of SoxS that reduce complex formation in the Y2H system would do so by interfering with protein-protein contacts between SoxS as bait and σ70 R4 as prey, the defect in transcription activation conferred by these substitutions in E. coli would almost certainly be due to an alteration in protein-protein interaction between SoxS and σ70 R4. Lastly, if the SoxS-σ70 R4 interaction that occurs in solution in yeast is disrupted by positive control mutants of SoxS, the corresponding amino acids in wild type SoxS are likely to be involved in forming the binary complexes between SoxS and RNAP that play a critical role in the pre-recruitment mechanism.
Since the interaction of SoxS with σ70 R4 could use the class I/II surface, the class II surface or both surfaces, and since the individual alanine substitutions are leaky, we used the same three pGILDA plasmids carrying multiple positive control substitutions for this analysis as we used previously in our characterization of the interaction between SoxS and the αCTD 60. The SoxS protein in plasmid pGILDA-SoxS-class I/II− carries alanine substitutions D9A, K30A, S31A and V45A of the class I/II surface and was shown to be totally defective in transcription activation of both class I and class II promoters in E. coli 60. The SoxS protein in pGILDA-SoxS-class II− carries substitutions F74A, D75A, M78A, D79A and Q85A and is totally defective in transcription activation of class II promoters in E. coli but activates transcription of class I promoters nearly as well as wild type SoxS 60. Plasmid pGILDA-SoxS-classI/II− + class II− carries the nine substitutions listed above plus K5A of the class I/II domain; it is totally defective in transcription activation of both classes of promoter in E. coli 60. Importantly, the three multiply mutant proteins display the same over-expression toxicity in E. coli as wild type SoxS, an indication that they retain normal DNA binding in vivo 63, and as purified proteins they bind and bend DNA in vitro as well as wild type 60. These latter two tests indicate that the conformation of the proteins with the multiple positive control substitutions is like that of wild type SoxS 60.
Each of the three pGILDA-SoxS plasmids carrying multiple positive control substitutions of SoxS were co-transformed with pB42AD-σ70 R4 into strain EGY48[p8oplacZ]. The plate phenotypes of the respective strains and the induction ratios of β-galactosidase expression from the reporter gene in the three strains were determined as described above. The results are shown in Table 3 and are compared to the data obtained with wild type SoxS. No interaction with σ70 R4 was observed when the SoxS moiety carried the five substitutions of the class I/II surface and the five substitutions of the class II surface, as evidenced by formation of white-colored colonies and an induction ratio as low as that obtained with the two empty vectors. Thus, one or both of these surfaces of SoxS is required for the interaction with σ70 R4. The properties conferred by the other two mutant LexA-SoxS proteins enabled us to determine that only the class I/II surface is required for the SoxS-σ70 R4 interaction. Since co-expression of pGILDA-SoxS-class I/II− with pB42AD-σ70 R4 produced white-colored colonies and an induction ratio of 2, the class I/II surface is required for the interaction. In contrast, co-expression of pGILDA-SoxS-class II− with pB42AD-σ70 R4 had no effect on the interaction, since the colonies formed were of the same light blue color as with wild type LexA-SoxS and the induction ratio was the same as with wild type SoxS.
Thus, these data show that the interaction between SoxS and σ70 R4 is specific, because it is disrupted by a set of positive control mutants. In addition, the interaction depends on amino acid residues within the class I/II surface but not those within the class II surface; in turn, these data indicate that the substitutions of the class I/II surface that reduce transcription activation in E. coli do so by disrupting protein-protein interactions. Moreover, since interactions in the Y2H system occur in solution and in the absence of specific DNA binding, the ability of positive control substitutions of the class I/II surface to disrupt the interaction suggests that in E. coli the reduction in transcription activation by these positive control substitutions could be due to the disruption of interactions between SoxS and the σ70 R4 region of RNAP that occur in solution and in the absence of specific DNA binding. Accordingly, the positive control mutants could be interfering with the formation of the SoxS-RNAP binary complexes required for activation by pre-recruitment. Remarkably, positive control mutants of SoxS have the same effect on the interaction of SoxS with the αCTD 60 as with σ70 R4: positive control mutants of the class I/II surface disrupt the interactions with both the αCTD and σ70 R4 whereas positive control mutants of the class II surface have no effect. Thus, by this criterion, the same class I/II surface on SoxS makes protein-protein contact with both the αCTD and σ70 R4. Moreover, the existence of the two complexes in yeast suggests that the same two, mutually exclusive binary complexes form in E. coli, one through interaction between the class I/II surface and the αCTD and the other through interaction between the class I/II surface and σ70 R4. Logically, one binary complex might be used to find and bind to class I promoters while the other finds and binds to class II promoters.
The failure of the positive control mutants of the class II surface to disrupt the SoxS-σ70 R4 interaction in the Y2H system is also important with respect to the steps in transcription activation in E. coli. Since the class II mutants reduce transcription activation of class II promoters in E. coli but have no effect on the SoxS-σ70 R4 interaction in yeast, and since epistatic interactions were found in E. coli between some of these class II mutants and mutants of σ70 R4, we infer that the interactions between σ70 R4 and the class II surface of SoxS occur on SoxS-activatable promoter DNA.
Determination of whether single alanine substitutions of σ70R4 that reduce transcription activation in E. coli interfere with the interaction of SoxS and σ70 R4 in the Y2H system
Seven amino acid residues of σ70 R4 reduce SoxS-dependent transcription activation at the class II or class II* promoters in E. coli (Table 1). Although epistatic interactions in E. coli were found for only five of the σ70 R4 substitutions (Table 2), we wanted to determine which of the seven substitutions that reduce transcription activation affect the SoxS-σ70 R4 interaction in the Y2H system. Finding such an effect in yeast would identify σ70 R4 substitutions that are necessary for SoxS-σ70 R4 interactions in solution in E. coli and thus could be involved in the formation of binary complexes that would lead to transcription activation at one or more class II or class II* promoters.
To conduct this experiment, we first introduced the seven substitutions of σ70 R4 into prey plasmid pB42AD-σ70 R4 by site-directed mutagenesis and we transformed these plasmids into reporter strain EGY48[p8oplacZ] along with wild type bait plasmid pGILDA-SoxS. We then determined their plate phenotypes as described above. None of the plate phenotypes conferred by the respective mutant strains differed significantly from that of the strain co-expressing the wild type fusion proteins, i.e., all formed light blue-colored colonies, although some were slightly lighter and some were slightly darker than the strain with wild type σ70 R4.
To determine whether the σ70 R4 substitutions have a quantitative but not a qualitative effect on the SoxS-σ70 R4 interaction, we measured β-galactosidase activity under inducing conditions. Figure 3 shows that five of the σ70 R4 substitutions reduced transcription of the reporter to levels significantly below that of the wild type while substitutions K593A and R603A enhanced transcription of the reporter by 76% and 36%, respectively. The effects of the mutants were not due to differences in the abundance of the respective σ70 R4 proteins, as shown by qualitative western blots that detect the HA tag appended to B42AD (Figure 3). These data indicate that in E. coli amino acids R596, R599, E605, R608 and D612 of σ70 R4 are probably required for interaction with SoxS in solution, i.e., in binary complex formation. Because K593A and R603A produce enhanced interaction in yeast, they probably relieve inhibitory interactions conferred by the wild type amino acid residues. Nonetheless, the fact that they alter the interaction between SoxS and σ70 R4 in the Y2H system suggests that they too can function in solution. As discussed below, amino acid substitutions that produce values higher than that of wild type give inconclusive results in epistasis tests.
Figure 3.
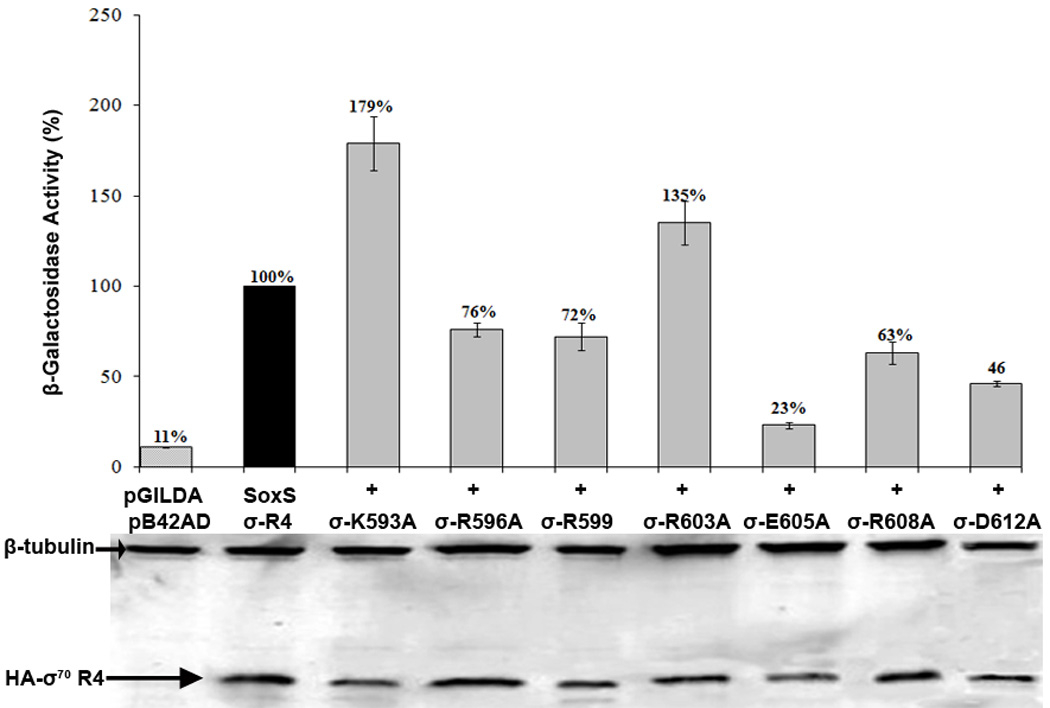
Effects in the Y2H system of single alanine substitutions of σ70 R4 that cause a defect in transcription activation at SoxS-dependent promoters in E. coli. Cultures of strain EGY48[p8oplacZ] carrying pGILDA-SoxS and pB42AD-σ70 R4 with wild type and mutant alleles of σ70 R4 were grown under inducing conditions and assayed for β-galactosidase activity as described in Materials and Methods. The specific activity of the strain expressing wild type σ70 R4 was taken as 100% and the activity values of the strains expressing the mutant alleles are given relative to wild type. As depicted at the bottom of the figure, cell extracts of each culture were subjected to western blotting with antibodies against the HA tag fused to B42AD-σ70 R4 and antibodies against β-tubulin, which served as an internal loading control.
Determination of whether single alanine substitutions of the class I/II surface of SoxS that are epistatic in E. coli to substitutions of σ70R4 interfere with the interaction of SoxS and σ70 R4 in the Y2H system
We wanted to conduct epistasis tests in the Y2H system to identify amino acid residues of the class I/II surface of SoxS that interact in solution with amino acid residues of σ70 R4. Since the work described in Table 3 only tested multiple positive control mutants in the Y2H system, we needed to introduce into pGILDA single alanine substitutions that were shown in the epistasis experiments conducted in E. coli to have an interacting partner in σ70 R4. We only used substitutions of the class I/II surface of SoxS and not those of the class II surface because the data in Table 3 show that the introduction of five substitutions of the class II surface into prey fusion B42AD-σ70 R4 has no effect on the interaction between SoxS and σ70 R4 in the Y2H system. Accordingly, we used site-directed mutagenesis to introduce the single alanine substitutions D9A, K30A and S31A into bait plasmid pGILDA-SoxS. We then co-transformed these singly mutant plasmids along with prey plasmid pB42AD-σ70 R4 into reporter strain EGY48[p8oplacZ]. After determining that the single positive control mutants have little effect on the plate phenotype, we then determined their effect on activated transcription of the reporter. We also conducted qualitative western blots with anti-SoxS antibodies and determined that the positive control substitutions do not substantially alter the abundance of SoxS in yeast.
As shown in Figure 4, substitutions K30A and S31A reduce activated transcription to 78% and 62% of wild type SoxS, respectively, and thus they play a role in the protein-protein interaction between SoxS and σ70 R4 in solution. Moreover, they are candidates to be tested for epistasis in the Y2H system. On the other hand, D9A enhances activated transcription by about 50%, probably by relieving an inhibitory interaction conferred by D9. As discussed below, conclusive results in epistasis tests cannot be obtained if one of the two substitutions produces a level of activated transcription significantly above that of wild type. Nonetheless, since D9A affects the interaction in the Y2H system between SoxS and σ70 R4, we infer that in E. coli it likely functions in solution (and perhaps also on-DNA).
Figure 4.
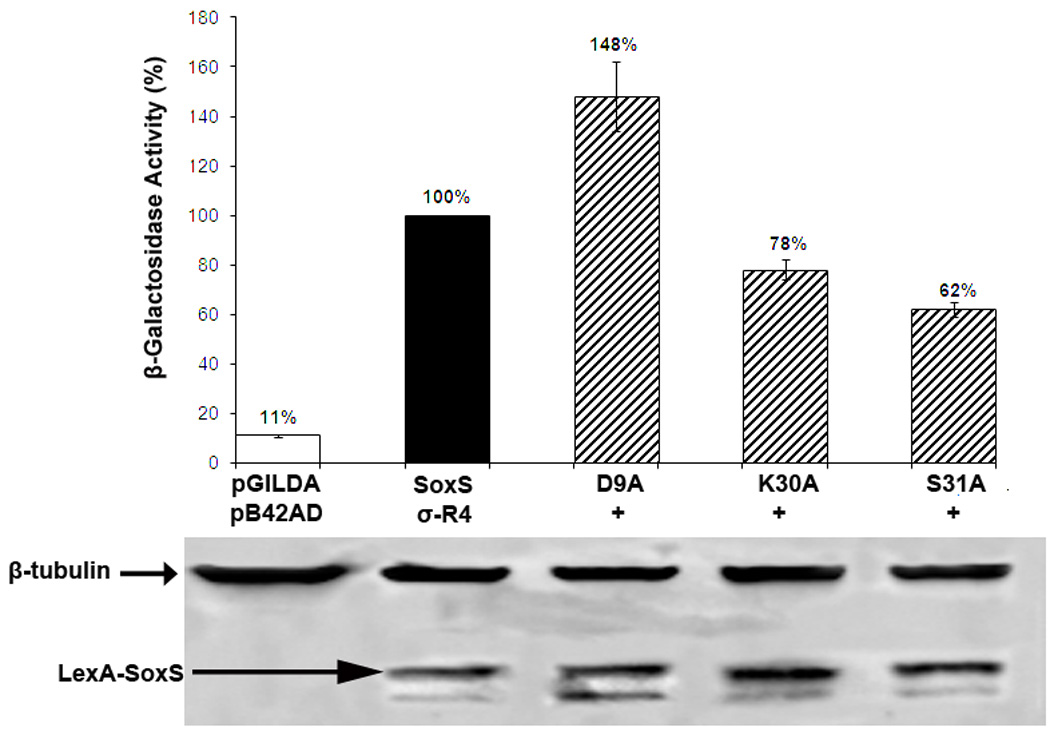
Effects in the Y2H system of single alanine substitutions of the class I/II surface of SoxS that confer defects in transcription activation at SoxS-dependent promoters and which display epistasis in E. coli with alanine substitutions of σ70 R4. Cultures of strain EGY48[p8oplacZ] carrying pB42AD-σ70 R4 and pGILDA-SoxS with wild type and mutant alleles of SoxS were grown under inducing conditions and assayed for β-galactosidase activity as described in Materials and Methods. The specific activity of the strain expressing wild type SoxS was taken as 100% and the activity values of the strains expressing the mutant alleles are given relative to the wild type. Cell extracts of each culture were subjected to western blotting with affinity-purified antibodies against SoxS and antibodies against β-tubulin, which served as an internal loading control.
Epistasis in the Y2H system
Having identified single alanine substitutions of SoxS and single alanine substitutions of σ70 R4 that interact epistatically in E. coli and also affect the interaction between SoxS and σ70 R4 in the Y2H system, we wanted to see whether the substitutions display epistasis in the Y2H system. If so, we could argue that an epistatic interaction that occurs in both yeast and E. coli likely occurs off-DNA in E. coli, i.e., in solution. Accordingly, our candidates for epistasis tests in the Y2H system were the single alanine substitutions of the class I/II surface of SoxS and the single alanine substitutions of σ70 R4 that participated with them in epistatic interactions in E. coli (Table 2) and which conferred defects in β-galactosidase expression from the reporter gene of the Y2H system (Table 3 and Figure 4).
In E. coli, epistasis involving positive control mutants of the class I/II surface was obtained with S31A + K593A, D9A + R599A, D9A + R608A and K30A + D612A. However, in the Y2H system, substitutions D9A of SoxS and K593A of σ70 R4 both enhance activated transcription of the lacZ reporter to levels considerably above that of the strain co-expressing the wild type fusion proteins and as such epistasis with them cannot be evaluated. This is because the activity produced by bait and prey substitutions acting non-epistatically, with one, e.g., D9A, producing an activity higher (e.g., 148%) than that of the wild type activity, and the other, e.g., R599A, producing an activity below (e.g., 72%) that of wild type, will be greater than the least active of the two (i.e., 107% vs. 72%); as such, a non-epistatic interaction cannot be distinguished from an epistatic interaction. Accordingly, we only conducted epistasis tests with the combination of LexA-SoxS-K30A and B42AD-σ70 R4-D612A; as a negative control, we also conducted the test on the combination of LexA-SoxS-S31A and B42AD-SoxS-D612A that was expected to be non-epistatic. Since the single mutants of SoxS and σ70 R4 did not produce a significant effect on the plate phenotype, we only assessed epistasis by assay of β-galactosidase, a measure of activated transcription of the lacZ reporter gene.
Figure 5 shows that K30A of SoxS and D612A of σ70 R4 are indeed epistatic to one another in yeast, since the β-galactosidase activity produced by the strain co-expressing the bait and prey fusions carrying the K30A and the D612A substitutions, respectively, is not statistically different from the amount of activity produced by the strains co-expressing one wild fusion (e.g., LexA-SoxS) and one mutant fusion (e.g., B42AD-σ70 R4-K30A). As expected, S31A of SoxS and D612A of σ70 R4 are not epistatic to one another, since the β-galactosidase activity produced by the strain co-expressing the putative non-epistatic substitutions, S31A and D612A, respectively, produced a level of β-galactosidase that is statistically lower than either of the mutant fusions in combination with a wild type fusion (Figure 5). Thus, among the ten epistatic interactions in E. coli obtained between substitutions of SoxS and substitutions of σ70 R4 (Table 2), only the interaction between K30A of SoxS and D612A of σ70 R4 (e.g., 72%) also occurs in the Y2H system and therefore only this specific interaction has been demonstrated to occur in solution, i.e., off-DNA. As stated above, epistasis in E. coli that involves SoxS positive control mutants residing in the class II surface probably represents interactions that occur on promoter DNA, but substitutions of SoxS (e.g., D9A) or σ70 R4 (e.g., K593A) that reduce transcription in E. coli but enhance reporter gene transcription in yeast probably represent interactions that occur off-DNA in E. coli.
Figure 5.
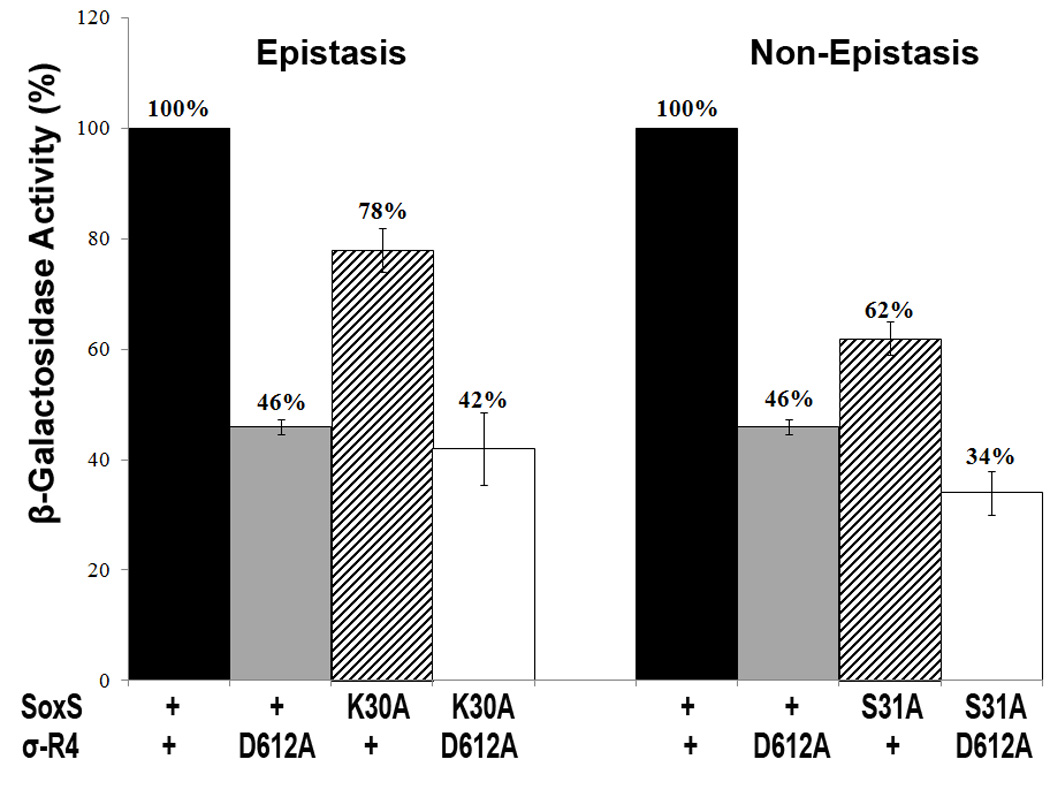
Representative epistatic and non-epistatic interactions in the Y2H system between alanine substitutions of SoxS and alanine substitutions of σ70 R4 that individually confer a defect in transcription activation of the lacZ reporter in the Y2H system. Cultures of strain EGY48[p8oplacZ] carrying plasmid pGILDA-SoxS with wild type and mutant alleles of SoxS and plasmid p42AD-σ70 R4 with wild type and mutant alleles of σ70 R4 were grown under inducing conditions and assayed for β-galactosidase activity as described in Materials and Methods. The specific activity of the strain expressing wild type SoxS and wild type σ70 R4 was taken as 100% and the activity values of the strains expressing the various mutant alleles are given relative to the wild type. An epistatic interaction was observed between σ70 substitution D612A and SoxS K30A, a mutant of the class I/II positive control surface. A two tailed t-test was carried out to determine whether the values for the double mutants are significantly different from the values for the corresponding single mutants. The value for the SoxS-K30A + σ70 D612A double mutant was not significantly different than that of the single mutant SoxS + σ70 D612A (p>0.05) and thus this interaction is epistatic; in contrast, the value for the SoxS-S31A + σ70 D612A double mutant was significantly different than that of the single mutant SoxS + σ70 D612A (p<0.05) and thus this interaction is non-epistatic.
It is important to note that even though only one epistatic interaction was found that occurs in both E. coli and yeast (K30A of SoxS with D612A of σ70 R4), this single example demonstrates that SoxS can make a specific protein-protein interaction with an amino acid of σ70 R4 in solution, i.e., off-DNA. Moreover, we argue that although only one specific interaction has been identified, we still know that SoxS interacts with σ70 R4 (Table 3) and full-length σ70 (see below) in solution, that this interaction requires amino acids of the class I/II of SoxS but not the class II surface (Table 3), that amino acid residues R596, R599, R605, R608 and R612 of σ70 R4 are required for the interaction (Figure 3), and that substitutions K593A and R603A enhance the strength of the interaction. In addition, we argue that even though the single epistatic interaction that has been shown to occur in solution does so in activating transcription from the atypical class II* zwf promoter (Table 2), the fact that amino acid substitutions K593A, R596A, R599A and R603A of σ70 R4 that reduce transcription activation of the fumC and micF promoters in E. coli also affect transcription activation of reporter gene expression in the Y2H system indicates that these residues play a role in binary complex formation in E. coli. As such, the data presented here make it clear that SoxS forms a binary complex off-DNA with σ70 R4 in yeast and with full-length σ70 in vitro (see below). Moreover, we point out that experiments supporting the conclusion that a given interaction occurs off-DNA do not rule out the possibility that the same interaction also occurs on-DNA.
Protein-protein interactions between SoxS and full-length σ70 demonstrated by AIA
The Y2H experiments (Figures 3–5) and the AIA (Figure 2) presented above have demonstrated specific protein-protein interactions between SoxS and σ70 R4 that occur in solution. Since the results could be an artifact of using σ70 R4 and not full length σ70 or because the results were obtained in yeast, a heterologous system, we conducted AIA using full-length σ70. Obtaining specific protein-protein interactions with SoxS and σ70 would not only rule out the possibility that the Y2H results are artifactual but would also provide independent verification of the interactions observed with σ70 R4 and the inferences drawn from them. In particular, if SoxS and full-length σ70 can be shown by AIA to make specific protein-protein contacts, then additional evidence will have been provided that the interactions can occur in solution and in the absence of SoxS-specific promoter DNA.
The following AIA experiments were carried out like those shown in Fig. 2 except that full-length his6-σ70 expressed from plasmid pHMK-his6-σ70 was used and it was unnecessary to run separate western blots to detect his6-σ70 and SoxS because they migrate to different positions during SDS-PAGE. Figure 6 shows that SoxS was present in the eluate when his6-σ70 was released from the Ni-NTA resin by treatment with a high concentration of imidazole. Thus, his6-σ70 immobilized on the Ni-NTA resin is able to capture wild type SoxS from an E. coli cell-free extract. Moreover, just as SoxS is able to interact with σ70 R4 in the Y2H system, so too is SoxS present in a crude cell extract able to interact with the full-length σ70 in AIA.
Figure 6.
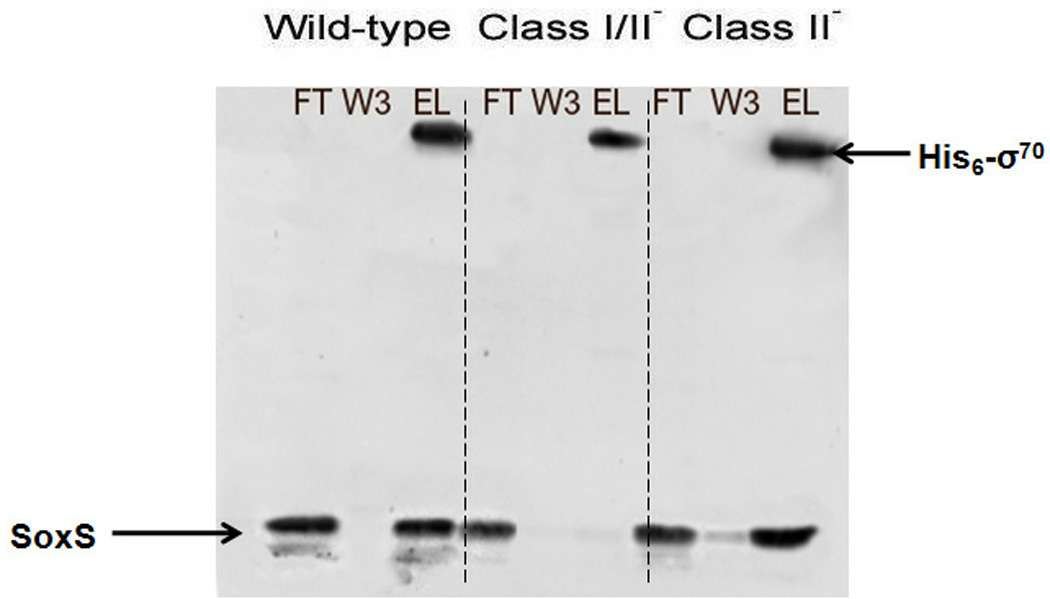
AIA of the protein–protein interactions between SoxS and full-length σ70 The immobilization of wild-type σ70 on Ni-NTA resin, the application of cell-free extracts containing wild type and SoxS proteins with multiple positive control substitutions, the recovery of the FT, W3 and EL fractions and western blotting of the fractions with antibodies directed against SoxS and the his6 tag of σ70 were carried out as described in Materials and Methods.
As a control for the specificity of the interaction between his6-σ70 and SoxS, we conducted AIA with the same three plasmids carrying multiple positive control substitutions of SoxS as were used in the Y2H assays (Table 3), i.e., (i) SoxS with the ten positive control substitutions that completely inactivate the class I/II surface and the class II surface; (ii) SoxS with the four positive control substitutions that completely inactivate the class I/II surface; and (iii) SoxS with the five positive control substitutions that completely inactivate the class II surface. The SoxS protein with the multiple mutations of the class I/II and class II surfaces was unable to interact with immobilized his6-σ70 (data not shown) as was the protein containing substitutions only of the class I/II surface (Figure 6). Importantly, the quintuple substitutions of the class II surface of SoxS did not interfere with the interaction (Figure 6). Thus, these results obtained with AIA fully agree with the corresponding assays conducted in the Y2H system. Moreover, they indicate that the class I/II domain is essential to the interaction with σ70 R4 even when it is part of full-length σ70. Indeed, when we prepared a mutant of his6-σ70 that carries two consecutive stop codons after position 529 and thus lacks region 4, i.e., his6-σ70(529), we found that after immobilization on Ni-NTA resin, it was unable to capture wild type SoxS in AIA (data not shown). This shows that σ70 R4 is necessary for the interaction in the AIA with full-length σ70 and the AIA experiment in which σ70 R4 was able to capture wild type SoxS (Figure 2) shows that σ70 R4 is sufficient for the interaction. Unfortunately, because of the relative weakness of the interaction between SoxS and σ70 R4, e.g., compared to the interaction of SoxS and αCTD, the effects of single alanine substitutions of σ70 R4 that reduce transcription in the Y2H system are too small to produce a significant effect in AIA (data not shown). Nonetheless, the data presented above demonstrate that SoxS makes protein-protein interactions with σ70 R4 and that the interaction requires the amino acids residues of the class I/II surface.
Concluding remarks
The data presented here and in the previous work by Shah and Wolf 60 are consistent with the pre-recruitment hypothesis in that they show that SoxS can indeed form binary complexes with RNAP in solution and in the absence of specific DNA binding. Of course, just because a binary complex between SoxS and RNAP can form in solution does not mean that it does form and then scans the chromosome for SoxS-dependent promoters. Hence, it will be interesting to determine whether activators known to function by recruitment, e.g., MelR 79, can also form binary complexes with RNAP in solution.
The data presented here and in the paper by Shah and Wolf 60 also indicate that SoxS makes two protein-protein contacts in solution with RNAP, one through an interaction with the αCTD and the other through an interaction with σ70 R4. However, since the same class I/II surface of SoxS is used for both interactions, the two interactions must lead to two mutually exclusive binary complexes. Thus, we infer that if SoxS functions by pre-recruitment, then one binary complex, e.g., the complex formed by the interaction with the αCTD, would scan the chromosome for class I SoxS-dependent promoters, while the other binary complex, e.g., the complex formed by the interaction with σ70 R4, would scan the chromosome for class II and class II* SoxS-dependent promoters. (We chose this partitioning because mutants of σ70 R4 have no effect on activation of class I promoters.)
Although it now appears that SoxS forms two different, off-DNA, binary complexes with RNAP as an initial step in transcription activation and that amino acids of SoxS, the αCTD and σ70 R4 participating in this initial step have been identified, determining the nature of the complexes in the subsequent steps leading to transcription initiation remains a challenge for future work.
Materials and Methods
Plasmid library of mutants of σ70 R4
Table 1 shows the library of 16 single alanine substitutions of σ70 R4 that was used for this study; the substitutions lie in the region of σ70 between positions 590 and 613. The gene encoding σ70 amino acids from positions 8 to 613 and its mutant derivatives was carried in plasmid pGEX-2T-σ70 and was obtained from C.A. Gross via S.M. Egan 69; 70 . After sequencing σ70 R4 of each member of the library at the UMBC DNA Sequencing Facility and finding several incorrect clones, we received replacements from V. A. Rhodius and W. Ross.
Determining the effect of single alanine substitutions of σ70 R4 on SoxS-dependent transcription in E. coli
The σ70 R4 library of single alanine substitutions was transformed into Δ(araBAD) 714 derivatives of strain N7840 ( Δ(argF-lac)U169 Δmar carrying transcriptional fusions of lac to the zwf, fpr, fumC and micF promoters 63 and plasmid pBAD33-his6-SoxS; the transformants were recovered by plating on LB plates containing ampicillin (100 µg/ml) and chloramphenicol (25 µg/ml). σ70 is expressed constitutively from the pGEX-2T plasmids while the expression of his6-SoxS is under control of the arabinose-inducible araBAD promoter. Plasmid pBAD33-his6-SoxS was prepared by subcloning the XbaI/HindIII fragment of high-copy plasmid pBAD18-his6-SoxS 63, which contains his6-SoxS, into the low-medium copy plasmid pBAD33 80 that had been digested with the same two enzymes. Triplicate cultures of the four fusion strains carrying the respective members of the σ70 R4 library and pBAD33-his6-SoxS were grown overnight at 37 °C in LB medium containing the two antibiotics in 96-well polypropylene blocks 81. Each overnight culture was diluted 1:200 in triplicate into the same medium and grown until the A600 of the cultures reached 0.1–0.2, at which point SoxS synthesis was induced with 0.02% arabinose, a concentration previously determined to be sufficient for maximum induction 63. Then, when the A600 of the cultures reached 0.6–0.8, samples were removed from each culture and the β-galactosidase activity was determined by our previously described high-throughput method 81. At least three independent experiments were carried out on each strain and the average values in Miller units for each experiment were averaged. The average values obtained with the mutants are presented as a percentage of the value obtained with the corresponding strain carrying the wild type plasmid. Standard errors of the means were calculated.
Epistasis tests in E. coli
The method described above for determining the effect of single alanine substitutions of σ70 R4 on SoxS-dependent activation of transcription of a specific promoter-lac fusion was also used for identifying epistatic interactions between σ70 R4 mutants and positive control mutants of SoxS. Thus, in these experiments, the strains harbored pBAD33-his6-SoxS carrying a positive control mutant of SoxS as well as pVR-σ70 carrying either the wild type or a mutant allele of σ70 82. Since all of the single alanine substitutions of SoxS that confer a positive control phenotype were prepared in pBAD18-his6-SoxS 63, we subcloned each substitution into pBAD33 as an XbaI/HindIII fragment as described above. Assays of β-galactosidase activity were done as described above and epistatic interactions were identified as described in the text.
Interaction between wild type and positive control mutants of SoxS and σ70 R4 or full-length σ70 as assessed in vitro by AIA
AIA was carried out by the method used previously for studying the interaction between wild type and mutant SoxS proteins and the full-length a subunit of RNAP 60. In the first set of AIA experiments, σ70 R4 was expressed with an N-terminal his6 tag from plasmid pSKB2-σ70 R4, kindly provided by L. F. Westblade. Since expression of his6-σ70 R4 is under control of a phage T7 promoter, the plasmid resided in E. coli strain BL21 (λDE3) and was maintained by growth in the presence of kanamycin (50 µg/ml); expression of T7 RNAP and thus his6-σ70 R4 was induced by treatment of the culture with 1 mM IPTG when the A600 of the 50 ml culture reached 0.2. The rest of this phase of the experiment with his6-σ70 R4 was carried out as described previously 60, except that the cell extract containing his6-σ70 R4 was incubated for 90 min with 250 µl of Ni-NTA agarose resin (Qiagen).
In the second phase of the AIA with his6-σ70 R4, the cell extracts containing untagged, wild type SoxS and untagged SoxS carrying multiple alanine substitutions of the class I/II and/or class II surfaces were prepared and incubated with the Ni-NTA resin to which his6-σ70 R4 had been bound as described previously 60, except that 50 ml cultures were used and buffer A contained 50 mM NaCl. Unbound material was removed by centrifugation and saved as the flow-through (FT) fraction. The resin was then washed three times with the last wash being saved as a control (W3). The resin was then incubated with buffer containing 500 mM imidazole to release N-his6-σ70 R4 and any proteins bound to it, i.e., SoxS; this elution fraction (EL) was also saved. Since SoxS and his6-σ70 R4 have about the same MW, ∼13 kD, and thus cannot be easily separated by SDS-PAGE and identified by western blotting, duplicate aliquots of the FT, W3 and EL samples were loaded onto two halves of the same 15% gel and after electrophoresis, the gel was subjected to western blotting as described 60. After transfer, the membrane was cut in half and one side was probed with mouse monoclonal antibodies directed against the his6 tag (Sigma) while the other half was probed with rabbit polyclonal anti-SoxS serum 18. The blots were then treated with secondary antibodies and developed as described 60. The resulting chemifluorescent signals were detected with a Molecular Dynamics Storm 860 PhosphorImager in fluorescence mode with a blue (450 nm) laser.
In the second set of experiments, plasmid pHMK-his6-σ70 encoding full-length σ70, kindly provided by R.R. Burgess 83, was transformed into strain BL21(λDE3) and the AIA was carried out as described above for the experiments with his6-σ70 R4. As a negative control, QuikChange mutagenesis of pHMK-his6-σ70 was used to introduce dual stop codons (TAGTGA) after codon 529 of σ70. AIA was carried out with this construct as described above.
Y2H system
The Y2H system used here was the same as that used in our previous work except that the prey was σ70 R4 rather than the αCTD 60. Thus, the parental plasmids for the bait and prey fusions were pGILDA and pB42AD, respectively, and the plasmid carrying the lacZ reporter was p8oplacZ; all were obtained from Clonetech, as was strain EGY48 (MATα his3 trp1 ura3 LexAop(x6)-LEU2), the strain of Saccharomyces cerevisiae used for all the Y2H experiments, and pop8lacZ, the plasmid encoding the gene that reports the transcriptional activity conferred by the bait and prey fusions.
As described by Shah and Wolf 60, plasmid pGILDA-SoxS expresses the bait as an in-frame fusion of the DBD of LexA to the N-terminus of SoxS. The following plasmids with multiple positive control mutations served as specificity controls for the interaction of SoxS with σ70 R4: pGILDA-SoxS-Class I/Class II− + Class II− with ten alanine substitutions of the class I/II and class II surfaces; pGILDA-SoxS-Class I/Class II− with four alanine substitutions in the class I/II surface; and pGILDA-SoxS-Class II− with five alanine substitutions in the class II surface 60. The single positive control substitutions D9A, K30A and S31A were introduced into pGILDA-SoxS by the same methods used to prepare the plasmids with the multiple positive control substitutions.
Plasmid pB42AD-σ70 R4 expresses the prey as an in-frame fusion between the B42AD activation peptide and σ70 R4. The DNA segment encoding σ70 R4 (amino acids 530–613) of plasmid pGEX-2T-σ70 was amplified by PCR with sense primer Sσ70R4–5’-CTTAGTGAATTCCTGCCGCTGGATTCTGCGACC-3’ and antisense primer ASσ70R4–5’-TACCGTCTCGAGCGATTAATCGTCCAGGAA-3’. The primers add-on restriction sites EcoRI and XhoI, respectively, whose recognition sequences are underlined. PCR was carried out for 30 cycles of 95 °C for 1 min, 55 °C for 30 sec and 72°C for 1 min. The PCR product was purified with a PCR cleanup kit (Qiagen) and digested with restriction enzymes EcoR and XhoI (New England BioLabs). The digestion product was subjected to gel electrophoresis and the DNA was extracted and purified with a Wizard Miniprep kit (Promega). The purified fragment was ligated to plasmid pB42AD digested with the same two enzymes and an aliquot of the ligation mixture was used to transform strain DH5α, which was plated onto LB agar medium containing ampicillin (50 µg/ml) and incubated at 37 °C. Colony PCR with vector-specific primers was used to identify clones containing insert fragments of the correct size. Then, plasmids were purified from these clones with Wizard minicolumns (Promega) and the DNA sequence of the candidates was determined. A clone with the correct sequence was named pB42AD-σ70 R4.
Six of the seven single alanine substitutions of σ70 R4 that reduce transcription activation in E. coli (K593A, R596A, R599A, R603A, E605A and R608A) were also PCR-amplified with the same primers as above from the members of the pGEX2T-σ70 library carrying the respective mutations; they were then cloned and their sequences determined as described above. Preparation of pB42AD-σ70 R4 carrying the D612A substitution required a different antisense primer, ASσ70R4-D612A-5’-TACCGTCTCGAGCGATTAATCAGCCAGGAA-3’, but otherwise the cloning, identification, and sequencing used the same procedures as for the other mutants.
The procedures for growth and manipulation of yeast were those described in Clonetech’s Matchmaker system manual as modified by Shah and Wolf 60. S. cerevisiae strain EGY48 was grown at 30 °C in SD medium, a complete minimal medium prepared to contain all the nutrients required for growth of yeast or to lack specific nutrients to allow selection for nutritional markers. Thus, EGY48 carrying the reporter plasmid pop8lacZ and plasmids pGILDA and pB42AD and their various wild type and mutant bait and prey derivatives were grown in SD medium lacking uracil, histidine and tryptophan (“triple drop-out medium”) and containing 2% (w/v) glucose as the carbons source for non-inducing conditions or 2% (w/v) gal plus 1% (w/v) raf for growth under inducing conditions.
Determination of plate phenotypes and assay of β-galactosidase in the Y2H system
The method for determining on plates whether co-expression of a given combination of bait and prey fusions produced a functional activator able to activate expression of the lacZ reporter was carried out as described previously by Shah and Wolf 60, as was quantification of the extent of lacZ reporter gene transcription under inducing and non-inducing conditions. The plate phenotypes were determined on SD/gal/raf/-uracil/-histidine/-tryptophan/X-gal plates. Quantification of reporter gene expression following co-expression of the various bait and prey fusions was done by determining the specific activities of β-galactosidase in Miller units from duplicate assays of samples taken from duplicate cultures, with the final average values calculated from at least three experiments conducted on different days. The induction ratios were calculated by dividing the average specific activity values obtained for cells grown under inducing conditions by the average specific activity values obtained for cells grown under non-inducing conditions.
Statistical Analyses
The analysis toolkit of Microsoft Excel was used to carry out two-tailed t-tests of the significance of differences in the effects of mutations on transcriptional activation. For example, a t-test was conducted to determine whether the effect of a double mutant combination of putative epistatic partners, e.g., σ70 D612A + SoxS K30A, differs significantly from the effect of the single mutant of the pair that conferred the greatest defect e.g., D612A. The null hypothesis was that the effects of the single mutant and the double mutant combination are not significantly different at a confidence interval of 95% (α=0.05). For epistatic interactions, the calculated value was p>0.05 and therefore the null hypothesis was accepted. For the non-epistatic interactions, the calculated value was p<0.05 and therefore the null hypothesis was rejected.
Western blotting of extracts prepared from strains used for reporter gene assays in the Y2H system
Western blots were used to determine qualitatively whether the effect of mutations of SoxS or σ70 R4 that either reduced or enhanced reporter gene expression in the Y2H system was due to a corresponding alteration in the abundance of the mutant protein compared to the wild type. The procedure employed was that described by Vallabhaneni and Farabaugh 84. Cultures were grown under conditions identical to those used when the various strains were grown for determination of the effect of mutations of SoxS and σ70 R4 on reporter gene expression. Thus, an overnight culture of each strain grown in non-inducing medium was diluted 1:200 into 10 ml of inducing medium and the culture was grown to A600∼0.8. The cells from each culture were collected by centrifugation at 5000g, resuspended in 1 ml water, transferred to a 1.5 ml microfuge tube and collected again by centrifugation. After adding acid-washed glass beads (425–600 mm) (Sigma-Aldrich) in an amount approximately equal to the volume of the cell pellet, the mixture was resuspended in 100 µl of lysis buffer (25mM Tris pH=7.5, 1mM DTT, 0.1% Triton X-100, 0.15M NaCl, 1mM EDTA, and a protease inhibitor cocktail (10 µl/ml) (Sigma-Aldrich). The mixture was subjected to 7 rounds of vortexing for 30 sec with rests of 30 sec on ice between each treatment. The cell lysate was cleared by centrifugation at 16000g for 5 min at 4 °C in a Heraeus Instruments Biofuge with a PP2/97 rotor. The supernatant fluid was transferred to a 1.5 ml microfuge tube and the protein concentration of each sample was determined by Bradford assay (Sigma). Equal amounts of protein (40 µg) were subjected to SDS-PAGE on 12% gels and western blotting was carried out as described above using the following primary antibodies: rabbit antibodies against SoxS 18; mouse monoclonal antibodies directed against the HA tag (Invitrogen) appended to the B42AD peptide fused to σ70 R4; and rabbit antibodies against β-tubulin (ANASPEC), which served as a loading control. The procedure for development of the blots was done as described 60. The blots were then treated with secondary antibodies and developed as described 60. The resulting chemifluorescent signals were detected with a Molecular Dynamics Storm 860 PhosphorImager in fluorescence mode with a blue (450 nm) laser.
Acknowledgements
We are grateful to our many colleagues who provided the strains that made this work possible: R. R. Burgess, S. M. Egan, C. A. Gross, V. A. Rhodius, W. Ross, and L. F. Westblade. We thank P. J. Farabaugh for help with the statistical analyses and H. Vallabhaneni for sharing her method for western analysis in yeast prior to publication. We also thank P. J. Farabaugh and K. L. Griffith for helpful discussions and N. Sanchez-Alberola for comments on the manuscript. This work was supported by Public Health Service grant GM27113 from the National Institutes of Health awarded to R.E.W.
Abbreviations used
- RNAP
RNA polymerase
- CAP
catabolite activator protein
- αCTD
C-terminal domain of RNAP α subunit
- αNTD
N-terminal domain of RNAP α subunit
- σ70 R4
region 4 of σ70
- Y2H
yeast two-hybrid system
- AIA
affinity immobilization assays
- his6
hexa-histidine tag
- FT
flow-through
- W3
third wash
- EL
eluate
- DBD
DNA binding domain
- AD
activation domain
- raf
raffinose
- gal
galactose
- X-gal
5-bromo-4-chloro-3-indolyl-β-D-galactoside
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Burgess RR. Separation and characterization of the subunits of RNA polymerase. J. Biol. Chem. 1969;244:6168–6178. [PubMed] [Google Scholar]
- 2.Zhang G, Campbell EA, Minakhin L, Richter C, Severinov K, Darst SA. Crystal structure of Thermus aquaticus core RNA polymerase at 3.3 A resolution. Cell. 1999;98:811–824. doi: 10.1016/s0092-8674(00)81515-9. [DOI] [PubMed] [Google Scholar]
- 3.Burgess RR, Travers AA, Dunn JJ, Bautz EKF. Factor stimulating transcription by RNA polymerase. Nature. 1969;221:43–46. doi: 10.1038/221043a0. [DOI] [PubMed] [Google Scholar]
- 4.Murakami KS, Masuda S, Campbell EA, Muzzin O, Darst SA. Structural basis of transcription initiation: an RNA polymerase holoenzyme-DNA complex. Science. 2002;296:1285–1290. doi: 10.1126/science.1069595. [DOI] [PubMed] [Google Scholar]
- 5.Korzheva N, Mustaev A. Transcription elongation complex: structure and function. Curr. Opin. Microbiol. 2001;4:119–125. doi: 10.1016/s1369-5274(00)00176-4. [DOI] [PubMed] [Google Scholar]
- 6.Murakami KS, Darst SA. Bacterial RNA Polymerases: the wholo story. Curr. Op. Struct. Biol. 2003;13:31–39. doi: 10.1016/s0959-440x(02)00005-2. [DOI] [PubMed] [Google Scholar]
- 7.McClure WR. Mechanism and control of transcription initiation in prokaryotes. Annu. Rev. Biochem. 1985;54:171–204. doi: 10.1146/annurev.bi.54.070185.001131. [DOI] [PubMed] [Google Scholar]
- 8.Kumar A, Malloch RA, Fujita N, Smillie DA, Ishihama A, Hayward RS. The minus 35-recognition region of Escherichia coli sigma 70 is inessential for initiation of transcription at an "extended minus 10" promoter. Journal of Molecular Biology. 1993;232:406–418. doi: 10.1006/jmbi.1993.1400. [DOI] [PubMed] [Google Scholar]
- 9.Ross W, Gosink KK, Salomon J, Igarashi K, Zou C, Ishihama A, Severinov K, Gourse RL. A third recognition element in bacterial promoters: DNA binding by the alpha subunit of RNA polymerase. Science. 1993;262:1407–1413. doi: 10.1126/science.8248780. [DOI] [PubMed] [Google Scholar]
- 10.Rhodius VA, Busby SJW. Positive activation of gene expression. Curr. Opin. Microbiol. 1998;1:152–159. doi: 10.1016/s1369-5274(98)80005-2. [DOI] [PubMed] [Google Scholar]
- 11.Rojo F. Repression of transcription initiation in bacteria. J Bacteriol. 1999;181:2987–2991. doi: 10.1128/jb.181.10.2987-2991.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ptashne M, Gann A. Transcriptional activation by recruitment. Nature. 1997;386:569–577. doi: 10.1038/386569a0. [DOI] [PubMed] [Google Scholar]
- 13.de Crombrugghe B, Chen B, Anderson WB, Gottesman ME, Perlman RL, Pastan I. Role of cyclic 3', 5'-monophosphate and the cyclic adenosine 3', 5'-monophosphate receptor protein in the initiation of lac transcription. J. Biol. Chem. 1971;246:7242–7248. [PubMed] [Google Scholar]
- 14.Majors J. Specific binding of CAP factor to lac promoter DNA. Nature. 1975;256:672–674. doi: 10.1038/256672a0. [DOI] [PubMed] [Google Scholar]
- 15.Benoff B, Yang H, Lawson CL, Parkinson G, Liu J, Blatter E, Ebright YW, Berman HM, Ebright RH. Structural basis of transcription activation: the CAP-αCTD-DNA complex. Science. 2002;297:1562–1566. doi: 10.1126/science.1076376. [DOI] [PubMed] [Google Scholar]
- 16.Haugen SP, Ross W, Gourse RL. Advances in bacterial promoter recognition and its control by factors that do not bind DNA. Nature Rev. Microbiol. 2008;6:507–529. doi: 10.1038/nrmicro1912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Minchin SD, Busby SJW. Analysis of mechanisms of activation and repression at bacterial promoters. Methods. 2008;47:6–12. doi: 10.1016/j.ymeth.2008.10.012. [DOI] [PubMed] [Google Scholar]
- 18.Griffith KL, Shah IM, Myers TE, O'Neill MC, Wolf RE., Jr Evidence for "pre-recruitment" as a new mechanism for transcription activation in Escherichia coli: the large excess of SoxS binding sites per cell relative to the number of SoxS molecules per cell. Biochem. Biophys. Res. Commun. 2002;291:979–986. doi: 10.1006/bbrc.2002.6559. Erratum. Biochem. Biophys. Res. Commun. 294:1191. [DOI] [PubMed] [Google Scholar]
- 19.Griffith KL, Wolf RE., Jr Genetic evidence for pre-recruitment as the mechanism of transcription activation by SoxS of Escherichia coli: the dominance of DNA binding mutations of SoxS. J. Mol. Biol. 2004;344:1–10. doi: 10.1016/j.jmb.2004.09.007. [DOI] [PubMed] [Google Scholar]
- 20.Martin RG, Gillette WK, Martin NI, Rosner JL. Complex formation between activator and RNA polymerase as the basis for transcription activation by MarA and SoxS in Escherichia coli. Mol. Microbiol. 2002;43:355–370. doi: 10.1046/j.1365-2958.2002.02748.x. [DOI] [PubMed] [Google Scholar]
- 21.Gallegos M-T, Schleif R, Bairoch A, Hofmann K, Ramos JL. AraC/XylS family of transcriptional regulators. Microbiol. Molec. Biol. Rev. 1997;61:393–410. doi: 10.1128/mmbr.61.4.393-410.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Griffith KL, Becker SM, Wolf RE., Jr Characterization of TetD as a transcriptional activator of a subset of genes of the Escherichia coli SoxS/MarA/Rob regulon. Mol. Microbiol. 2005;56:1103–1117. doi: 10.1111/j.1365-2958.2005.04599.x. [DOI] [PubMed] [Google Scholar]
- 23.Jair K-W, Yu X, Skarstad K, Thöny B, Fujita N, Ishihama A, Wolf RE., Jr Transcriptional activation of promoters of the superoxide and multiple antibiotic resistance regulons by Rob, a binding protein of the Escherichia coli origin of chromosomal replication. J. Bacteriol. 1996;178:2507–2513. doi: 10.1128/jb.178.9.2507-2513.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kwon HJ, Bennik MHJ, Demple B, Ellenberger T. Crystal structure of the Escherichia coli Rob transcription factor in complex with DNA. Nature Struct. Biol. 2000;7:424–430. doi: 10.1038/75213. [DOI] [PubMed] [Google Scholar]
- 25.Rhee S, Martin RG, Rosner JL, Davies DR. A novel DNA-binding motif in MarA: the first structure for an AraC family transcriptional activator. Proc. Natl. Acad. Sci. USA. 1998;95:10413–10418. doi: 10.1073/pnas.95.18.10413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Fawcett WP, Wolf RE., Jr Purification of a MalE-SoxS fusion protein and identification of the control sites of Escherichia coli superoxide-inducible genes. Mol. Microbiol. 1994;14:669–679. doi: 10.1111/j.1365-2958.1994.tb01305.x. [DOI] [PubMed] [Google Scholar]
- 27.Wood TI, Griffith KL, Fawcett WP, Jair K-W, Schneider TD, Wolf RE., Jr Interdependence of the position and orientation of SoxS binding sites in the transcriptional activation of the class I subset of Escherichia coli superoxide-inducible promoters. Mol. Microbiol. 1999;34:414–430. doi: 10.1046/j.1365-2958.1999.01598.x. [DOI] [PubMed] [Google Scholar]
- 28.Martin RG, Gillette WK, Rhee S, Rosner JL. Structural requirements for marbox function in transcriptional activation of mar/sox/rob regulon promoters in Escherichia coli: sequence, orientation and spatial relationship to the core promoter. Mol. Microbiol. 1999;34:431–441. doi: 10.1046/j.1365-2958.1999.01599.x. [DOI] [PubMed] [Google Scholar]
- 29.Griffith KL, Wolf RE., Jr Systematic mutagenesis of the DNA binding sites for SoxS in the Escherichia coli zwf and fpr promoters: identifying nucleotides required for DNA binding and transcription activation. Mol. Microbiol. 2001;40:1141–1154. doi: 10.1046/j.1365-2958.2001.02456.x. Erratum Mol. Microbiol. 42:571. [DOI] [PubMed] [Google Scholar]
- 30.Barbosa TM, Levy SB. Differential expression of over 60 chromosomal genes in Escherichia coli by constitutive expression of MarA. J. Bacteriol. 2000;182:3467–3474. doi: 10.1128/jb.182.12.3467-3474.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Martin RG, Rosner JL. Genomics of the marA/soxS/rob regulon of Escherichia coli: identification of directly activated promoters by application of molecular genetics and informatics to microarray data. Mol. Microbiol. 2002;44:1611–1624. doi: 10.1046/j.1365-2958.2002.02985.x. [DOI] [PubMed] [Google Scholar]
- 32.Pomposiello PJ, Bennik MH, Demple B. Genome-wide transcriptional profiling of the Escherichia coli responses to superoxide stress and sodium salicylate. J. Bacteriol. 2001;183:3890–3902. doi: 10.1128/JB.183.13.3890-3902.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ariza RR, Li Z, Ringstad N, Demple B. Activation of multiple antibiotic resistance and binding of stress-inducible promoters by Escherichia coli Rob protein. J. Bacteriol. 1995;177:1655–1661. doi: 10.1128/jb.177.7.1655-1661.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Amábile-Cuevas CF, Demple B. Molecular characterization of the soxRS genes of Escherichia coli: two genes control a superoxide stress regulon. Nucleic Acids Res. 1991;19:4479–4484. doi: 10.1093/nar/19.16.4479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Cohen SP, Hächler H, Levy SB. Genetic and functional analysis of the multiple antibiotic resistance (mar) locus in Escherichia coli. J. Bacteriol. 1993;175:1484–1492. doi: 10.1128/jb.175.5.1484-1492.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Asako H, Nakajima H, Kobayashi K, Kobayashi M, Aono R. Organic solvent tolerance and antibiotic resistance increased by overexpression of marA in Escherichia coli. Appl. Environ. Microbiol. 1997;63:1428–1433. doi: 10.1128/aem.63.4.1428-1433.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Nakajima H, Kobayashi K, Kobayashi M, Asako H, Aono R. Overexpression of the robA gene increases organic solvent tolerance and multiple antibiotic and heavy metal ion resistance in Escherichia coli. Appl. Environ. Microbiol. 1995;61:2302–2307. doi: 10.1128/aem.61.6.2302-2307.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.White DG, Goldman JD, Demple B, Levy SB. Role of the acrAB locus in organic solvent tolerance mediated by expression of marA, soxS, or robA in Escherichia coli. J. Bacteriol. 1997;179:6122–6126. doi: 10.1128/jb.179.19.6122-6126.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wu J, Weiss B. Two divergently transcribed genes, soxR and soxS, control a superoxide response regulon of Escherichia coli. J. Bacteriol. 1991;173:2864–2871. doi: 10.1128/jb.173.9.2864-2871.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rosenberg EY, Bertenthal D, Nilles ML, Bertrand KP, Nikaido H. Bile salts and fatty acids induce the expression of Escherichia coli AcrAB multidrug efflux pump through their interaction with Rob regulatory protein. Mol. Microbiol. 2003;48:1609–1619. doi: 10.1046/j.1365-2958.2003.03531.x. [DOI] [PubMed] [Google Scholar]
- 41.Nikaido H. Multidrug efflux pumps of gram-negative bacteria. J. Bacteriol. 1996;178:5853–5859. doi: 10.1128/jb.178.20.5853-5859.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lee E-H, Collatz E, Podglajen I, Gutman L. A rob-like gene of Enterobacter cloacae affecting porin synthesis and susceptibility to multiple antibiotics. Antimicrob. Agents Chemother. 1998;40:6299–6313. doi: 10.1128/aac.40.9.2029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Jair K-W, Fawcett WP, Fujita N, Ishihama A, Wolf RE., Jr Ambidextrous transcriptional activation by SoxS: requirement for the C-terminal domain of the RNA polymerase alpha subunit in a subset of Escherichia coli superoxide-inducible genes. Mol. Microbiol. 1996;19:307–317. doi: 10.1046/j.1365-2958.1996.368893.x. [DOI] [PubMed] [Google Scholar]
- 44.Jair K-W, Martin RG, Rosner JL, Fujita N, Ishihama A, Wolf RE., Jr Purification and regulatory properties of MarA protein, a transcriptional activator of Escherichia coli multiple antibiotic and superoxide resistance promoters. J. Bacteriol. 1995;177:7100–7104. doi: 10.1128/jb.177.24.7100-7104.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Martin RG, Gillette WK, Rosner JL. Promoter discrimination by the related transcriptional activators MarA and SoxS: differential regulation by differential binding. Mol. Microbiol. 2000;35:623–634. doi: 10.1046/j.1365-2958.2000.01732.x. [DOI] [PubMed] [Google Scholar]
- 46.Nunoshiba T, Hidalgo E, Amábile Cuevas CF, Demple B. Two-stage control of an oxidative stress regulon: the Escherichia coli SoxR protein triggers redox-inducible expression of the soxS regulatory gene. J. Bacteriol. 1992;174:6054–6060. doi: 10.1128/jb.174.19.6054-6060.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wu J, Weiss B. Two-stage induction of the soxRS (superoxide response) regulon of Escherichia coli. J. Bacteriol. 1992;174:3915–3920. doi: 10.1128/jb.174.12.3915-3920.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ding H, Demple B. In vivo kinetics of a redox-regulated transcriptional switch. Proc. Natl. Acad. Sci. USA. 1997;94:8445–8449. doi: 10.1073/pnas.94.16.8445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ding H, Hidalgo E, Demple B. The redox state of the [2Fe-2S] clusters in SoxR protein regulates its activity as a transcription factor. J. Biol. Chem. 1996;271:33173–33175. doi: 10.1074/jbc.271.52.33173. [DOI] [PubMed] [Google Scholar]
- 50.Gaudu P, Moon N, Weiss B. Regulation of the soxRS oxidative stress regulon. Reversible oxidation of the Fe-S centers of SoxR in vivo. J. Biol. Chem. 1997;272:5082–5086. doi: 10.1074/jbc.272.8.5082. [DOI] [PubMed] [Google Scholar]
- 51.Gaudu P, Weiss B. SoxR, a [2Fe-2S] transcription factor, is active only in its oxidized form. Proc. Natl. Acad. Sci. USA. 1996;93:10094–10098. doi: 10.1073/pnas.93.19.10094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Koo MS, Lee JH, Rah SY, Yeo WS, Lee JW, Lee KL, Koh YS, Kang SO, Roe JH. A reducing system of the superoxide sensor SoxR in Escherichia coli. EMBO J. 2003;22:2614–2622. doi: 10.1093/emboj/cdg252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Azam TA, Iwata A, Nishimura A, Ueda S, Ishihama A. Growth phase-dependent variation in protein composition of the Escherichia coli nucleoid. J. Bacteriol. 1999;181:6361–6370. doi: 10.1128/jb.181.20.6361-6370.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Skarstad K, Thöny B, Hwang D, Kornberg A. A novel binding protein of the origin of the Escherichia coli chromosome. J. Biol. Chem. 1993;268:5365–5370. [PubMed] [Google Scholar]
- 55.Rosner JL, Dangi B, Gronenborn AM, Martin RG. Posttranscriptional activation of the transcriptional activator Rob by dipyridyl in Escherichia coli. J. Bacteriol. 2002;184:1407–1416. doi: 10.1128/JB.184.5.1407-1416.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Azam TA, Hiraga S, Ishihama A. Two types of localization of the DNA-binding proteins with the Escherichia coli nucleoid. Genes Cells. 2000;5:613–626. doi: 10.1046/j.1365-2443.2000.00350.x. [DOI] [PubMed] [Google Scholar]
- 57.Griffith KL, Fitzpatrick MM, Keen EF, III, Wolf RE., Jr Two functions of the C-terminal domain of Escherichia coli Rob: mediating "sequestration-dispersal" as a novel off-on switch for regulating Rob's activity as a transcription activator and preventing degradation of Rob by Lon protease. J. Mol. Biol. 2009;388:415–430. doi: 10.1016/j.jmb.2009.03.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Browning DF, Busby SJW. The regulation of bacterial transcription initiation. Nature Rev. Microbiol. 2004;2:1–10. doi: 10.1038/nrmicro787. [DOI] [PubMed] [Google Scholar]
- 59.Dove SL, Darst SA, Hochschild A. Region 4 of s as a target for transcription regulation. Mol. Microbiol. 2003;48:863–874. doi: 10.1046/j.1365-2958.2003.03467.x. [DOI] [PubMed] [Google Scholar]
- 60.Shah IM, Wolf RE., Jr Novel protein-protein interaction between Escherichia coli SoxS and the DNA binding determinant of the RNA polymerase a subunit: SoxS functions as a co-sigma factor and redeploys RNA polymerase from UP-element-containing promoters to SoxS-dependent promoters during oxidative stress. J. Mol. Biol. 2004;343:513–532. doi: 10.1016/j.jmb.2004.08.057. [DOI] [PubMed] [Google Scholar]
- 61.Dangi B, Gronenborn AM, Rosner JL, Martin RG. Versatility of the carboxy-terminal domain of the a subunit of RNA polymerase in transcriptional activation: use of the DNA contact site as a protein contact site for MarA. Mol. Microbiol. 2004;54:45–59. doi: 10.1111/j.1365-2958.2004.04250.x. [DOI] [PubMed] [Google Scholar]
- 62.Gaal T, Ross W, Blatter EE, Tang H, Jia X, Krishnan VV, Assa-Munt N, Ebright RH, Gourse RL. DNA-binding determinants of the alpha subunit of RNA polymerase: novel DNA-binding domain architecture. Genes Dev. 1996;10:16–26. doi: 10.1101/gad.10.1.16. [DOI] [PubMed] [Google Scholar]
- 63.Griffith KL, Wolf RE., Jr A comprehensive alanine scanning mutagenesis of the Escherichia coli transcriptional activator SoxS: identifying amino acids important for DNA binding and transcription activation. J. Mol. Biol. 2002;322:237–257. doi: 10.1016/s0022-2836(02)00782-9. [DOI] [PubMed] [Google Scholar]
- 64.Egan SM. Growing repertoire of AraC/XylS activators. J. Bacteriol. 2002;184:5529–5532. doi: 10.1128/JB.184.20.5529-5532.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Bhende PM, Egan SM. Genetic evidence that transcription activation by RhaS involves specific amino acid contacts with sigma 70. J. Bacteriol. 2000;182:4959–4969. doi: 10.1128/jb.182.17.4959-4969.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wickstrum JR, Egan SM. Amino acid contacts between sigma 70 domain 4 and the transcription activators RhaS and RhaR. J. Bacteriol. 2004;186:6277–6285. doi: 10.1128/JB.186.18.6277-6285.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Grainger DC, Webster CL, Belyaeva TA, Hyde EI, Busby SJW. Transcription activation at the Esherichia coli melAB promoter: interactions of MelR with its DNA target site and with domain 4 of the RNA polymerase sigma subunit. Mol. Microbiol. 2004;51:1297–1310. doi: 10.1111/j.1365-2958.2003.03929.x. [DOI] [PubMed] [Google Scholar]
- 68.Fields S, Song O. A novel genetic system to detect protein-protein interaction. Nature. 1989;340:245–246. doi: 10.1038/340245a0. [DOI] [PubMed] [Google Scholar]
- 69.Dombroski AJ, Walter WA, Record MT, Jr, Siegele DA, Gross CA. Polypeptides containing highly conserved regions of transcription initiation factor σ70 exhibit specificity of binding to promoter DNA. Cell. 1992;70:501–512. doi: 10.1016/0092-8674(92)90174-b. [DOI] [PubMed] [Google Scholar]
- 70.Lonetto MA, Rhodius V, Lamberg K, Kiley P, Busby S, Gross C. Identification of a contact site for different transcription activators in region 4 of the Escherichia coli RNA polymerase σ70 subunit. J. Mol. Biol. 1998;284:1353–1365. doi: 10.1006/jmbi.1998.2268. [DOI] [PubMed] [Google Scholar]
- 71.Chen H, Tang H, Ebright RH. Functional interaction between RNA polymerase a subunit C-terminal domain and σ70 in UP-element and activator-dependent transcription. Mol. Cell. 2003;11:1621–1633. doi: 10.1016/s1097-2765(03)00201-6. [DOI] [PubMed] [Google Scholar]
- 72.Landini P, Busby SJW. The Escherichia coli Ada protein can interact with two distinct determinants in the σ70 subunit of RNA polymerase according to promoter architecture: identification of the target of Ada activation at the alkA promoter. J Bacteriol. 1999;181:1524–1529. doi: 10.1128/jb.181.5.1524-1529.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Nickels BD, Dove SL, Murakami KS, SA D, Hochschild A. Protein-protein and protein-DNA interactions of σ70 region 4 involved in transcription activation by λcI. J. Mol. Biol. 2002;324:17–34. doi: 10.1016/s0022-2836(02)01043-4. [DOI] [PubMed] [Google Scholar]
- 74.Zhou Y, Merkel TJ, Ebright RH. Characterization of the activating region of Escherichia coli catabolite gene activator protein (CAP): II. Role at Class I and Class II CAP-dependent promoters. J. Mol. Biol. 1994;243:603–610. doi: 10.1016/0022-2836(94)90035-3. [DOI] [PubMed] [Google Scholar]
- 75.Kuldell N, Hochschild A. Amino acid substitutions in the −35 recognition motif of sigma 70 that result in defects in phage lambda repressor-stimulated transcription. J. Bacteriol. 1994;176:2991–2998. doi: 10.1128/jb.176.10.2991-2998.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Li M, Moyle H, Susskind MM. Target of the transcriptional activation function of the phage l cI protein. Science. 1994;263:75–77. doi: 10.1126/science.8272867. [DOI] [PubMed] [Google Scholar]
- 77.Ebright RH, Kolb A, Buc H, Kunkel TA, Krakow JS, Beckwith J. Role of glutamic acid-181 in DNA-sequence recognition by the catabolite gene activator protein (CAP) of Escherichia coli: altered DNA-sequence-recognition properties of [Val181]CAP and [Leu181]CAP. Proc. Natl. Acad. Sci. USA. 1987;84:6083–6087. doi: 10.1073/pnas.84.17.6083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Westblade LF, Ilag LL, Powell AK, Kolb A, Robinson CV, Busby SJ. Studies of the Escherichia coli Rsd-sigma70 complex. J. Mol. Biol. 2004;335:685–692. doi: 10.1016/j.jmb.2003.11.004. [DOI] [PubMed] [Google Scholar]
- 79.Grainger DC, Overton TW, Reppas N, Wade JT, Tamai E, Hobman JL, Constantinidou C, Struhl K, Church G, Busby SJW. Genomic studies with Escherichia coli MelR protein: applications of chromatin immunoprecipitation and microarrays. J. Bacteriol. 2004;186:6938–6943. doi: 10.1128/JB.186.20.6938-6943.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Guzman L-M, Belin D, Carson MJ, Beckwith JR. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 1995;177:4121–4130. doi: 10.1128/jb.177.14.4121-4130.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Griffith KL, Wolf RE., Jr Measuring β-galactosidase activity in bacteria: cell growth, permeabilization, and enzyme assays in 96-well arrays. Biochem. Biophys. Res. Commun. 2002;290:397–402. doi: 10.1006/bbrc.2001.6152. [DOI] [PubMed] [Google Scholar]
- 82.Rhodius VA, Busby SJ. Interactions between activating region 3 of the Escherichia coli cyclic AMP receptor protein and region 4 of the RNA polymerase sigma70 subunit: application of suppression genetics. J. Mol. Biol. 2000;299:311–324. doi: 10.1006/jmbi.2000.3737. [DOI] [PubMed] [Google Scholar]
- 83.Arthur T, Burgess RR. Localization of a sigma70 binding site on the N terminus of the Escherichia coli RNA polymerase beta' subunit. J Biol Chem. 1998;273:31381–31387. doi: 10.1074/jbc.273.47.31381. [DOI] [PubMed] [Google Scholar]
- 84.Vallabhaneni H, Farabaugh PJ. Accuracy modulating mutations of the ribosomal protein S4-S5 interface do not necessarily destabilize the rps4-rps5 protein-protein interaction. RNA. 2009;15:1100–1109. doi: 10.1261/rna.1530509. [DOI] [PMC free article] [PubMed] [Google Scholar]