

Abstract
The presence of the cationic phospholipid lysyl-phosphatidylglycerol (lysyl-PG) in staphylococcal cytoplasmic membranes has been linked to increased resistance to cationic compounds, including antibiotics such as daptomycin as well as host defense antimicrobial peptides. We investigated the effects of lysyl-PG on binding of 6W-RP-1, a synthetic antimicrobial peptide, to lipid vesicles and on peptide-induced membrane permeabilization. Unexpectedly, physiological lysyl-PG concentrations only minimally reduced membrane binding of 6W-RP-1. In contrast, 6W-RP-1-induced dye leakage was severely inhibited by lysyl-PG, suggesting that lysyl-PG primarily impacts membrane defect formation.
Aminoacylated phosphatidylglycerols are common lipids in the cytoplasmic membranes of Gram-positive bacteria, most notably in the genera Staphylococcus, Bacillus, Clostridium, Lactobacillus, Listeria, and Streptococcus (7, 11, 15). Esterification of the phosphatidylglycerol (PG) headgroup occurs with either lysine (lysyl-PG), alanine (alanyl-PG), or, less commonly, ornithine (ornithyl-PG) or arginine (arginyl-PG). Such modifications may serve to modulate the cytoplasmic membrane charge, as the adduct converts the anionic PG into a net cationic or zwitterionic lipid, depending on the amino acid involved (13). In Staphylococcus aureus, this reaction is mediated by the multiple peptide resistance factor F (MprF), an integral membrane protein that catalyzes the transfer of the aminoacyl group from Lys-tRNALys to the sn-3 position of the PG headgroup (14).
In S. aureus, resistance to killing by mammalian cationic antimicrobial peptides (CAPs) has been attributed to reduced peptide binding to the bacterial cell surface (16). Furthermore, resistance to daptomycin, a calcium-dependent, cationic lipopeptide, has been suggested to have coevolved with reduced susceptibility to host defense CAPs (8, 9). The presence of lysyl-PG in the bacterial membrane has been clearly linked to these phenomena in S. aureus and Listeria (8, 17). For instance, reduced levels of lysyl-PG were found to increase the susceptibility of S. aureus to many antibiotics, including some CAPs (10, 18, 19), and mutant S. aureus strains that do not produce lysyl-PG exhibit reduced virulence in a variety of in vivo models (18). Of note, the observed correlation between PG lysinylation and reduced susceptibility to cationic antibiotics (e.g., daptomycin) and CAPs has been attributed to a relative decrease in the negative surface charge of the target bacterial cytoplasmic membrane as a consequence of PG modification (8, 10, 12, 13, 18). However, no detailed analyses of the mechanism(s) by which lysyl-PG protects the organism from cationic agent-induced killing have been carried out in either whole bacterial cells or model membranes. For this reason, we employed model membrane systems composed of various PG-to-lysyl-PG ratios and the synthetic peptide 6W-RP-1 to further probe this interaction. 6W-RP-1 is a fluorescent derivative of RP-1, a synthetic peptide modeled in part on the C-terminal helical and microbicidal domain of the mammalian platelet factor 4 (PF-4) family of molecules (20).
The peptide 6W-RP-1 (ALYKKWKKKLLKSLKRLG), a tryptophan-substituted congener of the parent peptide, RP-1 (20), was synthesized and authenticated as described previously (20). Large unilamellar vesicles (LUVs) were prepared by mixing chloroform solutions of lipids in the appropriate proportions (1). To mimic the cytoplasmic membrane of S. aureus, a 70:30 mixture of 1-palmitoyl-2-oleoyl-sn-glycero-3-[phospho-rac-(1-glycerol)] (POPG) and 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) was used instead of pure POPG to increase vesicle stability. The impact of lysyl-DOPG on peptide activity was investigated by replacing various fractions of POPG with 1,2-dioleoyl-sn-glycero-3-[phospho-rac-(3-lysyl(1-glycerol))] (lysyl-DOPG). For lipid vesicles used in the binding studies, the fluorescent lipid 7MC-POPE (1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoethanolamine [POPE], derivatized with 7MC through an amide bond to the amino group of the ethanolamine headgroup) was synthesized as previously described (5) and added in a chloroform solution to the unlabeled lipids.
The association kinetics of 6W-RP-1 with lipid vesicles were measured by Förster resonance energy transfer (FRET) from the Trp residue in the peptide to the acceptor fluorophore, 7MC-POPE, embedded in the bilayer at a concentration of 2 mol% (5). Peptide binding leads to an increase in FRET and can thus be monitored through the increased fluorescence emission from the acceptor. The reaction was followed in a stopped-flow fluorimeter (SX.18MV; Applied Photophysics) as previously described (1). The peptide concentration was held constant at 0.5 μM in all FRET experiments, and the lipid concentration was varied between 25 and 200 μM.
If the initial peptide binding to lipid vesicles is well separated in time from other bilayer-perturbing events, peptide binding is described by the simple equilibrium equation
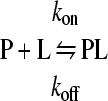 |
where P stands for free peptide in solution, L represents lipid, PL represents lipid-bound peptides, and kon and koff are the rate constants for peptide association and dissociation, respectively (1). The coupled differential equations that describe this equilibrium are d[P]/dt = −kon [L] [P] + koff [PL] d[PL]/dt = kon [L] [P] − koff [PL].
As the lipid concentration does not change throughout the experiment, the product of kon and [L] remains constant. Hence, the kinetics of peptide binding follow a pseudo-first-order rate law and are characterized by an apparent rate constant, kapp, equal to (kon [L] + koff), which can be obtained from a fit of the experimental traces to an exponential function (Fig. 1 A). Since the magnitude of kapp depends on the lipid concentration, a plot of kapp against [L] will be linear (Fig. 1B). The molecular rate constants of peptide binding, kon values, were obtained from the slope of a linear regression to the data points and koff from the y intercept. The dissociation constant, KD, was then obtained from the koff/kon ratio.
FIG. 1.

Binding of 6W-RP-1 to vesicles composed of POPG and POPC (70:30 ratio). (A) Experimental binding kinetics recorded using 50 μM lipid and 0.5 μM peptide. The experimental data are the averages of five individual traces to improve the signal-to-noise ratio; the solid line is a single exponential fit to the data. (B) The apparent rate constant (kapp) was plotted against the lipid concentration to obtain kon and koff, as explained in the text. The error bars represent the standard deviations from five sets of experiments.
We determined kapp as a function of lysyl-DOPG content and found that at a given lipid concentration, kapp varied only slightly with a lysyl-DOPG content between 0 and 30 mol% but decreased significantly at lysyl-DOPG concentrations exceeding ∼30 mol% (Fig. 2 A). By using the dependence of kapp on lipid concentration to obtain the molecular rate constants kon and koff for each lysyl-DOPG concentration, we were able to show that this effect is caused primarily by a change in koff. The rate constant for 6W-RP-1 binding to lipid vesicles varied linearly with lysyl-DOPG content (Fig. 2B). However, the dependence was relatively weak, dropping to about 60% of its original value at 40% lysyl-DOPG. In contrast, koff was found to remain essentially constant up to an overall lysyl-DOPG content of ∼30 mol% (Fig. 2C), which encompasses the lysinylation profile of most clinical S. aureus isolates (6, 12, 13). Only at a lysyl-DOPG concentration of >30 mol% did koff become notably faster, leading to a significantly reduced net association and a correspondingly higher KD (Fig. 2D). We propose that this observed lack of correlation between the lysyl-DOPG concentration and peptide binding until the lysyl-DOPG content exceeds 30 mol% is related to peptide-induced lipid domain formation that results in the separation of anionic PG from the cationic lysyl-DOPG and the remaining membrane lipids. In a homogeneous bilayer, the Gibbs free energy (ΔGo) of binding can be expected to be largely a linear function of the lysyl-DOPG fraction. In that case, we would predict the dissociation constant, KD, to depend exponentially on the fraction of lysyl-DOPG in the mixture, according to the equation KD = exp(ΔGo/RT), which it clearly did not in our studies (Fig. 2D).
FIG. 2.

6W-RP-1 interaction with lipid vesicles as a function of the lysyl-DOPG content. The vesicle composition is based on a POPG:POPC ratio of 70:30, in which a fraction of the POPG was replaced by lysyl-DOPG. (A) The apparent rate constant, kapp, as a function of lysyl-PG content, measured at a lipid concentration of 100 μM. (B to D) The molecular rate constant for peptide binding, kon (B), the molecular rate constant for 6W-RP-1 desorption, koff (C), and the dissociation constant, KD (D). The peptide concentration was 0.5 μM in all experiments. The error bars are the standard deviations for a minimum of two and usually three sets of experiments.
In contrast to the data on 6W-RP-1 binding, the extent of peptide-induced membrane permeabilization of lipid vesicles was strongly inhibited as a function of lysyl-DOPG content, even at low lysyl-DOPG concentrations. Dye leakage was assessed by encapsulating a self-quenching concentration of carboxyfluorescein (CF) in LUVs by methods described previously (1). A 5 μM peptide solution was allowed to interact with a lipid suspension, and the rate and extent of dye leakage were monitored. The kinetics of CF leakage, measured by the relief of fluorescence self-quenching, were recorded in a stopped-flow fluorimeter to ensure rapid and complete mixing of the reactants. We found that inclusion of 20 mol% lysyl-DOPG decreased both the extent and rate of dye leakage, and at 30 mol%, at which peptide binding is still largely unaffected, dye leakage was essentially abolished (Fig. 3). Although unanticipated, this finding is consistent with a biological function of lysyl-DOPG that derives in part from the inhibition of membrane perturbation and leakage, rather than the inhibition of CAP binding. Thus, while daptomycin-resistant clinical S. aureus strains often exhibit increased lysyl-PG membrane content and an enhanced surface positive charge (8), the current findings suggest that such phenotypes are more likely to stabilize membrane integrity than repel cationic antimicrobial agents.
FIG. 3.

Carboxyfluorescein leakage induced by 6W-RP-1 (5 μM) from vesicles (50 μM lipid) composed of POPG:POPC:lysyl-DOPG (70:30:0; dashed line), POPG:POPC:lysyl-DOPG (50:30:20; dotted line), POPG:POPC:lysyl-DOPG (40:30:30; dash-dotted line), and POPG:POPC:lysyl-DOPG (30:30:40; solid line). All curves are normalized to the maximum dye release, which was determined by adding Triton X-100 to the vesicles at a final concentration of 1% to dissolve the vesicles (1).
Figure 4 represents a working model that interprets our experimental findings. In this view, binding of 6W-RP-1 occurs from an unstructured state in solution to a bilayer in which the three lipid types (PG, lysyl-PG, and PC) are homogeneously distributed. The preferential interaction of the peptide with anionic PG (Fig. 4, red lipids) leads to the formation of lipid domains enriched in PG (Fig. 4, right) and the exclusion of cationic lysyl-PG (Fig. 4, blue lipids). We postulate that at a high lysyl-PG content, the remaining fraction of PG becomes limiting and the peptides no longer bind efficiently. It is not clear why this process per se should lead to an inhibition of dye leakage from vesicles; it is conceivable that the presence of the positively charged lysyl-PG leads to an overall stabilization of a membrane rich in anionic PG. The possibility of PG domain formation in mixed lipid vesicles induced by the binding of a cationic peptide has been suggested previously (2-4).
FIG. 4.

Cartoon showing peptide binding to a homogeneous bilayer (left) and desorption from a bilayer with PG domains induced by 6W-RP-1 binding (right). Red headgroups represent unmodified PG; blue headgroups represent lysyl-PG; white headgroups represent PC. In the left panel, peptides are drawn as gray, random structures above the bilayer, representing the unfolded state in solution; in the right panel, they are shown as gray rods to indicate their more ordered conformation in association with the bilayer-bound state.
Acknowledgments
Support for these studies was provided in part by the following grants from the National Institutes of Health: AI-39108 (to A.S.B.), AI-39001 and AI-48031 (to M.R.Y.), and AI-088567 (to A.P.).
M.R.Y. is a shareholder of NovaDigm Therapeutics, Inc., and has received prior research funding from Pfizer, Inc., Cubist, Inc., and Novazymes, Inc. None of these entities provided support for the current studies.
Footnotes
Published ahead of print on 26 July 2010.
REFERENCES
- 1.Almeida, P. F., and A. Pokorny. 2010. Binding and permeabilization of model membranes by amphipathic peptides. Methods Mol. Biol. 618:155-169. [DOI] [PubMed] [Google Scholar]
- 2.Arouri, A., M. Dathe, and A. Blume. 2009. Peptide induced demixing in PG/PE lipid mixtures: a mechanism for the specificity of antimicrobial peptides towards bacterial membranes? Biochim. Biophys. Acta 1788:650-659. [DOI] [PubMed] [Google Scholar]
- 3.Epand, R. F., G. Wang, B. Berno, and R. M. Epand. 2009. Lipid segregation explains selective toxicity of a series of fragments derived from the human cathelicidin LL-37. Antimicrob. Agents Chemother. 53:3705-3714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gehman, J. D., F. Luc, K. Hall, T. H. Lee, M. P. Boland, T. L. Pukala, J. H. Bowie, M. I. Aguilar, and F. Separovic. 2008. Effect of antimicrobial peptides from Australian tree frogs on anionic phospholipid membranes. Biochemistry 47:8557-8565. [DOI] [PubMed] [Google Scholar]
- 5.Gregory, S. M., A. Cavenaugh, V. Journigan, A. Pokorny, and P. F. Almeida. 2008. A quantitative model for the all-or-none permeabilization of phospholipid vesicles by the antimicrobial peptide cecropin A. Biophys. J. 94:1667-1680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hachmann, A. B., E. R. Angert, and J. D. Helmann. 2009. Genetic analysis of factors affecting susceptibility of Bacillus subtilis to daptomycin. Antimicrob. Agents Chemother. 53:1598-1609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Houtsmuller, U. M., and L. van Deenen. 1963. Identification of a bacterial phospholipid as an O-ornithine ester of phosphatidyl glycerol. Biochim. Biophys. Acta 70:211-213. [DOI] [PubMed] [Google Scholar]
- 8.Jones, T., M. R. Yeaman, G. Sakoulas, S.-J. Yang, R. A. Proctor, H.-G. Sahl, J. Schrenzel, Y. Q. Xiong, and A. S. Bayer. 2008. Failures in clinical treatment of Staphylococcus aureus infection with daptomycin are associated with alterations in surface charge, membrane phospholipid asymmetry, and drug binding. Antimicrob. Agents Chemother. 52:269-278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mishra, N. N., S. J. Yang, A. Sawa, A. Rubio, C. C. Nast, M. R. Yeaman, and A. S. Bayer. 2009. Analysis of cell membrane characteristics of in vitro-selected daptomycin-resistant strains of methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 53:2312-2318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nishi, H., H. Komatsuzawa, T. Fujiwara, N. McCallum, and M. Sugai. 2004. Reduced content of lysyl-phosphatidylglycerol in the cytoplasmic membrane affects susceptibility to moenomycin, as well as vancomycin, gentamicin, and antimicrobial peptides, in Staphylococcus aureus. Antimicrob. Agents Chemother. 48:4800-4807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.O'Leary, W. M., and S. G. Wilkinson. 1988. Gram-positive bacteria, p. 117-201. In C. Ratledge and S. G. Wilkinson (ed.), Microbial lipids, vol. 1. Academic Press, San Diego, CA. [Google Scholar]
- 12.Peschel, A., R. W. Jack, M. Otto, L. V. Collins, P. Staubitz, G. Nicholson, H. Kalbacher, W. F. Nieuwenhuizen, G. Jung, A. Tarkowski, K. P. van Kessel, and J. A. van Strijp. 2001. Staphylococcus aureus resistance to human defensins and evasion of neutrophil killing via the novel virulence factor MprF is based on modification of membrane lipids with l-lysine. J. Exp. Med. 193:1067-1076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.RajBhandary, U. L., and D. Söll. 2008. Aminoacyl-tRNAs, the bacterial cell envelope, and antibiotics. Proc. Natl. Acad. Sci. U. S. A. 105:5285-5286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Roy, H., and M. Ibba. 2008. RNA-dependent lipid remodeling by bacterial multiple peptide resistance factors. Proc. Natl. Acad. Sci. U. S. A. 105:4667-4672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Short, S. A., and D. C. White. 1971. Metabolism of phosphatidylglycerol, lysyl-phosphatidylglycerol, and cardiolipin of Staphylococcus aureus. J. Bacteriol. 108:219-226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Staubitz, P., H. Neumann, T. Schneider, I. Wiedemann, and A. Peschel. 2004. MprF-mediated biosynthesis of lysylphosphatidylglycerol, an important determinant in staphylococcal defensin resistance. FEMS Microbiol. Lett. 231:67-71. [DOI] [PubMed] [Google Scholar]
- 17.Thedieck, K., T. Hain, W. Mohamed, B. J. Tindall, M. Nimtz, T. Chakraborty, J. Wehland, and L. Jänsch. 2006. The MprF protein is required for lysinylation of phospholipids in listerial membranes and confers resistance to cationic antimicrobial peptides (CAMPs) on Listeria monocytogenes. Mol. Microbiol. 62:1325-1339. [DOI] [PubMed] [Google Scholar]
- 18.Weidenmaier, C., A. Peschel, V. A. Kempf, N. Lucindo, M. R. Yeaman, and A. S. Bayer. 2005. DltABCD- and MprF-mediated cell envelope modifications of Staphylococcus aureus confer resistance to platelet microbicidal proteins and contribute to virulence in a rabbit endocarditis model. Infect. Immun. 73:8033-8038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Xiong, Y. Q., K. Mukhopadhyay, M. R. Yeaman, J. Adler-Moore, and A. S. Bayer. 2005. Functional interrelationships between cell membrane and cell wall in antimicrobial peptide-mediated killing of Staphylococcus aureus. Antimicrob. Agents Chemother. 49:3114-3121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Yeaman, M. R., K. D. Gank, A. S. Bayer, and E. P. Brass. 2002. Synthetic peptides that exert antimicrobial activities in whole blood and blood-derived matrices. Antimicrob. Agents Chemother. 46:3883-3891. [DOI] [PMC free article] [PubMed] [Google Scholar]