

Abstract
Recovery of consciousness following severe brain injuries may occur over long time intervals. Importantly, evolving cognitive recovery can be strongly dissociated from motor recovery in some individuals, resulting in underestimation of cognitive capacities. Common mechanisms of cerebral dysfunction that arise at the neuronal population level may explain slow functional recoveries from severe brain injuries. This review proposes a “mesocircuit” model that predicts specific roles for different structural and dynamic changes that may occur gradually during recovery. Recent functional neuroimaging studies that operationally identify varying levels of awareness, memory and other higher brain functions in patients with no behavioral evidence of these cognitive capacities are discussed. Measuring evolving changes in underlying brain function and dynamics post-injury and post-treatment frames future investigative work.
Recovery of conscious awareness and cognitive function following severe brain injuries can occur over surprisingly long time intervals of months, years and rarely decades [1-5]. Moreover, recovery of consciousness may significantly lag or be entirely dissociated from expressed motor behavior [6]. It is increasing recognized that very limited evidence of behavioral responsiveness at the bedside (or rarely, even a lack of any evidence) does not accurately predict underlying brain function. As a result, significant ambiguity can be present when encountering behavioral features consistent with clinical diagnoses ranging from vegetative state (no behavioral evidence of self or environmental awareness), minimally conscious state (at least some behavioral evidence of awareness), and up to and including patients in locked-in state (full consciousness with limited to no motor control). Importantly, bedside behavioral assessment cannot alone provide insight into likelihood of further recovery, avenues for specific intervention, or level of consciousness and cognitive capacity. The underlying mechanisms accounting for this wide variance in recovery patterns are unknown and provide a compelling scientific challenge for further understanding.
This review considers aspects of current research aimed at understanding recovery of consciousness after brain injury. To best organize this advancing knowledge, a model at the neuronal population level is proposed that accounts for observed neuroimaging findings and response to treatments in the context of pathophysiological mechanisms associated with severe brain injury. This ‘mesocircuit’ model provides a parsimonious explanation of observations of recovery of consciousness after severe brain injuries and predicts several seemingly unrelated findings. From this vantage point recent research advances are reviewed, including: 1) interventional studies using pharmacological and electrical stimulation methods to improve function in patients with longstanding disorder of consciousness. 2) new functional neuroimaging techniques that reliably, and operationally, identify levels of awareness, memory and other higher brain functions in patients who show no behavioral evidence of these capacities, and 3) structural neuroimaging studies that identify changes in brain structure that may play a key role in the recovery process.
A short primer on severe brain injuries
Disorders of Consciousness
Figure 1 organizes the relationships among several clinical syndromes often lumped into the category of ‘disorders of consciousness’. Coma and vegetative state (VS) are both considered unconscious brain states as judged by the bedside behavioral exam. In both syndromes, patients are entirely unresponsive to environmental stimuli and fail to initiate goal-directed behaviors. Comatose patients show no state variation and usually have closed eyes and no response to the most vigorous stimulation. In VS, patients have a cycling of irregular periods of eye opening and eye closure which does not correlate with identifiable electroencephalographic (EEG) features of either sleep or normal wakefulness [7]. In the minimally conscious state (MCS) [8] patients demonstrate unequivocal but inconsistent evidence of awareness of self or the environment through a wide variety of behavioral response patterns that can be demonstrated at the bedside [9]. The functional boundary indicating emergence from MCS is the demonstration of reliable verbal or gestural communication.
Figure 1. Correspondence of cognitive and motor impairment associated with disorders consciousness arising following severe brain injuries.
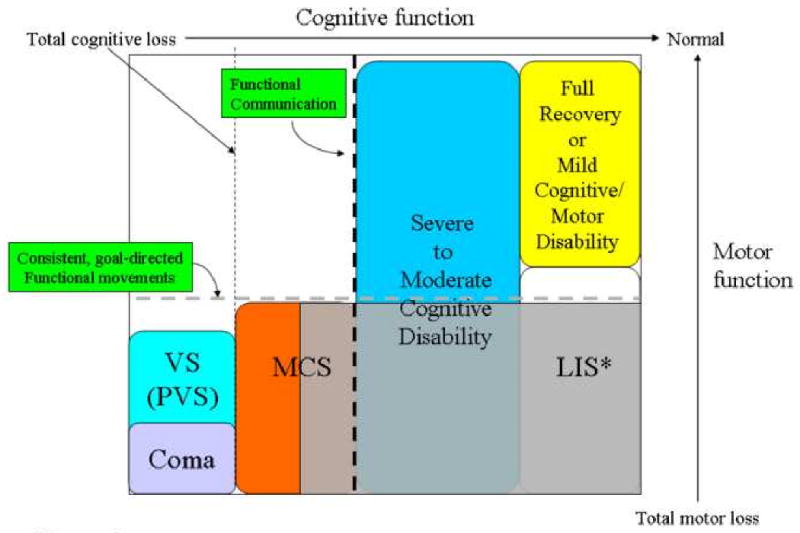
The distinctions among clinical disorders of consciousness can be best captured on a two-dimensional axis by comparing degree of impaired cognitive function against degree of motor function. At the bottom left of Figure the functional equivalence of coma and vegetative state (VS) as unconscious brain states is indicated by their placement to the left of the vertical dotted line indicating total loss of cognitive function. The large grey box in Figure 1, is placed to mark the high-degree of uncertainty associated with identifying the cognitive capacities of patients with no controllable motor output channel whose clinical bedside examination may range from minimally conscious state (MCS) to locked-in state (LIS), Note the * indicates that the locked-in state is not a disorder of consciousness and LIS patients retain normal cognitive function by definition. Establishing a true cognitive level for many patients who behaviorally cannot reliably signal through controlled goal-directed movements (dashed horizontal line) is possible at present. Such patients may retain varying levels of cognitive processing capabilities, awareness, memory and other higher brain functions without detection. Disentangling the potential for cognitive function in setting of severe limitations of motor control and sensorimotor integration mechanisms is among the most important challenges presented by new understanding of the recovery process following severe brain injuries.
Some fully conscious patients display a behavioral profile completely consistent with deep coma: eyes closed and unresponsive to any external stimuli as judged by a bedside examination. This condition is defined as the locked-in state (LIS; far right bottom of Figure 1). LIS is not a disorder of consciousness; by definition, LIS patients retain total preservation of cognitive function. LIS typically arises from neurological injuries that selectively disrupt the motor pathways or slowly reduce motor neuron function raising the probability of this diagnosis. The complexity of many brain injuries, however, creates a highly problematic set of patients who are unable to produce consistent goal-directed movements that allow for communication. Such individuals may retain significant cognitive capacity near the normal range of cognitive function and yet be indistinguishable from MCS patients.
Pathological findings in disorders of consciousness following severe brain injury
Anatomic pathologies associated with vegetative state, minimally conscious state, and severe to moderate cognitive disability following severe injuries have several common features. Autopsy studies of both traumatic and non-traumatic injuries resulting in permanent VS (a prognostic assessment rather than diagnosis, see [10]) identify widespread neuronal death throughout the thalamus in patients [11]. Importantly, the evident severe bilateral thalamic damage after either trauma or anoxia in permanent VS is not invariably associated with diffuse neocortical neuronal cell death. Moreover, the observation indicates the key functional role for the thalamus for integrative function of the forebrain corticothalamic systems.
Recent studies have shown that specific subnuclei of the thalamus demonstrate greater neuronal cell loss as a result of such global and multi-focal cerebral injuries [12]. The nuclei within the central thalamus (the intralaminar nuclei and related paralaminar nuclei) are most involved typically and the degree of neuronal loss observed within these neuronal aggregates grades with outcome [12]. In patients with only moderate disability following severe traumatic brain injury, neuronal loss is primarily identified within the anterior intralaminar nuclei (central lateral nucleus, central medial, paracentralis). Patients with progressively severe disabilities demonstrate neuronal loss involving more ventral and lateral nuclei of the central thalamus (posterior intralaminar group) as diagrammed in Figure 2A. These observations are likely a consequence of the unique geometry of connections of the central thalamus. Neurons in these subnuclei have wide point to point connectivity across the cerebral hemisphere and are thus likely to integrate neuronal cell death across these large territories [13,14].
Figure 2. Comparison of regions of central thalamus involved in focal and diffuse injuries producing global impairments of consciousness.
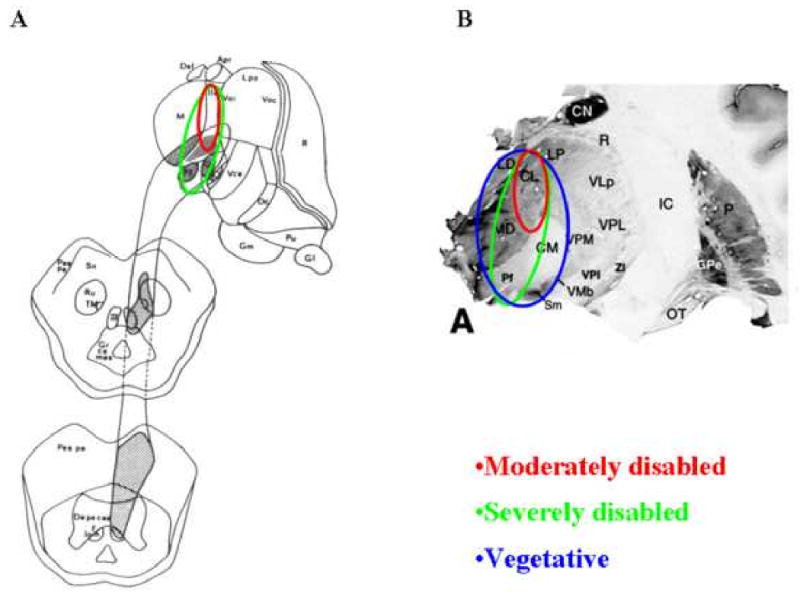
A. Focal injury patterns in the central thalamus associated with coma, vegetative state, and minimally conscious state (adapted from [15]). Red circle indicates anterior intralaminar nuclei and surrounding regions), green circle includes area of red circle and more caudal and medial components of the posterior intralaminar region. B. Regional neuronal cell loss in central thalamus following severe traumatic brain injuries indexed by functional outcomes [12]. Moderately disabled patients have cell loss restricted to the anterior intralaminar regions (red circle). Severely disabled patients have neuronal loss in more caudal and medial components of the central thalamus including the medial aspects of the posterior intralaminar nuclei. Permanent VS is associated with broad loss of central thalamic neurons including the large lateral component of the posterior intralaminar group (the centromedian nucleus) [14, 24]. Figure element of thalamic anatomy adapted from [74] with permission.
Importantly, the same selected thalamic subpopulations are known to produce global disorders of consciousness (coma, VS and minimally conscious state) following bilateral focal injuries [15, 16]. Figure 2B diagrams the overlap of the neuronal populations that undergo progressive deafferentation with increasingly severe multi-focal brain injuries and those typically involved in strokes producing initial coma and variable periods of VS and MCS [15,16]. As a consequence of diffuse brain injuries, considerable impact of either focal injury or deafferentation of these central thalamic neurons on forebrain function likely reflects their key contribution to normal mechanisms of arousal regulation [17]. Moreover, deafferentation and dysfunction of these neurons likely plays an important role in producing deficits even when injuries are not severe enough to produce broad neuronal death.
Functional specializations of the central thalamus
Neuromaging and electrophysiological studies demonstrate the selective activation of the central thalamus for tasks that require a short-term shift of attention [18. 19], sustained cognitive demands of high vigilance [20], or holding information in memory over extended time periods [19, 21]. The central thalamus is uniquely situated to support these broad ‘executive’ functions in the forebrain. Central thalamic neurons are strongly innervated by ascending projections from the brainstem/basal forebrain ‘arousal systems’ that control the activity of many cortical and thalamic neurons during the sleep-wake cycle and descending projections from frontal cortical systems that organize goal-directed behaviors and adjust the level of arousal associated with generalized alertness and variations in cognitive effort, stress, sleep deprivation, and other variables affecting the wakeful state [14, 18, 20, 22, 23 reviewed in 17]. The neurons within the central thalamus are further specialized, anatomically and physiologically, by their diffuse projections to supragranular layers of the cerebral cortex [14, 24-28] and striatal neurons [29-31]. Both the anterior (CL, Pc) and posterior intralaminar nuclei (centromedian-parafasicularis complex, Cm-Pf) and the mesencephalic reticular neurons that monosynaptically project to these neurons [32] activate during the short-term shifting of attention component of a forewarned reaction-time tasks. Activity in the central thalamus covaries with the anterior cingulate cortex (ACC) as well as the pontomesecephalon [20]. The ACC is similarly recruited by a wide range of cognitive demands and shows graded activity with increasing cognitive load, suggesting that this component of the frontal executive systems may drive, or reciprocally increase activity along with, the central thalamus in response to increasing demands of cognitive effort [33].
The central thalamus and the frontal lobe are closely linked through their direct corticothalamic connections, including supplementary motor, anterior cingular, premotor and prefrontal cortex [34] and indirect links through the frontal cortical-striatopallidal-thalamocortical loop systems [14, 24]. Behavioral fluctuations following central thalamic and frontal lobe injuries show strong quantitative and qualitative similarities in experimental behavioral lesion studies in rodents [35]. Similarly, notable fluctuations of behavioral response arise from both direct injuries to the central thalamus (either unilateral [36] or bilateral lesions and very closely resemble the typical behavioral fluctuations seen in patients and animals with frontal lobe lesions [38].
Linking time frames of recovery following severe brain injury to underlying pathophysiological mechanisms
To date the majority of longitudinal studies of recovery of consciousness after severe brain injury have focused on metrics that seek to predict the likelihood that a person will not recover past the VS after an initial coma (see [39,40] for reviews of the literature). This focus can be understood in light of important concerns of resource allocation in intensive care units and the high probability of death or permanent VS in coma following cardiac arrest or very severe traumatic brain injuries [40]. However, as immediate in-the-field care for patients with all types of severe brain injury has improved, increasingly large numbers of individuals not only survive their injuries but preserve correspondingly larger numbers of neurons and neuronal connections after the initial injury.
Well-established statistics guide the likelihood of permanent VS over time following some patterns of injury [10]. However, similar attempts to link time periods to outcome in patients who demonstrate the limited recovery patterns associated with MCS have shown a poor correlation of long-term outcome with comparable time-frames [1,41]. Most patients who demonstrate evidence of recovery to MCS within the first 3 months after injuries will recover past MCS by 10 months. Two to five year outcomes can include recovery past the level of severe disability even for patients who remain in MCS for greater than 6 months or a year. Rare cases demonstrate endpoints of very late recovery from MCS are documented including reemergence of higher functional levels of spoken conversation, autobiographical memory, and motor control after years and even decades [2, 42].
In part the differences in time frames for recovery reflect the differences in underlying pathology present in MCS and related outcomes of severe disability following brain injuries (compared with permanent VS). In these conditions, a mix of effects of neuronal death, deafferentation and dysfunction of remaining neuronal populations play a larger and considerably less well characterized role. The observations of late recovery from MCS indicate that brain networks may retain functional capacity without expression leaving an important possibility that marked changes in cognitive function may occur without bedside evidence either spontaneously or in response to interventions [2, 6].
Role of changes in brain structure in the recovery process
Recent studies provide evidence that late recovery of function following severe brain injury may involve structural changes within the brain. Structural magnetic resonance imaging studies of a man who at age 40 spontaneously recovered full expressive and receptive language, after remaining in MCS for 19 years following a severe traumatic brain injury suffered in a motor vehicle accident, revealed evidence of ongoing structural modifications [2]. Using diffusion tensor magnetic resonance imaging (DTI), a technique that quantifies the anisotropy of proton diffusion and thus is a proxy for axonal fiber integrity, extensive cerebral and subcortical atrophy was noted in brainstem and frontal lobes. Despite evidence of widespread white matter injury, a longitudinal DTI study of the brain identified regions that showed significant change over time. Notably, the midline cerebellar white matter showed increased fractional anisotropy in a second study that correlated with clinical improvements in motor control. In a recent prospective cohort study of severely brain injured patients followed for a year following initial injury, similar changes in DTI measured fractional anisotropy were identified in the patients who recovered neurological function [43]. In the aggregate, these observations suggest that the normal recovery process also includes a component of structural remodeling that may plausibly relate to reestablishment of goal-behaviors and driving of learning and memory mechanisms. However, why such processes may arise at late intervals or not at all requires an examination of potential mechanisms underlying large-scale changes in forebrain dynamics following severe injuries.
A ‘mesocircuit’ hypothesis
As reviewed above, regularities in the anatomic pathology of different types of severe brain injury suggest that large-scale forebrain dysfunction may arise as a result of at least three general mechanisms: 1) widespread death of forebrain neurons (i.e. sufficient to produce brain death or permanent VS), 2) widespread deafferentation and disconnection of neurons, and 3) “circuit”-level functional disturbances due to the loss of these neuronal connections [3, 17, 42, 44]. While the first mechanism is clearly irreversible, some evidence (as reviewed above) suggests that late structural alterations in the brain may arise, altering the effects of the second mechanism. Alterations at the third “mesocircuit” [45] level may arise as a result of global decreases of excitatory neurotransmission producing overall changes in cerebral background activity levels (as produced by anesthesia or direct effects on the function of certain cell types; e.g. hypoxia, see discussion below).
Figure 3 illustrates a key vulnerability of the anterior forebrain in the setting of widespread deafferentation and neuronal cell loss that may represent the common denominator in disturbances of consciousness in severe brain injuries. The primary result of disturbances of this network may be to effectively produce a broad decrease in background synaptic activity and excitatory neurotransmission (e.g. diffuse axonal injury, anoxia, hypoxia-ischemia, multi-focal infarction following cerebral vasospam, encephalitis, etc; see [40]). At a neuronal subpopulation level, the medium spiny neurons (MSN) of the striatum have a key role in maintaining activity in the anterior forebrain through their inhibitory projections to the globus pallidus interna which in turn inhibits the central thalamus [29, 46]. Activation of MSN projections, de facto, results in a disinhibition of central thalamic neurons, reestablishes the outflow of thalamocortical transmission and likely promotes a rebound of high frequency thalamocortical activity [75]. The thalamocortical projections from the central thalamus strongly innervate the frontal cortex and have in some instances a joint thalamostriatal projection back to the MSNs [29]; recent studies demonstrate that thalamocortical projections to cortex have a stronger impact on driving excitation within the cortex than cortico-cortical projections [47] and down-regulation of thalamic output can be expected to have broad effects across cortical regions.
Figure 3. Proposed “mesocircuit” model underlying forebrain dysfunction and interventions in severe brain injuries.
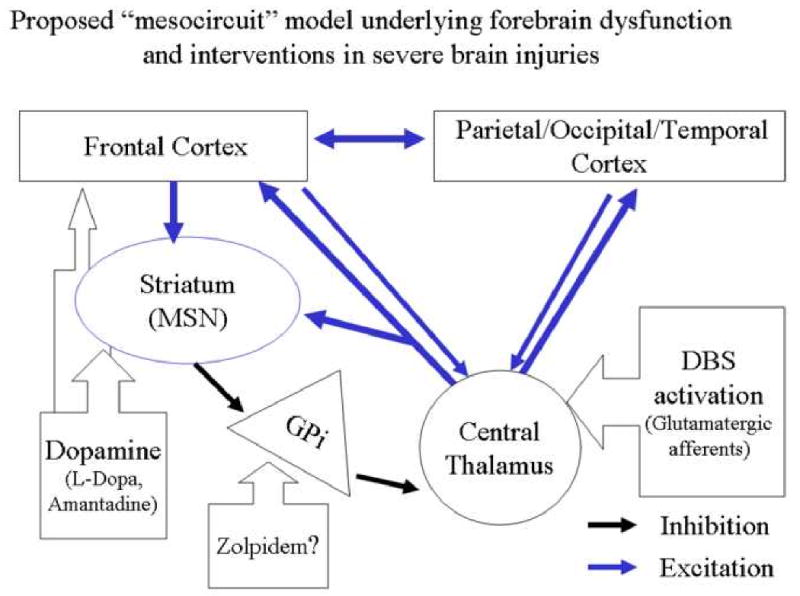
A proposed ‘mesocircuit’ that explains the vulnerability of the anterior forebrain (frontal/prefrontal cortical-striatopallidal thalamocortical loop systems) following multi-focal brain injuries that produce widespread deafferentation or neuronal cell loss. The thalamocortical projections of the central thalamus are proposed to play an important role in observed reduction of cerebral metabolism in this mesocircuit following different mechanisms of brain injury [42, 48]; these projections have a strong activating role strongly driving both cortical and striatal neurons [47, 30,31]. The medium spiny neurons (MSN) of the striatum which send inhibitory projections to the globus pallidus interna require high levels of background synaptic activity and dopaminergic neuromodulation to maintain firing rates [46]. Without MSN output the globus pallidus interna tonically inhibits the central thalamus potentially catalyzing a shut down of the anterior forebrain. Down-regulation of activity within the mesocircuit is predicted to have a broad modulatory impact on the global dynamics of the dominant corticothalamic system [25, 26, 28, 32, 73]; specific changes within the cortico-striatopallidal thalamocortical system identified with alterations of consciousness associated with sleep and anesthesia support this inference [55, 58, 61]. The mesocircuit model also economically accounts for the mix of interventions that have been noted in some patients to restore functions associated with these forebrain systems (e.g. dopaminergic agents, zolpidem, and electrical brain stimulation; see text for further discussion).
Neurons from the central thalamus (both central lateral nucleus and parafasicularis nucleus) strongly project to the MSNs [30] and diffusely innervate the striatum [29]. These thalamostriatal projections use glutamate transmitter proteins with a high probability of synaptic release [31] and may have a strong role in modulating background activity in the striatum. The MSNs have a ‘high threshold’ UP state that keeps them below their firing threshold unless sufficient levels of dopamine neuromodulation are present and there is a high level of spontaneous background synaptic activity arising from excitatory corticostriatal and thalamostriatal inputs [46]. Thus, diffuse brain injuries may lead to a sharp reduction of MSN output as diffuse deafferentation produces withdrawal of both direct excitatory striatal projections from the central thalamus [30] and down-regulation of the frontocortical regions that provide the main corticostriatal input. Among frontal cortical regions, the anterior cingulate cortex may play an essential role as it receives strong inputs for the anterior intralaminar nuclei (central lateral nucleus, [34]) and provide a very diffuse regulatory input across large territories of the rostral striatum [47].
Implications of the mesocircuit model for recovery of consciousness after severe brain injury
The mesocircuit model in Figure 3 organizes and rationalizes recent observations of the response of severely brain-injured subjects to pharmacological and electrophysiological interventions as well as some aspects of normal brain function, as reviewed below. The primary implication of the model in Figure 3 is that frontocortico-striatopallidal-thalamocortical loop frontal systems are selectively vulnerable at the ‘circuit’ level in many types of multi-focal brain injury. This accounts for the observations that selective metabolic depression of the anterior forebrain specifically grades with severity of behavioral impairment following diffuse axonal injury [48]. In addition, the well-known response to dopaminergic agents of severely brain-injured patients with markedly slowed behavioral response following either mesial frontal lobe, basal forebrain, or thalamic/midbrain injuries is consistent with the mesocircuit model [49, 50]. Behavioral features of these patients range from extreme poverty of movement (‘akinetic mutism”) to severe disability characterized by very slow but nonetheless accurate responses that allow communication [37, 51]. Dopaminergic facilitation of the output of the MSNs or direct modulation of mesial frontal cortical neurons would explain the restoration of anterior forebrain activity within the loop connections of the frontal cortex, striatum, pallidum and central thalamus.
Zolpidem induced paradoxical arousal in severe brain injury
Of particular interest, the mesocircuit model also offers an explanation of a surprising and puzzlingly paradoxical phenomenon recently described that zolpidem (a non-benzodiazepine hypnotic that potentiates GABA A receptors, also known as ‘Ambien’) can improve alertness and behavioral responsiveness in some severely brain injured patients [42, 52-54]. Brefel Courbon et al [42] reported an MCS patient who recovered spoken language, eating and ambulation with zolpidem administration. Figure 4 shows a marked increase in anterior forebrain metabolism associated with zolpidem administered condition compared to the patient's off drug state. Similar observations in another zolpidem responsive patient [55] link increases of cerebral metabolism in the frontal cortex, striatum and thalamus to changes in the shape of the spectral content of the EEG (removing abnormal low frequency component) and the coherence architecture (reducing marked low frequency coherence in the off drug state). Consonant with the model of Figure 3, Schiff and Posner [44] proposed the following mechanism for this paradoxical response. Under normal circumstances, the MSN's dis-inhibit the central thalamus via the GPi (see Figure 3). Thus, when MSN activity is reduced as a consequence of brain injury, central thalamic activity is also reduced. Since zolpidem directly inhibits the GPi, it can substitute for the normal inhibition of the GPi from MSN's, and thus permit a more normal level of central thalamic activity. The GABA-A alpha-1 subunit is expressed in large quantities in the globus pallidus interna and experimental studies support this mechanism of action [72]. Of note, the MSNs are uniquely vulnerable to cellular dysfunction after hypoxia [56] and several of the reported cases of paradoxical response have followed hypoxic-ischemic injuries [42, 53-55].
Figure 4. Changes in cerebral metabolism associated with zolpidem administration in severe brain-injury.
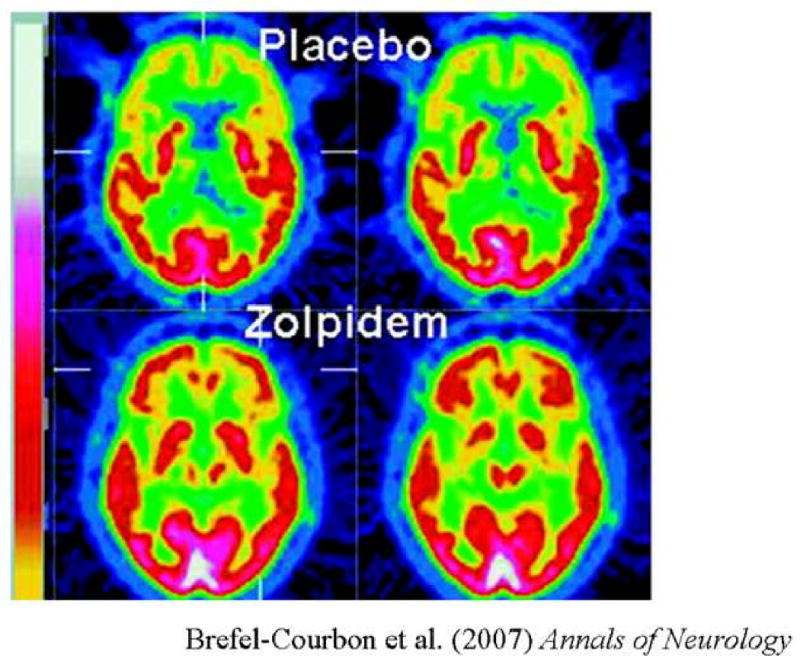
Fluorodeoxyglucose positron emission tomography studies by Brefel-Courbon et al. [42] of a severe brain injured patient in minimally conscious state before and after administration of the sedative agent zolpidem (‘Ambien’). In the off drug state (top panels) marked anterior forebrain hypometabolism is noted bilaterally in frontal/prefrontal cortex, thalami, and striatum. Following zolpidem administration broad increases of metabolic rates are observed in these regions.
In addition to accounting for the paradoxical response to zolpidem, the mesocircuit model provides a plausible framework for related observations in normal subjects. Of note, the model provides an explanation for the observation that the most robust changes in regional cerebral blood flow during the transitions during the sleep-wake cycle are in the striatum [57]. Specifically, increases during the transition from slow wave sleep to rapid eye movement sleep (REM) and decreases in the transition from wakefulness to non-REM sleep. Similar, slightly less significant changes also occur in the ‘centrencephalic’ components of the thalamus and cerebral cortex [57]. The ‘circuit breaker’ effect of withdrawal of cortical and thalamic excitation from the MSN suggests an economical explanation for this otherwise puzzling contribution of the striatum. Similar recovery patterns in metabolic activity of the anterior forebrain are seen during early wakefulness as sleep inertia dissipates [58]. The model suggesting reactivation of the frontostriatal systems during sleep states may also provide an explanation for a variety of reports of unusual behaviors (somnambulism, amnestic hyperphagia—nocturnal binge eating without memory trace) arising during sleep specifically associated zolpidem treatment[59, 60]. Finally, this mesocircuit model can also account for the common finding of the early selective metabolic downregulation in the mesial frontal and thalamic systems with different anesthetics [61] and variety of specific changes across the induction and recovery from general anesthesia (see [46])
Electrical stimulation of central thalamus in minimally conscious state
Markedly depressed rates of global metabolism are seen in patients with MCS or severe disability ([62]) and may be produced either by volume loss of neurons and subsequent deafferentation of remaining cells or by neuronal functional impairments and low firing rates. The mesocircuit model shown in Figure 3 predicts that direct activation of excitatory output from the central thalamus in patients with chronically downregulated background synaptic activity following severe brain injury will tend to normalize normal cortico-striatopallidal-thalamocortical function. Direct activation of central thalamic neurons through electrical stimulation (‘deep brain stimulation’, DBS) has been proposed as an experimental therapeutic strategy that might produce consistent and sustained effects of maintaining the activity within this circuit [63]. A recent single-subject study demonstrated that central thalamic DBS restored arousal regulation and promoted improved behavioral responsiveness in a 38 year old man after remaining in MCS for 6 years [64]. The patient had remained unable to communicate reliably despite neuroimaging evidence of preservation of large scale cerebral language networks [65] that suggested a substrate for further recovery. Figure 5 illustrates overall design of the study, placement of the electrodes in the central thalamus, and main results.
Figure 5. Central thalamic DBS in the minimally conscious state.
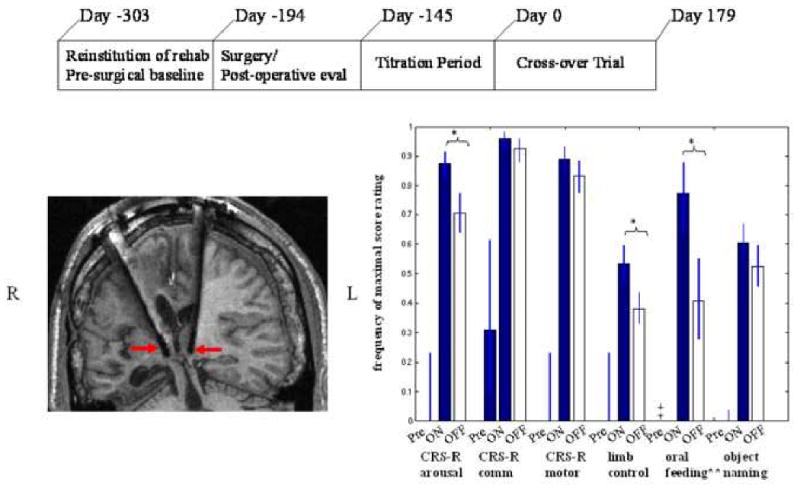
A. Timeline of single-subject study of deep brain stimulation in the central thalamus in a patient remaining in MCS for 6 years B. Location of electrode lead placements within central thalamus of patient's right (R) and left (L) hemispheres displayed in T1 weighted MRI coronal image and marked with red arrows. C. Comparison of pre-surgical baselines and DBS ON and DBS OFF periods during a six month cross-over trial. Behavioral baseline evaluations measured using a standardized quantitative assessment tool, the Coma Recovery Scale Revised (CRS-R) were obtained 4 months prior to surgery and for 2 months following surgery prior to titration testing of stimulation parameters [see Ref #64 for details] showed no change in behavioral responsiveness as compared to functional levels measured more than 2 years before the start of the trial. During the early titration testing of electrical stimulation the patient demonstrated immediate and accumulating effects of DBS that included the emergence of consistent and intelligible spoken language, recovery of limb control and the capacity for oral feeding (see [64] for details). Following a 5 month titration period of testing combinations of stimulation parameters the patient entered into a blinded six-month, 30 day alternating ON versus OFF (cross-over) study that demonstrated a robust overall effect on behavioral responsiveness measured by CRS-R subscales and supplementary behavioral rating scales. Significant ON versus OFF improvements with electrical were demonstrated for attentive behavior, oral feeding and limb control. All functional testing showed significant improvements when compared against the 6 month pre-stimulation baselines. Importantly, observed carryover effects of improvements from the ON to the OFF state were also identified as seen by the high frequency of OFF stimulation ratings compared to pre-stimulation baselines across all measurements. These effects are comparable to evidence of accumulating behavioral effects of central thalamic electrical stimulation as shown in a rodent model by Herrera and colleagues [66]. Figure elements adapted from Schiff et al [64] with permission.
Dissociation of expressed motor behavior and integrative cerebral function
The need to develop better models for understanding cognitive capacity after severe brain injury is dramatically illustrated by the study of Owen et al. [6]. Via fMRI, these authors demonstrated high-level cognitive function in a patient behaviorally judged to be in VS for 5 months following a severe traumatic brain injury. When asked to imagine playing tennis the patient exhibited significant fMRI measured brain activation in the supplementary motor areas and when asked to imagine walking through the rooms of her house showed activation in parahippocampal gyrus, posterior parietal cortex, and the lateral premotor cortex; both patterns are consistent with those seen in normal control subjects carrying out this task [67]. These observations provide unambiguous demonstrations of command following--a cardinal sign distinguishing VS from MCS [9].
The Owen study raises many important questions. The most critical question is what mechanisms might underlie the failure of the patient to exhibit goal-directed behavior despite the apparent integrity of motor pathways? Owen et al. interrogated the integrity of the patient's motor pathways using transcranial magnetic stimulation methods and ruled out an interruption of the outflow from the motor cortex to the skeletal muscles accounting for her lack of initiated movements [6]. Collectively, the observations indicate that although this patient could follow commands, she likely remained unable to organize motor responses to carry out goal-directed intentional behaviors because of functional disturbances of forebrain systems associated with motor preparation and action. As noted above, the mesocircuit model provides a parsimonious hypothesis that can be tested to explain these findings. Notably at the time of study the patient had collapsed regions of skull bilaterally across the frontal lobe visible in the imaging results, see [6] demonstrating a marked impact on the frontal systems and suggesting a possible mechanism for persistent dysfunction of the anterior forebrain [73] Clearly, better understanding of the mechanisms underlying these observations will lead to an improved ability to prospectively identify and risk-stratify patients to improve the likelihood of obtaining accurate diagnoses and facilitating recovery of communication.
Recent computational modeling studies provide potential insight into the functional role of these long loop frontal-striatopallidal-thalamocortical systems that appear to be selectively vulnerable to shut down after severe brain injury. Goldman [68] has shown that sequentially feedforward circuits (the striatopallidal components adds asymmetry to the standard corticothalamic circuit) provides a solution to organizing arbitrary temporal processing demands (such as flexibly reconfiguring sensorimotor contingencies) and holding this information over the long time scales associated with cognition. The model's feedforward architecture of connections produces response profiles similar to those recorded across frontal cortex [69], striatum [70], and central thalamus [19,21] during performance of ‘executive functions’ such as sustained attention, working memory, or motor preparation. This model provides an attractive first-order explanation of the central importance of the anterior forebrain mesocircuit in the global behavioral impairments seen after traumatic brain injury. Graded and variable recovery of function of the entire circuit may the variable that best indexes fluctuations in behavioral responsiveness. Reestablishing functional activity across these long loops over time is likely required for the minimal behavioral capacity required to advance motor behaviors beyond the MCS level. Gradually improving the integrity of normal activity patterns within the anterior forebrain may underlie the continuum of outcomes across severe disability whereby the probability of maintaining the anterior forebrain mesocircuit corresponds to different functional outcomes.
Future Directions
Understanding the circuit mechanisms associated with phases of recovery of consciousness following severe brain injuries will open many directions for future research including 1) the development of new diagnostic tools based on neuroimaging and electrophysiological measurements to guide longitudinal assessments of brain function, and 2) the development of novel interventions at the circuit and cellular level to aid recovery. A key overarching goal of these efforts is to identify the potential for communication and support this capacity.
Because of the intermittent nature of behaviors in MCS it is essential to develop tools to more accurately assess patients. Improving the consistency of observed behaviors, or the providing means for detecting potentially more reliable underlying neurophysiological signals that may enable consistent basic communication is important, even if cognitive capacity remains severely restricted. As Fins [71] has recently argued, functional communication represents a major milestone for all patients with severe brain injury across the diagnostic spectrum, their caregivers, and family members as it restores a fundamental aspect of the patient's capacity to reengage the human community and reestablish essential aspects of personhood.
Acknowledgments
This paper was initially presented as a Special Lecture at the Society of Neuroscience 38th Annual Meeting on November 18, 2008. The author thanks Jonathan Victor, Shawniqua Williams, Sudhin Shah, and Mary Conte. The support of the James S. McDonnell Foundation, the NIH-NICHD are gratefully acknowledged.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Lammi MH, Smith VH, Tate RL, Taylor CM. The minimally conscious state and recovery potential: a follow-up study 2 to 5 years after traumatic brain injury. Arch Phys Med Rehabil. 2005;86(4):746–54. doi: 10.1016/j.apmr.2004.11.004. [DOI] [PubMed] [Google Scholar]
- 2.Voss HU, et al. Possible axonal regrowth in late recovery from minimally conscious state. Journal of Clinical Investigation. 2006;116:2005–2011. doi: 10.1172/JCI27021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Burruss JW, Chacko RC. Episodically remitting akinetic mutism following subarachnoid hemorrhage. J Neuropsychiatry Clin Neurosci 1999. 1999;11(1):100–2. doi: 10.1176/jnp.11.1.100. [DOI] [PubMed] [Google Scholar]
- 4.McMillan TM, Herbert CM. Further recovery in a potential treatment withdrawal case 10 years after brain injury. Brain Inj. 2004;18(9):935–40. doi: 10.1080/02699050410001675915. [DOI] [PubMed] [Google Scholar]
- 5.Macniven JA, Poz R, Bainbridge K, Gracey F, Wilson BA. Emotional adjustment following cognitive recovery from ‘persistent vegetative state’: psychological and personal perspectives. Brain Inj. 2003;17(6):525–33. doi: 10.1080/0269905031000086254. [DOI] [PubMed] [Google Scholar]
- 6.Owen AM, et al. Detecting awareness in the vegetative state. Science. 2006;313(5792):1402. doi: 10.1126/science.1130197. [DOI] [PubMed] [Google Scholar]
- 7.Kobylarz EJ, Schiff ND. Neurophysiological correlates of persistent vegetative and minimally conscious states. Neuropsychological Rehabilitation. 2005;15:323–332. doi: 10.1080/09602010443000605. [DOI] [PubMed] [Google Scholar]
- 8.Giacino JT, Ashwal S, Childs N, Cranford R, Jennett B, Katz DI, Kelly JP, Rosenberg JH, Whyte J, Zafonte RD, Zasler ND. The minimally conscious state: definition and diagnostic criteria. Neurology. 2002;58:349–53. doi: 10.1212/wnl.58.3.349. [DOI] [PubMed] [Google Scholar]
- 9.Giacino JT, Whyte J. The vegetative state and minimally conscious state: current knowledge and remaining questions. J Head Trauma Rehabilitation. doi: 10.1097/00001199-200501000-00005. [DOI] [PubMed] [Google Scholar]
- 10.Jennett B. The Vegetative State. Cambridge University Press; 2002. [Google Scholar]
- 11.Adams JH, Graham DI, Jennett B. The neuropathology of the vegetative state after acute insult. Brain. 2000;123:1327–1338. doi: 10.1093/brain/123.7.1327. [DOI] [PubMed] [Google Scholar]
- 12.Maxwell WL, et al. Thalamic nuclei after human blunt head injury. J Neuropathol Exp Neurol. 2006;65(5):478–88. doi: 10.1097/01.jnen.0000229241.28619.75. [DOI] [PubMed] [Google Scholar]
- 13.Scannell JW, et al. The connectional organization of the cortico-thalamic system of the cat. Cereb Cortex. 1999;9(3):277–99. doi: 10.1093/cercor/9.3.277. [DOI] [PubMed] [Google Scholar]
- 14.Van der Werf YD, Witter MP, Groenewegen HJ. The intralaminar and midline nuclei of the thalamus. Anatomical and functional evidence for participation in processes of arousal and awareness. Brain Res Brain Res Rev. 2002;39(2-3):107–40. doi: 10.1016/s0165-0173(02)00181-9. [DOI] [PubMed] [Google Scholar]
- 15.Castaigne P, et al. Paramedian thalamic and midbrain infarcts: clinical and neuropathological study. Ann Neurol. 1981;10(2):127–48. doi: 10.1002/ana.410100204. [DOI] [PubMed] [Google Scholar]
- 16.Schiff ND, Plum F. The role of arousal and ‘gating’ systems in the neurology of impaired consciousness. Journal of Clin Neurophysiol. 2000;17:438–452. doi: 10.1097/00004691-200009000-00002. [DOI] [PubMed] [Google Scholar]
- 17.Schiff ND. Central thalamic contributions to arousal regulation and neurological disorders of consciousness. Annals of New York Academy of Sciences. 2008;1129:105–118. doi: 10.1196/annals.1417.029. [DOI] [PubMed] [Google Scholar]
- 18.Kinomura S, et al. Activation by attention of the human reticular formation and thalamic intralaminar nuclei. Science. 1996;271:512–515. doi: 10.1126/science.271.5248.512. [DOI] [PubMed] [Google Scholar]
- 19.Shah S, Baker J, Ryou JW, Purpura KP, Schiff ND. Modulation of arousal regulation with central thalamic deep brain stimulation. Conf Proc IEEE. 2009 doi: 10.1109/IEMBS.2009.5333751. in press. [DOI] [PubMed] [Google Scholar]
- 20.Paus T, et al. Time-related changes in Neural systems underlying attention and arousal during the performance of an auditory vigilance task. Jour Cognitive Neurosci. 1997;9:392–408. doi: 10.1162/jocn.1997.9.3.392. [DOI] [PubMed] [Google Scholar]
- 21.Wyder MT, Massoglia DP, Stanford TR. Contextual modulation of central thalamic delay-period activity: representation of visual and saccadic goals. J Neurophysiol. 2004;91(6):2628–48. doi: 10.1152/jn.01221.2003. [DOI] [PubMed] [Google Scholar]
- 22.Paus T, et al. Regional differences in the effects of task difficulty and motor output on blood flow response in the human anterior cingulate cortex: a review of 107 PET activation studies. Neuroreport. 1998;9(9):R37–47. doi: 10.1097/00001756-199806220-00001. [DOI] [PubMed] [Google Scholar]
- 23.Nagai Y, et al. Brain activity relating to the contingent negative variation: an fMRI investigation. Neuroimage. 2004;21(4):1232–41. doi: 10.1016/j.neuroimage.2003.10.036. [DOI] [PubMed] [Google Scholar]
- 24.Gronewegen H, Berendse H. The specificity of the ‘nonspecific’ midline and intralaminar thalamic nuclei. Trends in Neuroscience. 1994;17:52–66. doi: 10.1016/0166-2236(94)90074-4. Gronewegen and Berendse 1994. [DOI] [PubMed] [Google Scholar]
- 25.Llinas RR, Leznik E, Urbano FJ. Temporal binding via cortical coincidence detection of specific and nonspecific thalamocortical inputs: a voltage-dependent dye-imaging study in mouse brain slices. Proc Natl Acad Sci. 2002;99:449–54. doi: 10.1073/pnas.012604899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Jones EG. The thalamic matrix and thalamocortical synchrony. Trends Neurosci. 2001;24:595–601. doi: 10.1016/s0166-2236(00)01922-6. [DOI] [PubMed] [Google Scholar]
- 27.Purpura KP, Schiff ND. The thalamic intralaminar nuclei: role in visual awareness. Neuroscientist. 1997;3:8–14. Purpura and Schiff. [Google Scholar]
- 28.Steriade M, Curró Dossi R, Contreras D. Electrophysiological properties of intralaminar thalamocortical cells discharging rhythmic (approximately 40 HZ) spike-bursts at approximately 1000 HZ during waking and rapid eye movement sleep. Neuroscience. 1993;56(1):1–9. doi: 10.1016/0306-4522(93)90556-u. [DOI] [PubMed] [Google Scholar]
- 29.Deschenes M, Bourassa J, Parent A. Striatal and cortical projections of single neurons from the central lateral thalamic nucleus in the rat. Neuroscience. 1996;72(3):679–87. doi: 10.1016/0306-4522(96)00001-2. [DOI] [PubMed] [Google Scholar]
- 30.Lacey CJ, Bolam JP, Magill PJ. Novel and distinct operational principles of intralaminar thalamic neurons and their striatal projections. J Neurosci. 2007;27(16):4374–4384. doi: 10.1523/JNEUROSCI.5519-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Smith Y, Raju D, Nanda B, Pare JF, Galvan A, Wichmann T. The thalamostriatal systems: anatomical and functional organization in normal and parkinsonian states. Brain Res Bull. 2009;78(2-3):60–8. doi: 10.1016/j.brainresbull.2008.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Steriade M, Glenn LL. Neocortical and caudate projections of intralaminar thalamic neurons and their synaptic excitation from midbrain reticular core. J Neurophysiol. 1982;48:352–71. doi: 10.1152/jn.1982.48.2.352. [DOI] [PubMed] [Google Scholar]
- 33.Stuss DT, Alexander MP. Is there a dysexecutive syndrome? Philos Trans R Soc Lond B Biol Sci. 2007;362(1481):901–15. doi: 10.1098/rstb.2007.2096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Morel A, et al. Divergence and convergence of thalamocortical projections to premotor and supplementary motor cortex: a multiple tracing study in the macaque monkey. Eur J Neurosci. 2005;21(4):1007–29. doi: 10.1111/j.1460-9568.2005.03921.x. [DOI] [PubMed] [Google Scholar]
- 35.Mair RG, Burk JA, Porter MC. Lesions of the frontal cortex, hippocampus, and intralaminar thalamic nuclei have distinct effects on remembering in rats. Behav Neurosci. 1998;112(4):772–92. doi: 10.1037//0735-7044.112.4.772. [DOI] [PubMed] [Google Scholar]
- 36.Van Der Werf YD, et al. Neuropsychological correlates of a right unilateral lacunar thalamic infarction. J Neurol Neurosurg Psychiatry. 1999;66:36–42. doi: 10.1136/jnnp.66.1.36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Stuss DT, Guberman A, Nelson R, Larochelle S. The neuropsychology of paramedian thalamic infarction. Brain Cogn. 1988;8:348–378. doi: 10.1016/0278-2626(88)90059-0. [DOI] [PubMed] [Google Scholar]
- 38.Robertson IH, et al. ‘Oops!’: performance correlates of everyday attentional failures in traumatic brain injured and normal subjects. Neuropsychologia. 1997;35:747–758. doi: 10.1016/s0028-3932(97)00015-8. [DOI] [PubMed] [Google Scholar]
- 39.Young GB. Clinical practice. Neurologic prognosis after cardiac arrest. N Engl J Med. 2009;361(6):605–11. doi: 10.1056/NEJMcp0903466. [DOI] [PubMed] [Google Scholar]
- 40.Posner J, Saper C, Schiff N, Plum F. Plum and Posner's Diagnosis of Stupor and Coma. 4th. Oxford University Press; 2007. [Google Scholar]
- 41.Giacino JT, Kalmar K. The Vegetative and Minimally Conscious States: A Comparison of Clinical Features and Functional Outcome. Journal of Head Trauma Rehabilitation. 1997;12:36–51. [Google Scholar]
- 42.Brefel-Courbon C, et al. Clinical and imaging evidence of zolpidem effect in hypoxic encephalopathy. Ann Neurol. 2007;62(1):102–5. doi: 10.1002/ana.21110. [DOI] [PubMed] [Google Scholar]
- 43.Sidaros A, Engberg AW, Sidaros K, Liptrot MG, Herning M, Petersen P, Paulson OB, Jernigan TL, Rostrup E. Diffusion tensor imaging during recovery from severe traumatic brain injury and relation to clinical outcome: a longitudinal study. Brain. 2008;131:559–72. doi: 10.1093/brain/awm294. [DOI] [PubMed] [Google Scholar]
- 44.Schiff ND, Posner JP. Another “Awakenings”. Annals of Neurology. 2007;62:5–7. doi: 10.1002/ana.21158. [DOI] [PubMed] [Google Scholar]
- 45.Bohland JW, et al. A Proposal for a Coordinated Effort for the Determination of Brainwide Neuroanatomical Connectivity in Model Organisms at a Mesoscopic Scale. PLoS Comput Biol. 2009;5(3):e1000334. doi: 10.1371/journal.pcbi.1000334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Grillner S, et al. Mechanisms for selection of basic motor programs--roles for the striatum and pallidum. Trends Neurosci. 2005;28(7):364–70. doi: 10.1016/j.tins.2005.05.004. [DOI] [PubMed] [Google Scholar]
- 47.Rigas P, Castro-Alamancos MA. Thalamocortical Up states: differential effects of intrinsic and extrinsic cortical inputs on persistent activity. J Neurosci. 2007;27(16):4261–72. doi: 10.1523/JNEUROSCI.0003-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kato T, et al. Statistical image analysis of cerebral glucose metabolism in patients with cognitive impairment following diffuse traumatic brain injury. J Neurotrauma. 2007;24(6):919–26. doi: 10.1089/neu.2006.0203. [DOI] [PubMed] [Google Scholar]
- 49.Matsuda W, Matsumura A, Komatsu Y, Yanaka K, Nose T. Awakenings from persistent vegetative state: report of three cases with parkinsonism and brain stem lesions on MRI. J Neurol Neurosurg Psychiatry. 2003;74:1571–1573. doi: 10.1136/jnnp.74.11.1571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Meythaler JM, Brunner RC, Johnson A, Novack TA. Amantadine to improve neurorecovery in traumatic brain injury-associated diffuse axonal injury: a pilot double-blind randomized trial. J Head Trauma Rehabil. 2002;17(4):300–13. doi: 10.1097/00001199-200208000-00004. [DOI] [PubMed] [Google Scholar]
- 51.Katz DI, Alexander MP, Mandell AM. Dementia following strokes in the mesencephalon and diencephalon. Arch Neurol. 1987;44:1127–1133. doi: 10.1001/archneur.1987.00520230017007. [DOI] [PubMed] [Google Scholar]
- 52.Whyte J, Myers R. Incidence of clinically significant responses to zolpidem among patients with disorders of consciousness: a preliminary placebo controlled trial. Am J Phys Med Rehabil. 2009;88(5):410–8. doi: 10.1097/PHM.0b013e3181a0e3a0. [DOI] [PubMed] [Google Scholar]
- 53.Shames JL, Ring H. Transient reversal of anoxic brain injury-related minimally conscious state after zolpidem administration: a case report. Arch Phys Med Rehabi. 2008;89(2):386–8. doi: 10.1016/j.apmr.2007.08.137. [DOI] [PubMed] [Google Scholar]
- 54.Cohen SI, Duong TT. Increased arousal in a patient with anoxic brain injury after administration of zolpidem. Am J Phys Med Rehabil. 2008;87(3):229–31. doi: 10.1097/PHM.0b013e318161971b. [DOI] [PubMed] [Google Scholar]
- 55.Williams S, Conte MM, Kobylarz EJ, Hersh J, Victor JD, Schiff ND. Quantitative neurophysiologic characterization of a paradoxical response to zolpidem in a severely brain-injured human subject. Society for Neuroscience Meeting Abstract 2009 [Google Scholar]
- 56.Calabresi P, Centonze D, Bernardi G. Cellular factors controlling neuronal vulnerability in the brain: a lesson from the striatum. Neurology. 2000;55(9):1249–55. doi: 10.1212/wnl.55.9.1249. [DOI] [PubMed] [Google Scholar]
- 57.Braun AR, Balkin TJ, Wesenten NJ, Carson RE, Varga M, Baldwin P, Selbie S, Belenky G, Herscovitch P. Regional cerebral blood flow throughout the sleep-wake cycle. An H2(15)O PET study. Brain. 1997;120(Pt 7):1173–97. doi: 10.1093/brain/120.7.1173. [DOI] [PubMed] [Google Scholar]
- 58.Balkin TJ, Braun AR, Wesensten NJ, Jeffries K, Varga M, Baldwin P, Belenky G, Herscovitch P. The process of awakening: a PET study of regional brain activity patterns mediating the re-establishment of alertness and consciousness. Brain. 2002;125(Pt 10):2308–19. doi: 10.1093/brain/awf228. [DOI] [PubMed] [Google Scholar]
- 59.Yang W, Dollear M, Muthukrishnan SR. One rare side effect of zolpidem--sleepwalking: a case report. Arch Phys Med Rehabil. 2005;86(6):1265–6. doi: 10.1016/j.apmr.2004.11.022. [DOI] [PubMed] [Google Scholar]
- 60.Najjar M. Zolpidem and amnestic sleep related eating disorder. Clin Sleep Med 2007. 2007 Oct 15;3(6):637–8. [PMC free article] [PubMed] [Google Scholar]
- 61.Alkire MT, Hudetz AG, Tononi G. Consciousness and anesthesia. Science. 2008;322(5903):876–80. doi: 10.1126/science.1149213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Laureys S, Owen AM, Schiff ND. Brain function in coma, vegetative state, and related disorders. Lancet Neurol. 2004;3(9):537–46. doi: 10.1016/S1474-4422(04)00852-X. [DOI] [PubMed] [Google Scholar]
- 63.Schiff ND, Purpura KP. Towards a neurophysiological basis for cognitive neuromodulation through deep brain stimulation. Thalamus and Related Systems. 2002;2:55–69. [Google Scholar]
- 64.Schiff ND, Giacino JT, Kalmar K, Victor JD, Baker K, Gerber M, Fritz B, Eisenberg B, Biondi T, O'Connor J, Kobylarz EJ, Farris S, Machado A, McCagg C, Plum F, Fins JJ, Rezai AR. Behavioral improvements with thalamic stimulation after severe traumatic brain injury. Nature 2007. 2007;448:600–603. doi: 10.1038/nature06041. [DOI] [PubMed] [Google Scholar]
- 65.Schiff ND, et al. fMRI reveals large-scale network activation in minimally conscious patients. Neurology. 2005;64:514–523. doi: 10.1212/01.WNL.0000150883.10285.44. [DOI] [PubMed] [Google Scholar]
- 66.Shirvalkar P, et al. Cognitive enhancement with central thalamic electrical stimulation. Proceedings of the National Academy of Sciences. 2006;103(45):17007–17012. doi: 10.1073/pnas.0604811103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Boly M, Coleman MR, Davis MH, Hampshire A, Bor D, Moonen G, Maquet PA, Pickard JD, Laureys S, Owen AM. When thoughts become action: an fMRI paradigm to study volitional brain activity in non-communicative brain injured patients. Neuroimage. 2007;36(3):979–92. doi: 10.1016/j.neuroimage.2007.02.047. [DOI] [PubMed] [Google Scholar]
- 68.Goldman M. Memory without feedback in a neural network. Neuron. 2009;61(4):621–34. doi: 10.1016/j.neuron.2008.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Messinger A, Lebedev MA, Kralik JD, Wise SP. Multitasking of attention and memory functions in the primate prefrontal cortex. J Neurosci. 2009;29(17):5640–53. doi: 10.1523/JNEUROSCI.3857-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Hikosaka O, Sakamoto M, Usui S. Functional properties of monkey caudate neurons. III. Activities related to expectation of target and reward. J Neurophysiol. 1989;61(4):814–32. doi: 10.1152/jn.1989.61.4.814. [DOI] [PubMed] [Google Scholar]
- 71.Fins JJ. Being conscious of their burden: severe brain injury and the two cultures challenge. Ann N Y Acad Sci. 2009;1157:131–47. doi: 10.1111/j.1749-6632.2009.04473.x. [DOI] [PubMed] [Google Scholar]
- 72.Chen L, Savio Chan C, Yung WH. Electrophysiological and behavioral effects of zolpidem in rat globus pallidus. Exp Neurol. 2004;186:212–20. doi: 10.1016/j.expneurol.2003.11.003. [DOI] [PubMed] [Google Scholar]
- 73.Akins PT, Guppy KH. Sinking skin flaps, paradoxical herniation, and external brain tamponade: a review of decompressive craniectomy management. Neurocrit Care. 2008;9(2):269–76. doi: 10.1007/s12028-007-9033-z. [DOI] [PubMed] [Google Scholar]
- 74.Munkle MC, Waldvogel HJ, Faull RL. The distribution of calbindin, calretinin and parvalbumin immunoreactivity in the human thalamus. J Chem Neuroanat. 2000;19:155–73. doi: 10.1016/s0891-0618(00)00060-0. [DOI] [PubMed] [Google Scholar]
- 75.Llinás RR, Steriade M. Bursting of thalamic neurons and states of vigilance. J Neurophysiol. 2006;95(6):3297–308. doi: 10.1152/jn.00166.2006. [DOI] [PubMed] [Google Scholar]