

Abstract
Background
Previous studies have revealed that the response of prey species to predatory risk comprised either freezing (when the prey remained immobile), or fleeing (when it ran frantically in order to remove itself from the vicinity of the predator). Other studies, however, have suggested that the prey will adjust its behavior to risk level. The present study was designed to follow the attacks of a barn owl (Tyto alba) on common spiny mice (Acomys cahirinus) and social voles (Microtus socialis guntherei), in order to reveal the correspondence between the behavior of the owl, the risk level at each phase of the owl's attack, and the defensive behavior of the rodents.
Results
Spiny mice dramatically increased the traveled distance upon the appearance of the owl, and kept moving during its attack while taking long trajectories of locomotion. Defensive response in voles dichotomized: in some voles traveled distance dropped when the owl appeared, reaching zero during its attack. In other voles, traveled distance dramatically increased once the owl appeared and further increased under its attack. These defensive responses developed by gradual tuning of normal locomotor behavior in accordance with the level of risk.
Conclusions
The phenotypic difference in defensive behavior between voles and spiny mice probably stems from their different habitats and motor capacities. Agility and running capacity, together with a relatively sheltered natural habitat, make fleeing the most appropriate response for spiny mice during owl attack. Clumsiness and relatively limited motor capacities, together with an open natural habitat, account for the dichotomy to freezing or fleeing in voles. Thus, the apparent species-specific anti-predator response in spiny mice and voles is based on species-specific normal locomotor behavior, which depends on the species-specific ecology and motor capacity, and behaviors like defensive attack or escape jump that are specific to life threat. The latter behaviors are brief, and irregularly inlaid in the ongoing locomotor behavior. Finally, our results show that in both voles and spiny mice there is a gradual transition from normal to defensive behavior in accordance with the increase in risk level.
Background
Predation generates ecological effects ranging from defensive responses of individuals [1,2], through habitat selection [3,4] and local population changes [5-7], to the ecosystem level [8,9]. This range primarily depends on the individual response of the prey when encountering a predator. However, the majority of studies on predator-prey interactions have not scrutinized real encounters. Rather, simulation of a predatory risk is used, assuming that a few stimuli and sometimes even a single stimulus that is unequivocally related to the predator will suffice to result in defensive behavior by the prey. Among these stimuli, scent materials and vocalizations are typically used to simulate predation risks [10]. Other studies employ disclosure of a prey to a predator through a physical or procedural barrier that prevents predation. For example, a caged cat was placed next to a caged rodent, with direct visual and olfactory stimulation but without physical contact [11]. Although this latter study and others on simulating threat were accompanied by adequate controls that unequivocally indicated that a predation risk had been perceived by the prey, they did not include a group with real encounters. This left open the question of when, how, and even whether the defensive patterns observed under simulation of predation also occur under actual predation. The present study was designed to scrutinize real owl-rodent encounters in order to monitor the attack pattern and the corresponding defensive responses. The behavior of rodents was studied in non-sheltered space, since rodents were attacked and captured by owls more frequently in the open ([12]).
The particular behavior of the predator may have a conspicuous impact on the response of the prey. For example, slightly moving a stuffed (taxidermised) predator dramatically increased the defensive response of prey compared with the response to a stationary stuffed predator [13]. Other components of an encounter, such as direction of an approaching threat in relation to location of a shelter [14] or access to a refuge [11] may also dictate the response of the prey. Moreover, in nature, different predators utilize a variety of hunting patterns ranging from continuous active pursuit to waiting in ambush. For example, voles are heavily predated upon by tawny owls (Strix aluco) that usually wait on a fixed perch and then swoop down on the nearby prey, and by barn owls (Tyto alba) that may hunt either from a perch or on the wing [15]. Consequently, prey species must employ different defensive strategies and develop differentiating mechanisms for the most appropriate response to the hunting strategy of a particular predator [10,13,16-22]. Indeed, rodents have been shown, for example, to display differential foraging and activity patterns following exposure to an owl or a snake [23,24]. Traditionally, the immediate behavioral responses of a prey animal facing a predator have been classified as: i) freezing – the prey remains immobile, typically crouching and sometimes also relying on camouflage, in order to avoid attention [25-28]; ii) fleeing – the prey gallops in order to remove itself from the vicinity of the predator [29,30]; and iii) fighting (or defensive threat) – the prey advances on the predator and threatens it in return in an attempt to prevent predation (this occurs only when the prey cannot avoid encounter). These defenses were previously described for various occasions and in a variety of prey species, ranging from hermit crab to deer [16,27,31-33].
Freeze and flee also characterized the response of social (Gunther's) voles (Microtus socialis guntherei) to playback of recorded owl calls [1,27]: some voles decreased activity and hid in an accessible shelter or corner (freezing) whereas other voles increased activity, running frenetically in the apparatus (fleeing). In contrast, when common spiny mice (Acomys cahirinus) were exposed to recorded owl calls [1,27], they continued their ongoing activities and ignored a shelter that was accessible in the test apparatus. Nonetheless, in both spiny mice and voles, owl calls induced increase in corticosteroids, which is a reliable indicator of stress, with higher levels recorded in voles that responded by freezing compared with those that fled [1]. The puzzling lack of behavioral response in spiny mice and the conspicuous response in voles guided us to choose these two rodent species for the present study (see Methods for more information on spiny mice and voles).
Most studies that utilize abstract threat have classified the response within one of the above three defense behaviors (freezing, fleeing, fighting), probably due to the sharp distinction of freeze and flee, allowing the prey to take only one of these responses at a given moment [34]. Other studies, however, focused on the differentiating mechanisms of defensive response, revealing freezing in the face of a distant threat but fleeing away from a closer threat [35]. This was also seen in field observations on the dynamic nature of predator-prey interaction [36], where the prey was avoiding unnecessary movements that might expose it during less critical phases of the attack, conserving energy for the more dangerous phases [34,37-40]. These and other studies demonstrate the necessity to track the behavior of the predator in order to understand the response of the prey [41].
The present study was designed to follow the attack of a freely moving live barn owl (Tyto alba) on common spiny mice and social (Gunther's) voles, in order to reveal the correspondence between the behavior of the owl and the defensive behavior of the rodents. Specifically, we set out to determine the phases in owl attack, the risk level in each of these phases, and whether the rodents adjust their behavior in accordance with changes in risk levels. Assuming that the defensive response also depends on the motor capacities and habitat structure of the prey, we compared the response in the agile and nimble rock-dwelling spiny mice with the response in the slower and clumsier burrow-dwelling voles. Finally, we examined whether access to a shelter could prevent predation.
Results
Four sequential phases in owl attack
Analysis of the videotaped owl attacks revealed three critical points: i) when the owl was first perceived by the rodent; ii) when the owl initiated its first attack on the rodent; and iii) when the rodent was caught. In accordance with these time-points, owl attack comprised four phases, as schematically illustrated in Figure 1. During the Pre-appearance phase the owl was not visible, typically being stationary in a corner or inside tree foliage thus hidden from the camera and the rodent. This phase varied from a few minutes to more than two hours, comprising behavior that mainly served as reference for the following phases. Therefore, in the present analysis the duration of this phase was arbitrarily set to the last 5 min before the first appearance of the owl. Five minutes were enough to provide a reliable basis for the behavior of the rodent and the owl before the onset of the attack, behavior that was compared with subsequent phases. The Appearance phase began when the owl moved out from its hide for the first time, and was now exposed to the camera. It was assumed that this was also the first time that the rodent noticed the presence of the owl. While this assumption was based only on the behavior of the owl regardless of the rodent's behavior, the latter matched this assumption, as shown in the results on the behavior of the rodents in each phase. It should be noted that the change in the behavior of rodents upon owl appearance was abrupt and notable (See additional files #1 and #2 with video-clips of the behavior during the four phases of owl attack in spiny mouse and vole). Considering the quiet owl flight [42,43] and the dark aviary, we presume that the remarkable response of the rodents to owl appearance was induced by various stimuli (e.g. – auditory, visual) that were generated at the time of the appearance of the owl on camera. The Attack phase started when the owl swooped down toward the rodent. However, only one spiny mouse and five voles were caught in the first attempt made by the owl. The other rodents (12 spiny mice and 8 voles) managed to evade the owl, necessitating definition of a Post-attack phase, starting when the owl landed on the ground of the rodent's enclosure in the first missed attack, and continuing until the rodent was caught. Post-attack phase comprised periods in which the owl remained chasing the rodent in its enclosure, and periods between attacks when the owl flew out from the enclosure to a perch and later initiated another attack. There was no difference in the mean time that the owl spent chasing spiny mice or voles inside the apparatus (42.5 ± 17.2 and 58.3 ± 22.6 sec, respectively). In the intervals that the owl was inside the enclosure, it could be stationary or attacking the rodent, but there was no difference in attacking periods (mean duration of owl attacking periods was 22.9 ± 7.8 sec on spiny mice and 19.5 ± 6.9 sec on voles). However, post attack intervals in which the owl re-perched outside the rodent enclosure were considerably longer in voles compared with spiny mice (287.0 ± 204.5 and 60.6 ± 30.5 sec, respectively). Obviously, the sequence of phases in owl attack was identical whether attacking a spiny mouse or a vole, since by definition the owl cannot attack before appearing or post-attack before attacking. While the duration of Pre-appearance was arbitrarily set to 5 min, and the duration of Attack was relatively short (0.6–3.0 sec), the duration of the Appearance and Post-attack greatly varied in a wide range (Figure 1).
Figure 1.
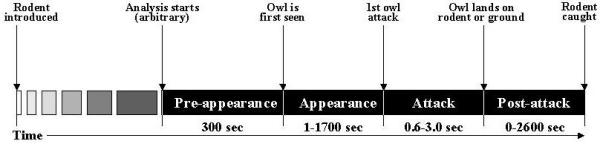
Schematic representation of four phases in barn-owl attack. Phases are described horizontally along a time axis, from left to right, with the range of the duration of each phase and the event that marks transition from phase to phase.
Differential traveling speed in the rodents during the phases of owl attack
The behavior of the rodents underwent substantial changes during the different phases of the owl attack. To illustrate the impact of the attack phases on the corresponding behavior of individual rodents, we calculated the speed of traveling in each phase and ranked these speeds together from low to high (Figure 2). As shown, the speeds of individuals in each phase aggregated in adjacent ranks, indicating different locomoting speeds during the different phases of the attack.
Figure 2.
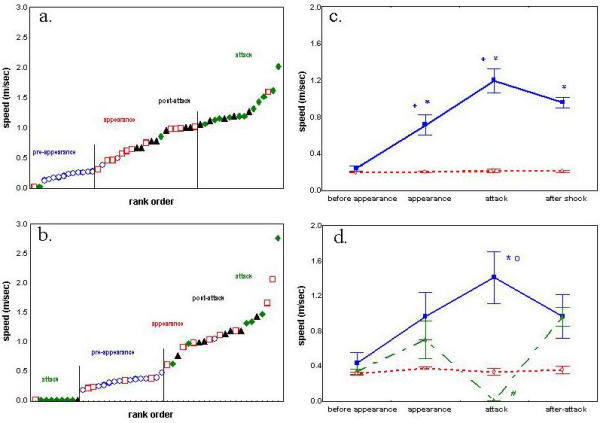
i. Rank order of the speed of locomotion of individual spiny mice in each of the four phases of barn-owl attack. The speed was calculated for each rodent by dividing the distance it traveled in each phase by the locomoting time at that phase. The speeds of all spiny mice in all four phases were then pooled and ranked from low to high and are depicted in this figure with a different symbol for each phase. As shown, individual spiny mice at the same phase (same symbols) aggregated to the same range, indicating that each phase has a typical differential speed. Specifically, spiny mice traveled at low speed during Pre-appearance (○), they increased speed during Appearance (□), and further increased it during Attack (◆). During Post-attack (▲) they attained a speed that was higher than Appearance and lower than during Attack. ii. Rank order of the speed of locomotion of individual voles for each of the four phases of barn-owl attack. Data were calculated and depicted as explained above for spiny mice. As shown, the speed during Pre-appearance (○), either increased or decreased during Appearance (□), and moved further in one of these opposite directions during Attack (◆), to zero in voles that froze (left ranks) and voles that fled (in the right third of the graph). During Post-attack (▲) most voles traveled at a relatively high speed regardless of their previous behavior (freeze or flee). iii. The mean (± SEM) in spiny mice during the four phases of owl attack. As shown, the control group (◇, dotted line) maintained the same speed throughout all phases, whereas spiny mice that were exposed to owls (■, solid line) started at the control level during Pre-appearance, increased the speed during Appearance, further increased it during Attack, and slightly decreased during Post attack. There was significant difference between control and owl groups (F1,21 = 388.6; p < 0.001), between phases (F3,36 = 43.7; p < 0.001) and significant interaction of group × phase (F3,63 = 39.4; p < 0.001). Further comparisons in Tukey test are indicated by * for difference compared with control and + for difference compared with the previous phase.iv. The mean (± SEM) speed in voles during the four phases of owl attack. Three groups are shown: i) controls, not exposed to owls (◇, dotted line); ii) voles that fled during owl attack (■, solid line); and iii) voles that displayed freeze response during owl attack (▲, dashed line). As shown, the control group maintained the same speed throughout all phases, whereas voles that were exposed to owls started at the control level during Pre-appearance, and dichotomized to freeze (speed = 0) and flee during Attack. Both these groups revealed the same speed during Post-attack. Indeed, there was significant difference between groups (F2,16 = 8.6; p = 0.002), between phases (F3,48 = 4.8; p = 0.004), and in the interaction of group × phase (F6,48 = 5.9; p < 0.001). Further Tukey comparisons are depicted by * for difference compared with control level; ○ for difference compared with the respective phase in voles that froze; # for difference between Attack phase in freeze group compared with Appearance, Attack and Post-attack in the voles that fled.
Spiny mice (Figure 2a) traveled at a relatively low speed during Pre-appearance phase (○). They increased speed during Appearance (□), and further increased it during Attack phase (◆), except for one spiny mouse that remained immobile (speed = 0). During Post-attack (▲), speed ranged at levels similar to the high ranks of the Appearance and low ranks of Attack.
Voles (Figure 2b), traveled at a relatively low speed during Pre-appearance (○). During Appearance (□), the speed measured in the different individuals was scattered across a wide range compared with its aggregated scattering during Pre-appearance. During Attack (◆), some voles remained immobile (freeze response) whereas others further increased their speed (flee response). Thus, the response during owl attack dichotomized to freeze (speed = 0) and flee (high speed). According to the response in this phase, in subsequent analyses voles were classified into 'Freeze' group, and 'Flee' group. During the Post-attack (▲), voles continued to travel at a relatively high speed except for one vole that remained stationary.
Figure 2c depicts the mean speed (± SEM) for each phase, illustrating that the change in speed measured in spiny mice significantly differed between successive phases. In voles, the increase in speed was not significant during Appearance, but significantly differed during Attack in both voles that froze and voles that fled. Post-attack speed was an intermediate of the Appearance and Attack phases in both rodent species.
The above changes in speed represent thorough changes in the behavior of the rodents, which occurred differentially during the different phases of the owl attack, as summarized in Table 1 for spiny mice and Table 2 for voles. These changes are based on three aspects of the rodents' locomotor behavior: i) the amount of activity; ii) the temporal organization of activity; and iii) the spatial distribution of activity (see 'Methods'). As revealed by these data, in terms of activity the behavior of spiny mice during Pre-appearance of the owl resembled that of control spiny mice that were not exposed to the owl, indicating that Pre-appearance behavior was not affected by the presence of the unseen owl in the aviary. Traveled distance dramatically increased upon Appearance of the owl and spiny mice kept moving continuously during the Attack phase. Trip-length dramatically increased in the Post-attack phase with a concomitant increase in inter-stop distance, indicating that spiny mice were taking longer trajectories. Spiny mice typically spent about 40% of the time along the perimeter of the enclosure and the counterpart of 60% of the time at the center, except for the short duration of the Attack when they were mainly in the center and only 16% of the time along the perimeter.
Table 1.
Changes in locomotor behavior of spiny mice during the four phases of the barn-owl attack, compared with controls that were not exposed to the owl. Data (means ± SEM) are arranged according to activity, temporal structure, and spatial distribution (see 'Methods'). Results of two-way ANOVA are depicted for Groups (Control vs. Owl), Phases (of owl attack) and the interaction Groups × Phases. Distance was normalized per minute, and locomoting time was calculated as percent of the respective phase to bypass the different duration of each phase in each individual. [Statistical comparisons were not carried out on activity parameters since they are linked with speed, as animals were divided according to their speed].
| Behavior | Phase | Control | Owl | Groups F1,21; P | Phases F3,63; P | Interaction F3,63; P |
| ACTIVITY | ||||||
| Distance (m./min.) | Pre-appearance | 5.97 ± 0.25 | 4.40 ± 0.35 | |||
| Appearance | 4.45 ± 0.64 | 14.15 ± 4.22 | ||||
| Attack | 4.43 ± 0.98 | 57.74 ± 8.15 | ||||
| Post-attack | 3.11 ± 0.83 | 36.26 ± 7.33 | ||||
| Locomoting time (% of phase) | Pre-appearance | 51.99 ± 1.42 | 34.85 ± 3.65 | |||
| Appearance | 37.13 ± 4.11 | 31.35 ± 8.72 | ||||
| Attack | 35.86 ± 6.73 | 74.23 ± 8.04 | ||||
| Post-attack | 26.12 ± 6.25 | 58.74 ± 10.23 | ||||
| TEMPORAL STRUCTURE | ||||||
| Trip length (m.) | Pre-appearance | 8.72 ± 2.67 | 4.27 ± 1.56 | 3.06; ns | 3.88; ns | 7.94; <0.001 |
| Appearance | 7.81 ± 2.01 | 3.67 ± 0.87 | ||||
| Attack | 5.45 ± 0.98 | 1.10 ± 0.17 | ||||
| Post-attack | 3.94 ± 0.65 | 8.80 ± 2.01 | ||||
| # Stops per trip | Pre-appearance | 8.61 ± 2.61 | 4.62 ± 1.31 | 16.84; <0.001 | 3.85; ns | 1.52; ns |
| Appearance | 9.47 ± 2.37 | 2.99 ± 0.88 | ||||
| Attack | 5.82 ± 0.79 | 0.23 ± 0.12 | ||||
| Post-attack | 5.73 ± 1.04 | 3.52 ± 0.92 | ||||
| Inter-stop distance (m.) | Pre-appearance | 1.01 ± 0.05 | 0.87 ± 0.05 | 7.83; ns | 3.47; ns | 10.33; <0.001 |
| Appearance | 0.83 ± 0.09 | 1.30 ± 0.39 | ||||
| Attack | 0.97 ± 0.16 | 1.10 ± 0.17 | ||||
| Post-attack | 0.78 ± 0.13 | 2.94 ± 0.54 | ||||
| SPATIAL DISTRIBUTION | ||||||
| Time at perimeter (% of phase) | Pre-appearance | 33.65 ± 2.64 | 39.30 ± 4.20 | 3.94; ns | 1.98; ns | 7.15; <0.001 |
| Appearance | 43.27 ± 4.74 | 41.79 ± 10.62 | ||||
| Attack | 53.49 ± 6.57 | 16.92 ± 9.57 | ||||
| Post-attack | 51.92 ± 8.12 | 37.15 ± 8.77 | ||||
Table 2.
Changes in locomotor behavior of voles during the four phases of the barn-owl attack, compared with controls that were not exposed to the owl. Voles under owl attack were classified into 'freeze' and 'flee' groups, with the former minimizing and the latter increasing their speed during owl attack.Data (means ± SEM) are arranged according to activity, temporal structure, and spatial distribution (see 'Methods'). The results of two-way ANOVA are depicted for Groups (Control vs. Owl), Phases (the four phases of owl attack) and the interaction Groups × Phases. Distance was normalized per minute, and locomoting time was calculated as percent of the respective phase to bypass the different duration of each phase in each individual. [Statistical comparisons were not carried out on activity parameters since they are linked with speed, as animals were divided according to their speed].
| Behavior | Phase | Control | Freeze | Flee | Groups F2,16; P | Phases F3,48; P | Interaction F6,48; P |
| ACTIVITY | |||||||
| Distance (m./min.) | Pre-appearance | 6.01 ± 0.50 | 5.21 ± 1.09 | 3.29 ± 0.85 | |||
| Appearance | 5.15 ± 0.68 | 0.86 ± 0.26 | 2.90 ± 1.78 | ||||
| Attack | 4.86 ± 0.89 | 0.0 ± 0.0 | 59.78 ± 10.62 | ||||
| Post-attack | 4.37 ± 0.89 | 4.11 ± 3.04 | 21.60 ± 9.05 | ||||
| Locomoting time (% of phase) | Pre-appearance | 33.47 ± 2.72 | 25.02 ± 4.30 | 15.14 ± 4.54 | |||
| Appearance | 23.02 ± 3.00 | 3.55 ± 2.13 | 5.85 ± 2.92 | ||||
| Attack | 22.40 ± 3.86 | 0.0 ± 0.0 | 100.0 ± 0.0 | ||||
| Post-attack | 19.63 ± 3.96 | 6.47 ± 4.33 | 29.18 ± 12.61 | ||||
| TEMPORAL STRUCTURE | |||||||
| Trip length (m.) | Pre-appearance | 8.53 ± 2.26 | 6.28 ± 2.11 | 5.57 ± 1.80 | 3.14; ns | 6.74; <0.001 | 2.61; ns |
| Appearance | 6.28 ± 0.90 | 2.18 ± 0.93 | 4.70 ± 1.44 | ||||
| Attack | 7.02 ± 2.17 | 0.0 ± 0.0 | 1.20 ± 0.30 | ||||
| Post-attack | 6.41 ± 2.28 | 6.43 ± 2.17 | 5.30 ± 1.84 | ||||
| # Stops per trip | Pre-appearance | 12.24 ± 3.39 | 8.30 ± 2.35 | 6.97 ± 0.85 | 5.99; ns | 8.23; <0.001 | 3.16; 0.01 |
| Appearance | 9.15 ± 1.64 | 4.27 ± 1.70 | 5.42 ± 2.19 | ||||
| Attack | 10.13 ± 3.30 | 1.00 ± 0.0 | 0.0 ± 0.0 | ||||
| Post-attack | 9.26 ± 2.98 | 3.10 ± 0.24 | 3.17 ± 0.58 | ||||
| Inter-stop distance (m.) | Pre-appearance | 0.70 ± 0.03 | 0.77 ± 0.10 | 0.81 ± 0.29 | 2.43; ns | 10.16; <0.001 | 6.70; <0.001 |
| Appearance | 0.72 ± 0.05 | 0.45 ± 0.09 | 1.77 ± 1.10 | ||||
| Attack | 0.65 ± 0.07 | 0.0 ± 0.0 | 1.20 ± 0.30 | ||||
| Post-attack | 0.61 ± 0.08 | 2.00 ± 0.53 | 1.42 ± 0.49 | ||||
| SPATIAL DISTRIBUTION | |||||||
| Time at perimeter (% of phase) | Pre-appearance | 66.67 ± 4.65 | 59.00 ± 8.03 | 62.84 ± 13.31 | 0.696; ns | 5.07; 0.003 | 2.54; ns |
| Appearance | 67.27 ± 5.43 | 82.99 ± 11.10 | 87.29 ± 7.17 | ||||
| Attack | 69.67 ± 6.75 | 85.71 ± 14.29 | 33.33 ± 21.08 | ||||
| Post-attack | 71.84 ± 5.91 | 94.63 ± 2.91 | 85.21 ± 8.72 | ||||
In voles that displayed freeze response, traveled distance dropped when the owl appeared, reaching zero during owl Attack, where locomoting time, trip-length and inter-stop distance were also zero, and there was only one stop (the incidence of freezing). Once the owl appeared, these voles spent more than 80% of the time along the perimeter, reaching more than 95% of the time during the Post-attack. In voles that displayed flee response, traveled distance dramatically increased during Attack and Post-attack, with these voles locomoting throughout the Attack phase. Inter-stop distance increased once the owl appeared, indicating that fleeing voles took longer trajectories between successive stops. Like the freezing voles, fleeing voles also clung to the walls once the owl appeared and during the Post-attack phase. However, during the Attack phase they abandoned the walls, tending to run frantically mainly in the center (like spiny mice), spending only 33% of the time along the perimeter.
Defensive attacks and escape jumps
Defensive attacks were rare, observed in only one spiny mouse and four voles, with an overall incidence of two in spiny mice (both by the same spiny mouse) and an incidence of six in four voles (1–2 per individual; See additional file #3 with video clip of defensive attack). Escape jumps were not observed during Pre-appearance and Attack in either voles or spiny mice. In spiny mice, escape jumps were observed in low frequencies during Appearance (3.25 ± 1.35; mean incidence ± SEM), but were prevalent in Post-attack (11.31 ± 5.00; mean incidence ± SEM). In voles, escape jumps were rare in Appearance (0.62 ± 0.40; mean incidence ± SEM), and frequent during Post-attack (8.78 ± 2.84; mean incidence ± SEM; see additional file #4 with video clip of escape jump).
The effect of access to a shelter
In order to assess the effect of the lack of shelter in the above experiments, six voles were tested under the same procedure used in the above experiments, but this time with a shelter available inside the test apparatus. Each of these voles was placed inside the shelter at the beginning of the observation (see 'Methods'), and thus could actually avoid the owl. However, as shown in Table 3, access to the shelter delayed but did not prevent predation. The voles soon left the shelter and traveled in the open area, even after noticing or encountering the owl, as shown in Figure 3 for two representative voles. The effect of access to a shelter was therefore in adding an initial response of running into the shelter, but once in the shelter voles either remained motionless inside the shelter or resumed locomotion in the outside.
Table 3.
A comparison of the distance traveled, speed, and time until being caught by the owl with/without shelter. The distance (m/min) describes the metric distance traveled by the rodent in each phase, comprising locomoting and non-locomoting periods. However, since the duration of phases was different for each rodent, data are "normalized" per minute of observation. This measure is not interchangeable with speed (m/sec) that refers only to the locomoting periods, and was calculated by dividing the distance traveled with the duration of traveling that distance (excluding all non-locomoting periods).
| Behavior | Phase of owl attack | Without shelter | With shelter |
| Distance traveled (m./min.) | Pre-appearance | 4.32 ± 1.6 | 4.16 ± 1.62 |
| Appearance | 1.80 ± 1.7 | 0.36 ± 0.68 | |
| Attack | 27.6 ± 6.0 | 52.3 ± 6.8 | |
| Post-attack | 15.04 ± 4.24 | 5.6 ± 2.07 | |
| Speed (m/sec) | Pre-appearance | 0.37 ± 0.46 | 0.54 ± 0.01 |
| Appearance | 0.82 ± 0.78 | 0.54 ± 0.05 | |
| Attack | 0.65 ± 0.93 | 0.83 ± 0.86 | |
| Post-attack | 0.96 ± 0.66 | 0.58 ± 0.02 | |
| Time until being caught (min) | 38.3 ± 6.8 | 83 ± 28.7 | |
Figure 3.
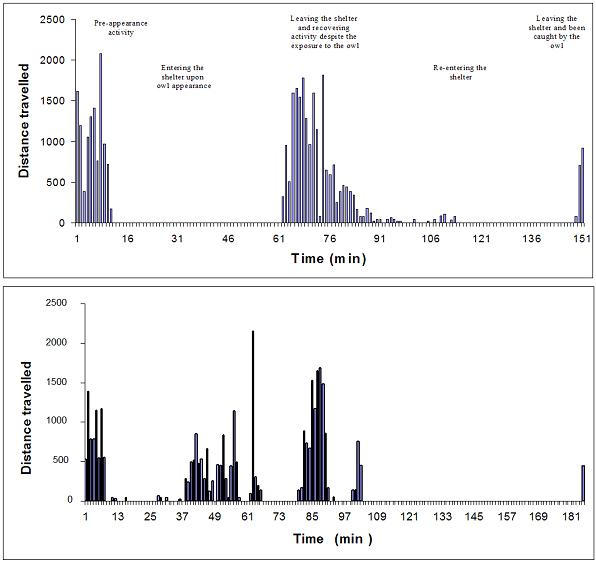
Behavior with access to shelter. Each graph represents the activity of one vole that was tested with a shelter. The traveled distance (in cm.) is shown for each minute of observation. In the top graph, activity during Pre-appearance (left portion of the graph) ceased to zero when the owl appeared and the vole entered the shelter (11 min). At 63 min the vole left the shelter for another period of activity, matching the level of the Pre-appearance period, despite the exposure to the owl. The vole then re-entered the shelter, taking only frequent short-distance trips (risk assessments) to the outside (90–110 min), followed by inactivity (110–148 min). The vole then again left the shelter (110) and was then caught during the first attack by the owl (151 min after beginning of observation). A second vole with the same behavioral pattern is shown in the bottom graph. Its initial activity (1–13 min) ceased when the owl appeared and the voles stayed in the shelter with few stretch attempts to the outside (13–37 min). It then resumed activity in the outside (37–101 min), followed by a long pause in activity (101–184 min). The vole then left the shelter and was caught by the owl.
Discussion
The present study shows that barn-owl attack comprises four sequential phases during which the owl emerges from a hide to a perch, then swoops down on the rodent, and finally, if the rodent evades the first attack, re-attacks until a successful catch is gained. Both spiny mice and voles adjust their behavior in accordance with the differential risk level of each phase. However, while spiny mice mostly flee from the attacking owl, the response in voles dichotomizes to freeze and flee. In the following discussion we first describe how defensive behavior developed from tuning normal behavior according to the increased threat. We then use the continuum between normal and defensive locomotor behavior to explain previous reports on a lack of behavioral response in spiny mice that were exposed to owl calls. Finally, we discuss how the behavioral response is affected by access to a shelter.
When encountering a predator, the prey is required to discern between lethal and less dangerous phases, and react according to the risk level embedded in each attack phase. When danger is low, unnecessary movement (response) may expose the prey and consume energy that may be essential for escape efforts during more critical moments [34,37-39]. Indeed, the present results demonstrate that at different phases of owl attack, spiny mice and voles change their level of activity, the structure of the trips they take in the environment, and the spatial distribution of their locomotion. At first, risk level was low and the owl was not visible, perching motionless in hidden parts of the aviary (Pre-appearance phase). Risk level increased once the owl moved forward to a visible roost, exposing itself to the rodent (Appearance). Risk level then peaked when from this or another perch the owl swept down on the rodent (Attack). Risk level slightly declined if the owl failed to catch the rodent in the first attack and executed repeated attacks until a successful catch (Post-attack). In accordance with these different risk levels, the behavior of the rodents differed, resembling control levels during Pre-appearance and peaked during Attack.
During Pre-appearance, behavior of the rodents was similar to behavior of control rodents that were not exposed to an owl, indicating that the rodents were probably not aware of the presence of a hidden owl in the aviary. Our previous studies with tamed wild rodents that were introduced into an empty arena (e.g. – [27,44,45]) revealed that they were relatively relaxed, and after a few minutes of exploration tended to rest in one place for extended periods, except for spiny mice that kept continuously on the move. Therefore, we assume that the rodents were not stressed by being introduced to the empty arena. This was further supported by the finding that transferring voles and spiny mice to a small open field does not induce increase in corticosteriods [1], which would indicate stress. Thus, we consider the behavior of the rodents during Pre-appearance phase as normal, as indicated by controls that were not exposed to an owl, and as compared with their different behavior once the owl appeared.
The more risky phase of Appearance involved a decrease in activity, probably in order not to expose the location of the rodent to the owl. Indeed, prey species are capable of recognizing and reacting to sound, odor, or silhouette of predators [46-48], and intensifying the defensive response once the threat (either real predator of predator taxidermy) moves [23]. In spiny mice, periods of locomotion during Appearance comprised shorter trips executed at higher speed compared with Pre-appearance. The behavior of the voles during Appearance started to dichotomize, with some of them decreasing activity, trip length, and travel speed ('freeze' response) and others decreasing activity and trip length but traveling at a higher speed ('flee' response). This dichotomy became more obvious when predatory risk peaked during Attack: voles that froze remained immobile, probably presuming that the owl had not identified their location, whereas voles that fled ran frantically, trying to evade the attacking owl. Attack phase was relatively short, followed by a longer Post-attack phase, when the owl might leave the rodent enclosure, fly to a perch, and then re-attack. Overall risk level during Post-attack was therefore higher than in Appearance but lower than in Attack. In accordance, the rodents adopted in this phase a behavior that was intermediate between that seen in Appearance and Attack. In certain bouts of Post-attack, the owl would remain nearby the rodent and chase it. In this case, life threat was close and immediate, and rodents not only froze or fled but also sporadically executed jumps either to evade the owl (escape jumps; see additional file #3) or attack it (defensive attack; see additional file #4) [11,49]. It should be noted, however, that in the present study rodents were constrained within the enclosure and their movement might attract the owl to remain in the enclosure and chase them, whereas in the wild, barn owls would probably be unsuccessful in getting an individual prey if they miss it in the first swoop. To our point, nonetheless, the Post-attack phase reflects intermediate level between Appearance and Attack, when considered only from the perspective of risk level as explained above. Thus, in real encounters with owls, rodents have a repertoire of defensive behaviors from which they choose their response according to the level of risk, as suggested in previous studies [34,37-39].
Escape jumps, defensive attack, freezing and frantic running, are behaviors that were not seen in the control groups or during the Pre-appearance phase in the behavior of rodents that were exposed to owls. These behaviors are therefore specific to immediate life threat (Appearance, Attack and Post-attack phases), yet they are typically brief, irregularly inlaid in the ongoing locomotor behavior. We suggest here that locomotor behavior under owl attack is an intensified form of ordinary locomotor behavior of spiny mice and voles that are not exposed to owls. Indeed, spiny mice that were introduced into 100 cm × 100 cm arena in a quiet isolated room with no threat or obstruction (Eilam, unpublished results) tended to travel with frequent changes in the direction of progression, as did the control spiny mice in the present study. Spiny mice under owl attack show the same locomotor behavior, but with more frequent changes in the direction of progression due to their higher traveling speed. Voles exploring a 100 cm × 100 cm arena in a quiet isolated room with no threat or obstruction ([45]; Eilam, unpublished results) traveled mainly along the walls of the arena, crossing the center in relatively straight segments that extended from wall to wall, as did control voles in the present study and voles before the appearance of the owl. This form of locomotor behavior was intensified in voles that fled while frequently crossing the center from wall to wall, and was moderated in voles that froze while clinging to the walls. Thus, it is suggested that locomotion under owl attack developed by gradual tuning of normal locomotor behavior in accordance with the level of risk, with incorporation of responses that were specific to life threat. In consequence, a species-specific anti-predator response may be regarded as consisting of: i) species-specific locomotor behavior which depends on the species-specific morphology, ecology and motor capacity; and ii) more general forms of defensive behaviors (defensive jump, extended duration of freezing, escape jump) that are irregularly ingrained into the species-specific locomotor behavior.
Field studies with spiny mice revealed that, like other rodents, they decrease activity and foraging in open spaces under moonlit compared with dark nights [6]. However, laboratory studies [1,27] revealed that unlike other rodent species, spiny mice do not alter behavior when exposed to playback of recorded owl calls. This lack of behavioral response was puzzling, considering that the same spiny mice that did not change behavior had increased levels of cortisol [1], which is a reliable indicator of stress [50-52]. In other words, while owl calls are perceived as a threat by spiny mice, they do not necessarily require a behavioral change. It was suggested that this lack of behavioral defensive response is due to the small size of the test apparatus, which could well have offered the perception of shelter, as do the spaces and crevices under and between boulders where spiny mice prefer to forage, being protected from aerial predators [1]. In the present study, behavioral response in spiny mice was discerned as soon as predation risk increased during Appearance (see additional file #1). This marked response supports the surmise that the small-sized apparatus (40 cm × 40 cm) used in previous studies [1,27] accounts for the lack of behavioral response in spiny mice. In light of the above suggestion on the continuity between normal and defensive behavior, it is probable that spiny mice perceived playback of recorded owl calls as a threat, as indicated by the increased corticosteroid level. However, deviation from their ongoing locomotor behavior was minimal under this sole stimulus, whereas exposure to the attacking owl in the present study induced marked intensification in their locomotor behavior. It should be noted that when spiny mice and voles were exposed to owl calls, they were also provided with access to a shelter ('artificial' burrow) [1,27]. However, while some of the voles hid in the shelter, all the spiny mice ignored the burrow and kept moving in the open space of the apparatus [1,27], attesting to their uniqueness among murid species in not building a 'nest' or permanent home shelter.
A major difference between the locomotor response of spiny mice and voles is that spiny mice mostly flee whereas voles dichotomize to those that freeze and those that flee. These differences, however, match the motor capacities and habitat structure of each species. Spiny mice are agile and fast runners as compared with voles [53,54]. In addition, spiny mice live and forage in crevices and spaces between and under boulders, spending little time in the open [55]. Thus, agility, fast running, and a habitat that is relatively protected from aerial predators, make fleeing the more appropriate response for a spiny mouse during owl attack. This follows a previous experiment in which spiny mice increased activity and fled when exposed to a stuffed (taxidermised) predator [56]. Conversely, voles are burrow-dwellers that forage for vegetation and seeds in relatively open spaces, where they are heavily predated upon by both terrestrial and aerial predators [57]. The motor capacity of voles is basic and they rarely gallop [53], making fleeing less effective for a vole under owl attack. Freezing, on the other hand, may have several advantages for voles, as follows. Owls initially detect prey by means of their sensitive auditory nervous system [58]. The noise generated by moving may therefore expose the location of the rodent while freezing prevents this. Having aurally located a prey, the owl then pinpoints its location also by means of vision. The brown fur of the vole blends in with the color of the heavy soil of their habitat, making it hard to distinguish them once motionless. Finally, voles have a relatively small home range, where they travel in the vicinity of their burrows in familiar and relatively fixed routes [57]. These features of voles are tailored in their response to threat as revealed in the field, where they initially freeze and then bolt for a burrow hole [59] and in the present observations on voles with access to a shelter. Without shelter, voles equally freeze or flee [1,16,27]. In the present study, the incidence of freeze and flee was seven and six, respectively, in response to the first catching attempt by the owl, reflecting equal use of both these defensive strategies. Linkage between a specific defensive response, the motor capacities of the prey and its habitat structure, was found in comparing two deer species [60]. The white-tailed deer (Odocoileus virginianus), which inhabits forest and is a fast runner, tends to flee when encountering coyotes (Canis latrans), whereas the mule deer (Odocoileus hemionus), which lives in relatively open spaces and is a moderate runner, tends to freeze or flee. Moreover, upon detecting a predator, white-tailed deer freeze when they are in dense vegetation, but flee when they are in sparse vegetation [33]. Thus, as in spiny mice and voles, better motor capacities and a sheltered habitat account for fleeing, while limited speed and open habitat account for the dichotomy of freeze and flee.
When exposed to predatory risk, prey species extend the time they spend in burrows or among boulders and dense vegetation, shifting their activity to more secure spaces [34,59,61,62]. It has been shown that when threatened, prey species favor entering an accessible refuge [11,14] where they may stay for extensive periods [63]. This extended interruption of vital activities such as foraging and mating has led to the assumption that re-emergence from the shelter depends on the physical condition of the prey, with a shorter latency in subordinates. In the present study, providing satiated voles with access to a shelter delayed but did not prevent predation. These voles remained in the shelter for periods ranging from two minutes to more than two hours, but ultimately left the shelter and were caught by the owl. This was especially remarkable in voles that had managed to evade the owl and hide in the shelter where they were protected from the owl, but nonetheless left the shelter later. A possible explanation is that the defensive response declined once the owl had moved away and perched motionless, thus the voles might no longer notice the owl and resume activity outside the shelter (Figure 3). This pattern was seen in field observations on gerbils, revealing that they decreased activity when a tamed owl flew above them, but once flight had ceased the gerbils reverted to the initial level of activity [23]. Access to the shelter, however, had a unifying effect [64] on the initial response to the presence of the owl, with all six voles first performing a short freeze after which they fled to the shelter, as also observed in the field [59]. In contrast, there was no specific order or direction in the response of voles tested without shelter, and they fled in different directions or froze in various places.
Lima (2002) criticized studies of predator-prey interactions that consider the predator as an abstract source of threat rather than a participant in a larger behavioral interaction. This was exemplified in the radical change shown in the behavior of a prey simply by moving a stuffed (taxidermized) predator compared with the response to a stationary stuffed predator [13]. By illustrating how the behavior of prey coincides with the differential risk imposed at different phases of predator's attack, the present study follows the suggestion to put the predator factor back into studies of behavioral predator-prey interaction [41].
Conclusions
Owl attack comprises several phases, each with a different level of risk. Spiny mice and voles adjust their locomotion in accordance with the risk level of each phase of the attack. The gradual adjustment in locomotor behavior indicates that defensive behavior emerges by incremental tuning of the behavior that was ongoing before the exposure to the owl, with sporadic episodes of behaviors that are specific to life threat (defensive attack or escape jumps). In consequence, defensive locomotor behavior, like normal locomotor behavior, depends on motor capacities and habitat structure. Indeed, agility and running capacity, together with sheltered habitat that is available for spiny mice in the wild, make fleeing their appropriate response during owl attack. In contrast, clumsiness and relatively limited motor capacities, together with relatively open natural habitat, account for the dichotomy to freezing or fleeing in voles.
Methods
Study animals
Spiny mice
The common spiny mouse (Acomys cahirinus) weighs 38–44 g and is 11 cm long, plus a 10-cm tail. Spiny mice are an exceptional genus among murid rodents (Muridae) in being precocial and not building a nest. They differ from rats and mice in many respects (see [65] for review); noteworthy are differences in depth perception [66,67], distance perception [68], exploration [56,69] and excitability [70]. The common spiny mouse is a nocturnal omnivore [55] that occurs in rocky environments where it nimbly moves, foraging in the crevices between and under rocks and boulders [71-73] where the complex habitat structure provides shelter and escape from avian predators. Nonetheless, the spines on its rump, a histological mechanism for tail-loss [74], and relative resistance to snake venom (Weissenberg et al. 1997, Bouskila A., Dayan T., and Weissenberg S., unpublished data) attest to the predation pressure on this species. When placed in an open laboratory arena, spiny mice tend to progress continuously with frequent and irregular changes in the direction of progression (Eilam, unpublished data).
Voles
The social (Guenther's) vole (Microtus socialis guentheri) weighs 37–50 g and is 11 cm long, plus a 2-cm tail. It is a burrow-dwelling rodent that feeds on seeds and green vegetation. Social voles inhabit areas with green vegetation and grain fields where they dig system of burrows to which they escape by traveling along relatively fixed routes. High year-round fecundity and early maturation results in large vole populations that are heavily predated upon by owls and other predators. For example, voles comprise 40–70% (sometimes over 90%) of the diet of barn owls and tawny owls (Strix aluco) [15,57,58,75,76]. When placed in an open laboratory arena, social voles tend to progress along the walls or quickly cross the center from wall to wall. Their behavior in the arena is highly organized as a set of roundtrips that start and end in the same place, where they stop for extended periods [45].
We obtained 19 voles and 13 spiny mice bred in captivity in colonies at the research zoo of Tel-Aviv University. Another group of 11 voles and a group of 11 spiny mice were used as controls, and these were returned to their colonies after testing. In studying behavior of rodents that were born in captivity we relied on our past studies with wild captive rodents [1,27,44,45,53,77-80] where we found that the behavior of wild rodents that were born in captivity appeared advantageous over the use of rodents that were caught in the wild. While the former were relatively tame, the latter were frantic and did not seem to display any sensible behavior that could reflect their behavior in the wild.
Several weeks before testing, the rodents of each species were housed in groups of 5–10, in metal cages measuring 40 cm × 70 cm and 25 cm, located outdoors in the zoo yard under natural (uncontrolled) temperature and light conditions. Overturned ceramic pots and wooden boxes were placed each cage to provide shelter. Seeds and diced fresh vegetables were provided ad lib. Spiny mice were also provided with live fly larvae. Based on years of experience in maintaining colonies of voles in our zoo, provision of water is unnecessary when sufficient fresh vegetables are provided.
Barn owls
The barn owl (Tyto alba) weighs 250–315 gr and is 31.5–36 cm long with 28 cm span of elliptic wing. Dorsal feathers are golden-yellow and the ventral feathers are white with scattered brown dots. Barn owls are efficient raptors that feed mainly on rodents. The initial detection of prey location relies on hearing the sounds generated by prey movement, and is followed by visually pinpointing the prey with sharp night-vision. Barn owls then swoop down on the prey from a perch or on the wing, catching it with their spiked talons and killing it quickly. A colony of barn owls is kept in the research zoo of Tel-Aviv University, and is fed with freshly killed (from time to time also live) chicks and mice, obtained from surplus stock of the University animal quarters and from chicken-incubators. Thus, these captive barn owls were accustomed to preying on live rodents. Because of the high fecundity of voles and spiny mice (early maturation, short weaning period, frequent all-year-round breeding in captivity), surplus of these rodents is also used to feed the owls. One adult male barn owl was selected as the predator in the present experiment. We chose this specific barn owl since preliminary observations on the barn-owl aviary revealed that it had relatively short latency in attacking live prey. Throughout the experimental period the owl was provided with one rodent per night, which is the regular feeding schedule at the zoo.
Ethical consideration
This study was carried out under the regulations and approval of the institutional committee for animal experimentation (permit #L-02-40). The design carefully followed the Guidelines for the treatment of animals in behavioural research and teaching, ANIMAL BEHAVIOUR, 2001, 61, 271–275, with special emphasis on the guidelines for staged encounters as outlined by [81].
Apparatus
Observations took place in a 5 × 5 × 4 m aviary with roosts and a small tree, where the barn owl was housed alone several weeks before testing. A 1 × 1 m plywood enclosure with 60 cm high walls, open top, and a transparent glass front was placed in the center of the aviary, used as the rodent's enclosure. The floor of the enclosure was covered with a thin layer of brown-yellowish sand, and the walls were painted dark blue (except for the transparent front). In this setting, all the roosts in the aviary were visible from the floor of the rodent's enclosure, subject to the level of light. In other words, a rodent in the enclosure could see the owl unless it was hidden in the tree, was on the floor of the aviary, or when the level of light was low. Two infrared-sensitive video cameras (Sony TRV23E; Ikegami ICD47E) that allow vivid shots in complete dark were installed in the aviary, one encompassing the entire cage, providing information on the location of the owl, and the other focusing on the enclosure, providing a close view of the rodent. Two IR lights (Tracksys, IR LED Illuminator; UK) that emit light in a range invisible to owls and rodents, followed the directions of the cameras. A video mixer (Panasonic WJ-AVE5) was fed with the video signals of both cameras, providing a single composed picture, showing the attacking owl and the defending rodent in the same frame (= same time), thus enabling us to monitor the behavior of the owl and the immediate response of the rodent from moment to moment. A time-code generator (Telcom Research, Canada; T-5010+T800) that marks every video frame (25 frames/sec) was added to the composed video signal, which was stored in a VHS format VCR (LG C20W).
Procedure
Observations (one per night) took place between 11 pm and 2 am, since in these hours the latency for owl attack is relatively short. At these hours, the apparatus was entirely dark and light level was 0.0425 Lux (measured with Profisix Sbc, Gossen). At the beginning of the observation, an experimenter switched on the equipment, released a rodent into the enclosure, and left the aviary area. The next morning, the videocassette (180 min) was collected and scanned for the owl-rodent encounter, which was then analyzed frame-by-frame. Control rodents underwent the same procedure but without the owl (empty aviary). Thirteen spiny mice and thirteen voles were tested with no shelter available in the enclosure. Another group of six voles was tested with an owl in the aviary and a shelter in the rodent's enclosure. The shelter was an overturned ceramic pot that was taken from the voles' cage, where it was also used as shelter. Thus, the voles were familiar with the odor and the functional use of this shelter. Spiny mice were not tested with shelter since previous studies [1,27] had revealed that they ignored the shelter and kept moving in the open apparatus despite predatory threat.
Behavioral analysis
The owl-rodent encounters were each analyzed during playback of the videocassettes, using a custom designed software. This allowed us to score the behavior of the two experimental animals, providing the time the rodents spent locomoting or stationary, the distance they traveled, the places in which they stopped, and the incidence and duration of stopping in each place [82]. Data from the owl were used to identify its location and the duration it spent in each roost. Rodent data were used to reconstruct their activity, its temporal structure, and its spatial distribution. For these, the following parameters were scored.
Activity
Distance traveled
Overall distance (m.) traveled by a rodent during the observation.
Locomoting time
Overall duration (sec.) of locomoting periods in the observation. Since the periods of locomotion greatly varied between individuals, locomoting time was also described as percent of the entire period (= locomoting + non-locomoting intervals).
Speed
The average speed at which the rodent was traveling was calculated by dividing the distance traveled by locomoting time (m/sec).
Temporal organization
The above measures describe the amount of rodent activity, but not how this activity was temporally organized. Our previous studies of rodents that were introduced into an empty arena as in the present study, demonstrated that their locomotor behavior was organized in relation to a home base, which is the place where a rodent spends the longest cumulative non-locomoting periods [44,45]. From the home base the rodent takes round trips in the environment [44]. Rodents adjust their activity to the space available for locomotion by changing the length of round trips and scaling the distance between consecutive stops, but preserve the average number of stops in a round trip [45,80]. Using these characteristics of the temporal (sequential) organization of locomotor behavior we measured in this study the following parameters:
Round-trip length
The average metric distance traveled in a round-trip, calculated by dividing the total traveled distance by the number of trips.
Stops per round-trip
The average number of stops between two successive visits to the home base, calculated by dividing the total number of stops by the number of round trips.
Inter-stops distance
The average metric distance traveled between two consecutive stops, calculated by dividing the total traveled distance by the total number of stops.
Spatial distribution
The above measures describe the amount of activity and its temporal structure but not where it occurred. For example, the same amount of activity and temporal structure could be executed along the perimeter of the arena, or only in the center of the arena. For this, a representation of the spatial distribution is required. In this study, we only compared the amount of activity that took place along the perimeter with that occurring in the center of the arena by measuring the Time spent along perimeter of the enclosure as percent of the overall time of observation (= time along perimeter + time in the center). To be in the perimeter, the entire trunk of the vole had to be within 12 cm from the walls of the enclosure.
Defensive attack
This was scored when the rodent leapt toward the owl, with or without attempt to bite it.
Escape Jump
This was scored when the rodent leapt, either vertically or away from the owl (but not toward the owl as in defensive attack), as if trying to evade the attacking owl or move away from it. However, escape jumps also seemed to occur regardless of the direct behavior of the owl. For example, leaping occurred when the rodent bumped into a wall of the enclosure while escaping the owl.
Statistics
Unless otherwise indicated we analyzed raw data using two-way ANOVA, with follow-up comparisons of individual group differences using Tukey HSD for unequal N (Spjotvoll-Stoline test). The two rodent species, or control vs. test groups, were the between group factor. Phases in owl attack were the within group factor, and the repetitions were the individual rodents. When data deviated significantly from a normal distribution (Kolmogorof-Smirnov test for normality) we carried out an ANOVA on ln [X+0.01]. Data calculated as proportions were transformed to the arcsine of the square-root-transformed raw data. A Bonferroni correction was applied to set alpha level to 0.007, due to possible dependence of the data on locomotor behavior. Otherwise, alpha level was set to 0.05.
Authors' contributions
Both authors equally contributed in all phases of this study.
Additional files
Each clip comprises two simultaneous pictures. One of the entire cage and a second (left bottom quarter of the frame) is a close view on the rodent enclosure (also seen in the picture of the entire cage). Numerals above the rodent enclosure describe the time from the onset of observation (hours, minutes, seconds, and frames; 25 frames/sec). It should be noted that the clips were videotaped in darkness (0.0425 Lux measured with Profisix Sbc, Gossen) under infrared light that is not visible to both the owl and the rodent. Therefore, the glaring eyes of the rodents were not visible to the owl as they are to the camera lens.
Supplementary Material
Spiny mouse & barn owl. The spiny mouse is moving around until the owl appears 11 minutes after the spiny mouse was introduced (00:11:02:00). It then moves to the corner and remains immobile for a while, resuming locomotion in the center with relatively long pauses, until the owl launches its attack 21 minutes after the spiny mice was introduced (00:21:41:00. The spiny mouse responds in fleeing and orienting to the owl, until being caught by the owl (00:22:07:00).
Vole & barn owl. At first the owl is hidden and the vole moving freely. Once the owl appears and progresses to exposed roost (00:07:31:00), the vole leaps and runs to a corner, where it remains freezing until being caught by the owl (00:09:21:00).
Vole & owl – Defensive attack. The owl is following the vole inside the enclosure. The vole escapes but then approached the owl and leaps toward the owl (00:30:50:00).
Spiny mouse & barn owl – Escape jump. The spiny mouse escapes first to evade the attacking owl (00:14:01:00) but the it also leaps when bumping to a corner (00:14:02:13 and 00:14:05:13) or the front wall (00:14:07:00)
Acknowledgments
Acknowledgments
We are grateful to the zookeepers of the I. Meier Segals Gardens for Zoological Research in Tel-Aviv University for providing the animals, to N. Paz for editing this manuscript, and to T. Eckert for thoughtful comments.
Contributor Information
Shahaf Edut, Email: shahaf.edut@teva.co.il.
David Eilam, Email: eilam@post.tau.ac.il.
References
- D Eilam, Dayan T, Ben-Eliyahu S, Schulman I, Shefer G, Hendrie CA. Differential behavioural and hormonal responses of voles and spiny mice to owl calls. Animal Behaviour. 1999;58:1085–1093. doi: 10.1006/anbe.1999.1224. [DOI] [PubMed] [Google Scholar]
- Kieffer JD. The influence of apparent predation risk on the foraging behavior of eastern chipmunks (Tamias striatus) Canadian Journal of Zoology. 1991;69:2349–2351. [Google Scholar]
- Dickman CR. Predation and habitat shift in the house mouse, Mus domesticus. Ecology. 1992;73:313–322. [Google Scholar]
- Redpath SM. Impact of habitat fragmentation on activity and hunting behavior in the tawny owl, Strix aluco. Behavioral Ecology. 1995;6:410–415. [Google Scholar]
- Berger J. Pregnancy incentives, predation constraints and habitat shifts – experimental and field evidence for wild bighorn sheep. Animal Behaviour. 1991;41:61. [Google Scholar]
- Jones M, Mandelik Y, Dayan T. Coexistence of temporally partitioned spiny mice: Roles of habitat structure and foraging behavior. Ecology. 2001;82:2164–2176. [Google Scholar]
- Wooster D, Sih A. A review of the drift and activity responses of stream prey to Predator presence. Oikos. 1995;73:3–8. [Google Scholar]
- Abrams PA. Implications of dynamically variable traits for identifying, classifying, and measuring direct and indirect effects in ecological communities. American Naturalist. 1995;146:112–134. [Google Scholar]
- Turner AM. Contrasting short-term and long-term effects of predation risk on consumer habitat use and resources. Behavioral Ecology. 1997;8:120–125. [Google Scholar]
- Jedrzejewski W, Rychlik L, Jedrzejewska B. Responses of bank voles to odours of seven species of predators: experimental data and their relevance to natural predator-vole relationship. Oikos. 1993;68:251–257. [Google Scholar]
- Blanchard RJ, Blanchard DC. Antipredator defensive behaviors in a visible burrow system. Journal of Comparative Psychology. 1989;103:70–82. doi: 10.1037/0735-7036.103.1.70. [DOI] [PubMed] [Google Scholar]
- Longland W, Price MV. Direct observations of owls and heteromyid rodents – can predation risk explain microhabitat use. Ecology of Disease. 1991;72:2261–2273. [Google Scholar]
- Pongracz P, Altbacker V. Ontogeny of the responses of European rabbits (Oryctolagus cuniculus) to aerial and ground predators. Canadian Journal of Zoology. 2000;78:655–665. [Google Scholar]
- Kramer DL, Bonenfant M. Direction of predator approach and the decision to flee to a refuge. Animal Behaviour. 1997;54:289–295. doi: 10.1006/anbe.1996.0360. [DOI] [PubMed] [Google Scholar]
- Martin GR. Birds by Night. Carlton: Poyser. 1990.
- Fentress JC. Interrupted ongoing behaviour in two species of vole (Microtus agrestis and Clethrionomys britannicus) Animal Behaviour. 1968;16:135–153. doi: 10.1016/0003-3472(68)90124-3. [DOI] [PubMed] [Google Scholar]
- Erlinge S, Bergsten B, Kristiansson H. The stoat and its prey-hunting behaviour and fugitive reactions. Fauna Flora. 1974;69:203–211. [Google Scholar]
- Seyfarth RM, Cheney DL, Marler P. Monkey responses to three diffrenet alarm calls: evidence of predator classification and semantic communication. Science. 1980;210:801–803. doi: 10.1126/science.7433999. [DOI] [PubMed] [Google Scholar]
- Kotler BP, Brown JS, Slotow RH, Goodfriend WL, Strauss M. The influence of snakes on the foraging behavior of gerbils. Oikos. 1993;67:309–316. [Google Scholar]
- Macwhirter RB. Vocal and escape responses of Columbian ground squirrels to simulated terrestrial and aerial predator attacks. Ethology. 1992;91:311–325. [Google Scholar]
- Randall JA, Hatch SM, Hekkala ER. Interspecific variation in antipredator behavior in sympatric species of kangaroo rat. Behavioral Ecology and Sociobiology. 1995;36:243–250. [Google Scholar]
- Krause J, Cheng DJS, Kirkman E, Ruxton GD. Species-specific patterns of refuge use in fish: the role of metabolic expenditure and body length. Behaviour. 2000;137:1113–1127. [Google Scholar]
- Abramsky Z, Strauss E, Subach A, Kotler BP, Riechman A. The effect of barn owls (Tyto alba) on the activity and microhabitat selection of Gerbillus allenbyi and G. pyramidum. Oecologia. 1996;105:313–319. doi: 10.1007/BF00328733. [DOI] [PubMed] [Google Scholar]
- Kotler BP, Blaustein L, Brown JS. Predator facilitation: the combined effect of snakes and owls on the foraging behavior of gerbils. Annales Zoologici Fennici. 1992;29:199–206. [Google Scholar]
- Desy EA, Batzli GO, Liu J. Effects of food and predation on behavior of prairie voles – a field experiment. Oikos. 1990;58:159–168. [Google Scholar]
- Hendrie CA, Weiss SM. The development of an animal model of panic with predictive and face validity. In: Cooper SJ, Hendrie CA, editor. Ethology and Psychopharmacology. England: John Wiley and Sons Ltd; 1994. pp. 111–132. [Google Scholar]
- Hendrie CA, Weiss SM, Eilam D. Behavioural response of wild rodents to the calls of an owl: a comparative study. Journal of Zoology. 1998;245:439–446. [Google Scholar]
- Ronkainen H, Ylonen H. Behavior of cyclic bank voles under risk of mustelid predation – do females avoid copulations. Oecologia. 1994;97:377–381. doi: 10.1007/BF00317328. [DOI] [PubMed] [Google Scholar]
- Bolles RC. Species-specific defense reaction and avoidance learning. Psychology Reviews. 1970;77:32–48. [Google Scholar]
- Driver PM, Humphries DA. Protean Behavior. Ethology and Sociobiology. 1989;10:393–394. [Google Scholar]
- King CM. Interactions between woodland rodents and their predators. Symposium of the Zoological Society of London. 1985;55:219–247. [Google Scholar]
- Scarratt AM, Godin JGJ. Foraging and antipredator decisions in the hermit-crab Pagurus acadianus (Benedict) Journal of Experimental Marine Biology and Ecology. 1992;156:225–238. [Google Scholar]
- Smith WP. Ontogeny and adaptiveness of tail flagging behavior in white-tailed deer. American Naturalist. 1991;138:190–200. [Google Scholar]
- Lima SL, Dill LM. Behavioral decisions made under the risk of predation – a review and prospectus. Canadian Journal of Zoology. 1990;68:619–640. [Google Scholar]
- Blanchard RJ, Blanchard DC, Hori K. An ethoexperimental approach to the study of defense. In: Blanchard RJ, Brain PF, Blanchard DC, Parmigiani S, editor. An Ethoexperimental Approach to the Study of Behavior. Kluwer, Dordrecht, The Netherlands: Kluwer Academic Publishers; 1989. pp. 114–136. [Google Scholar]
- Lingle S, Wilson WF. Detection and avoidance of predators in white-tailed deer (Odocoileus virginianus) and mule deer (O. hemionus) Ethology. 2001;107:125–147. [Google Scholar]
- Ydenberg RC, Dill LM. The economics of fleeing from predators. Advances in the Study of Behavior. 1986;16:229–249. [Google Scholar]
- Robinson SR. Antipredator behaviour and predator recognition in Belding's ground squirrels. Animal Behaviour. 1980;28:840–852. [Google Scholar]
- Stanford CB. Avoiding predators: Expectations and evidence in primate antipredator behavior. International Journal of Primatology. 2002;23:741–757. [Google Scholar]
- Lima SL, Bednekoff PA. Temporal variation in danger drives antipredator behavior: The predation risk allocation hypothesis. American Naturalist. 1999;153:649–659. doi: 10.1086/303202. [DOI] [PubMed] [Google Scholar]
- Lima SL. Putting predators back into behavioral predator-prey interactions. Trends in Ecology & Evolution. 2002;17:70–75. [Google Scholar]
- Graham RR. The silent flight of owls. Journal of the Royal Aeronautical Society. 1934;38:837–843. [Google Scholar]
- Thorpe WH, Griffin DR. The lack of ultrasonic component in the flight noise of owls compared with other birds. Ibis. 1962;104:256–257. doi: 10.1038/193594b0. [DOI] [PubMed] [Google Scholar]
- Eilam D, Golani I. Home base behavior of rats (Rattus norvegicus) exploring a novel environment. Behavioural Brain Research. 1989;34:199–211. doi: 10.1016/s0166-4328(89)80102-0. [DOI] [PubMed] [Google Scholar]
- Eilam D. Open field behavior withstands drastic changes in arena size. Behavioral Brain Research. 2003;142:53–62. doi: 10.1016/s0166-4328(02)00382-0. [DOI] [PubMed] [Google Scholar]
- Curio E. Proximate and developmental aspects of antipredator behavior. Advances in the Study of Behavior. 1993;22:135–238. [Google Scholar]
- Carere C, Casetti R, de Acetis L, Perretta G, Cirulli F, Alleva E. Behavioural and nociceptive response in male and female spiny mice (Acomys cahirinus) upon exposure to snake odour. Behavioural Processes. 1999;47:1–10. doi: 10.1016/s0376-6357(99)00048-0. [DOI] [PubMed] [Google Scholar]
- Kavaliers M, Choleris E. Antipredator responses and defensive behavior: ecological and ethological approaches for the neurosciences. Neuroscience and Biobehavioral Reviews. 2001;25:577–586. doi: 10.1016/s0149-7634(01)00042-2. [DOI] [PubMed] [Google Scholar]
- Blanchard RJ, Flannelly KJ, Blanchard DC. Defensive behaviors of laboratory and wild Rattus norvegicus. Journal of Comparative Psychology. 1986;100:101–107. [PubMed] [Google Scholar]
- Selye H. The Physiology and Pathology of Exposure to Stress: A Treatise Based on the Concept of the General-Adaptation-Syndrome and the Diseases of Adaptation. Montreal: Acta Medica Publ. 1950.
- Blanchard RJ, Nikulina JN, Sakai RR, Mckittrick C, Mcewen B, Blanchard DC. Behavioral and endocrine change following chronic predatory stress. Physiology and Behavior. 1998;63:561–569. doi: 10.1016/s0031-9384(97)00508-8. [DOI] [PubMed] [Google Scholar]
- Levy A, Grauer E, Ben-Nathan D, Kloet ERd. New Frontiers in Stress Research: Modulation of Brain Function. Harwood Academic Publishers. 1998.
- Eilam D. Postnatal development of body architecture and gait in several rodent species. Journal of Experimental Biology. 1997;200:1339–1350. doi: 10.1242/jeb.200.9.1339. [DOI] [PubMed] [Google Scholar]
- Oron U, Maltz L, Shefer G, Eilam D. Histology and enzymatic activity in the postnatal development of limb muscles in rodents. Physiology & Behavior. 1998;63:651–657. doi: 10.1016/s0031-9384(97)00489-7. [DOI] [PubMed] [Google Scholar]
- Kronfeld-Schor N, Dayan T. The dietary basis for temporal partitioning: food habits of coexisting Acomys species. Oecologia. 1999;121:123–128. doi: 10.1007/s004420050913. [DOI] [PubMed] [Google Scholar]
- Birke LIA, D'Udine B, Albonetti ME. Exploratory behavior of two species of murid rodents, Acomys cahirinus and Mus musculus: a comparative study. Behavioral and Neural Biology. 1985;43:143–161. doi: 10.1016/s0163-1047(85)91337-8. [DOI] [PubMed] [Google Scholar]
- Mendelssohn H, Yom-Tov Y. Fauna Palaestina: Mammalia of Israel. Jerusalem: Keterpress Enterprises. 1999.
- Mikkola H, Willis I. Owls of Europe. England: T and A D Poyser Ltd. 1983.
- Koivunen V, Korpimaki E, Hakkarainen H. Refuge sites of voles under owl predation risk: priority of dominant individuals? Behavioral Ecology. 1998;9:261–266. [Google Scholar]
- Lingle S, Pellis SM. Fight or flight? Antipredator behavior and the escalation of coyote encounters with deer. Oecologia. 2002;131:154–164. doi: 10.1007/s00442-001-0858-4. [DOI] [PubMed] [Google Scholar]
- Sih A, McCarthy TM. Prey responses to pulses of risk and safety: testing the risk allocation hypothesis. Animal Behaviour. 2002;63:437–443. [Google Scholar]
- Steele MA. Effects of shelter and predators on reef fishes. Journal of Experimental Marine Biology and Ecology. 1999;233:65–79. [Google Scholar]
- Blanchard RJ, Parmigiani S, Bjornson C, Masuda C, Weiss SM, Blanchard DC. Antipredator behavior of Swiss-Webster mice in a visible burrow system. Aggressive Behavior. 1995;21:123–136. [Google Scholar]
- Ellard CG. Organization of escape movements from overhead threats in the Mongolian gerbil (Meriones unguiculatus) Journal of Comparative Psychology. 1993;107:242–249. doi: 10.1037/0735-7036.107.3.242. [DOI] [PubMed] [Google Scholar]
- Brunjes PC. The precocial mouse, Acomys cahirinus. Psychobiology. 1990;18:339–350. [Google Scholar]
- Greenberg G. Depth-perception in Mongolian gerbils (Meriones unguiculatus) and spiny mice (Acomys russatus and Acomys cahirinus) Journal of Comparative Psychology. 1986;100:81–84. [PubMed] [Google Scholar]
- Birke LIA, Sadler D. Maternal discrimination and the development of sex differences in exploratory-behavior in infant spiny mice (Acomys cahirinus) Quarterly Journal of Experimental Psychology Section B – Comparative and Physiological Psychology. 1991;43:403–430. doi: 10.1080/14640749108401277. [DOI] [PubMed] [Google Scholar]
- Goldman M, Skolnick AJ, Hernandez TP, Tobach E. Distance perception in the spiny mouse Acomys cahirinus – vertical jumping. Perceptual and Motor Skills. 1992;75:883–895. doi: 10.2466/pms.1992.75.3.883. [DOI] [PubMed] [Google Scholar]
- Birke LIA, Sadler D. Effects of odor familiarity on the development of systematic exploration in the spiny mouse, Acomys cahirinus. Developmental Psychobiology. 1987;20:627–640. doi: 10.1002/dev.420200608. [DOI] [PubMed] [Google Scholar]
- Marona-Lewicka D, Michaluk J, Antekiewicz-Michaluk L, Vetulani JA. A comparison of locomotor responses to some psychotropic drugs and cerebral receptors in the Acomys cahirinus and the laboratory mouse. Polish Journal of Pharmacology. 1987;39:293–302. [PubMed] [Google Scholar]
- Shkolnik A. Diurnal activity in a small desert rodents. International Journal of Biometeor. 1971;15:115–120. doi: 10.1007/BF01803884. [DOI] [PubMed] [Google Scholar]
- Kronfeld N, Shkolnik M. Adaptation to life in the desert in the brown hare (Lepus capensis) Journal of Mammalogy. 1996;77:171–178. [Google Scholar]
- Shargal E, Kronfeld-Schor N, Dayan T. Population biology and spatial relationships of coexisting spiny mice (Acomys) in Israel. Journal of Mammalogy. 2000;81:1046–1052. [Google Scholar]
- Shargal E, Rath-Wolfson L, Kronfeld N, Dayan T. Ecological and histological aspects of tail loss in spiny mice (Rodentia : Muridae, Acomys) with a review of its occurrence in rodents. Journal of Zoology. 1999;249:187–193. [Google Scholar]
- Selaas V. A comparison of the diet of sympatric owls in Aust-Agder county, Southern Norway. Fauna-Norvegia. 1993;26:7–13. [Google Scholar]
- Tome D. Diet composition of the long-eared owl in center Slovenia: seasonal variations in prey use. Journal of Raptor Research. 1994;28:253–258. [Google Scholar]
- Eilam D, Golani I. Home base behavior in amphetamine-treated tame wild rats (Rattus norvegicus) Behavioural Brain Research. 1990;36:161–170. doi: 10.1016/0166-4328(90)90170-j. [DOI] [PubMed] [Google Scholar]
- Eilam D, Golani I. Amphetamine induced stereotypy in rats: Its morphogenesis in locale space from normal exploration. In: Cooper SJ, Hendrie CA, editor. Ethology and Pharmacology. Chichester: John Wiley & Sons; 1994. pp. 241–266. [Google Scholar]
- Blumberg-Feldman H, Eilam D. Postnatal development of synchronous stepping in the gerbil (Gerbillus dasyurus) Journal of Experimental Biology. 1995;198:363–372. doi: 10.1242/jeb.198.2.363. [DOI] [PubMed] [Google Scholar]
- Eilam D, Dank M, Maurer R. Vole scale locomotion to the size of the open-field by adjusting the distance between stops: A possible link to path integration. Behavioural Brain Research. 2003;141:73–81. doi: 10.1016/s0166-4328(02)00322-4. [DOI] [PubMed] [Google Scholar]
- Huntingford FA. Some ethical issues raised by studies of predation and aggression. Animal Behaviour. 1984;32:210–215. [Google Scholar]
- Ben-Pazi A, Szechtman H, Eilam D. The morphogenesis of motor rituals in rats treated chronically with the dopamine agonist quinpirole. Behavioral Neuroscience. 2001;115:1301–1317. doi: 10.1037//0735-7044.115.6.1301. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Spiny mouse & barn owl. The spiny mouse is moving around until the owl appears 11 minutes after the spiny mouse was introduced (00:11:02:00). It then moves to the corner and remains immobile for a while, resuming locomotion in the center with relatively long pauses, until the owl launches its attack 21 minutes after the spiny mice was introduced (00:21:41:00. The spiny mouse responds in fleeing and orienting to the owl, until being caught by the owl (00:22:07:00).
Vole & barn owl. At first the owl is hidden and the vole moving freely. Once the owl appears and progresses to exposed roost (00:07:31:00), the vole leaps and runs to a corner, where it remains freezing until being caught by the owl (00:09:21:00).
Vole & owl – Defensive attack. The owl is following the vole inside the enclosure. The vole escapes but then approached the owl and leaps toward the owl (00:30:50:00).
Spiny mouse & barn owl – Escape jump. The spiny mouse escapes first to evade the attacking owl (00:14:01:00) but the it also leaps when bumping to a corner (00:14:02:13 and 00:14:05:13) or the front wall (00:14:07:00)