

Abstract
Compared with DR and DQ, knowledge of the binding repertoires and specificities of HLA-DP alleles is somewhat limited. However, a growing body of literature has indicated the importance of DP-restricted responses in the context of cancer, allergy, and infectious disease. In the current study, we developed high-throughput binding assays for the five most common HLA-DPB1 alleles in the general worldwide population. Using these assays on a comprehensive panel of single-substitution analogs and large peptide libraries, we derived novel detailed binding motifs for DPB1*0101 and DPB1*0501. We also derived more detailed quantitative motifs for DPB1*0201, DPB1*0401, and DPB1*0402, which were previously characterized on the basis of sets of eluted ligands and/or limited sets of substituted peptides. Unexpectedly, all five DP molecules, originally selected only on the basis of their frequency in human populations, were found to share largely overlapping peptide motifs. Testing panels of known DP epitopes and a panel of peptides spanning a set of Phleum pratense Ags revealed that these molecules also share largely overlapping peptide-binding repertoires. This demonstrates that a previously hypothesized DP supertype extends far beyond what was originally envisioned and includes at least three additional very common DP specificities. Taken together, these DP supertype molecules are found in >90% of the human population. Thus, these findings have important implications for epitope-identification studies and monitoring of human class II-restricted immune responses.
T cells recognize a complex between a specific MHC type and a particular Ag-derived epitope. Thus, a given epitope elicits a response only in individuals who express an MHC molecule capable of binding that particular epitope (1-4). MHC molecules are extremely polymorphic, with hundreds of different variants known in humans (5, 6). Much of the observed polymorphism is concentrated in the residues hypothesized to line the peptide-binding groove and form the specific pockets that engage the amino acid side chains of the peptide ligand. Therefore, addressing multiple MHC-binding specificities is required to allow coverage of the human patient populations targeted by an epitope-specific diagnostic or therapeutic intervention.
The issue of population coverage in relation to MHC polymorphism is further complicated by the fact that different MHC types are expressed at dramatically different frequencies in different ethnicities. Thus, without careful consideration, an epitope set associated with an ethnically biased population could result in decreased applicability and usefulness for any diagnostic immunoprophylactic or -therapeutic applications (7, 8). One means of circumventing the problem of MHC polymorphism relies on the selection of epitopes restricted by multiple MHC types. In the case of HLA class I, the identification of promiscuous epitopes is facilitated by the use of MHC supertypes, which defines sets of A and B class I molecules associated with largely overlapping peptidebinding repertoires (9-12).
In the context ofHLAclass II, four locimustbe considered:DRB1, DRB3/4/5,DQ,andDP. LikeHLAclass I, each locus is polymorphic, with specific alleles occurring with often widelyvariable frequencies in different ethnicities. Several studies suggested the existence of class II supertypes encompassing several DR and DQ specificities that, like class I supertypes, describe sets of moleculessharing largely overlapping peptide-binding repertoires (13-26).
Unlike the situation withHLA-DRand-DQ, forwhich hundreds of T cell epitopes have been identified and reported in the literature, relatively few HLA-DP epitopes have been described [e.g., see the listings compiled by the Immune Epitope Database: www.iedb.org (27)]. Similarly, knowledge of the binding repertoires and specificities ofHLA-DP alleles is, by comparison, somewhat limited. This is due, in part, to the common assumption that DP molecules are less important in the immune response than DR or DQ molecules, because the cell surface expression level of DP molecules seems to be ~10-fold lower than forDRorDQ(28-30). This lowexpression level also makes DP more difficult to isolate, purify, and characterize, which may have limited the number of studies conducted with these molecules. However, a growing body of literature indicates that DP encodes fully functional molecules, restricting epitope responses in the context of cancer, allergy, and infectious disease (31-44).
A putative motif for DPB1*0201, present in ~20% of the general population, has been defined using peptide-elution methodology (45, 46). The most frequent DPB alleles are DPB1*0401 and DPB1*0402, both of which are present in about one third of the individuals in the general population. Using peptide-binding studies, Maillere and coworkers defined motifs for both of these DPB1 subtypes (47, 48). Interestingly, these investigators showed that DPB1*0401 and DPB1*0402 share very similar peptide-binding specificity, with nearly identical main anchor motifs, leading them to hypothesize the existence of an HLA-DP supertype. The same investigators also noted that DPB1*0402 and DPB1*0201 differ by only a single residue in the P4 secondary pocket and that the two molecules share identical specificity at the main P1 and P6 anchor positions. Accordingly, they hypothesize that DP4 and DP2 molecules will also share a common repertoire of peptides. In contrast, Dong et al. (34) described a motif for DP9 that differs considerably from that recognized by DP4 and DP2 molecules.
In the current study, using comprehensive panels of single substitution analogs and large peptide libraries, we derived novel detailed binding motifs for two additional HLA-DP molecules: DPB1*0101 and DPB1*0501. We also provide more detailed motifs for DPB1*0201, DPB1*0401, and DPB1*0402, previously characterized on the basis of sets of eluted ligands and/or limited sets of substituted peptides. Unexpectedly, all five DP molecules, originally selected only on the basis of their frequency in human populations, were found to share largely overlapping peptide motifs. This led to the hypothesis that these molecules might also share largely overlapping peptide-binding repertoires. To test this hypothesis, we used a large set of binding data, encompassing several hundred nonredundant peptides, to examine the cross-reactivity between these five common DP molecules. Our findings demonstrate that the DP supertype, originally hypothesized by Malliere and coworkers, extends far beyond what was originally envisioned and includes at least three additional very common DP specificities. These DP supertype molecules are found in >90% of the human population; thus, these findings have important implications for epitope-identification studies and the monitoring of human class IIrestricted immune responses.
Materials and Methods
Peptide synthesis
Peptides for screening studies were purchased from Mimotopes (Clayton, Victoria, Australia) and/or A & A Labs (San Diego, CA) as crude material on a small (1-mg) scale. Peptides used as radiolabeled ligands were synthesized on a larger scale by A & A Labs and purified (>95%) by reversed-phase HPLC.
MHC purification
MHC molecules were purified from EBV-transformed homozygous cell lines by mAb-based affinity chromatography, essentially as described in detail elsewhere (49). Briefly, cells were maintained in vitro by culture in RPMI 1640 medium (Flow Laboratories, McLean, VA), supplemented with 2 mM l-glutamine (Life Technologies, Grand Island, NY), 100 U (100 mg/ml) penicillin-streptomycin solution (Life Technologies), and 10% heat-inactivated FCS (Hazelton Biologics, Lenexa, KS). Large-scale cultures were maintained in roller bottles. HLA molecules were purified from cells lysed at a concentration of 108 cells/ml in 50 mM Tris-HCl (pH 8.5), containing 1% (v/v) Nonidet P-40 (NP-40), 150 mM NaCl, 5 mM EDTA, and 2 mM PMSF. Lysates were passed through 0.45-μm filters and cleared of nuclei and debris by centrifugation at 10,000 × g for 20 min.
For affinity purification, columns of inactivated Sepharose CL4B and Protein A Sepharose were used as precolumns. HLA-DR, -DQ, and -DP molecules were captured by repeated passage of lysates over LB3.1 (anti–HLA-DR), SPV-L3 (anti–HLA-DQ), and B7/21 (anti–HLA-DP) columns. After two to four passages of the lysate, Ab columns were washed with 10-column volumes of 10 mM Tris-HCl (pH 8) with 1% (v/v) NP-40, 2-column volumes of PBS, and 2-column volumes of PBS containing 0.4% (w/v) n-octylglucoside. MHC molecules were then eluted with 50 mM diethylamine in 0.15 M NaCl containing 0.4% (w/v) n-octylglucoside (pH 11.5). A 1/26 volume of 2.0 M Tris (pH 6.8) was added to the eluate to reduce the pH to 8. Eluates were concentrated by centrifugation in Centriprep 30 microconcentrators at 2000 rpm (Millipore, Bedford, MA). Protein purity, concentration, and the effectiveness of depletion steps were monitored by SDS-PAGE and bicinchoninic acid assay.
MHC/peptide-binding assays
Assays to quantitatively measure peptide binding to class IIMHCmolecules are based on the inhibition of binding of a high-affinity radiolabeled peptide to purifiedMHCmolecules. This assay systemwasdescribed in detail elsewhere (49), but it is briefly described in general terms. The optimal assay conditions for each DP molecule tested in the current study are summarized in the Results. To measure the capacity of peptide ligands to bindMHC molecules, 0.1–1nMradiolabeled peptide was coincubated at room temperature or 37°C with 1 μM to 1 nM purified MHC in the presence of a mixture of protease inhibitors (EDTA, pepstatin A, N-ethylmaleimide, 1,10-phenanthroline, PMSF, and Nα-p-tosyl-l-lysine chloromethyl ketone). Following a 2–4 d incubation, the percentage of MHC-bound radioactivity was determined by capturing MHC/peptide complexes on LB3.1 (DR), L243 (DR), HB180 (DR/ DQ/DP), SPV-L3 (DQ), or B7/21 (DP) Ab-coated Optiplates (Packard Instrument, Meriden, CT) and determining bound cpm using the TopCount (Packard Instrument) microscintillation counter. For competitive assays, the concentration of peptide yielding 50% inhibition of the binding of a radiolabeled standard probe peptide are calculated. Under the conditions used, where [label] < [MHC] and IC50 ≥ [MHC], the measured IC50 values are reasonable approximations of the true Kd values (50, 51). Each competitor peptide was tested at six concentrations covering a 100,000-fold dose range in three or more independent experiments. As a positive control, the unlabeled version of the radiolabeled probe was also tested in each experiment.
Bioinformatic analyses
The peptide-binding repertoire (R) of each MHC molecule was defined as the set of peptides that bind that molecule with an affinity ≥1000 nM. The relationship between two molecules is measured by determining crossreactivity and repertoire overlap. Cross-reactivity is defined as the fraction of peptides that bind one MHC molecule (i) that also bind a second (ii): (Ri and Rz)/(Ri) × 100%. Repertoire overlap is defined as the fraction of peptides binding either molecule that bind both: (Ri and Rz)/(Ri or Rz) × 100%.
Population coverage was calculated as previously described (52). Gene frequencies (gf) for each HLA allele were calculated from population frequencies obtained from the MHC database [dbMHC; National Center for Biotechnology Information (NCBI), (53)]. Phenotypic frequencies (pf) were calculated using the binomial distribution formula: pf = 1 − (1−Σgf)2. To obtain total potential population coverage, no linkage disequilibrium was assumed.
Analysis of the single amino acid substitution (SAAS) binding data was performed essentially as described previously for analysis of positional scanning combinatorial library data (40). Briefly, the IC50 nanomolar value of each substituted peptide was standardized relative to the geometric mean IC50 nanomolar value of the wild-type peptide. Relative binding values were then represented in a 20 × 15-aa position matrix. Next, an average (geometric) relative binding affinity (ARB) was calculated for each position, encompassing all 20 possible residues. Finally, for each position, the ratio of ARB for the entire library/position-specific ARB was derived. We denominated this ratio, which describes the factor by which the geometric average binding affinity associated with all 20 residues at a specified position differs from that of the average binding affinity of the entire library, as the specificity factor (SF). Positions with the greatest specificity have the greatest SF value. Primary anchor positions were defined as those with SF ≥ 2.4. This criterion identifies positions where the majority of residues are associated with significant decreases in binding capacity.
Results
Selection of the DP allelic variants most common worldwide
HLA class II polymorphism is very extensive, and different MHC types are expressed at different frequencies in different ethnicities. Although DP α- and β-chains are polymorphic, polymorphism of the α-chain is generally limited to a few residues; for the most part, based on HLA-DR crystal structures, these residues are in regions not likely to affect binding specificity (54-56). Furthermore, DPA is overrepresented by just a few alleles, where DPA*0103 represents ~60% of all DPA genes, and four alleles account for ~90%. For these reasons, most studies focus on HLA-DP β-chains as the main determinant of HLA-DP peptide-binding specificity. The frequency of DPB1 allelic variants expressed in various ethnicities [dbMHC NCBI, (53)] was compiled with the goal of selecting a panel of DP molecules affording coverage of the majority of the human population, irrespective of ethnicity (Table I).
Table I.
Phenotypic frequencies of HLA-DPB1 alleles
| Allele | Phenotypic Frequency |
|||||||
|---|---|---|---|---|---|---|---|---|
| Australia | Europe | North America | Oceania | South America | Southeast Asia | Sub-Saharan Africa | Average | |
| DPB1*0401 | 19.8 | 68.1 | 51.5 | 34.0 | 18.3 | 31.3 | 10.7 | 33.4 |
| DPB1*0402 | 0.0 | 27.5 | 71.1 | 4.4 | 82.0 | 5.6 | 34.3 | 32.1 |
| DPB1*0501 | 76.8 | 3.4 | 4.0 | 49.2 | 0.0 | 43.4 | 0.5 | 25.3 |
| DPB1*0201 | 25.7 | 27.2 | 18.4 | 24.5 | 6.9 | 5.6 | 20.3 | 18.4 |
| DPB1*0101 | 1.5 | 9.3 | 1.7 | 32.7 | 3.0 | 5.6 | 52.1 | 15.1 |
| DPB1*0301 | 4.4 | 17.7 | 13.5 | 8.6 | 16.4 | 14.7 | 12.2 | 12.5 |
| DPB1*1301 | 1.5 | 2.1 | 0.0 | 13.6 | 0.0 | 24.9 | 8.7 | 7.3 |
| DPB1*1401 | 0.0 | 2.1 | 1.7 | 6.3 | 27.1 | 7.5 | 1.6 | 6.6 |
| DPB1*2201 | 30.8 | 0.0 | 0.0 | 0.3 | 0.0 | 0.0 | 0.2 | 4.5 |
| DPB1*2801 | 0.0 | 0.0 | 0.0 | 0.6 | 0.0 | 19.9 | 0.0 | 2.9 |
| DPB1*1801 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.9 | 2.0 |
| DPB1*1701 | 0.0 | 4.3 | 0.3 | 0.0 | 0.0 | 1.9 | 6.5 | 1.9 |
| DPB1*1101 | 1.5 | 2.6 | 2.0 | 0.0 | 0.0 | 0.0 | 4.7 | 1.5 |
| DPB1*1501 | 3.7 | 1.3 | 0.3 | 0.3 | 0.0 | 1.9 | 1.4 | 1.3 |
| DPB1*0202 | 0.0 | 1.3 | 0.0 | 0.3 | 0.0 | 5.6 | 0.2 | 1.1 |
| Study panel | 94.6 | 95.8 | 98.8 | 96.3 | 92.3 | 75.5 | 88.6 | 91.7 |
Phenotypic frequencies derived from data available at dbMHC (NCBI). Alleles included in the study panel are highlighted by bold type.
Accordingly, we selected a panel of five HLA-DPB1 molecules (DPB1*0101, DPB1*0201, DPB1*0401, DPB1*0402, and DPB1*0501) representing the most common allelic variants in major populations worldwide. Each DPB1 allele selected is encountered with an average phenotypic frequency >15% across the seven main populations for which frequency data are available (Australia, Europe, North America, Oceania, South America, Southeast Asia, and sub-Saharan Africa). In aggregate, these molecules cover ~92% of the average population at the DPB1 locus. Importantly, they afford equally high and balanced population coverage, irrespective of the ethnicity considered (range 75–99%, depending on the specific ethnicity considered). Of the targeted specificities, quantitative peptide-binding assays were reported by Maillere and coworkers for DPB1*0401 and DPB1*0402 (47, 48). To establish a complete panel covering all of the common specificities, we undertook development of binding assays for DPB1*0101, DPB1*0201, and DPB1*0501.
Establishment of HLA-DP binding assays
To establish assays for the DPB1*0101, DPB1*0201, and DPB1*0501 molecules, we first assembled a panel of potential binders, including peptides known to be HLA-DP epitopes or endogenous ligands presented by these specificities or known to bind other HLA-DP or class II molecules with high affinity. Candidate peptides were radiolabeled and tested in direct-binding assays for their capacity to bind DP molecules purified as described in Materials and Methods.
We found that a two-residue C-terminal truncation of a previously described high-affinity DP4 ligand [human aminopeptidase 285–299, sequence EKKYFAATQFEPLAA (47)] was the optimal radiolabeled ligand for DPB1*0101 purified molecules, whereas a two-residue N-terminal truncation of the same peptide (i.e., residues 287–301, sequence KYFAATQFEPLAARL) was optimal for DPB1*0201. The optimal radiolabeled ligand for DPB1*0501 was an M11 to Y, C7 to T, and R15 to A analog of a DP5 epitope [(57); sequence IGRIAETILGYNPSA]. The specificity of each assay was verified by assessing capture with various HLA class II locus-specific Abs (Supplemental Fig. 1).
Various assay conditions were examined to assess the effects of pH, temperature, duration of incubation, and MHC capture time. A representative analysis is shown in Supplemental Fig. 1. These experiments allowed us to identify optimal conditions for each specific molecular assay (Table II). In general, optimal assay conditions were similar to those used for HLA-DR assays (15, 58, 59). However, signals in DP-binding assays were substantially amplified by using an extended MHC-Ab capture step of up to 20–24 h. These assay conditions also proved optimal for the DPB1*0401 and DPB1*0402 molecules and were used in subsequent competitive-inhibition assays.
Table II.
Optimized conditions for HLA-DP binding assays
| Specificity |
Conditions |
|||||||
|---|---|---|---|---|---|---|---|---|
| DPB1 moleculea | Serological Ag | Preferred Cell Line | Sequence | Sourceb | Temperaturec | pH | Assay (h) | MHC Capture (h) |
| DPB1*0101 | DPw1 | VAVY | EKKYFAATQFEPLAA | A | RT | 5.5 | 72 | 24 |
| DPB1*0201 | DPw2 | JY | KYFAATQFEPLAARL | B | RT | 5.5 | 48 | 24 |
| DPB1*0401 | DPw4 | Pitout | KYFAATQFEPLAARL | B | RT | 7 | 48 | 24 |
| DPB1*0402 | DPw4 | AMAI | EKKYFAATQFEPLAA | A | 37°C | 7 | 72 | 24 |
| DPB1*0501 | DPw5 | HO301 | IGRIAETILGYNPSA | C | 37°C | 5.5 | 72 | 24 |
MHC purification and capture assays for all alleles were performed as described in Materials and Methods using the B7/21 mAb. All assays were performed in a final NP-40 concentration of 0.15% and with enough labeled peptide (0.1–1 nM) to provide 15,000 cpm of radioactivity.
For assays using the preferred cell line, the corresponding DPA1* chains were 0201, 0103, 01, 0301, and 0201 for DPB1*0101, 0201, 0401, 0402, and 0501, respectively.
A, human aminopeptidase 285–299; B, human aminopeptidase 287–301; C, human chimeric protein analog 1–15.
RT, room temperature.
Biological validation: binding capacity of reported DP epitopes
Following the biochemical validation of the assays described above, we sought to further validate them biologically by examining the correlation between assay specificity and the binding capacity of known HLA-restricted epitopes. We queried the Immune Epitope Database [www.iedb.org (27)] and the published literature (26, 60, 61) and selected a panel of 27 epitopes and endogenous ligands presented by one or more of the five common DP specificities selected in the current study (Table III). To control for locus specificity, we also selected a panel of 24 HLA-DQ–restricted epitopes.
Table III.
HLA-DP ligands and epitopes exhibit broad DP-binding capacity
| Peptidea | Source | HLA Restriction | HLA-DP Binding Capacity (IC50 nM) |
DP Bound (n) | ||||
|---|---|---|---|---|---|---|---|---|
| DPB1*0101 | DPB1*0201 | DPB1*0401 | DPB1*0402 | DPB1*0501 | ||||
| EKIQKAFDDIAKYFSK | H. sapiens TSA-2 | DPB1*0101 | 786 | 75 | 230 | 506 | 11 | 5 |
| LPSQAFEYILYNKG | H. sapiens cathepsin H | DPB1*0201 | 5.7 | 1.0 | 4.7 | 9.6 | 235 | 5 |
| VPDHVVWSLFNTL | H. sapiens CD225 | DPB1*0201 | 10 | 1.1 | 4.4 | 8.4 | 245 | 5 |
| GGFMTTAFQYIIDNKG | H. sapiens cathepsin S | DPB1*0201 | 14 | 1.4 | 2.0 | 11 | 534 | 5 |
| NNKYAASSYLSLTPE | H. sapiens Ig | DPB1*0201 | 8.6 | 2.4 | 4.8 | 42 | 707 | 5 |
| LQSLVSQYFQTVADY | B. taurus ApoA-II | DPB1*0201 | 1,058 | 2.7 | 6.4 | 12 | 929 | 4 |
| SGFLGPLLVLQAGFFLLTR | Hepatitis B S | DPB1*0201 | 4.0 | 3.3 | 5.5 | 4.8 | 421 | 5 |
| AFILDGDNLFPKV | B. pendula Bet v 1-A | DPB1*0201 | 65 | 15 | 45 | 140 | 360 | 5 |
| SVRFSWLSLLVPFVQWF | Hepatitis B S | DPB1*0201 | 25 | 36 | 52 | 179 | 24 | 5 |
| DKKETVWHLE | H. sapiens DP α-chain | DPB1*0201 | 785 | 93 | 665 | 3,743 | 18,382 | 3 |
| LTGYSLFQKEKMVLN | P. falciparum Merz S | DPB1*0201 | 25 | 180 | 215 | 1,271 | 180 | 4 |
| DTLRSYYADWYQQKPG | H. sapiens Ig | DPB1*0201 | 728 | 188 | 677 | 1,548 | 6,485 | 3 |
| ASDVETAEGGEIHELLRLQ | Measles V | DPB1*0201 | 318 | 560 | 841 | 289 | 8,591 | 4 |
| SDVGEFRAVTELG | H. sapiens DP β-chain | DPB1*0201 | 720 | 619 | 766 | 4,053 | 6,188 | 3 |
| SLLMWITQCFLPV | H. sapiens ESO | DPB1*0401 | 8.2 | 1.3 | 4.8 | 6.1 | 1,367 | 4 |
| NAQRFGISNYCQI | D pter Der p 1 | DPB1*0401, DPB1*0402, DPB1*0501 | 110 | 7.8 | 15 | 365 | 8,696 | 4 |
| WITQCFLPVFLAQPPSGQRR | H. sapiens ESO | DPB1*0401 | 8.3 | 12 | 22 | 25 | 297 | 5 |
| KIFGSLAFLPESFDGDPA | H. sapiens erbB 2 | DPB1*0401 | 8.2 | 9.0 | 26 | 143 | 289 | 5 |
| RELWWVFYAAD | Herpesvirus UL21 | DPB1*0401 | 331 | 291 | 113 | 1,652 | 979 | 4 |
| VLMEWLKTRPILSPLTKGIL | Influenza A M | DPB1*0401 | 9.3 | 58 | 115 | 12 | 6.4 | 5 |
| DKISDVSTIVPYIGPALNIV | C tetani tet toxin | DPB1*0401 | 162 | 165 | 151 | 173 | 3,007 | 4 |
| QSCRRPNAQRFGISNYCQI | D pter I29 | DPB1*0401 | 474 | 278 | 153 | 11,357 | 10,225 | 3 |
| DEPTLLYVLFEVFDV | Adenoviridae hexon | DPB1*0401 | 3,158 | 5,224 | 2,613 | 2,533 | 7,987 | 0 |
| FFLLTRILTIPQSLD | Hepatitis B S | DPB1*0402 | 0.88 | 0.30 | 0.60 | 1.2 | 59 | 5 |
| VFLQTHIFAEVLKDAIKDL | Herpesvirus EBNA 1 | DPw5 | 5.4 | 32 | 17 | 5.6 | 2.5 | 5 |
| THIFAEVLKD | Herpesvirus nuc ag 1 | DPw5 | 162 | 4,724 | 3,106 | 6,655 | 292 | 2 |
| RSQPGLCNMYKDSHHPARTA | H. sapiens MBP | DPB1*0501 | 2,017 | 4,588 | 13,310 | 7,779 | 1,389 | 0 |
| KPVSKMRMATPLLMQALP | H. sapiens Ii chain | DQB1*0201 | 52 | 161 | 291 | 138 | 22 | 5 |
| YQSYGPSGQYTHEFD | H. sapiens DQA | DQB1*0201 | 1,165 | 18,622 | 39,787 | 5,702 | 2,435 | 0 |
| TEDQAMEDIKQMEAESIS | B. taurus a-S1 casein | DQB1*0201 | 124 | 4,769 | 17,243 | 2,466 | 7,022 | 1 |
| DVKFPGGGQIVGGVYLLPRR | Hepatitis C polyprotein | DQB1*0301 | 47 | 16 | 9.1 | 33 | 4,042 | 4 |
| MGDDGVLACAIATHAKIRD | D pter Der p 2 | DQB1*0301 | 1,700 | 8,791 | 8,977 | 2,602 | 238 | 1 |
| HGSEPCIIHRGKPFQLEAV | D pter Der p 2 | DQB1*0301 | 135 | 820 | 3,092 | 1,409 | 6,127 | 2 |
| EYLNKIQNSLSTEWSPCSVT | P. falciparum CS | DQB1*0301 | 98 | 436 | 314 | 921 | 216 | 5 |
| VLVPGCHGSEPCIIHR | D pter Der p 2 | DQB1*0301 | 2,686 | 4,584 | 8,149 | 32,165 | 22,321 | 0 |
| CDGERPTLAFLQDVM | H. sapiens GAD65 | DQB1*0302 | 1,029 | 32,754 | 36,088 | 15,226 | 8,929 | 0 |
| DSNIMNSINNVMDEIDFFEK | P. falciparum 101 kDa | DQB1*0302 | 1,441 | 3,959 | 15,803 | 1,173 | 343 | 1 |
| DCLLCAYSIEFGTNISKEHD | G. gallus ovomucoid | DQB1*0302 | 85 | 343 | 372 | 232 | 827 | 5 |
| RMMEYGTTMVSYQPL | H. sapiens GAD65 | DQB1*0302 | 3,966 | 725 | 534 | 1,314 | 1,955 | 2 |
| FDRSTKVIDFHYPNE | H. sapiens GAD65 | DQB1*0302 | 15,792 | — | — | 30,395 | 13,699 | 0 |
| TPTEKDEYCARVNH | H. sapiens B2-m | DQB1*0302 | 36,584 | — | — | 13,994 | 19,231 | 0 |
| KDILEDERAAVDTYC | H. sapiens DRB1*0402 | DQB1*0401 | 1,733 | 10,468 | 26,251 | 4,195 | 9,690 | 0 |
| SDDELPYIDPNMEPV | Herpesvirus nuc | DQ5 | 2,157 | 3,811 | 17,339 | 3,091 | 3,382 | 0 |
| PLYRYLGGSFSHVL | C. trachomatis enolase | DQB1*0501 | 9.9 | 13 | 10 | 149 | 8,130 | 4 |
| EELKSLNSVQAQYA | C. trachomatis CT579 | DQB1*0501 | 18 | 310 | 318 | 975 | 8,741 | 4 |
| INEPTAAAIAYGLDR | H. sapiens HSP70 | DQB1*0602 | 5,690 | 4,146 | 7,276 | 11,599 | 29,412 | 0 |
| PPLYATGRLSQAQLMPSPPM | Herpesvirus UL48 | DQB1*0602 | 356 | 14,120 | 12,273 | 855 | 999 | 3 |
| NNYGSTIEGLLD | Herpesvirus VP16 | DQB1*0602 | 3,379 | 5,098 | 9,708 | 5,875 | 49,505 | 0 |
| RGYFKMRTGKSSIMRS | Influenza A HA | DQB1*0602 | 911 | 7,119 | 6,565 | 1,527 | 806 | 2 |
| NPRDAKACVVHGSDLK | H. sapiens ATPase | DQB1*0602 | 3,147 | 2,359 | 8,295 | 6,402 | 15,773 | 0 |
| EEVDMTPADALDDFD | Herpesvirus UL48 | DQB1*0201, DQB1*0302 | 3,489 | 5,798 | 22,322 | 26,042 | — | 0 |
Bold font highlights known epitopes binding to their reported restricting allele with an affinity less than 1000 nM.
Each of the peptides was tested for its capacity to bind DPB1*0101, DPB1*0201, DPB1*0401, DPB1*0402, and DPB1*0501. The resulting IC50 nanomolar values are shown in Table III. In 90% of the cases, a DP-restricted epitope bound its corresponding restricting allele with a binding affinity ≥1000 nM; >50% of the cases were associated with affinities ≥100 nM (Fig. 1).
FIGURE 1.
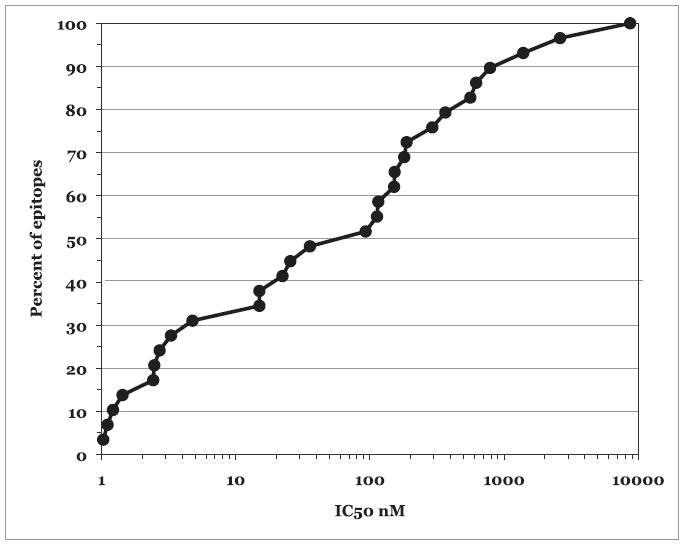
HLA-DP binding affinity of DP epitopes and ligands. The IC50 nanomolar binding capacity of known HLA-DP epitopes and ligands is shown as a function of the fraction of known epitopes and ligands that have the same or better binding affinity for their respective restricting HLA-DP allele.
A striking degree of cross-reactivity was also noted between the different DP allelic variants in the context of the panel of HLA-DP–restricted epitopes and endogenous ligands, with the majority (71.4%) of peptides binding four or more of the five molecules with an affinity ≥1000 nM (Fig. 2). This might have been predicted based on the fact that a similar radiolabeled ligand was found to be suitable for four of the DP assays studied herein, as well as previous observations by Maillere and coworkers regarding the existence of a potential HLA-DP supertypic specificity (47). In contrast, only occasional DP binding and cross-reactivities were detected with the panel of DQ-restricted epitopes and ligands. In conclusion, the data presented in this section support the biological relevance of the assays developed.
FIGURE 2.
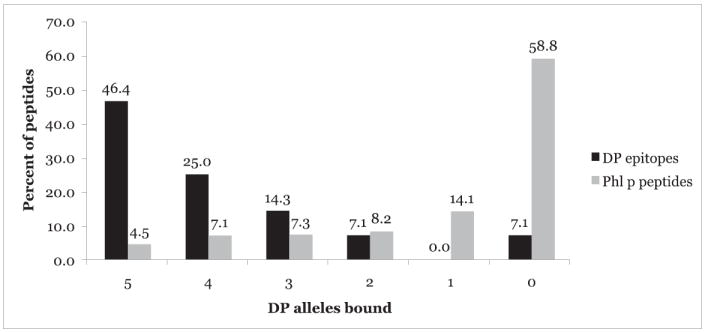
HLA-DP epitopes are frequently promiscuous DP supertype binders. The percentage of peptides in a panel of known HLA-DP epitopes and ligands or a panel of 425 nonredundant peptides spanning a set of P. pratense (Phl p) pollen Ags that bound the specified number of DP molecules.
Definition of peptide-binding motifs for HLA-DPB1*0101, -DPB1*0201, and -DPB1*0501 molecules
Next, we sought to use the newly developed assays to define motifs for DPB1*0101, DPB1*0201, and DPB1*0501. To accomplish this task, we took advantage of the observation that the human aminopeptidase 285–299 peptide (sequence EKKYFAATQFEPLAA) used as the ligand for the DP*0101 and DP*0402 assays, fortuitously bound, albeit with varying affinities (0.66–8.3 nM), to the each of the various DP molecules in consideration. A full set of SAAS analogs was synthesized; all 19 possible amino acid substitutions were represented for each position (1-15) of the wild-type peptide.
The entire panel of analogswas then tested for binding in each of the novel DP assays. Binding data were evaluated as described in Materials and Methods and summarized asARBs in allele-specific 20 × 15-aa position matrices. The role of each position was determined by calculating an SF for each position, which describes the factor by which the geometric average binding affinity associated with all 20 residues at a specified position differs from that of the average affinity of the entire library (62, 63). Positions with the greatest specificity have the greatest SF value. Primary anchor positions were defined as those with SF ≥ 2.4.
In a set of preliminary experiments, we tested the panel of SAASs in the DPB1*0401 and DPB1*0402 assays, for which Maillere and coworkers defined a motif based on two hydrophobic/ aromatic main anchors at positions 1 and 6 of a prototypical class II nine-residue core (47). Our results confirmed these observations. For DPB1*0401, positions F5 and F10, corresponding to the P1 and P6 anchors, had the greatest influence on binding capacity. These positions were associated with SFs of 15 and 23, respectively; 13 substitutions at position F5 and 14 substitutions at F10 affected binding by ≥10-fold (Supplemental Table IA). No other positions were associated with an SF > 2.0 or had more than two substitutions with a ≥10-fold effect on binding. For DPB1*0402, positions F5, F10, and L13, corresponding to the P1, P6, and P9 anchors, had SFs of 7.7, 8.0, and 3.4, respectively. At positions F5 and F10, 11 substitutions affected binding by ≥10-fold, as did 7 substitutions at L13 (Supplemental Table IB). For both alleles, the most preferred residues at the P1 and P6 anchors were found to be aromatic or hydrophobic. Taken together, these results confirm the observations of Maillere’s group and further validate the assays established.
When the same panel of SAASs was tested for DPB1*0101 binding, positions F5 and F10 had SFs of 3.7 and 2.4, respectively, and nine and seven substitutions at these positions affected binding by ≥10-fold (Table IV). These main anchor positions are identical to the P1 and P6 anchor positions similarly defined for the DPB1*0401 and 0402 molecules. In addition, a very prominent role was demonstrated for position N-2, corresponding to residue K3, for which an SF of 9.4 was calculated, and 13 of the substitutions affected binding to DPB1*0101. No other position was associated with SF > 1.8 or more than three substitutions with >10-fold effects on DPB1*0101 binding.
Table IV.
SAAS analysis of the binding specificity of HLA-DPB1*0101
| E | K | K | Y | F | A | A | T | Q | F | E | P | L | A | A | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Substitution | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
| A | 0.18 | 0.89 | 0.38 | 0.066 | 0.13 | 1.0 | 1.0 | 1.1 | 1.7 | 0.27 | 2.3 | 2.7 | 1.4 | 1.0 | 1.0 |
| C | 0.59 | 0.80 | 0.038 | 0.83 | 0.082 | 0.37 | 0.54 | 0.35 | 1.8 | 0.049 | 0.51 | 0.95 | 0.18 | 1.9 | 3.9 |
| D | 0.43 | 0.49 | 0.017 | 2.0 | 0.022 | 0.19 | 2.2 | 0.20 | 1.2 | 0.041 | 1.4 | 1.5 | 0.27 | 1.6 | 0.98 |
| E | 1.0 | 0.52 | 0.030 | 0.54 | 0.018 | 1.5 | 0.75 | 0.90 | 2.2 | 0.42 | 1.0 | 0.45 | 2.8 | 2.8 | 2.4 |
| F | 0.23 | 0.48 | 0.040 | 0.73 | 1.0 | 2.1 | 4.4 | 2.7 | 3.3 | 1.0 | 2.7 | 1.7 | 0.56 | 1.3 | 1.8 |
| G | 0.70 | 0.85 | 0.037 | 0.34 | 0.057 | 2.8 | 1.6 | 2.6 | 0.43 | 0.087 | 3.6 | 1.3 | 0.33 | 0.62 | 2.4 |
| H | 0.54 | 0.31 | 0.027 | 0.15 | 0.097 | 0.62 | 1.2 | 0.70 | 1.0 | 0.024 | 0.87 | 0.74 | 0.036 | 4.5 | 2.0 |
| I | 1.7 | 0.11 | 0.027 | 0.45 | 0.092 | 8.4 | 3.0 | 0.72 | 1.3 | 0.73 | 3.3 | 0.79 | 0.84 | 1.7 | 0.83 |
| K | 1.0 | 1.0 | 1.0 | 0.12 | 0.024 | 0.55 | 0.27 | 0.30 | 1.1 | 0.028 | 0.91 | 0.48 | 0.025 | 1.9 | 0.20 |
| L | 0.7 | 0.21 | 0.053 | 0.86 | 0.99 | 2.9 | 1.7 | 20.0 | 3.5 | 2.5 | 1.2 | 5.3 | 1.0 | 1.7 | 1.2 |
| M | 1.0 | 0.24 | 0.060 | 0.21 | 0.87 | 1.3 | 1.9 | 2.8 | 1.1 | 2.7 | 1.3 | 4.2 | 0.18 | 1.4 | 0.10 |
| N | 1.2 | 0.0035 | 0.0069 | 0.53 | 0.046 | 0.39 | 2.1 | 3.4 | 0.57 | 0.53 | 0.085 | 0.9 | 1.1 | 0.42 | 0.53 |
| P | 0.49 | 0.15 | 0.066 | 0.55 | 0.21 | 0.18 | 1.5 | 0.40 | 1.0 | 0.61 | 1.7 | 1.0 | 0.40 | 1.4 | 1.4 |
| Q | 0.29 | 0.59 | 0.014 | 1.0 | 0.15 | 0.74 | 3.8 | 2.4 | 1.0 | 0.35 | 1.9 | 3.0 | 0.63 | 1.3 | 0.71 |
| R | 0.43 | 0.30 | 0.011 | 0.0030 | 0.39 | 5.2 | 1.5 | 0.15 | 1.8 | 0.031 | 1.4 | 1.5 | 0.025 | 4.8 | 0.40 |
| S | 0.25 | 0.45 | 0.24 | 0.91 | 0.56 | 1.1 | 2.1 | 0.85 | 1.7 | 0.16 | 1.6 | 5.0 | 0.75 | 1.5 | 0.28 |
| T | 0.22 | 0.51 | 0.14 | 0.87 | 0.079 | 2.1 | 1.9 | 1.0 | 1.4 | 0.10 | 0.66 | 2.8 | 1.1 | 2.6 | 1.0 |
| V | 0.23 | 0.57 | 0.29 | 0.91 | 0.85 | 1.1 | 2.8 | 1.5 | 2.9 | 0.39 | 2.2 | 1.7 | 1.9 | 0.60 | 0.43 |
| W | 1.1 | 0.49 | 0.29 | 0.97 | 0.35 | 1.5 | 3.1 | 0.35 | 2.2 | 0.35 | 0.88 | 1.1 | 0.37 | 1.8 | 0.43 |
| Y | 2.7 | 1.1 | 0.46 | 1.0 | 0.63 | 6.0 | 4.1 | 5.5 | 2.2 | 2.6 | 0.66 | 2.3 | 0.93 | 3.6 | 0.44 |
| Summary | |||||||||||||||
| Geomeana | 0.57 | 0.35 | 0.066 | 0.40 | 0.16 | 1.2 | 1.7 | 1.1 | 1.5 | 0.25 | 1.2 | 1.5 | 0.41 | 1.6 | 0.79 |
| SFb | 1.1 | 1.8 | 9.4 | 1.5 | 3.7 | 0.51 | 0.36 | 0.57 | 0.42 | 2.4 | 0.52 | 0.40 | 1.5 | 0.38 | 0.78 |
| 10-fold effectsc | 0 | 1 | 13 | 2 | 9 | 0 | 0 | 1 | 0 | 7 | 1 | 0 | 3 | 0 | 1 |
Values reflect the relative binding capacity of a peptide bearing the indicated residue substituted in the specified position of peptide EKKYFAATQFEPLAA. Bold type represents geomean < 0.25, SF > 2.4, or ≥10-fold effects on binding.
Geomean: geometric mean relative binding capacity of all substitutions at the indicated position.
Ratio of the geomean of all substitutions at the specified position to the geomean of all 300 possible substitutions.
10-fold effects: number of substitutions at each position with a 10-fold increase or decrease in binding capacity.
The SAAS panel was next tested for DPB1*0201 binding. At residues F5 and F10, again corresponding to the P1 and P6 anchors, SFs of 9.1 and 13 were calculated, respectively, and 12 and 14 substitutions affected binding by ≥10-fold, respectively (Table V). In addition, a very prominent role was once again demonstrated for position N-2, corresponding to residue K3; an SF of 3.1 was calculated, and eight of the substitutions affected DPB1*0201 binding by > 10-fold. No other position was associated with an SF > 2.1 or more than five substitutions with > 10-fold effects on DPB1*0201 binding.
Table V.
SAAS analysis of the binding specificity of HLA-DPB1*0201
| E | K | K | Y | F | A | A | T | Q | F | E | P | L | A | A | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Substitution | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
| A | 0.95 | 0.41 | 0.36 | 0.20 | 0.081 | 1.0 | 1.0 | 2.8 | 1.4 | 0.035 | 0.93 | 6.3 | 0.65 | 1.0 | 1.0 |
| C | 0.58 | 0.43 | 0.087 | 0.54 | 0.14 | 0.38 | 0.51 | 0.24 | 2.2 | 0.072 | 1.5 | 3.0 | 0.20 | 2.0 | 1.8 |
| D | 0.37 | 0.12 | 0.19 | 0.73 | 0.063 | 0.36 | 0.91 | 1.2 | 2.5 | 0.0022 | 0.64 | 0.75 | 0.21 | 2.1 | 0.71 |
| E | 1.0 | 0.14 | 0.056 | 0.56 | 0.0060 | 0.41 | 0.65 | 1.3 | 0.77 | 0.0023 | 1.0 | 0.68 | 8.2 | 2.3 | 1.2 |
| F | 0.30 | 0.29 | 0.37 | 0.42 | 1.0 | 1.4 | 1.6 | 1.5 | 3.2 | 1.0 | 1.9 | 2.8 | 0.69 | 1.2 | 1.2 |
| G | 1.9 | 0.30 | 0.12 | 0.30 | 0.0058 | 0.47 | 1.1 | 1.6 | 0.97 | 0.010 | 2.2 | 0.91 | 0.25 | 0.25 | 1.0 |
| H | 0.45 | 0.14 | 0.070 | 0.78 | 0.045 | 0.66 | 0.61 | 0.59 | 0.98 | 0.0051 | 1.2 | 0.96 | 0.29 | 1.6 | 0.81 |
| I | 0.80 | 0.14 | 0.052 | 0.26 | 0.27 | 0.89 | 2.3 | 0.64 | 1.6 | 0.32 | 1.8 | 1.0 | 0.57 | 4.0 | 0.72 |
| K | 1.1 | 1.0 | 1.0 | 0.30 | 0.0045 | 0.63 | 0.73 | 0.22 | 1.5 | 0.0071 | 0.94 | 1.1 | 0.069 | 1.1 | 0.40 |
| L | 0.87 | 0.22 | 0.017 | 1.2 | 0.56 | 1.1 | 1.5 | 98.0 | 1.2 | 1.7 | 2.0 | 61.0 | 1.0 | 0.50 | 1.4 |
| M | 1.5 | 0.24 | 0.13 | 0.29 | 0.25 | 0.67 | 0.72 | 0.67 | 0.86 | 0.28 | 1.1 | 34.0 | 0.51 | 0.72 | 0.086 |
| N | 0.66 | 0.21 | 0.027 | 2.2 | 0.017 | 0.81 | 0.82 | 0.98 | 1.3 | 0.018 | 0.21 | 4.5 | 0.95 | 0.60 | 0.38 |
| P | 0.47 | 0.37 | 0.069 | 0.64 | 0.024 | 0.037 | 1.1 | 5.7 | 0.69 | 0.012 | 0.64 | 1.0 | 0.88 | 1.8 | 0.84 |
| Q | 0.33 | 0.19 | 0.11 | 1.8 | 0.019 | 0.82 | 8.5 | 2.3 | 1.0 | 0.034 | 0.93 | 12.0 | 0.82 | 3.8 | 0.56 |
| R | 0.51 | 0.21 | 0.033 | 0.67 | 0.028 | 3.5 | 1.2 | 0.33 | 1.2 | 0.0017 | 1.6 | 4.6 | 0.32 | 6.0 | 0.28 |
| S | 0.53 | 0.34 | 1.1 | 0.81 | 0.013 | 0.85 | 8.6 | 1.4 | 2.0 | 0.026 | 1.2 | 43.0 | 0.34 | 2.3 | 0.43 |
| T | 0.38 | 0.24 | 0.81 | 0.56 | 0.0068 | 1.0 | 0.74 | 1.0 | 1.5 | 0.012 | 0.90 | 13.0 | 0.40 | 1.3 | 0.14 |
| V | 0.28 | 0.52 | 0.74 | 1.6 | 0.61 | 1.1 | 1.1 | 1.1 | 0.57 | 0.075 | 38.0 | 1.9 | 0.96 | 0.92 | 1.0 |
| W | 0.33 | 0.18 | 1.2 | 0.55 | 0.72 | 1.2 | 0.99 | 0.28 | 2.2 | 1.4 | 1.3 | 2.6 | 0.58 | 0.97 | 0.74 |
| Y | 0.53 | 0.46 | 1.9 | 1.0 | 0.43 | 1.5 | 8.6 | 1.6 | 2.5 | 1.2 | 3.2 | 1.8 | 5.4 | 2.3 | 0.54 |
| Summary | |||||||||||||||
| Geomeana | 0.59 | 0.27 | 0.18 | 0.62 | 0.062 | 0.72 | 1.3 | 1.2 | 1.4 | 0.042 | 1.4 | 3.5 | 0.58 | 1.4 | 0.62 |
| SFb | 0.95 | 2.1 | 3.1 | 0.90 | 9.1 | 0.78 | 0.42 | 0.47 | 0.41 | 13 | 0.41 | 0.16 | 0.96 | 0.40 | 0.91 |
| 10-fold effectsc | 0 | 0 | 8 | 0 | 12 | 1 | 0 | 1 | 0 | 14 | 1 | 5 | 1 | 0 | 1 |
Values reflect the relative binding capacity of a peptide bearing the indicated residue substituted in the specified position of peptide KKYFAATQFEPLAA. Bold type represents geomean < 0.25, SF > 2.4, or ≥10-fold effects on binding.
Geomean: geometric mean relative binding capacity of all substitutions at the indicated position.
Ratio of the geomean of all substitutions at the specified position to the geomean of all 300 possible substitutions.
10-fold effects: number of substitutions at each position with a 10-fold increase or decrease in binding capacity.
Finally, the panel of SAASs was tested for DPB1*0501 binding. Again, the most significant influences on binding were detected at residues F5 and F10, corresponding to the P1 and P6 anchors, for which SFs of 9.0 and 9.6 were found, respectively; 13 substitutions for each site affected binding by ≥10-fold (Table VI). Position N-4, corresponding to residue E1, had an SF of 2.3, just below the threshold we used to define main anchors, and seven of the substitutions tested affected binding by >10-fold, suggesting that this position has at least some secondary influence on DPB1*0501 binding capacity.
Table VI.
SAAS analysis of the binding specificity of HLA-DPB1*0501
| E | K | K | Y | F | A | A | T | Q | F | E | P | L | A | A | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Substitution | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
| A | 0.72 | 0.17 | 0.48 | 0.070 | 0.028 | 1.0 | 1.0 | 0.75 | 1.0 | 0.053 | 0.40 | 3.2 | 2.7 | 1.0 | 1.0 |
| C | 0.32 | 0.18 | 0.31 | 0.11 | 0.057 | 0.30 | 1.6 | 0.65 | 1.5 | 0.067 | 1.5 | 1.2 | 0.61 | 2.0 | 1.0 |
| D | 0.32 | 0.16 | 0.43 | 0.24 | 0.0083 | 0.13 | 0.47 | 0.54 | 0.72 | 0.0047 | 0.22 | 0.99 | 0.39 | 1.2 | 1.4 |
| E | 1.0 | 0.11 | 0.28 | 0.16 | 0.039 | 1.7 | 0.39 | 1.2 | 1.5 | 0.0029 | 1.0 | 2.6 | 0.18 | 0.86 | 3.5 |
| F | 0.059 | 0.12 | 1.1 | 0.75 | 1.0 | 2.7 | 3.4 | 0.74 | 13.0 | 1.0 | 0.70 | 6.5 | 2.6 | 1.2 | 1.8 |
| G | 0.10 | 0.16 | 1.0 | 0.068 | 0.011 | 0.078 | 5.5 | 0.24 | 0.16 | 0.027 | 0.40 | 1.8 | 0.80 | 1.2 | 2.1 |
| H | 0.10 | 0.13 | 0.50 | 0.057 | 0.013 | 0.12 | 1.2 | 0.19 | 0.79 | 0.012 | 0.11 | 2.2 | 0.34 | 2.1 | 1.5 |
| I | 0.084 | 0.094 | 0.65 | 0.16 | 0.10 | 4.6 | 0.12 | 0.78 | 1.1 | 0.17 | 2.1 | 1.5 | 1.6 | 2.8 | 0.42 |
| K | 0.094 | 1.0 | 1.0 | 0.078 | 0.59 | 1.8 | 0.45 | 1.1 | 0.95 | 0.15 | 0.17 | 2.0 | 0.12 | 1.6 | 0.69 |
| L | 0.058 | 0.62 | 0.86 | 0.35 | 0.19 | 2.5 | 1.6 | 2.9 | 29.0 | 0.19 | 5.5 | 2.4 | 1.0 | 2.6 | 1.2 |
| M | 0.030 | 0.46 | 2.5 | 0.12 | 0.098 | 0.75 | 0.71 | 0.86 | 1.3 | 0.16 | 7.2 | 1.0 | 0.33 | 0.58 | 0.15 |
| N | 0.032 | 0.25 | 0.17 | 0.51 | 0.0024 | 0.39 | 0.69 | 0.15 | 1.5 | 0.024 | 0.36 | 0.82 | 1.0 | 0.98 | 0.66 |
| P | 0.031 | 0.11 | 0.38 | 0.63 | 0.0024 | 0.078 | 0.23 | 0.53 | 0.62 | 0.027 | 0.082 | 1.0 | 2.1 | 1.7 | 1.2 |
| Q | 0.87 | 0.26 | 1.5 | 0.75 | 0.015 | 1.4 | 1.5 | 1.1 | 1.0 | 0.010 | 1.0 | 2.1 | 0.23 | 1.4 | 0.15 |
| R | 0.58 | 0.26 | 0.18 | 0.21 | 0.30 | 1.9 | 1.0 | 0.22 | 1.2 | 0.0025 | 1.0 | 3.4 | 0.11 | 27.0 | 0.032 |
| S | 0.45 | 0.40 | 0.62 | 0.27 | 0.024 | 1.1 | 1.5 | 0.93 | 1.9 | 0.057 | 0.34 | 2.2 | 1.9 | 1.9 | 0.051 |
| T | 0.50 | 0.32 | 0.21 | 0.58 | 0.048 | 0.23 | 1.6 | 1.0 | 1.1 | 0.044 | 0.15 | 1.5 | 1.8 | 1.3 | 0.048 |
| V | 0.46 | 0.44 | 0.22 | 0.68 | 0.27 | 3.9 | 3.2 | 1.2 | 3.4 | 0.037 | 2.0 | 3.6 | 3.0 | 1.6 | 0.039 |
| W | 0.31 | 0.21 | 0.66 | 0.87 | 0.14 | 4.4 | 3.5 | 0.13 | 2.3 | 0.54 | 0.13 | 2.0 | 1.5 | 0.60 | 0.054 |
| Y | 0.79 | 0.40 | 0.95 | 1.0 | 0.13 | 2.1 | 7.6 | 0.20 | 2.3 | 0.26 | 0.72 | 3.7 | 1.3 | 8.0 | 0.070 |
| Summary: | |||||||||||||||
| Geomeana | 0.20 | 0.24 | 0.54 | 0.26 | 0.050 | 0.82 | 1.2 | 0.57 | 1.5 | 0.047 | 0.58 | 2.0 | 0.78 | 1.7 | 0.37 |
| SFb | 2.3 | 1.9 | 0.84 | 1.7 | 9.0 | 0.55 | 0.39 | 0.80 | 0.30 | 9.6 | 0.79 | 0.23 | 0.59 | 0.26 | 1.2 |
| 10-fold effectsc | 7 | 1 | 0 | 4 | 13 | 2 | 0 | 0 | 2 | 13 | 1 | 0 | 0 | 1 | 6 |
Values reflect the relative binding capacity of a peptide bearing the indicated residue substituted in the specified position of peptide EKKYFAATQFEPLAA. Bold type represents geomean < 0.25, SF > 2.4, or ≥10-fold effects on binding.
Geomean: geometric mean relative binding capacity of all substitutions at the indicated position.
Ratio of the geomean of all substitutions at the specified position to the geomean of all 300 possible substitutions.
10-fold effects: number of substitutions at each position with a 10-fold increase or decrease in binding capacity.
Taken together, the data in this section defined comprehensive quantitative bindingmotifs for each of the fivemost common HLA-DP specificities in the general population. In addition to identifying specific main anchor positions and preferences, these motifs provide information about specificity at other positions and the effect on binding of each of the 20 naturally occurring amino acids.
Allelic-specific and consensus motif associated with DP peptide-binding specificity
A summary of the motif associated with each of the five common DP allelic variants is presented in Table VII. All five molecules are associated with a P1–P6 spacing of two main anchor residues, for which a prominent specificity for hydrophobic and aromatic residues is generally noted.
Table VII.
Summary of HLA-DP motifs and definition of a DP supermotif
| DPB1 Molecule | N-2 | P1 | P6 | P9 |
|---|---|---|---|---|
| DPB1*0101 | AKSVWYa | FLMPRSVWY | AEFILMNPQVWY | |
| DPB1*0201 | AFKSTVWY | FILMVWY | FILMWY | |
| DPB1*0401 | FIVWY | FLMWY | ||
| DPB1*0402 | FILMVWY | FILMWY | ADEFILMNTVW | |
| DPB1*0501 | FKRV | FWY | ||
| HLA-DP supermotifb | FILMVWY | FILMWY |
Positions are defined relative to the motif for HLA-DPB1*0401 and 0402, as described in the text. Anchor positions are defined as described in the text, highlighting positions where the SF was ≥ to 2.4.
Residues shown are preferred at the indicated anchor positions, and are associated with a relative affinity within 5-fold of the optimal residue.
The supermotif is defined by residues at the corresponding positions that are preferred by three or more of the HLA-DPB1 alleles studied.
However, each allele is also associated with it own unique binding specificity; in two cases, the position 2 residues to the N-terminal side of this core also seemed to significantly affect overall binding capacity. These results, indicating significant similarity in the mode and specificity of binding, are in agreement with the testing of the known epitopes described above, which showed a high degree of similarity in the binding pattern of different DP molecules and clearly demonstrated that each allele is associated with its own binding specificity.
In conjunction with the observations of Maillere and coworkers, these results define a broad DP supertype that includes several DP molecules with specificity for aromatic or hydrophobic residues at positions 1 and 6. This proposed shared specificity (the DP supermotif) is shown in Table VII.
Validation of the proposed supermotif by testing overlapping peptide sets for binding to the various DP allelic molecules
The data presented above demonstrated that the various allelic variants considered have a common motif, but they did not address to what extent corresponding overlaps in binding repertoire exist. To address this point, we tested a panel of 425 nonredundant peptides for its capacity to bind each DP molecule. The set of peptides consisted of 15-mers, overlapping by 10 residues, spanning the entire sequences of the Phleum pratense 1, 2, 3, 4, 5, 6, 7, 11, 12, and 13 pollen Ags, which are implicated in allergic reaction to timothy grass.
We found that 250 (58.8%) of the P. pratense peptides bound none of the DP molecules, and 60 (14.1%) bound only one (Fig. 2). Thirty-five (8.2%) other peptides bound two of the different DP molecules tested. Thirty-one (7.3%) peptides bound three DP molecules, 30 (7.1%) bound four DP molecules, and 19 (4.5%) bound all five DP molecules. Altogether, 80 (18.7%) peptides were promiscuous DP-binding peptides, binding three or more DP molecules. This incidence of promiscuity is appreciably higher than would be expected.
The shared specificities of these DP molecules was also demonstrated by measuring the degree of allele-to-allele cross-reactivity, defined as the fraction of peptides binding a specific DP allele that also bind a second DP allele (Table VIII). Overall, 35–87%(mean, 57.6%) of the peptides that bind one DP molecule also bind any given second DP molecule. The greatest cross-reactivity was seen between DPB1*0401 and DPB1*0201; for which 86.7% of the DPB1*0401-binding peptides also bound DPB1*0201. In general, the lowest cross-reactivities were found for DPB1*0501; an average of only 40% of the peptides binding one DP type also bound DPB1*0501.
Table VIII.
Extent of cross-reactivity between HLA-DP molecules with a set of nonbiased peptides spanning several P. pratense Ags
| Molecule | Binders (n)a | Binders (%) | Cross-Reactivity With (%):b |
|||||
|---|---|---|---|---|---|---|---|---|
| DPB1*0101 | DPB1*0201 | DPB1*0401 | DPB1*0402 | DPB1*0501 | Average | |||
| DPB1*0101 | 98 | 23.1 | — | 70.4 | 54.1 | 68.4 | 44.9 | 59.4 |
| DPB1*0201 | 109 | 25.6 | 63.3 | — | 59.6 | 47.7 | 34.9 | 51.4 |
| DPB1*0401 | 75 | 17.6 | 70.7 | 86.7 | — | 53.3 | 38.7 | 62.3 |
| DPB1*0402 | 95 | 22.4 | 70.5 | 54.7 | 42.1 | — | 43.2 | 52.6 |
| DPB1*0501 | 61 | 14.4 | 72.1 | 62.3 | 47.5 | 67.2 | — | 62.3 |
| Average | 20.6 | 69.2 | 68.5 | 50.8 | 59.2 | 40.4 | 57.6 | |
Peptides with IC50 ≤ 1000 nM.
Cross-reactivity defined as the fraction of peptides binding the allele heading the row that also bind the allele heading the column.
Repertoire overlap, defined as the fraction of peptides that bind either of two molecules in a pair that bind both molecules, is a second method for assessing similarities in binding specificity (Table IX). Overall, on average, ~40% of the repertoires of any pair of DP alleles studied overlap.
Table IX.
Extent of repertoire overlap between HLA-DP alleles with a set of nonbiased peptides spanning several P. pratense Ags
| Molecule | Binders (n)a | Binders (%) | Repertoire Overlap With (%):b |
|||||
|---|---|---|---|---|---|---|---|---|
| DPB1*0101 | DPB1*0201 | DPB1*0401 | DPB1*0402 | DPB1*0501 | Average | |||
| DPB1*0101 | 98 | 23.1 | — | 50.0 | 44.2 | 53.2 | 38.3 | 46.4 |
| DPB1*0201 | 109 | 25.6 | 50.0 | — | 54.6 | 34.2 | 28.8 | 41.9 |
| DPB1*0401 | 75 | 17.6 | 44.2 | 54.6 | — | 30.8 | 27.1 | 39.2 |
| DPB1*0402 | 95 | 22.4 | 53.2 | 34.2 | 30.8 | — | 35.7 | 38.5 |
| DPB1*0501 | 61 | 14.4 | 38.3 | 28.8 | 27.1 | 35.7 | — | 32.5 |
| Average | 20.6 | 46.4 | 41.9 | 39.2 | 38.5 | 32.5 | 39.7 | |
Peptides with IC50 ≤ 1000 nM.
Repertoire overlap is defined as the fraction of peptides binding either allele that bind both alleles.
Table X presents a list of the peptides binding four or more of the DP molecules with an affinity ≥1000 nM. The inferred main anchors, matching the DP supermotif from Table VII in nature and spacing, are highlighted in bold type. Overall, 34 of 49 (69.4%) of the promiscuous peptides carry the proposed supermotif (Fig. 3). Indeed, of 159 peptides carrying the DP supermotif, 53 (33.3%) bound more than two DP molecules; by contrast, only 27 of 266 (10.2%) peptides that did not carry the supermotif bound three or more DP molecules.
Table X.
Promiscuous HLA-DP binding peptides derived from P. pratense Ags
| Peptide Sequencea |
P. pratense Ag | Position | HLA-DP Binding Capacity (IC50 nM) |
DP Alleles Bound (n) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N Flank | P1 | Mid | P6 | C Flank | DPB1*0101 | DPB1*0201 | DPB1*0401 | DPB1*0402 | DPB1*0501 | |||
| V | WGQK | Y | FKGNFERLA | 4 | 486 | 62 | 7.6 | 15 | 202 | 338 | 5 | |
| DKFT | V | FEAA | F | NDAIK | 5 | 211 | 130 | 24 | 543 | 640 | 561 | 5 |
| APE | V | KYTV | F | ETALKK | 5 | 256 | 16 | 5.8 | 8.7 | 50 | 12 | 5 |
| Y | FKGN | F | ERLAITKGK | 4 | 491 | 348 | 281 | 329 | 81 | 313 | 5 | |
| MAVHQ | Y | TVAL | F | LAVA | 5 | 1 | 473 | 18 | 278 | 344 | 47 | 5 |
| A | Y | ESYK | F | IPALEAAV | 5 | 231 | 100 | 225 | 649 | 115 | 377 | 5 |
| IAF | F | RKEP | L | KECGGI | 11 | 116 | 84 | 49 | 61 | 252 | 295 | 5 |
| GAQLGE | L | YYAI | Y | KAS | 4 | 151 | 21 | 6.7 | 19 | 601 | 126 | 5 |
| E | L | YYAI | Y | KASPTLAF | 4 | 156 | 264 | 63 | 129 | 782 | 347 | 5 |
| RKGV | L | FNIQ | Y | VNYWF | 4 | 416 | 118 | 8.4 | 445 | 98 | 357 | 5 |
| Y | EAFV | L | HFSEALRII | 6 | 111 | 4.9 | 0.45 | 8.5 | 4.3 | 54 | 5 | |
| IEGITL | L | NAKF | F | HMN | 13 | 156 | 328 | 117 | 402 | 13 | 555 | 5 |
| LAK | Y | KANW | I | EIMRIK | 13 | 96 | 133 | 158 | 496 | 18 | 79 | 5 |
| CDDAL | I | EGIT | L | LNAK | 13 | 151 | 246 | 58 | 74 | 5.5 | 917 | 5 |
| KL | R | SAGE | L | ELQFRRV | 1 | 146 | 546 | 312 | 630 | 557 | 17 | 5 |
| GVWT | F | DSEE | P | LQGPF | 2 | 76 | 87 | 91 | 313 | 105 | 229 | 5 |
| AAKED | F | LGCL | V | KEIP | 4 | 31 | 27 | 58 | 33 | 256 | 499 | 5 |
| FLGCL | V | KEIP | P | RLLY | 4 | 36 | 237 | 712 | 754 | 626 | 174 | 5 |
| RTFVAT | F | GAAS | N | KAF | 5 | 91 | 714 | 130 | 103 | 785 | 492 | 5 |
| VVLFAV | F | LGSA | Y | GIP | 1 | 11 | 370 | 12 | 442 | 414 | 17,604 | 4 |
| GGN | F | AGGG | F | GMLLRK | 4 | 181 | 337 | 73 | 39 | 414 | 1,802 | 4 |
| QP | F | PKTV | W | EQILNTW | 4 | 371 | 71 | 267 | 150 | 3.9 | 1,519 | 4 |
| MAAHK | F | MVAM | F | LAVA | 6 | 1 | 11 | 15 | 6.9 | 6.7 | 7,881 | 4 |
| ARVT | V | KDVT | F | RNITG | 13 | 321 | 251 | 38 | 849 | 2,349 | 403 | 4 |
| LQPET | F | AVVD | L | NKMR | 4 | 121 | 106 | 595 | 171 | 14 | 1,076 | 4 |
| AT | F | EAMY | L | GTCKTLT | 4 | 296 | 719 | 1,482 | 115 | 560 | 771 | 4 |
| MSWQTY | V | DEHL | M | CEI | 12 | 1 | 122 | 45 | 164 | 89 | 5,482 | 4 |
| IPKGD | F | LTGP | L | NFTG | 13 | 61 | 417 | 298 | 3,119 | 108 | 299 | 4 |
| LVLD | F | CDDA | L | IEGIT | 13 | 146 | 411 | 177 | 959 | 86 | 9,713 | 4 |
| LSYRS | L | QPET | F | AVVD | 4 | 116 | 202 | 14 | 14 | 2,744 | 226 | 4 |
| VA | W | QVKL | L | PVPPTVT | 4 | 241 | 24 | 234 | 482 | 22 | 9,874 | 4 |
| CKTLTP | L | MSSK | F | PEL | 4 | 306 | 6.6 | 5.3 | 0.34 | 36 | 1,011 | 4 |
| SW | I | QSIP | F | VHLGHRD | 4 | 331 | 6.3 | 8.7 | 1.8 | 2.7 | 1,325 | 4 |
| GDGF | I | DFNE | F | ISFCN | 7 | 51 | 259 | 7.8 | 79 | 2,612 | 62 | 4 |
| N | Y | NCKI | L | PNTLVLDF | 13 | 136 | 304 | 365 | 1,391 | 357 | 342 | 4 |
| SKLT | Y | ENVK | M | EDVGY | 13 | 286 | 642 | 12 | 250 | 35 | 15,281 | 4 |
| VSEALR | I | IAGT | L | EVH | 5 | 156 | 240 | 98 | 569 | 208 | 5,487 | 4 |
| R | I | IAGT | L | EVHAVKPA | 5 | 161 | 404 | 248 | 477 | 63 | 2,413 | 4 |
| N | M | VVER | L | GDYLVEQG | 12 | 116 | 134 | 150 | 1,311 | 183 | 882 | 4 |
| SFGI | V | VAWQ | V | KLLPV | 4 | 236 | 0.22 | 22 | 223 | 0.096 | 1,742 | 4 |
| AA | P | ANDK | F | TVFEAAF | 5 | 206 | 262 | 16 | 87 | 4,696 | 362 | 4 |
| AT | V | ATAP | E | VKYTVFE | 5 | 251 | 791 | 53 | 416 | 11,126 | 286 | 4 |
| KYM | V | IQGE | P | GAVIRG | 12 | 71 | 103 | 13 | 773 | 317 | 7,118 | 4 |
| KGSNPN | Y | LALL | V | KYV | 1 | 176 | 505 | 357 | 2,906 | 610 | 598 | 4 |
| MYLGTC | K | TLTP | L | MSS | 4 | 301 | 114 | 1,991 | 612 | 33 | 968 | 4 |
| PADKYR | T | FVAT | F | GAA | 5 | 86 | 214 | 33 | 129 | 3,867 | 340 | 4 |
| KYKT | F | EAAF | T | VSSKR | 6 | 61 | 88 | 358 | 178 | 1,037 | 108 | 4 |
| KQQGIR | Y | ANPI | A | FFR | 11 | 106 | 248 | 492 | 156 | 1,527 | 49 | 4 |
| RYA | N | PIAF | F | RKEPLK | 11 | 111 | 121 | 399 | 4,667 | 42 | 14 | 4 |
Residues at P1 and P6 that match the putative HLA-DP supermotif defined in Table VII are in bold type.
FIGURE 3.

Frequency of peptides bearing the canonical DP supermotif is high among DP epitopes and promiscuous binding peptides. The percentage of peptides within various sets of peptides that bear the DP supermotif as defined herein is shown. The majority (≥69.4%) of DP-binding peptides bear the supermotif compared with a minority (22.4%) of DP nonbinders.
In conclusion, a significant overlap is present among the peptidebinding repertoire of the five DP molecules considered, and peptides binding these multiple DP molecules are associated with a specific DP supermotif.
Discussion
We studied the peptide-binding specificity of the five most common DPB1 HLA allelic variants. These alleles were found to share remarkable similarities in their associated binding specificity. The extent of similarity in binding specificity observed is surprising and goes well beyond the repertoire overlap expected based on the available data. Previous observations indicated that most HLADRB1 allelic variants have similar supertype specificity (15), and Malliere and coworkers demonstrated that DPB1*0401 and DPB1*0402 share a similar supertypic specificity (47, 48). The present results confirmed the existence of a hypothesized DP supertype (47) and demonstrate that it extends to at least three additional common DP alleles. These alleles are found in >90% of the human population, and the degree of cross-reactivity determined using a panel of known DP epitopes exceeds that seen for closely related DR and DQ molecules (15, 26, 64).
These observations were enabled by the development and validation of high-throughput assays for the five molecules analyzed. Overall, each of the DP molecules tested bound between 14% and 26% of the peptides tested at the 1000-nM level. These levels of binding are consistent, in terms of frequency and affinity, with other HLAclass II molecules encoded in theDRlocus (13, 15, 64, 65). We anticipate that these assays will be useful tools for epitope-identification studies. In addition, these assays will enable the generation of the large amount of binding data required for the development of bioinformatic predictions of peptide binders for these molecules.
The biochemical assays that we established and reported herein were validated at the biological level by demonstrating that known epitopes and natural ligands bound to the relevant DPB1 allelic variant. About 90% of known DP-restricted epitopes bound the respective molecule with an affinity ≥1000 nM. The binding rates and affinities detected are similar to those described for HLA-DR epitopes (15) and suggest that 1000 nM is a generally applicable threshold for biologically relevant binding in the context of HLA class II.
On average, ~40% of the repertoires of any pair of the DP alleles studied overlap. This level of cross-reactivity is even higher than what was noted among most HLA-DR supertype specificities (J. Sidney and A. Sette, unpublished observations). The biological relevance of the binding overlap is further emphasized by the fact that ~78% of the known DP epitopes were found to bear the DP supermotif defined herein. Also, remarkably, 89% of these epitopes were promiscuous DP binders, having the capacity to bind three or more of the DP molecules tested; 74% bound four or more.
The basic DPB1 supertype is associated with a canonical P1–P6 main anchor spacing. The primary specificity at these positions involves the presence of aromatic/hydrophobic residues. This is reminiscent of DRB1 molecules, in which a strong preference for an aromatic or hydrophobic residue is seen in P1 for most of the molecules studied. However, the requirement for an aromatic/hydrophobic residue at P6 is novel and unique to DP molecules, because DRB1 molecules are usually associated with a small or charged residue in this position.
Although all five molecules studied shared a common P1–P6 main anchor spacing and motif, each was associated with its own unique specificity. In particular, in two cases, effects were detected in position N-2, presumably outside of the main binding core. Crystal structures of DP–ligand complexes are required to explain these effects and firmly establish the spatial relationship of the proposed anchor positions with the binding cleft of the DP molecules.
However, in the absence of such structures, it is still possible to speculate on the reasons for the striking degree of similarity of binding specificity between the molecules studied herein and the corresponding high incidence of promiscuous binding. Analysis of the primary sequences of DPB1 alleles reveals that, compared with DRB1 and DQB1, polymorphism is some what limited. Indeed, based on the DR/DP pocket residues defined previously (47, 55, 66), it seems that the main P6 pocket structure of all of the DP molecules studied herein are identical. The P4, P7, and P9 pockets are also, if not identical, largely conserved in most cases in terms of chemical specificity. Interestingly, the corresponding P1 pockets (and nearby residues) of the set of molecules we studied seem to separate into two types. These sets are represented by DPB1*0201, DPB1*0401, and DPB1*0402 on the one hand and DPB1*0101 and DPB1*0501 on the other, reflecting polymorphism in positions 84, 85, 86, and 87. Although the resulting overall specificity of these molecules was similar at P1, the presence of the negatively charged residues D and E in positions 84 and 85 of DPB1*0101 and DPB1*0501 may explain why these molecules tolerated positively charged residues in the corresponding position of their ligands and why these residues were deleterious for DPB1*0201, DPB1*0401, and DPB1*0402.
From the motifs defined in the current study, it seems that the P10 side chain does not play the same role in DP as it does in DR (67), despite the fact that the residues hypothesized to be involved in this interaction are largely conserved between DRB1 and DPB1. The reason for this is not possible to rationalize on the basis of our data. It is possible that the orientation of the DP binding groove makes this interaction less possible, or perhaps the energy of binding imparted by the large hydrophobic P1 and P6 main anchors renders the P10 interaction less crucial. Again, the availability of DP crystal structures would be very helpful in elucidating these physical chemical mechanisms of DP binding.
The majority of DR molecules are associated with the main DR supertypic specificity. However, it was also demonstrated that a less common supertype, associated with a radically different DRB1*0301- like binding motif/specificity, exists. For DP molecules, all of the most common alleles investigated herein were found to share a similar specificity. Given the relatively low level of polymorphism at the DPB1 locus, compared with DRB1 and DQB1, the degree of similarity in binding specificity observed might not be surprising. However, because DP molecules do play important roles in Ag recognition in the context of allergy, infectious disease, and autoimmunity, demonstration of this characteristic is of importance for diagnostic and epitope-discovery studies. At the sametime, it is possible that less commonly encountered molecules might be associated with a rather different specificity. Indeed, Dong et al. (34) proposed a motif for DP9 molecules that specifies the presence of positively charged residues in position 1. In our own unpublished studies, we identified several epitopes restricted by DP alleles other than the ones studied herein that do not bind any of the five alleles that we have characterized. One of these alleles (DP14) shares with DP9 the presence of histidine in position 9 and leucine in position 11, hypothesized to form part of the P9 and P6 pockets, respectively. Neither of these residues is present in any of the DP molecules characterized in our study, nor are they chemically conserved with the corresponding residues. Thus, it is possible that these molecules, as well as several others that share this L/H P6-P9 polymorphism, define a second DP supertype. Studies to examine the specificity of these molecules are being undertaken in our laboratory.
It is also likely that several additional DP molecules might eventually be shown to share this supertype specificity, based on the fact that the five molecules studied herein were chosen solely on the basis of their frequency. These observations raise the question of whether this supertype specificity is reflective of common ancestry or independent evolution. Indeed, it was demonstrated that both mechanisms play a role in the origin and maintenance of class I supertypes. Future studies will address this point in more detail, also in relation to the binding specificities associated with HLA-DR and -DQ molecules.
The observation by Malliere and coworkers that the primary anchor specificity of DP molecules exhibits features also found in most DR molecules (47) is supported by the current study. Taken together, these data suggest the possibility that epitopes may be identified that exhibit interlocus specificity, as well as intralocus cross-reactivity. Indeed, it has long been known that certain epitopes are capable of being recognized in the context of multiple DR, DQ, or DP types and even across loci (68-81). These observations include epitopes associated with common allergens, such as bee venom (82, 83) and the rye grass lol p5a Ag (84), for which recognition across multiple DQ or DP types and loci has been reported.
It is also possible that instances of epitopes exhibiting intralocus or interlocus cross-reactivity may not always be due to the presence of a single promiscuous core region, but rather may be caused by multiple overlapping core regions, each binding different HLA molecules. This type of cross-reactivity might be related to the previously reported phenomenon of epitope/motif clustering, or “hot spots” (36, 85-90). It was reported that certain protein regions are targets of T cells restricted by multiple HLA specificities that recognize overlapping, yet distinct, peptides. It was hypothesized that this clustering occurs significantly more frequently than would be expected by random chance. The available data suggest that it may be possible to identify highly promiscuous Th cell epitopes capable of mediating the activation of T cells restricted by multiple HLA molecules within a locus (DR, DQ, or DP) or even across multiple loci.
In conclusion, the current article described assays and peptide-binding motifs for the most common DPB1 molecules. These motifs are in general agreement with those described previously for DPB1*0201, DPB1*0401, and DPB1*0402 (45-48) but also represent more detailed quantification of the specificity of these alleles. To the best of our knowledge, the motifs for DPB1*0101 and DPB1*0501 are novel. Our experiments revealed an unexpected high degree of overlap between the peptide-binding repertoires of these molecules and led to the definition of a broad DP supertype. We anticipate that the data presented herein will facilitate molecular studies that identify and investigate, in more detail, the specificity of DP-restricted epitopes, which, despite their contribution to HLA class II responses, have received much less attention than their DR-restricted counterparts.
Supplementary Material
Acknowledgments
We thank Carla Oseroff and Howard Grey for helpful discussions.
This work was supported with funds from National Institute of Allergy and Infectious Diseases National Institutes of Health contracts N01-AI-40023, N01-AI-40024, HHSN266200400006C, and N01-700048C (all to A.S.).
Abbreviations in this paper
- ARB
average relative binding affinity
- dbMHC
MHC database
- NCBI
National Center for Biotechnology Information
- RT
room temperature
- SAAS
single amino acid substitution
- SF
specificity factor
Footnotes
Disclosures
The authors have no financial conflicts of interest.
The online version of this article contains supplemental material.
References
- 1.Doherty PC, Zinkernagel RM. A biological role for the major histocompatibility antigens. Lancet. 1975;1:1406–1409. doi: 10.1016/s0140-6736(75)92610-0. [DOI] [PubMed] [Google Scholar]
- 2.Zinkernagel RM, Doherty PC. H-2 compatability requirement for T-cell-mediated lysis of target cells infected with lymphocytic choriomeningitis virus. Different cytotoxic T-cell specificities are associated with structures coded for in H-2K or H-2D. J Exp Med. 1975;141:1427–1436. doi: 10.1084/jem.141.6.1427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Doherty PC, Zinkernagel RM. H-2 compatibility is required for T-cell-mediated lysis of target cells infected with lymphocytic choriomeningitis virus. J Exp Med. 1975;141:502–507. doi: 10.1084/jem.141.2.502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Zinkernagel RM, Doherty PC. The discovery of MHC restriction. Immunol Today. 1997;18:14–17. doi: 10.1016/s0167-5699(97)80008-4. [DOI] [PubMed] [Google Scholar]
- 5.Klein J. Natural History of the Major Histocompatibility Complex. Wiley; New York: 1986. [Google Scholar]
- 6.Hughes AL, Nei M. Maintenance of MHC polymorphism. Nature. 1992;355:402–403. doi: 10.1038/355402b0. [DOI] [PubMed] [Google Scholar]
- 7.Ferbas J, Kaplan AH, Hausner MA, Hultin LE, Matud JL, Liu Z, Panicali DL, Nerng-Ho H, Detels R, Giorgi JV. Virus burden in long-term survivors of human immunodeficiency virus (HIV) infection is a determinant of anti-HIV CD8+ lymphocyte activity. J Infect Dis. 1995;172:329–339. doi: 10.1093/infdis/172.2.329. [DOI] [PubMed] [Google Scholar]
- 8.Cao Y, Qin L, Zhang L, Safrit J, Ho DD. Virologic and immunologic characterization of long-term survivors of human immunodeficiency virus type 1 infection. N Engl J Med. 1995;332:201–208. doi: 10.1056/NEJM199501263320401. [DOI] [PubMed] [Google Scholar]
- 9.Sette A, Sidney J. Nine major HLA class I supertypes account for the vast preponderance of HLA-A and -B polymorphism. Immunogenetics. 1999;50:201–212. doi: 10.1007/s002510050594. [DOI] [PubMed] [Google Scholar]
- 10.Sette A, Sidney J. HLA supertypes and supermotifs: a functional perspective on HLA polymorphism. Curr Opin Immunol. 1998;10:478–482. doi: 10.1016/s0952-7915(98)80124-6. [DOI] [PubMed] [Google Scholar]
- 11.Sidney J, Peters B, Frahm N, Brander C, Sette A. HLA class I supertypes: a revised and updated classification. BMC Immunol. 2008;9:1. doi: 10.1186/1471-2172-9-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lund O, Nielsen M, Kesmir C, Petersen AG, Lundegaard C, Worning P, Sylvester-Hvid C, Lamberth K, Røder G, Justesen S, et al. Definition of supertypes for HLA molecules using clustering of specificity matrices. Immunogenetics. 2004;55:797–810. doi: 10.1007/s00251-004-0647-4. [DOI] [PubMed] [Google Scholar]
- 13.Doolan DL, Southwood S, Chesnut R, Appella E, Gomez E, Richards A, Higashimoto YI, Maewal A, Sidney J, Gramzinski RA, et al. HLADR- promiscuous T cell epitopes from Plasmodium falciparum pre-erythrocyticstage antigens restricted by multiple HLA class II alleles. J Immunol. 2000;165:1123–1137. doi: 10.4049/jimmunol.165.2.1123. [DOI] [PubMed] [Google Scholar]
- 14.Wilson CC, Palmer B, Southwood S, Sidney J, Higashimoto Y, Appella E, Chesnut R, Sette A, Livingston BD. Identification and antigenicity of broadly cross-reactive and conserved human immunodeficiency virus type 1-derived helper T-lymphocyte epitopes. J Virol. 2001;75:4195–4207. doi: 10.1128/JVI.75.9.4195-4207.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Southwood S, Sidney J, Kondo A, del Guercio MF, Appella E, Hoffman S, Kubo RT, Chesnut RW, Grey HM, Sette A. Several common HLA-DR types share largely overlapping peptide binding repertoires. J Immunol. 1998;160:3363–3373. [PubMed] [Google Scholar]
- 16.Chelvanayagam G. A roadmap for HLA-DR peptide binding specificities. Hum Immunol. 1997;58:61–69. doi: 10.1016/s0198-8859(97)00185-7. [DOI] [PubMed] [Google Scholar]
- 17.Doytchinova IA, Flower DR. In silico identification of supertypes for class II MHCs. J Immunol. 2005;174:7085–7095. doi: 10.4049/jimmunol.174.11.7085. [DOI] [PubMed] [Google Scholar]
- 18.Gibert M, Balandraud N, Touinssi M, Mercier P, Roudier J, Reviron D. Functional categorization of HLA-DRB1 alleles in rheumatoid arthritis: the protective effect. Hum Immunol. 2003;64:930–935. doi: 10.1016/s0198-8859(03)00186-1. [DOI] [PubMed] [Google Scholar]
- 19.Gibert M, Sanchez-Mazas A. Geographic patterns of functional categories of HLA-DRB1 alleles: a new approach to analyse associations between HLA-DRB1 and disease. Eur J Immunogenet. 2003;30:361–374. doi: 10.1046/j.1365-2370.2003.00418.x. [DOI] [PubMed] [Google Scholar]
- 20.Ou D, Mitchell LA, Décarie D, Tingle AJ, Nepom GT. Promiscuous T-cell recognition of a rubella capsid protein epitope restricted by DRB1*0403 and DRB1*0901 molecules sharing an HLA DR supertype. Hum Immunol. 1998;59:149–157. doi: 10.1016/s0198-8859(98)00006-8. [DOI] [PubMed] [Google Scholar]
- 21.Ou D, Mitchell LA, Tingle AJ. HLA-DR restrictive supertypes dominate promiscuous T cell recognition: association of multiple HLA-DR molecules with susceptibility to autoimmune diseases. J Rheumatol. 1997;24:253– 261. [PubMed] [Google Scholar]
- 22.Ou D, Mitchell LA, Tingle AJ. A new categorization of HLA DR alleles on a functional basis. Hum Immunol. 1998;59:665–676. doi: 10.1016/s0198-8859(98)00067-6. [DOI] [PubMed] [Google Scholar]
- 23.Hammer J, Valsasnini P, Tolba K, Bolin D, Higelin J, Takacs B, Sinigaglia F. Promiscuous and allele-specific anchors in HLA-DR-binding peptides. Cell. 1993;74:197–203. doi: 10.1016/0092-8674(93)90306-b. [DOI] [PubMed] [Google Scholar]
- 24.Raddrizzani L, Sturniolo T, Guenot J, Bono E, Gallazzi F, Nagy ZA, Sinigaglia F, Hammer J. Different modes of peptide interaction enable HLA-DQ and HLA-DR molecules to bind diverse peptide repertoires. J Immunol. 1997;159:703–711. [PubMed] [Google Scholar]
- 25.Sinigaglia F, Hammer J. Motifs and supermotifs for MHC class II binding peptides. J Exp Med. 1995;181:449–451. doi: 10.1084/jem.181.2.449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sidney J, del Guercio MF, Southwood S, Sette A. The HLA molecules DQA1*0501/B1*0201 and DQA1*0301/B1*0302 share an extensive overlap in peptide binding specificity. J Immunol. 2002;169:5098–5108. doi: 10.4049/jimmunol.169.9.5098. [DOI] [PubMed] [Google Scholar]
- 27.Peters B, Sidney J, Bourne P, Bui HH, Buus S, Doh G, Fleri W, Kronenberg M, Kubo R, Lund O, et al. The immune epitope database and analysis resource: from vision to blueprint. PLoS Biol. 2005;3:e91. doi: 10.1371/journal.pbio.0030091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Edwards JA, Durant BM, Jones DB, Evans PR, Smith JL. Differential expression of HLA class II antigens in fetal human spleen: relationship of HLA-DP, DQ, and DR to immunoglobulin expression. J Immunol. 1986;137:490–497. [PubMed] [Google Scholar]
- 29.Hauber I, Gulle H, Wolf HM, Maris M, Eggenbauer H, Eibl MM. Molecular characterization of major histocompatibility complex class II gene expression and demonstration of antigen-specific T cell response indicate a new phenotype in class II-deficient patients. J Exp Med. 1995;181:1411–1423. doi: 10.1084/jem.181.4.1411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Guardiola J, Maffei A. Control of MHC class II gene expression in autoimmune, infectious, and neoplastic diseases. Crit Rev Immunol. 1993;13:247–268. [PubMed] [Google Scholar]
- 31.Agrewala JN, Wilkinson RJ. Differential regulation of Th1 and Th2 cells by p91-110 and p21-40 peptides of the 16-kD alpha-crystallin antigen of Mycobacterium tuberculosis. Clin Exp Immunol. 1998;114:392–397. doi: 10.1046/j.1365-2249.1998.00724.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Caccamo N, Barera A, Di Sano C, Meraviglia S, Ivanyi J, Hudecz F, Bosze S, Dieli F, Salerno A. Cytokine profile, HLA restriction and TCR sequence analysis of human CD4+ T clones specific for an immunodominant epitope of Mycobacterium tuberculosis 16-kDa protein. Clin Exp Immunol. 2003;133:260–266. doi: 10.1046/j.1365-2249.2003.02201.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.de Waal L, Yüksel S, Brandenburg AH, Langedijk JP, Sintnicolaas K, Verjans GM, Osterhaus AD, de Swart RL. Identification of a common HLA-DP4-restricted T-cell epitope in the conserved region of the respiratory syncytial virus G protein. J Virol. 2004;78:1775–1781. doi: 10.1128/JVI.78.4.1775-1781.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Dong RP, Kamikawaji N, Toida N, Fujita Y, Kimura A, Sasazuki T. Characterization of T cell epitopes restricted by HLA-DP9 in streptococcal M12 protein. J Immunol. 1995;154:4536–4545. [PubMed] [Google Scholar]
- 35.Fossum B, Gedde-Dahl T, 3rd, Hansen T, Eriksen JA, Thorsby E, Gaudernack G. Overlapping epitopes encompassing a point mutation (12 Gly—>Arg) in p21 ras can be recognized by HLA-DR, -DP and -DQ restricted T cells. Eur J Immunol. 1993;23:2687–2691. doi: 10.1002/eji.1830231045. [DOI] [PubMed] [Google Scholar]
- 36.Higgins JA, Thorpe CJ, Hayball JD, O’Hehir RE, Lamb JR. Overlapping T-cell epitopes in the group I allergen of Dermatophagoides species restricted by HLA-DP and HLA-DR class II molecules. J Allergy Clin Immunol. 1994;93:891–899. doi: 10.1016/0091-6749(94)90383-2. [DOI] [PubMed] [Google Scholar]
- 37.Kurane I, Dai LC, Livingston PG, Reed E, Ennis FA. Definition of an HLA-DPw2-restricted epitope on NS3, recognized by a dengue virus serotype-cross-reactive human CD4+ CD8− cytotoxic T-cell clone. J Virol. 1993;67:6285–6288. doi: 10.1128/jvi.67.10.6285-6288.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mandic M, Castelli F, Janjic B, Almunia C, Andrade P, Gillet D, Brusic V, Kirkwood JM, Maillere B, Zarour HM. One NY-ESO-1-derived epitope that promiscuously binds to multiple HLA-DR and HLA-DP4 molecules and stimulates autologous CD4+ T cells from patients with NY-ESO-1- expressing melanoma. J Immunol. 2005;174:1751–1759. doi: 10.4049/jimmunol.174.3.1751. [DOI] [PubMed] [Google Scholar]
- 39.Okamoto YI, Kurane AM, Leporati AM, Ennis FA. Definition of the region on NS3 which contains multiple epitopes recognized by dengue virus serotype- cross-reactive and flavivirus-cross-reactive, HLA-DPw2-restricted CD4+ T cell clones. J Gen Virol. 1998;79:697–704. doi: 10.1099/0022-1317-79-4-697. [DOI] [PubMed] [Google Scholar]
- 40.Qian F, Gnjatic S, Jäger E, Santiago D, Jungbluth A, Grande C, Schneider S, Keitz B, Driscoll D, Ritter G, et al. Th1/Th2 CD4+ T cell responses against NY-ESO-1 in HLA-DPB1*0401/0402 patients with epithelial ovarian cancer. Cancer Immun. 2004;4:12. [PubMed] [Google Scholar]
- 41.Tang J, Olive M, Champagne K, Flomenberg N, Eisenlohr L, Hsu S, Flomenberg P. Adenovirus hexon T-cell epitope is recognized by most adults and is restricted by HLA DP4, the most common class II allele. Gene Ther. 2004;11:1408–1415. doi: 10.1038/sj.gt.3302316. [DOI] [PubMed] [Google Scholar]
- 42.Voo KS, Fu T, Heslop HE, Brenner MK, Rooney CM, Wang RF. Identification of HLA-DP3-restricted peptides from EBNA1 recognized by CD4+ T cells. Cancer Res. 2002;62:7195–7199. [PubMed] [Google Scholar]
- 43.Yang J, Huston L, Berger D, Danke NA, Liu AW, Disis ML, Kwok WW. Expression of HLA-DP0401 molecules for identification of DP0401 restricted antigen specific T cells. J Clin Immunol. 2005;25:428–436. doi: 10.1007/s10875-005-6095-6. [DOI] [PubMed] [Google Scholar]
- 44.Zeng G, Wang X, Robbins PF, Rosenberg SA, Wang RF. CD4+ T cell recognition of MHC class II-restricted epitopes from NY-ESO-1 presented by a prevalent HLA DP4 allele: association with NY-ESO-1 antibody production. Proc Natl Acad Sci USA. 2001;98:3964–3969. doi: 10.1073/pnas.061507398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Díaz G, Canñas B, Vazquez J, Nombela C, Arroyo J. Characterization of natural peptide ligands from HLA-DP2: new insights into HLA-DP peptide-binding motifs. Immunogenetics. 2005;56:754–759. doi: 10.1007/s00251-004-0735-5. [DOI] [PubMed] [Google Scholar]
- 46.Chicz RM, Graziano DF, Trucco M, Strominger JL, Gorga JC. HLA-DP2: self peptide sequences and binding properties. J Immunol. 1997;159:4935–4942. [PubMed] [Google Scholar]
- 47.Castelli FA, Buhot C, Sanson A, Zarour H, Pouvelle-Moratille S, Nonn C, Gahery-Ségard H, Guillet JG, Ménez A, Georges B, Maillére B. HLA-DP4, the most frequent HLA II molecule, defines a new supertype of peptide-binding specificity. J Immunol. 2002;169:6928–6934. doi: 10.4049/jimmunol.169.12.6928. [DOI] [PubMed] [Google Scholar]
- 48.Cohen WM, Pouvelle-Moratille S, Wang XF, Farci S, Munier G, Charron D, Meénez A, Busson M, Maillère B. Scanning the HIV genome for CD4+ T cell epitopes restricted to HLA-DP4, the most prevalent HLA class II molecule. J Immunol. 2006;176:5401–5408. doi: 10.4049/jimmunol.176.9.5401. [DOI] [PubMed] [Google Scholar]
- 49.Sidney J, Southwood S, Oseroff C, Del Guercio MF, Sette A, Grey H. Measurement of MHC/peptide interactions by gel filtration. Curr Protoc Immunol. 1998;26:18.3.1–18.3.19. doi: 10.1002/0471142735.im1803s31. [DOI] [PubMed] [Google Scholar]
- 50.Gulukota K, Sidney J, Sette A, DeLisi C. Two complementary methods for predicting peptides binding major histocompatibility complex molecules. J Mol Biol. 1997;267:1258–1267. doi: 10.1006/jmbi.1997.0937. [DOI] [PubMed] [Google Scholar]
- 51.Cheng Y, Prusoff WH. Relationship between the inhibition constant (K1) and the concentration of inhibitor which causes 50 per cent inhibition (I50) of an enzymatic reaction. Biochem Pharmacol. 1973;22:3099–3108. doi: 10.1016/0006-2952(73)90196-2. [DOI] [PubMed] [Google Scholar]
- 52.Sidney J, Grey HM, Southwood S, Celis E, Wentworth PA, del Guercio MF, Kubo RT, Chesnut RW, Sette A. Definition of an HLAA3- like supermotif demonstrates the overlapping peptide-binding repertoires of common HLA molecules. Hum Immunol. 1996;45:79–93. doi: 10.1016/0198-8859(95)00173-5. [DOI] [PubMed] [Google Scholar]
- 53.Meyer D, Singe RM, Mack SJ, Lancaster A, Nelson MP, Erlich H, Fernandez-Vina M, Thomson G. Single locus polymorphism of classical HLA genes. In: Hansen J, editor. Immunobiology of the Human MHC: Proceedings of the 13th International Histocompatibility Workshop and Conference. IHWG Press; Seattle, WA: 2007. pp. 653–704. [Google Scholar]
- 54.Ghosh P, Amaya M, Mellins E, Wiley DC. The structure of an intermediate in class II MHC maturation: CLIP bound to HLA-DR3. Nature. 1995;378:457–462. doi: 10.1038/378457a0. [DOI] [PubMed] [Google Scholar]
- 55.Stern LJ, Brown JH, Jardetzky TS, Gorga JC, Urban RG, Strominger JL, Wiley DC. Crystal structure of the human class II MHC protein HLA-DR1 complexed with an influenza virus peptide. Nature. 1994;368:215–221. doi: 10.1038/368215a0. [DOI] [PubMed] [Google Scholar]
- 56.Madden DR. The three-dimensional structure of peptide-MHC complexes. Annu Rev Immunol. 1995;13:587–622. doi: 10.1146/annurev.iy.13.040195.003103. [DOI] [PubMed] [Google Scholar]
- 57.Yun C, Senju S, Fujita H, Tsuji Y, Irie A, Matsushita S, Nishimura Y. Augmentation of immune response by altered peptide ligands of the antigenic peptide in a human CD4+ T-cell clone reacting to TEL/AML1 fusion protein. Tissue Antigens. 1999;54:153–161. doi: 10.1034/j.1399-0039.1999.540206.x. [DOI] [PubMed] [Google Scholar]
- 58.Kaufmann DE, Bailey PM, Sidney J, Wagner B, Norris PJ, Johnston MN, Cosimi LA, Addo M, Lichterfeld M, Altfeld M, et al. Comprehensive analysis of human immunodeficiency virus type 1-specific CD4 responses reveals marked immunodominance of gag and nef and the presence of broadly recognized peptides. J Virol. 2004;78:4463–4477. doi: 10.1128/JVI.78.9.4463-4477.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Sidney J, Southwood S, Oseroff C, del Guercio MF, Sette A, Grey HM. Measurement of MHC/peptide interactions by gel filtration. Curr Protoc Immunol. 2001;26:18.3.1–18.3.19. doi: 10.1002/0471142735.im1803s31. [DOI] [PubMed] [Google Scholar]
- 60.Falk K, Rötzschke O, Stevanovicć S, Jung G, Rammensee HG. Pool sequencing of natural HLA-DR, DQ, and DP ligands reveals detailed peptide motifs, constraints of processing, and general rules. Immunogenetics. 1994;39:230–242. doi: 10.1007/BF00188785. [DOI] [PubMed] [Google Scholar]
- 61.Minohara M, Ochi H, Matsushita S, Irie A, Nishimura Y, Kira J. Differences between T-cell reactivities to major myelin protein-derived peptides in opticospinal and conventional forms of multiple sclerosis and healthy controls. Tissue Antigens. 2001;57:447–456. doi: 10.1034/j.1399-0039.2001.057005447.x. [DOI] [PubMed] [Google Scholar]
- 62.Sidney J, Assarsson E, Moore C, Ngo S, Pinilla C, Sette A, Peters B. Quantitative peptide binding motifs for 19 human and mouse MHC class I molecules derived using positional scanning combinatorial peptide libraries. Immunome Res. 2008;4:2. doi: 10.1186/1745-7580-4-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Sidney J, Peters B, Moore C, Pencille TJ, Ngo S, Masterman KA, Asabe S, Pinilla C, Chisari FV, Sette A. Characterization of the peptide-binding specificity of the chimpanzee class I alleles A 0301 and A 0401 using a combinatorial peptide library. Immunogenetics. 2007;59:745–751. doi: 10.1007/s00251-007-0243-5. [DOI] [PubMed] [Google Scholar]
- 64.Sidney J, Oseroff C, Southwood S, Wall M, Ishioka G, Koning F, Sette A. DRB1*0301 molecules recognize a structural motif distinct from the one recognized by most DR beta 1 alleles. J Immunol. 1992;149:2634–2640. [PubMed] [Google Scholar]
- 65.Sette A, Sidney J, Oseroff C, del Guercio MF, Southwood S, Arrhenius T, Powell MF, Coloón SM, Gaeta FC, Grey HM. HLA DR4w4-binding motifs illustrate the biochemical basis of degeneracy and specificity in peptide-DR interactions. J Immunol. 1993;151:3163–3170. [PubMed] [Google Scholar]
- 66.Díaz G, Amicosante M, Jaraquemada D, Butler RH, Guillén MV, Sánchez M, Nombela C, Arroyo J. Functional analysis of HLA-DP polymorphism: a crucial role for DPbeta residues 9, 11, 35, 55, 56, 69 and 84-87 in T cell allorecognition and peptide binding. Int Immunol. 2003;15:565–576. doi: 10.1093/intimm/dxg057. [DOI] [PubMed] [Google Scholar]
- 67.Zavala-Ruiz Z, Strug I, Anderson MW, Gorski J, Stern LJ. A polymorphic pocket at the P10 position contributes to peptide binding specificity in class II MHC proteins. Chem Biol. 2004;11:1395–1402. doi: 10.1016/j.chembiol.2004.08.007. [DOI] [PubMed] [Google Scholar]
- 68.Roche PA, Cresswell P. High-affinity binding of an influenza hemagglutinin-derived peptide to purified HLA-DR. J Immunol. 1990;144:1849–1856. [PubMed] [Google Scholar]
- 69.Busch R, Strang G, Howland K, Rothbard JB. Degenerate binding of immunogenic peptides to HLA-DR proteins on B cell surfaces. Int Immunol. 1990;2:443–451. doi: 10.1093/intimm/2.5.443. [DOI] [PubMed] [Google Scholar]
- 70.Ceppellini R, Frumento G, Ferrara GB, Tosi R, Chersi A, Pernis B. Binding of labelled influenza matrix peptide to HLA DR in living B lymphoid cells. Nature. 1989;339:392–394. doi: 10.1038/339392a0. [DOI] [PubMed] [Google Scholar]
- 71.Panina-Bordignon P, Demotz S, Corradin G, Lanzavecchia A. Study on the immunogenicity of human class-II-restricted T-cell epitopes: processing constraints, degenerate binding, and promiscuous recognition. Cold Spring Harb Symp Quant Biol. 1989;54:445–451. doi: 10.1101/sqb.1989.054.01.053. [DOI] [PubMed] [Google Scholar]
- 72.Panina-Bordignon P, Tan A, Termijtelen A, Demotz S, Corradin G, Lanzavecchia A. Universally immunogenic T cell epitopes: promiscuous binding to human MHC class II and promiscuous recognition by T cells. Eur J Immunol. 1989;19:2237–2242. doi: 10.1002/eji.1830191209. [DOI] [PubMed] [Google Scholar]
- 73.Sinigaglia F, Guttinger M, Kilgus J, Doran DM, Matile H, Etlinger H, Trzeciak A, Gillessen D, Pink JR. A malaria T-cell epitope recognized in association with most mouse and human MHC class II molecules. Nature. 1988;336:778–780. doi: 10.1038/336778a0. [DOI] [PubMed] [Google Scholar]
- 74.O’Sullivan D, Arrhenius T, Sidney J, Del Guercio MF, Albertson M, Wall M, Oseroff C, Southwood S, Colón SM, Gaeta FC, et al. On the interaction of promiscuous antigenic peptides with different DR alleles. Identification of common structural motifs. J Immunol. 1991;147:2663–2669. [PubMed] [Google Scholar]
- 75.Hill CM, Hayball JD, Allison AA, Rothbard JB. Conformational and structural characteristics of peptides binding to HLA-DR molecules. J Immunol. 1991;147:189–197. [PubMed] [Google Scholar]
- 76.Diepolder HM, Gerlach JT, Zachoval R, Hoffmann RM, Jung MC, Wierenga EA, Scholz S, Santantonio T, Houghton M, Southwood S, et al. Immunodominant CD4+ T-cell epitope within nonstructural protein 3 in acute hepatitis C virus infection. J Virol. 1997;71:6011–6019. doi: 10.1128/jvi.71.8.6011-6019.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Alexander J, Sidney J, Southwood S, Ruppert J, Oseroff C, Maewal A, Snoke K, Serra HM, Kubo RT, Sette A, et al. Development of high potency universal DR-restricted helper epitopes by modification of high affinity DR-blocking peptides. Immunity. 1994;1:751–761. doi: 10.1016/s1074-7613(94)80017-0. [DOI] [PubMed] [Google Scholar]
- 78.Wucherpfennig KW, Sette A, Southwood S, Oseroff C, Matsui M, Strominger JL, Hafler DA. Structural requirements for binding of an immunodominant myelin basic protein peptide to DR2 isotypes and for its recognition by human T cell clones. J Exp Med. 1994;179:279–290. doi: 10.1084/jem.179.1.279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Ferrari C, Bertoletti A, Penna A, Cavalli A, Valli A, Missale G, Pilli M, Fowler P, Giuberti T, Chisari FV, et al. Identification of immunodominant T cell epitopes of the hepatitis B virus nucleocapsid antigen. J Clin Invest. 1991;88:214–222. doi: 10.1172/JCI115280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Valli A, Sette A, Kappos L, Oseroff C, Sidney J, Miescher G, Hochberger M, Albert ED, Adorini L. Binding of myelin basic protein peptides to human histocompatibility leukocyte antigen class II molecules and their recognition by T cells from multiple sclerosis patients. J Clin Invest. 1993;91:616–628. doi: 10.1172/JCI116242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Lamonaca V, Missale G, Urbani S, Pilli M, Boni C, Mori C, Sette A, Massari M, Southwood S, Bertoni R, et al. Conserved hepatitis C virus sequences are highly immunogenic for CD4+ T cells: implications for vaccine development. Hepatology. 1999;30:1088–1098. doi: 10.1002/hep.510300435. [DOI] [PubMed] [Google Scholar]
- 82.Texier C, Pouvelle S, Busson M, Hervé M, Charron D, Ménez A, Maillère B. HLA-DR restricted peptide candidates for bee venom immunotherapy. J Immunol. 2000;164:3177–3184. doi: 10.4049/jimmunol.164.6.3177. [DOI] [PubMed] [Google Scholar]
- 83.Texier C, Pouvelle-Moratille S, Buhot C, Castelli FA, Pecquet C, Ménez A, Leynadier F, Maillère B. Emerging principles for the design of promiscuous HLA-DR-restricted peptides: an example from the major bee venom allergen. Eur J Immunol. 2002;32:3699–3707. doi: 10.1002/1521-4141(200212)32:12<3699::AID-IMMU3699>3.0.CO;2-V. [DOI] [PubMed] [Google Scholar]
- 84.de Lalla C, Sturniolo T, Abbruzzese L, Hammer J, Sidoli A, Sinigaglia F, Panina-Bordignon P. Cutting edge: identification of novel T cell epitopes in Lol p5a by computational prediction. J Immunol. 1999;163:1725–1729. [PubMed] [Google Scholar]
- 85.Arentz-Hansen H, McAdam SN, Molberg O, Fleckenstein B, Lundin KE, Jørgensen TJ, Jung G, Roepstorff P, Sollid LM. Celiac lesion T cells recognize epitopes that cluster in regions of gliadins rich in proline residues. Gastroenterology. 2002;123:803–809. doi: 10.1053/gast.2002.35381. [DOI] [PubMed] [Google Scholar]
- 86.Ahlers JD, Pendleton CD, Dunlop N, Minassian A, Nara PL, Berzofsky JA. Construction of an HIV-1 peptide vaccine containing a multideterminant helper peptide linked to a V3 loop peptide 18 inducing strong neutralizing antibody responses in mice of multiple MHC haplotypes after two immunizations. J Immunol. 1993;150:5647–5665. [PubMed] [Google Scholar]
- 87.Brown SA, Lockey TD, Slaughter C, Slobod KS, Surman S, Zirkel A, Mishra A, Pagala VR, Coleclough C, Doherty PC, Hurwitz JL. T cell epitope “hotspots” on the HIV Type 1 gp120 envelope protein overlap with tryptic fragments displayed bymass spectrometry. AIDSRes Hum Retroviruses. 2005;21:165–170. doi: 10.1089/aid.2005.21.165. [DOI] [PubMed] [Google Scholar]
- 88.Gupta V, Tabiin TM, Sun K, Chandrasekaran A, Anwar A, Yang K, Chikhlikar P, Salmon J, Brusic V, Marques ET, et al. SARS coronavirus nucleocapsid immunodominant T-cell epitope cluster is common to both exogenous recombinant and endogenous DNA-encoded immunogens. Virology. 2006;347:127–139. doi: 10.1016/j.virol.2005.11.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Meinl E, Weber F, Drexler K, Morelle C, Ott M, Saruhan-Direskeneli G, Goebels N, Ertl B, Jechart G, Giegerich G, et al. Myelin basic proteinspecific T lymphocyte repertoire in multiple sclerosis. Complexity of the response and dominance of nested epitopes due to recruitment of multiple T cell clones. J Clin Invest. 1993;92:2633–2643. doi: 10.1172/JCI116879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Muller CP, Bünder R, Mayser H, Ammon S, Weinmann M, Brons NH, Schneider F, Jung G, Wiesmüller KH. Intramolecular immunodominance and intermolecular selection of H2d-restricted peptides define the same immunodominant region of the measles virus fusion protein. Mol Immunol. 1995;32:37–47. doi: 10.1016/0161-5890(94)00132-k. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.