

Abstract
During the 2005 hurricane season, the storm surge and wave field associated with Hurricanes Katrina and Rita eroded 527 km2 of wetlands within the Louisiana coastal plain. Low salinity wetlands were preferentially eroded, while higher salinity wetlands remained robust and largely unchanged. Here we highlight geotechnical differences between the soil profiles of high and low salinity regimes, which are controlled by vegetation and result in differential erosion. In low salinity wetlands, a weak zone (shear strength 500–1450 Pa) was observed ∼30 cm below the marsh surface, coinciding with the base of rooting. High salinity wetlands had no such zone (shear strengths > 4500 Pa) and contained deeper rooting. Storm waves during Hurricane Katrina produced shear stresses between 425–3600 Pa, sufficient to cause widespread erosion of the low salinity wetlands. Vegetation in low salinity marshes is subject to shallower rooting and is susceptible to erosion during large magnitude storms; these conditions may be exacerbated by low inorganic sediment content and high nutrient inputs. The dramatic difference in resiliency of fresh versus more saline marshes suggests that the introduction of freshwater to marshes as part of restoration efforts may therefore weaken existing wetlands rendering them vulnerable to hurricanes.
Keywords: wetland loss, Mississippi Delta, shear stress, soil shear strength, vegetation roughness
Coastal wetlands are valued for their ability to protect landward areas from the impacts of storms: dampening wave energy, distancing the mainland from the open water, and potentially reducing storm surge height (1–5). Understanding and predicting wetland loss, and the consequences with respect to coastal populations, is of global interest. Present projections suggest that before the end of the century, as much as one third of the world’s coastal wetlands could be lost to sea-level rise (6). Furthermore, a recent study (7) underscores the vulnerability of the world’s largest deltas and the need to maintain the sediment supply to the vast wetlands that comprise these regions. In an effort to stabilize deltaic wetlands, riverine diversions have been introduced, designed to restore (preanthropogenic) conditions by reducing salinity and reintroducing sediment within these vulnerable deltas (8).
Much of the research pertaining to wetland loss considers the threat of submergence due to an inability to keep pace with sea level (2–4, 9). However, in their role as a coastal buffer, marshes may experience a range of damage including vegetation mortality as a result of wrack deposition during high waters, salt intrusion into freshwater wetlands, enhanced wave erosion at the marsh edge (10), or the bulk removal of the vegetation mat. The extent of this damage will affect recovery time and the health of the poststorm marsh and its effectiveness as a buffer, and it may lead to permanent wetland loss (5, 11). Arguably, cannibalization of sediment from one region of the marsh may provide the input necessary to bolster neighboring zones (3), but the result is still one of net loss.
In 2005, the storm surge and wave field associated with Hurricanes Katrina and Rita eroded 527 km2 of wetlands within the Louisiana coastal plain (11). Low salinity wetlands were preferentially eroded, while higher salinity wetlands were more resilient (Fig. 1). Salinity regimes were defined according to vegetation mapping (12, 13): “low salinity” describes a grouping of freshwater and intermediate marsh comparable to limnetic and olighohaline conditions; “high salinity” incorporates brackish and saline marsh and is equivalent to meso- and polyhaline conditions. (For further information on vegetation-salinity regimes see Table S1.) The high degree of spatial coincidence between hurricane-induced erosion and low salinity wetlands across coastal Louisiana (including parts of the Chenier plain and the Birdfoot, Wax Lake, and Atchafalaya deltas) suggests a regionally consistent process-response model. Furthermore, the fact that older landforms (the Chenier Plain and Breton Sound) experienced similar land loss trends to relatively modern landforms (Birdfoot and Wax Lake deltas) suggests that it is not age that is controlling land loss but salinity regime. In this paper we explore the ability of different wetland types to resist erosion by quantifying the shear strength of the soils and vegetation, and we compare these strengths to the forces imposed across the wetlands by hurricane-generated waves. In upper Breton Sound, a region that is heavily impacted by a freshwater diversion, we find that low salinity marshes have shallower rooting and significantly weaker soils. These conditions could be exacerbated by low inorganic sediment content (14) and high nutrient inputs (15, 16).
Fig. 1.

(A and B) Land loss in the Louisiana coastal plain. Low salinity (fresh and intermediate combined) marsh experienced more than twice as much land loss by percent than high salinity (brackish and saline combined) marsh. The failure of low salinity wetlands was focused in the interior regions of Breton Sound, the western chenier plain, and the more exposed regions of the Birdfoot and Wax Lake deltas. (C) Zoom in on Breton Sound. Vegetation type and conditions defining each zone are given in the bottom table (expanded in Table S1).
In the mid-twentieth century, levee construction along the Mississippi River effectively cut off wetlands from fluvial sedimentation and freshwater overland flows. To restore the preexisting salinity gradients and the extent of freshwater wetlands, the Caernarvon Diversion began operating in 1991, diverting Mississippi River water into the interior wetlands of Breton Sound. Additional diversions have been planned for other regions of the Mississippi River delta, which similarly lack a natural riverine freshwater input. However, the results of this study raise questions about the resilience of freshwater environs relative to saline regions.
Within our study site, upper Breton Sound, both low and high salinity wetlands experienced very similar hydrodynamic conditions during Hurricane Katrina (17), yet low salinity wetlands failed preferentially, providing a natural case to study causes of the observed land loss patterns. Furthermore, the freshwater diversion has been operating for more than 18 years, longer than at any other site in the delta, allowing the wetlands to adapt to the imposed conditions. The basin is defined by the Mississippi River levees along its western margin and the Mississippi River Gulf Outlet (MGRO) protection levee along the eastern margin, encompassing 1,100 km2 of wetlands that grade from freshwater to saline. Bayou Terre aux Beoufs, a distributary of the former St. Bernard delta, partitions the flow from the diversion resulting in extensive low salinity wetlands west of the bayou and higher salinity brackish and saline marshes to the east (18). The diversion has successfully increased the extent of low salinity wetlands west of the bayou, displacing the boundary between low and high salinity wetlands seaward by as much as 29 km since 1991.
As part of this study, ground and aerial surveys of the low salinity wetlands in Breton Sound conducted following Hurricane Katrina revealed extensive deposits of “marshballs,” uprooted masses of marsh grass consisting of a rooting mat and bound sediment. Similar observations have been made elsewhere (5). The marshballs typically had a rooting depth of 20–30 cm and were neutrally buoyant. Devegetated peat surfaces in the nearby marsh surface marked the source regions of the rafted marshballs. Field observations illustrated that the erosion occurred by scouring of the marsh surface, including the root mat, rather than by sediment resuspension as has been studied elsewhere (10, 19). A similar process was suggested by van de Plassche et al. (2004) to explain erosive contacts within the stratigraphy of a Connecticut wetland (20). In contrast to the low salinity regions, saline portions of the marsh were left largely intact. We hypothesize that wave shear stresses generated during the hurricanes exceeded the shear strength of the low salinity wetland soils, resulting in failure, whereas greater soil shear strength in the saline wetlands largely precluded erosion. Soil shear strength and the resistance of the soil to erosion are determined by the properties of the vegetation, the surrounding soil matrix, and their interaction. We propose that resistance to erosion is primarily a function of rooting characteristics, which depend on the dominant species of vegetation, as controlled by salinity (2, 13) (Table S1).
Results
Field-vane measurements illustrate that the brackish-saline soils are geotechnically distinct from the fresh-intermediate soils and consistently stronger within the upper soil column (P < 0.0001, α = 0.02) (Fig. 2A). The greatest scatter in shear strength is observed near the surface, which is attributed to variations in both rooting density and the strength of individual roots. Strengths in the two soil types converge with depth. Weak zones in the freshwater substrate have been identified at depths between 10–50 cm based on shear strength, characterized not only by low minimum strengths (1500 Pa) but also reduced maximum strengths (6800 Pa). In this case, the low minimum strengths reflect a low shear strength soil matrix, and the reduced maximum strengths generally reflect a lower rooting abundance. An analogue low strength layer was not observed in the saline marshes; they exhibit a more consistent strength profile with depth, although the decrease in live roots with depth does produce a decreasing trend in strength.
Fig. 2.

(A) A comparison of in situ shear strength measurements in low and high salinity wetlands (low salinity n = 53, from six sites; high salinity n = 47, from six sites) (see Fig. S1). The arrows indicate that the soil shear strength exceeded the vane maximum, as our objective was to examine minimum shear strengths. A less sensitive torque head was used to determine the maximum values occurring in the saline marsh at only one site. A two-tailed Welches t-test shows that the low and high salinity populations are significantly different (P < 0.0001, α = 0.02). (B) Shear strengths of the low salinity wetland including both lab (n = 30) and field data (n = 53). Note the minimum in average strengths around 30 cm depth. The arrows at the base of the diagram indicate wave shear stress from scenarios A, B, C, and D in Fig. 3.
The field-vane shear strength measurement often overestimates soil strength (21, 22) (Methods and SI Text A). Thus, in order to further quantify the weak layer observed in the low salinity wetland, laboratory vane measurements were made on undisturbed half cores collected in the field. The overall trend in shear strength is similar to the field tests, but the absolute values measured were consistently lower. In the weak layer, a minimum value of 500 Pa was observed and values ranged between 500–3000 Pa (Fig. 2B). The field-vane and lab-vane tests bracket and constrain the actual undrained strength of the soil.
Model simulations of Hurricane Katrina at the study site produced a peak surge of 4.4 m, a significant wave height of 2.1 m, with wave periods between 5–9 s. Using the two representative roughness lengths (Z0 = 0.17 & 0.30 m) (23, 24) wave shear stresses during the storm were calculated at locations surrounding our coring stations (25). Four model scenarios were used to investigate wave shear stress (Fig. 3 and Fig. S1). The maximum values of wave shear stress ranged between 425–3600 Pa, sufficient to cause failure within the low salinity wetland, which exhibited minimum soil strengths between 500–1450 Pa, but not in the high salinity regions where the soil strength exceeded 4500 Pa in all measurements. The potential failure of low strength, low salinity wetlands under wave shear stresses will be illustrated below with a scaling argument. In addition to individual wave events, which produced sufficient shear stress to exceed the soil strength of the low salinity wetland, there would have been long-term stress on the underlying substrate due to the repetitive passage of waves (leading to fatigue), as well as the impact of breaking waves.
Fig. 3.

Wave shear stresses generated during Hurricane Katrina at seven sites in Breton Sound (see also Fig. S1). Four scenarios are modeled with different combinations of two wave heights (H) and two roughness lengths (Z0). Hs is significant wave height and Hmax is maximum wave height (Hmax = 1.868Hs).
In Breton Sound, soil weakness in the low salinity wetlands correlates closely with an inorganic layer. The layer is identified by discontinuities in bulk density, gravimetric water content, and organic content (Fig. 4 and Fig. S2). The layer is interpreted as a possible crevasse splay or as storm related deposits (20) and is bracketed above and below by marsh peat. Most importantly, it lacks the abundant rooting structures observed in the peat layer above. Low salinity wetlands proximal to rivers and distributaries are exposed to periodic flooding and are expected to exhibit similar stratigraphy consisting of a sequence of peat interrupted by inorganic overbank flood deposits. Similar inorganic layers have been reported in both the Atchafalaya delta and upper-basin freshwater wetlands (26, 27). Hurricanes are also capable of depositing inorganic material across a marsh platform (3).
Fig. 4.

Geotechnical soil properties measured in the low salinity wetland (see also Figs. S1 and S2). A discontinuity exists at roughly 30 centimeters depth attributed to an inorganic layer that interrupts the rooting profile and displays markedly different soil properties.
The minimum freshwater soil strengths were concentrated near the top of the inorganic layer, approximately 30 cm below the marsh surface. The depth of the root mat within the post-Katrina marshballs (20–30 cm) supports the theory that the wetlands failed along this layer. The interruption in rooting that coincided with the inorganic sediment layer was likely exploited during the storm. Intact roots contribute to overall strength of a soil. Vegetation increases the strength of soils by providing mechanical reinforcement, which in turn is controlled by the rooting depth, diameter, and density (28, 29). The tensile force required to break individual roots and rhizomes has been shown to scale with diameter, and the overall shear strength of a soil increases proportionally to the cross sectional area of the roots crossing the shearing plane (28, 29). As a result, highly decayed rooting and unvegetated depths in the soil column display lower values of strength (30).
Spartina alterniflora is the most abundant species in high salinity wetland, characterized by extensive horizontal rhizomes and a dense stem-root network that reinforces the surrounding soil (31). The rooting depth of Sp. alterniflora is less sensitive to anoxic soil conditions than other species, allowing it to colonize lower elevations in the tidal regime (32). In contrast, many post-Katrina marshballs consisted of Spartina patens, one of the dominant species in intermediate wetlands (Table S1), which also has extensive rooting but of a smaller diameter. The plant is less tolerant to anoxic soil conditions, which likely limits the root network to shallower depths (32). Direct comparisons of the variation of belowground biomass with depth indicate that live rooting is shallower and less abundant in Sp. patens marshes than in Sp. alterniflora marshes (e.g., 33).
In unvegetated soils, a typical soil strength profile increases with depth and the greater degree of consolidation. The strength profile is fundamentally different in vegetated soils (28). In this case, strength decreases with depth as the result of a decrease in root abundance. The saline marsh strength measurements exhibit this trend, which illustrates the dominant role of vegetation in the uppermost meter of the soil column. The freshwater data reflect a combination of vegetated and unvegetated profile characteristics. In the near surface, the vegetation effect is reflected by relatively high values of strength. However, the profile rapidly decreases toward a minimum as rooting diminishes near the inorganic layer and transitions into the typical unvegetated profile of increasing strength with depth (Fig. 5). Based on our data, the minimum strength in a wetland soil profile occurs where the control of soil strength by roots is superseded by control of the substrate (i.e., unvegetated soil). This situation exists at a shallow depth in the low salinity wetland but was not clearly observed in the saline wetland, where the rooting continued to dominate the strength profile to greater depths. The convergence of strength profiles in the saline and low salinity soils at around 1 m in depth likely reflects the beginning of the transition from root to substrate control in the saline marsh.
Fig. 5.

A conceptual model of soil strength in freshwater and saline marshes based on field data. In the freshwater marsh, shallow roots dominate the strength profile in the uppermost portion of the soil. Beneath the roots, the strength is controlled by the unvegetated soil properties. The minimum values in strength occur at this transition, which defines the depth of the theoretical failure plane. In the saline marsh, roots dominate the strength profile to a greater depth. In addition, the roots in saline marsh appear stronger, shifting the vegetated profile to the right. As a result, the transition and theoretical failure plane occurs deeper in soil column, which results in a higher value of minimum strength and more resistance to erosion.
Discussion
To illustrate the effect of rooting depth (d) on soil failure, we compare the force of waves imparted on a block of soil and compare it with the forces resisting erosion. The soil fails if the force of the waves on the block is equal to or greater than the force resisting scour: Force = Stress × Area
Failure occurs if:
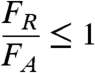 |
[1] |
Where FR is the resisting force, in this case the strength of the soil, and FA is the applied force, in this case the wave forcing.
Wave shear stress (τw) acts tangential to the surface of the marsh and is multiplied by the area over which it acts to calculate total wave force. The stress is assumed constant over the surface of the block if the length (l) and width (w) of the block is much less than the wavelength of the wave. In each wave cycle, there are two peak wave shear stress events: one with the passage of the trough and the other beneath the crest. These peak stresses occur in opposite directions. The undrained soil strength (τs) is assumed identical in each plane and acts to resist erosion. This is a simplification of nature, and of the processes occurring, but remains useful to illustrate the concept of rooting depth. To erode a rectangular block from an intact marsh surface, failure occurs on five planes (four vertical planes and one basal plane). In this case, the total force resisting erosion is the soil strength multiplied by the surface area of the failure planes. When the ratio of the soil forces to the wave forces are compared Eq. 1 becomes:
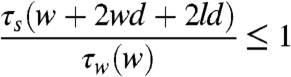 |
[2] |
Where τs is the undrained shear strength and τw is the wave shear stress. This expands to
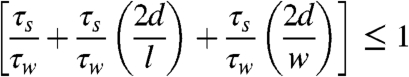 |
[3] |
For a given area over which the wave forces act (i.e., a given block dimensions l × w), the soil force resisting erosion is therefore proportional to the depth of the roots or the depth of any weak layer in the soil column. Using values of 30 cm for depth (based on our field observations of marshball dimensions) and unity for length scale (i.e., O ∼ 1 m), wave shear stress would need to exceed the soil strength by a factor of 1.6, to cause failure. Based on our measurements of minimum strength in low salinity wetland (500–1450 Pa), “failure” (scouring of the root mat) would require a wave shear stress of 800–2320 Pa, a range that closely agrees with our earlier estimates of shear during the storm (425–3600 Pa). In contrast, the saline marsh displayed a rooting depth closer to 1 m and minimum strength values of 4,500 Pa. Assuming the same length scale (O ∼ 1 m), wave shear stress would need to exceed soil strength by a factor of 2, requiring wave shear stresses exceeding 9,000 Pa, which exceeds the estimated storm-induced shear. The depth of the minimum soil strength is clearly a critical parameter in terms of soil failure. The resistance of a wetland to scouring is proportional to the depth of any weak layer. A shallow weakness will enhance failure, as was the case in low salinity wetlands of the Louisiana coastal plain.
The detailed physics of marsh failure during storms is more complicated than this simplified model, however, and the feedbacks involved are not yet fully understood or quantified. In addition to rooting properties, soil strength and the mode of wetland failure will also be affected by soil properties such as bulk density and water content. The bulk density of the soil matrix surrounding roots depends on the ratio of mineral to organic accumulation. Rates of mineral sedimentation in inland freshwater marshes are as much as five times lower than saline marshes (34), reducing overburden pressures, soil consolidation, and friction between the roots and the surrounding soil, which could change the mode of failure from root breakage to root slippage (28). When inorganic sedimentation is extremely low, a buoyant root mat develops (due to the low bulk density of organics) that diverges from the substrate below and creates a zone of weakness in the soil column. Further, exposure of marine clays to freshwater can also reduce soil matrix strengths by 40–60%; this effect cannot account for the maximum difference in shear strength observed between high and low salinity marshes within our data but must instead be attributed to rooting (35).
High concentrations of nutrients carried by the Mississippi and Atchafalaya rivers may also contribute to lower soil strength in areas receiving riverine input. Swarenski et al. (2008) concluded that river water input to freshwater marshes creates a more reducing soil environment, greater degradation of the root mat, and a weaker overall soil that is more susceptible to hurricanes; these observations are mirrored by our freshwater soil strength data in Breton Sound. Greater concentrations of nitrogen and phosphorus may also reduce the need of plant roots to forage deep into the substrate and thereby reduce the extent and depth of their rooting (15), a key factor in soil failure. Furthermore, Darby and Turner (2008) conclude that nitrogen and phosphorous loading increases aboveground biomass. If true, this development would enhance wave shear stress due to greater vegetation related roughness lengths, while reducing the belowground soil reinforcement by roots. A number of reasons can be presented to account for differences in rooting depth (or strength) between different species, including stratigraphic or biogeochemical controls, in any site where regions of vegetation have differing rooting depths; however, the conceptual model will hold, and a disparity in soil strength would be expected.
Our research identifies a key difference between the geotechnical strength profile of low and high salinity marshes. Saline marshes in Breton Sound were more able to resist storm erosion than low salinity wetlands due to higher soil strengths. The comparative difference in soil strength is the result of more robust, deeper rooting by saline marsh plants. Low salinity wetlands are vulnerable to bulk erosion of the marsh platform during storms due to weaker soil strengths and reduced rooting depths. The influence of high nutrient levels or unrooted layers may contribute to or compound this. With this in mind, proposed freshwater diversions should be reassessed not only for their ability to restore low salinity and introduce sediment but their efficacy in creating robust hurricane-resistant marsh, because the potential exists for the development of geotechnically weak soils and thus higher susceptibility to erosion during large magnitude storm events.
Methods
To quantify the comparative strengths of wetland soils, geotechnical measurements were taken along a transect through low and high salinity marshes. Half cores were retrieved (avoiding sediment compaction, at sites a minimum of 15 m from the marsh platform edge) and sampled with depth to obtain saturated bulk density, gravimetric water content, and organic content (Figs. S1 and S2). The undrained soil shear strength was measured in situ using a Seiken field-vane (low salinity n = 53; high salinity n = 47), and further measurements of weaker soils were carried out in the laboratory with a Wykeham–Farrance lab-vane (low salinity n = 30). Undrained strength is a measure of strength specifically employed when a rapid loading condition or force is applied to a fine-grained saturated soil, such that there is insufficient time for the soil to drain. This was the case when the Breton Sound region was inundated by the storm surge (the entire region was “open water”) and experienced wave forces. High and low salinity marshes were compared for significant difference using a two-tailed Welches t-test.
To establish the hydrodynamic conditions within the basin during storm conditions, numerical model results were obtained using the full plane steady state spectral wave model STWAVE run in a quasi-coupled mode with the unstructured coastal ocean circulation model ADCIRC (interfaced every 30 min), with spatially varying wind fields and spatially varying friction (15). Based upon the hydrodynamic conditions (significant wave height, Hs; peak period, Tp; and water depth, h) and an appropriate wave friction factor (fw), calculations were made to determine the maximum wave shear stresses acting on the vegetated marsh platform (25, 36). The wave friction factor parameterizes the drag related to vegetation and is a function of the roughness length (Z0), which is influenced by stem density, canopy height, and leaf geometry (37–41). Although, when emergent, vegetation dramatically reduces flow velocities (4, 40, 42), when fully submerged it produces a two-phase flow; speeds are reduced within the canopy, while a faster skimming flow occurs above the canopy (38, 41). When water depth becomes significantly deeper than the canopy height, as occurred during Hurricane Katrina, vegetation can be considered as a bed roughness element, retarding flow and increasing shear in the near-bed boundary layer (40, 41). Two characteristic roughness lengths (17 and 30 cm) were used to calculate maximum wave shear stresses, representing the upper and lower limits of vegetation conditions (37–40). Calculations were made at seven sites on either side of the Terre aux Boeuf distributary (see SI Text B and Table S2).
Supplementary Material
Acknowledgments.
We gratefully acknowledge field assistance received from M. Brown, P. McCarty, and J. Motti. This work was primarily funded by National Oceanic and Atmospheric Administration Pontchartrain Restoration Program Grant NA06 NOS4630026 awarded to the University of New Orleans and Boston University. U.S. Geological Survey Northern Gulf of Mexico (NGOM) Ecosystem Change and Hazard Susceptibility Project provided funding for the land loss mapping.
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.0914582107/-/DCSupplemental.
References
- 1.Moeller I, Spencer T, French JR. Wind wave attenuation over saltmarsh surfaces: Preliminary results from Norfolk, England. J Coastal Res. 1996;12:1009–1016. [Google Scholar]
- 2.Mitsch WJ, Gosselink JG. Wetlands. 3rd ed. New York: Wiley; [Google Scholar]
- 3.McKee KL, Cherry JA. Hurricane Katrina sediment slowed elevation loss in subsiding brackish marshes of the Mississippi River delta. Wetlands. 2009;29:2–15. [Google Scholar]
- 4.Reed DJ, Spencer T, Murray AL, French JR, Leonard L. Marsh surface sediment deposition and the role of tidal creeks: Implications for created and managed coastal marshes. J Coast Conservat. 1999;5:81–90. [Google Scholar]
- 5.Day JW, Jr, et al. Restoration of the Mississippi delta: Lessons learned from Hurricanes Katrina and Rita. Science. 2007;315:1679–1684.29. doi: 10.1126/science.1137030. [DOI] [PubMed] [Google Scholar]
- 6.Parry ML, Canziani OF, Palutikof JP, van der Linden PJ, Hanson CE, editors. IPCC. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK, and New York: Cambridge Univ Press; 2007. p. 976. [Google Scholar]
- 7.Syvitski JPM, et al. Sinking deltas due to human activities. Nat Geosci. 2009;2:681–686. [Google Scholar]
- 8.Blum MD, Roberts HH. Drowning of the Mississippi Delta due to insufficient sediment supply and global sea-level rise. Nat Geosci. 2009;2:488–491. [Google Scholar]
- 9.Cahoon DR, et al. In: Wetlands and Natural Resource Management. Ecological Studies. Verhoeven JTA, Beltman B, Bobboink R, Whigham D, editors. Vol. 190. Berlin Heidelberg: Springer-Verlag; 2006. pp. 271–292. [Google Scholar]
- 10.Feagin RA, et al. Does vegetation prevent wave erosion of salt marsh edges? Proc Natl Acad Sci USA. 2009;106:10109–10113. doi: 10.1073/pnas.0901297106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Barras JA, Bernier JC, Morton RA. Land area changes in coastal Louisiana—A multi decadal perspective (from 1956 to 2006) US Geological Survey Scientific Investigations Map 3019. 2008;14 scale 1∶250,000. [Google Scholar]
- 12.Visser JM, Sasser CE, Chabreck RH, Linscombe RG. Marsh vegetation types of the Mississippi River Deltaic Plain. Estuaries. 1998;21:818–828. [Google Scholar]
- 13.Visser JM, Sasser CE, Chabreck RH, Linscombe RG. Marsh vegetation types of the Chenier Plain USA. Estuaries. 2000;23:318–327. [Google Scholar]
- 14.Ravens TM, Thomas RC, Roberts KA, Santschi PH. Causes of salt marsh erosion in Galveston Bay, Texas. J Coast Res. 2009;25:265–272. [Google Scholar]
- 15.Darby FA, Turner RE. Below and aboveground biomass of Spartina alterniflora: Response to nutrient addition in a Louisiana salt marsh. Estuar Coast. 2008;31:326–334. [Google Scholar]
- 16.Swarzenski CM, Doyle TW, Fry B, Hargis TG. Biogeochemical response of organic-rich freshwater marshes in the Louisiana delta plain to chronic river water influx. Biogeochemistry. 2008;90:49–63. [Google Scholar]
- 17.Bunya S, et al. A high resolution coupled riverine flow, tide, wind, wind wave and storm surge model for southern Louisiana and Mississippi: Part I—Model development and validation. Monthly Weather Review. [(accepted 2009)].
- 18.Piazza BP, La Peyre MK. Restoration of the annual flood pulse in Breton Sound, Louisiana, USA: Habitat change and nekton community response. Aquat Biol. 2007;1:109–119. [Google Scholar]
- 19.van de Plassche O, et al. Salt-marsh erosion associated with hurricane landfall in southern New England in the fifteenth and seventeenth centuries. Geology. 2006;34:829–832. [Google Scholar]
- 20.Temmerman S, et al. Vegetation causes channel erosion in a tidal landscape. Geology. 2007;35:631–634. [Google Scholar]
- 21.Bjerrum L. Embankments on soft ground; Proc ASCE Specialty Conf Performance of Earth and Earth Supported Structures; Lafayette, IN: Purdue Univ; 1972. pp. l–54. [Google Scholar]
- 22.Day RW. Soil Testing Manual. New York: McGraw-Hill; 2001. [Google Scholar]
- 23.Klopstra D, Barneveld HJ, van Noortwijk JM, van Velzen EH. Analytical model for hydraulic roughness of submerged vegetation; Proceedings of the 27th Congress of the International Association for Hydraulic Research, San Francisco.; New York: Am Society of Civil Engineers; 1997. pp. 775–780. [Google Scholar]
- 24.Leonard LA, Croft AL. The effect of standing biomass on flow velocity and turbulence in Spartina alterniflora canopies. Estuar Coast Shelf S. 2006;69:325–336. [Google Scholar]
- 25.Soulsby RL. Dynamics of Marine Sands, a Manual for Practical Applications. Telford, UK: Thomas; 1997. p. 250. [Google Scholar]
- 26.Swarzenski CM, Swenson EM, Sasser CE, Gosselink JG. Marsh mat flotation in the Louisiana delta plain. J Ecol. 1991;79:999–1011. [Google Scholar]
- 27.Sasser CE, Gosselink JG, Swenson EM, Swarzenski CM, Leibowitz NC. Vegetation, substrate and hydrology in floating marshes in the Mississippi river delta plain wetlands, USA. Vegetatio. 1996;122:129–142. [Google Scholar]
- 28.Mickovski SB, et al. Mechanical reinforcement of soil by willow roots: Impacts of root properties and root failure mechanism. Soil Sci Soc Am J. 2009;73:1276–1285. [Google Scholar]
- 29.van Eerdt MM. The influence of vegetation on erosion and accretion in salt marshes of the Oosterschelde, The Netherlands. Vegetatio. 1985;62:367–373. [Google Scholar]
- 30.Huat BBK, Asadi A, Kazemian S. Experimental investigation on geomechanical properties of tropical organic soils and peat. Am J Eng App Sci. 2009;2:184–1888. [Google Scholar]
- 31.Chung C. Forty years of ecological engineering with Spartina plantations in China. Ecol Eng. 2006;27:49–57. [Google Scholar]
- 32.Bertness MD. Zonation of Spartina patens and Spartina alterniflora in a New England salt marsh. Ecology. 1991;72:138–148. [Google Scholar]
- 33.Connor RF, Chmura GL. Dynamics of above- and belowground organic matter in a high latitude macrotidal saltmarsh. Mar Ecol Prog Ser. 2000;204:101–110. [Google Scholar]
- 34.Bjerrum L, Rosenqvist IT. Some experiments with artificially sedimented clays. Geotechnique. 1956;6:124–136. [Google Scholar]
- 35.Nyman JA, Walters RJ, Delaune RD, Patrick WH., Jr Marsh vertical accretion via vegetative growth. Estuar Coast Shelf S. 2006;69:370–380. [Google Scholar]
- 36.Mathisen PP, Madsen OS. Waves and currents over a fixed rippled bed. 1. Bottom roughness experienced by waves in the presence and absence of currents. J Geophys Res. 1996;101:16533–16542. [Google Scholar]
- 37.Augustin LN, Irish JL, Lynett P. Laboratory and numerical studies of wave damping by emergent and near-emergent wetland vegetation. Coast Eng. 2009;56:332–340. [Google Scholar]
- 38.Neumeier U. Velocity and turbulence variations at the edge of salt marshes. Cont Shelf Res. 2007;27:1046–1059. [Google Scholar]
- 39.Neumeier U, Amos CL. The influence of vegetation on turbulence and flow velocities in European salt marshes. Sedimentology. 2006;53:259–277. [Google Scholar]
- 40.Neumeier U, Ciavola P. Flow resistance and associated sedimentary processes in a Spartina maritime salt marsh. J Coast Res. 2004;20:435–447. [Google Scholar]
- 41.Ghisalberti M, Nepf H. The structure of the shear layer over rigid and flexible canopies. Environ Fluid Mech. 2006;6:277–301. DOI: 10.1007/s10652-006-0002-4. [Google Scholar]
- 42.Brown SL. Black KS, Paterson DM, Cramp A, editors. Sedimentation on Humber saltmarsh. Sedimentary Processes in the intertidal zone, Geological Special Publication 139. 1998:69–84. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.