

Abstract
At present, 51 genes are already known to be responsible for Non-Syndromic hereditary Hearing Loss (NSHL), but the knowledge of 121 NSHL-linked chromosomal regions brings to the hypothesis that a number of disease genes have still to be uncovered. To help scientists to find new NSHL genes, we built a gene-scoring system, integrating Gene Ontology, NCBI Gene and Map Viewer databases, which prioritizes the candidate genes according to their probability to cause NSHL. We defined a set of candidates and measured their functional similarity with respect to the disease gene set, computing a score (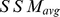 ) that relies on the assumption that functionally related genes might contribute to the same (disease) phenotype. A Kolmogorov-Smirnov test, comparing the pair-wise
) that relies on the assumption that functionally related genes might contribute to the same (disease) phenotype. A Kolmogorov-Smirnov test, comparing the pair-wise 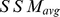 distribution on the disease gene set with the distribution on the remaining human genes, provided a statistical assessment of this assumption. We found at a p-value
distribution on the disease gene set with the distribution on the remaining human genes, provided a statistical assessment of this assumption. We found at a p-value
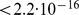 that the former pair-wise
that the former pair-wise 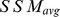 is greater than the latter, justifying a prioritization strategy based on the functional similarity of candidate genes respect to the disease gene set. A cross-validation test measured to what extent the
is greater than the latter, justifying a prioritization strategy based on the functional similarity of candidate genes respect to the disease gene set. A cross-validation test measured to what extent the 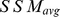 ranking for NSHL is different from a random ordering: adding 15% of the disease genes to the candidate gene set, the ranking of the disease genes in the first eight positions resulted statistically different from a hypergeometric distribution with a p-value
ranking for NSHL is different from a random ordering: adding 15% of the disease genes to the candidate gene set, the ranking of the disease genes in the first eight positions resulted statistically different from a hypergeometric distribution with a p-value
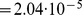 and a power
and a power
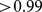 . The twenty top-scored genes were finally examined to evaluate their possible involvement in NSHL. We found that half of them are known to be expressed in human inner ear or cochlea and are mainly involved in remodeling and organization of actin formation and maintenance of the cilia and the endocochlear potential. These findings strongly indicate that our metric was able to suggest excellent NSHL candidates to be screened in patients and controls for causative mutations.
. The twenty top-scored genes were finally examined to evaluate their possible involvement in NSHL. We found that half of them are known to be expressed in human inner ear or cochlea and are mainly involved in remodeling and organization of actin formation and maintenance of the cilia and the endocochlear potential. These findings strongly indicate that our metric was able to suggest excellent NSHL candidates to be screened in patients and controls for causative mutations.
Introduction
Non Syndromic hereditary Hearing Loss (NSHL) is one of the most genetically heterogeneous disorders known. Indeed it can present an autosomal recessive, autosomal dominant, X-linked or mitochondrial pattern of inheritance; furthermore, mutations in the same gene may cause syndromic or non syndromic hearing loss, and recessive forms may be caused by a combination of two mutations in different genes from the same functional group [1].
Due to this tremendous genetic heterogeneity, the identification of genes and gene defects that affect the process of hearing is challenging [1]. At present 51 genes have been already identified to be responsible, if mutated, for this phenotype (see Table 1 for references); nevertheless not all these genes have been fully characterized. They usually are involved in the inner ear development or functionality, and their mutations generally cause hearing loss interfering in the process of the elaboration of sound.
Table 1. NSHL disease genes.
| Gene Symbol | Locus Name | Chromosomal Location | References |
| DIAPH1 | DFNA1 | [5q31.3c] | [43] |
| GJB3 | DFNA2 | [1p34.3f] | [44] |
| KCNQ4 | DFNA2 | [1p34.2c] | [45] |
| GJB2 | DFNA3/DFNB1A | [13q12.11a] | [2] |
| GJB6 | DFNA3/DFNB1B | [13q12.11b] | [3], [46] |
| MYH14 | DFNA4 | [19q13.33c] | [47] |
| DFNA5 | DFNA5 | [7p15.3a] | [48] |
| WFS1 | DFNA6/DFNA14 | [4p16.1f] | [7], [8] |
| TECTA | DFNA8/DFNA12/DFNB21 | [11q23.3h] | [49] |
| COCH | DFNA9 | [14q12e] | [5] |
| EYA4 | DFNA10 | [6q23.2c] | [50] |
| MYO7A | DFNA11/DFNB2 | [11q13.5c] | [51] |
| COL11A2 | DFNA13/DFNB53 | [6p21.32a] | [52] |
| POU4F3 | DFNA15 | [5q32d] | [53] |
| MYH9 | DFNA17 | [22q12.3d] | [54] |
| ACTG1 | DFNA20/DFNA26 | [17q25.3f] | [55], [56] |
| MYO6 | DFNA22/DFNB37 | [6q14.1a] | [57] |
| GRHL2 | DFNA28 | [8q22.3a-q22.3b] | [58] |
| TMC1 | DFNA36/DFNB7/DFNB11 | [9q21.13a] | [59] |
| CRYM | DFNA40 | [16p12.2b] | [60] |
| CCDC50 | DFNA44 | [3q28d] | [61] |
| MYO1A | DFNA48 | [12q13.3a] | [62] |
| KCNJ10 | DFNA49 | [1q23.2c] | [63] |
| MIRN96 | DFNA50 | [7q32.2a] | [64] |
| MYO15A | DFNB3 | [17p11.2g-7p11.2f] | [65] |
| SLC26A4 | DFNB4 | [7q22.3c] | [4] |
| TMIE | DFNB6 | [3p21.31a] | [66] |
| TMPRSS3 | DFNB8/DFNB10 | [21q22.3b] | [67] |
| OTOF | DFNB9 | [2p23.3b] | [68] |
| CDH23 | DFNB12 | [10q22.1d-10q22.1e] | [69] |
| STRC | DFNB16 | [15q15.3a] | [70] |
| USH1C | DFNB18 | [11p15.1d] | [37], [38] |
| OTOA | DFNB22 | [16p12.2a] | [71] |
| PCDH15 | DFNB23 | [10q21.1b-10q21.1c] | [72] |
| RDX | DFNB24 | [11q22.3d] | [73] |
| TRIOBP | DFNB28 | [22q13.1a] | [74], [75] |
| CLDN14 | DFNB29 | [21q22.13a] | [76] |
| MYO3A | DFNB30 | [10p12.1b] | [77] |
| WHRN(DFNB31) | DFNB31 | [9q32e] | [78] |
| ESRRB | DFNB35 | [14q24.3c] | [79], [80] |
| ESPN | DFNB36 | [1p36.31a] | [81] |
| HGF | DFNB39 | [7q21.11c-q21.11d] | [82] |
| KIAA1199 | DFNB48 | [15q25.1b] | [83] |
| MARVELD2 | DFNB49 | [5q13.2a] | [84] |
| PJVK(DFNB59) | DFNB59 | [2q31.2b] | [85] |
| SLC26A5 | DFNB61 | [7q22.1g] | [86] |
| LRTOMT | DFNB63 | [11q13.4] | [87] |
| LHFPL5 | DFNB66/DFNB67 | [6p21.31b] | [40], [88], [89] |
| PRPS1 | DFN2 | [Xq22.3b] | [90] |
| POU3F4 | DFN3 | [Xq21.1d] | [6] |
| ATP2B2 | [3p25.3b] | [91], [92] |
GeneIDs are from NCBI Entrez Gene database; gene symbols correspond to the official gene names as provided by HUGO Gene Nomenclature Committee (HGNC); locus names have been inferred from literature; chromosomal locations are derived from the file cyto_gene.md downloaded from the NCBI Entrez Gene ftp site and references are relative to the articles where the gene association to NSHL was identified.
About 50% of cases of NSHL are due to mutations of GJB2, a gene coding for a gap-junction protein called connexin 26, involved in the cell-cell communication process. Another important gene responsible for NSHL is GJB6, belonging to the same family of GJB2 and adjacent to it. The identification of these two genes highlighted the role of connexins, and therefore of the cochlear gap-junction ion channels, in the auditory function [2], [3].
However the biology of hearing is extremely complex and many other different classes of genes are involved in NSHL. For instance, SLC26A4, associated with autosomal recessive NSHL [4] and Pendred syndrome, is a gene coding for pendrin, a chloride/iodide transporter; COCH, responsible for autosomal dominant non syndromic post-lingual with a progressive onset in adulthood [5], encodes for cochlin, a component of the extracellular matrix of the inner ear; POU3F4, responsible for an X-linked non syndromic progressive and profound sensorineural hearing loss [6], encodes for a transcription factor; while WFS1 associated with autosomal recessive Wolfram syndrome and autosomal dominant low frequency NSHL [7], [8], is a gene coding for the glycoprotein wolframin.
Moreover, several linkage studies over the years have shown that many chromosomal regions are involved in NSHL. At present 121 loci are known to be involved in this phenotype [9], and for many of them the genes causing NSHL have not been identified yet. Due to their often extremely large dimensions – they can even contain several hundreds of genes – it is not feasible to experimentally validate all the genes contained in each locus. In addition, some loci might contain more than one disease gene, as in the case of DFNA3 that harbors GJB2 and GJB6.
In this scenario, a bioinformatic approach to narrow down the list of possible candidate genes is an essential requirement in order to experimentally validate first those genes most likely associated with the disease.
Many strategies have been devised to address this issue, mostly sharing the common prioritization idea of ranking the candidate genes on the basis of their similarity with a set of training genes – genes already associated to the phenotype – relying on the main assumption that genes whose dysfunction contributes to a disease phenotype tend to be functionally related (see [10] and references within).
Quantifying the functional relatedness between two genes is not trivial; often existing information about gene function are exploited to infer functional relationships among genes. In this kind of approach an excellent means is provided by Gene Ontology (GO, The Gene Ontology Consortium, 2001) [11], which is the golden standard ontology in the field of genes and gene products.
Indeed one of the advantages of having genes annotated with GO terms is the possibility to compare them not only from a qualitative point of view (e.g. by searching for common terms with which they are annotated), but also by defining an explicit semantic similarity measure which reflects the closeness in meaning between the terms with which they are annotated [12], [13]. This semantic similarity measure gives in turn a measure of the functional similarity of the annotated gene products, as extensively discussed in Pesquita et al [12].
Briefly, when comparing two terms in an ontology, two main approaches are generally distinguished, the edge-based, which counts the edges in the graph path between two terms [14]–[18], and the node-based, which looks at the properties of the terms, their ancestors and descendants [19]–[25]. Most of the node-based similarity measures are functions of the information content (IC) of each term, and their most informative common ancestors [25]. IC is the amount of information a term contains, meaning that a term contains less information if it occurs very often; in this context the similarity between two terms is quantified looking at the amount of information they share. Very often gene products are annotated with multiple GO terms, in this case maximum [26], [27], average [13], [18], [28] or sum [29] of the GO term similarities may be taken as the gene similarity.
Here we define a new Semantic Similarity Measure (SSM) between gene products by directly extending to sets of concepts (the gene annotations) the Lin's idea [25] of quantifying the similarity between two concepts in an ontology. Our metric provides a measure of the functional similarity between two genes and its reliability is tested in this paper in the context of gene prioritization for NSHL. Indeed the overall aim of this paper is (i) to support researchers in search of new genes responsible for NSHL and (ii) provide indications about the main biological processes, molecular functions and cellular components to be explored to study NSHL, by defining a procedure to computationally prioritize candidate genes for their association with this phenotype. The availability of a good training gene set for NSHL – 51 genes already associated with this phenotype (disease genes) – allows to select new genes most likely responsible for this phenotype estimating their similarity with the disease gene set.
Finally we define a systematic and unbiased statistical assessment to validate the obtained results.
Results
The candidate genes prioritized for NSHL in this study were selected as described in the Methods section. They were prioritized against all the genes already known to be responsible for NSHL (disease genes, see Methods section for details on their selection), according to a score which is function of the Semantic Similarity Measure (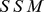 ) estimated for each candidate-disease gene pair. All candidate genes were ranked by computing the
) estimated for each candidate-disease gene pair. All candidate genes were ranked by computing the 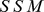 for each candidate-disease gene pair; the final score used for prioritizing each candidate was obtained as the mean of the scores estimated for that candidate against all the disease genes and was defined Semantic Similarity Measure Average (
for each candidate-disease gene pair; the final score used for prioritizing each candidate was obtained as the mean of the scores estimated for that candidate against all the disease genes and was defined Semantic Similarity Measure Average (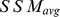 ).
).
Validation of the 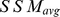 for NSHL gene prioritization
for NSHL gene prioritization
Before being able to assert that the ranking produced by 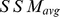 is worthy of attention and therefore evaluating it from a biological point of view, we wanted to evaluate two main aspects concerning our prioritization methodology. We first wanted to test whether the main hypothesis upon which this and most of the prioritization studies are based – genes whose dysfunction contributes to a disease phenotype tend to be functionally related – is quantifiable in terms of semantic similarity, especially in the particular case of NSHL, where the complexity of the hearing process and the complexity of the genetics of the disease both play an important role. Second aspect is whether our metric is able to catch this functional relatedness. To test these two aspects is equivalent to answer the following question: are the disease genes more functionally related than two generic human genes according to
is worthy of attention and therefore evaluating it from a biological point of view, we wanted to evaluate two main aspects concerning our prioritization methodology. We first wanted to test whether the main hypothesis upon which this and most of the prioritization studies are based – genes whose dysfunction contributes to a disease phenotype tend to be functionally related – is quantifiable in terms of semantic similarity, especially in the particular case of NSHL, where the complexity of the hearing process and the complexity of the genetics of the disease both play an important role. Second aspect is whether our metric is able to catch this functional relatedness. To test these two aspects is equivalent to answer the following question: are the disease genes more functionally related than two generic human genes according to 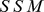 ? A positive answer would yield a positive result for both aspects at the same time, implying that the more a candidate gene obtains a high
? A positive answer would yield a positive result for both aspects at the same time, implying that the more a candidate gene obtains a high 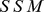 score respect to the disease gene set, the higher is its probability to cause NSHL when mutated. To address this issue, we estimated the pair-wise SSM distribution on the disease gene set, and compared it with the pair-wise SSM distribution estimated on the entire human gene set. In Figure 1 a population pyramid shows the pair-wise SSM distribution across the disease genes and All-human-genes sets in two back-to-back histograms. It provides the graphical evidence that the majority of the disease gene pairs assume
score respect to the disease gene set, the higher is its probability to cause NSHL when mutated. To address this issue, we estimated the pair-wise SSM distribution on the disease gene set, and compared it with the pair-wise SSM distribution estimated on the entire human gene set. In Figure 1 a population pyramid shows the pair-wise SSM distribution across the disease genes and All-human-genes sets in two back-to-back histograms. It provides the graphical evidence that the majority of the disease gene pairs assume 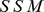 values in the range of 0.5–0.6, much greater than those assumed by the majority of all the remaining human genes (around 0.4). This clearly indicates that the NSHL genes are more functionally related in terms of
values in the range of 0.5–0.6, much greater than those assumed by the majority of all the remaining human genes (around 0.4). This clearly indicates that the NSHL genes are more functionally related in terms of 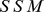 similarity than two generic human genes. In order to statistically support this result, we formulated the following test: the null hypothesis is that the pair-wise SSM distribution in the disease genes set is equal to the pair-wise SSM distribution in the All-human-genes set, while the alternative hypothesis is that the former is greater than the latter, i.e. the cdf (cumulative distribution function) of the former population is smaller than the cdf of the latter population. The test was performed using the bootstrap version of the Kolmogorov-Smirnov test (ks.boot), which allows ties and is included in the R package Matching [30]. We found a p-value
similarity than two generic human genes. In order to statistically support this result, we formulated the following test: the null hypothesis is that the pair-wise SSM distribution in the disease genes set is equal to the pair-wise SSM distribution in the All-human-genes set, while the alternative hypothesis is that the former is greater than the latter, i.e. the cdf (cumulative distribution function) of the former population is smaller than the cdf of the latter population. The test was performed using the bootstrap version of the Kolmogorov-Smirnov test (ks.boot), which allows ties and is included in the R package Matching [30]. We found a p-value
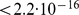 , confirming the hypothesis that the disease genes are indeed more similar according to
, confirming the hypothesis that the disease genes are indeed more similar according to 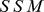 than two generic human genes. This evidence shows the ability of our metric in capturing the functional relatedness of NSHL genes respect to the rest of all human genes, justifying therefore a gene prioritization strategy for association with NSHL based on the
than two generic human genes. This evidence shows the ability of our metric in capturing the functional relatedness of NSHL genes respect to the rest of all human genes, justifying therefore a gene prioritization strategy for association with NSHL based on the 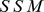 similarity of the candidate genes with respect to the disease gene set.
similarity of the candidate genes with respect to the disease gene set.
Figure 1. Similarity population pyramid.

Back-to-back histograms showing the asymmetry in frequencies of SSM values (in 0.1 bin interval between 0 and 1) among gene pairs, for disease genes (on the right) and the entire human gene set (on the left).
In order to validate the reliability of 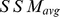 in ranking the candidate genes with respect to their probability to play a causative role in NSHL manifestation, we designed a specific cross-validation procedure that quantifies how much the ranking obtained with our metric differs from a random ordering of the candidate genes. Indeed, due to the specific context we are dealing with, i.e. the gene prioritization, we could not use the classical cross-validation procedure, we in fact added 15% of the disease genes randomly drawn for 10000 times from the disease gene set to the candidates, and counted each time the number of the diseases that fell in the top four windows of 100, 75, 50 and 8 genes. Here the candidate gene set was used exclusively to produce noise, as the positions of the candidates in the ranking were never evaluated during the cross-validation procedure. We in fact tested if the number of disease genes ranked in the top windows were significantly greater than expected when a random extraction of 100, 75, 50 and 8 genes was performed from the total (candidates plus 15% of disease genes) gene set. In Figure 2 we report the distributions obtained from the cross-validation procedure (in blue) applied to the four top windows. In this figure we compare these distributions with the hypergeometric ones (in red), which mimic the random extraction of 100, 75, 50 and 8 genes from the 8748 genes (8740 candidate genes plus 8 disease genes). In all four cases the two distributions are clearly distinct (i.e. the overlapping regions are small). Moreover the means of the distributions for the cross-validation (blue triangles in the figure) result always greater than the means of the hypergeometric distributions (red triangles in the figure). This confirms that the ranking computed by our gene scoring system is significantly different from a random ordering. This is equivalent to assert that our scoring system is able to put at the top of the ranking those genes which are functionally more related to the NSHL genes and thus, more likely, potentially to cause the disease when mutated. This evidence is statistically supported as the p-value and the power of the test for each of the four windows (see Methods section) resulted always smaller than 0.01 and greater than 0.99, respectively (Table 2).
in ranking the candidate genes with respect to their probability to play a causative role in NSHL manifestation, we designed a specific cross-validation procedure that quantifies how much the ranking obtained with our metric differs from a random ordering of the candidate genes. Indeed, due to the specific context we are dealing with, i.e. the gene prioritization, we could not use the classical cross-validation procedure, we in fact added 15% of the disease genes randomly drawn for 10000 times from the disease gene set to the candidates, and counted each time the number of the diseases that fell in the top four windows of 100, 75, 50 and 8 genes. Here the candidate gene set was used exclusively to produce noise, as the positions of the candidates in the ranking were never evaluated during the cross-validation procedure. We in fact tested if the number of disease genes ranked in the top windows were significantly greater than expected when a random extraction of 100, 75, 50 and 8 genes was performed from the total (candidates plus 15% of disease genes) gene set. In Figure 2 we report the distributions obtained from the cross-validation procedure (in blue) applied to the four top windows. In this figure we compare these distributions with the hypergeometric ones (in red), which mimic the random extraction of 100, 75, 50 and 8 genes from the 8748 genes (8740 candidate genes plus 8 disease genes). In all four cases the two distributions are clearly distinct (i.e. the overlapping regions are small). Moreover the means of the distributions for the cross-validation (blue triangles in the figure) result always greater than the means of the hypergeometric distributions (red triangles in the figure). This confirms that the ranking computed by our gene scoring system is significantly different from a random ordering. This is equivalent to assert that our scoring system is able to put at the top of the ranking those genes which are functionally more related to the NSHL genes and thus, more likely, potentially to cause the disease when mutated. This evidence is statistically supported as the p-value and the power of the test for each of the four windows (see Methods section) resulted always smaller than 0.01 and greater than 0.99, respectively (Table 2).
Figure 2. Cross-validation and hypergeometric distributions in case of (a)100, (b)75, (c)50, (d)8 window widths.

In red the hypergeometric distributions with their expectation values (red filled triangles); in blue the distributions, estimated by cross-validation, of disease genes in the top-ranked genes with their mean values (blue filled triangles).
Table 2. Gene scoring system cross-validation.
| Window Width | Mean Value | P-value | Power |
| 100 | 5.845 |
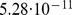
|
1 |
| 75 | 5.313 |
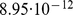
|
1 |
| 50 | 4.502 |
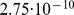
|
0.999 |
| 8 | 1.151 |
|
1 |
Window width indicates the number of top-ranked genes considered in the cross-validation procedure; mean value is the number of disease genes for each window averaged on the 10000 cross-validations; p-value and power are computed as described in the text.
Analysis of the top-ranked candidate genes
The candidates ranked according to 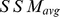 were then examined looking at their functions and expression sites. The twenty top-scored genes are reported in Table 3 together with a brief description of their functions. The number of 20 was arbitrarily chosen, mainly thinking about the intrinsic technical limitations of experimentally testing a great number of genes for disease association – this is actually the reason why such prioritization studies are becoming routine.
were then examined looking at their functions and expression sites. The twenty top-scored genes are reported in Table 3 together with a brief description of their functions. The number of 20 was arbitrarily chosen, mainly thinking about the intrinsic technical limitations of experimentally testing a great number of genes for disease association – this is actually the reason why such prioritization studies are becoming routine.
Table 3. Top-ranked candidate genes.
| Gene symbol | Gene description |
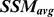
|
Ear expression | Gene Functions for NSHL |
| WDR1 | WD repeat domain 1 | 0.55 | H. sapiens (ear)a | regulation of hair |
| M. musculus (inner ear)a | cell actin dynamicsf | |||
| ALMS1 | Alström syndrome 1 | 0.53 | M. musculus (inner ear)a | normal function of cilia [93] |
| CD151 | CD151 molecule | 0.52 | possible human inner ear component [94] | inner ear ECM assembly [94] |
| (Raph blood group) | M. musculus (inner ear)a | |||
| CLRN1 | clarin 1 | 0.52 | M. musculus (inner ear)a | inner ear developmentf |
| widely expressed in humanb | F actin organizationf | |||
| protein traffickingf | ||||
| ABHD5 | abhydrolase domain containing 5 | 0.52 | M. musculus (inner ear)a | TG accumulationf |
| lipid homeostasisf | ||||
| USH1G | Usher syndrome 1G | 0.52 | H. sapiens (inner ear)b , c | cohesion of hair cell bundlesf |
| (ankyrin and pdz domains) | ||||
| ATP6V0A4 | ATPase H transporting transporting |
0.51 | H. sapiens (cochlea)b [95] | cochlear pH homeostasis [96] |
| lysosomal V0 subunit a4 | ||||
| PRCD | progressive rod-cone degeneration | 0.50 | no data | no evidence |
| KCNQ1 | potassium voltage-gated channel | 0.50 | M. musculus (inner ear)a | K cyclingf cyclingf
|
| KQT-like subfamily member 1 | ||||
| NUMB | numb homolog (Drosophila) | 0.50 | H. sapiens (ear)a | cell fate determination |
| M. musculus (inner ear)a | during developmentf | |||
| ZAR1 | zygote arrest 1 | 0.50 | M. musculus (cochlea, stria vascularis)g | no evidence |
| PTPLA | protein tyrosine phosphatase-like | 0.50 | H. sapiens (fetal cochlea)d | signal transductionf |
| (proline instead of catalytic arginine) | ||||
| member A | ||||
| FLII | flightless I homolog (Drosophila) | 0.50 | H. sapiens (fetal cochlea)d | actin remodeling |
| PTPN11 | protein tyrosine phosphatase | 0.49 | H. sapiens (ear)a | signal transductionf |
| non-receptor type 11 | M. musculus (inner ear)a | |||
| TBL1X | transducin (beta)-like 1X-linked | 0.49 | H. sapiens (fetal cochlea)d | signal transductionf |
| M. musculus (inner ear) | vescicular traffickingf | |||
| cytoskeleton assemblyf | ||||
| KCNE1L | KCNE1-like | 0.49 | M. musculus (inner ear)a | K cyclingf cyclingf
|
| TIMM8A | translocase of inner mitochondrial | 0.49 | no data | signal transductionf |
| membrane 8 homolog A (yeast) | protein transportf | |||
| ROM1 | retinal outer segment | 0.49 | H. sapiens (fetal cochlea)d | cell adhesionf |
| membrane protein 1 | ||||
| CC2D2A | coiled-coil and C2 | 0.49 | no data | Ca bindingf bindingf
|
| domain containing 2A | cilia formationf | |||
| BARHL1 | BarH-like homeobox 1 | 0.48 | M.musculus (inner ear)e | external sensory organ |
| fate determination [97] |
Gene expression information are taken from
NCBI Unigene [98],
UniProtKB [99],
HPRD database [100],
the table of gene expression in the developing ear from the Institute of Hearing Research [103],
Bgee dataBase for Gene Expression Evolution [104] and literature. Gene function information have been inferred from
NCBI Gene [39] and literature.
Half of them are reported in literature to be expressed in human inner ear or cochlea, despite the very limited availability of gene expression data for these tissues due to the technical difficulties of obtaining undamaged hair-cell samples for gene expression experiments. For the remaining genes, six are reported to be expressed in other organisms' inner ear or cochlea, mainly mouse or chicken, while four have no gene expression data for these tissues. Taken altogether, these are important indications supporting the goodness of the ranking we produced in respect to the NSHL, especially if we think that the initial candidate gene list was not a priori filtered by any criterium except that of being all annotated genes located in the susceptibility loci.
Moreover, looking at their functions, we found that most of the top-ranked genes play roles compatible with a possible involvement in NSHL phenotype. Among the most relevant, we identified a) processes of remodeling and organization of actin (WDR1, CLRN1, FLII), an essential component of the hair-cell bundle; b) formation and maintenance of cilia (ALMS1, USH1G, CC2D2A), the sensory organelles devoted to receive the mechanical stimulus; c) 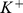 cycling and pH homeostasis in cochlear fluids (ATP6V0A4, KCNQ1, KCNE1L), essentials for the generation and maintenance of the endocochlear potential; d) signal transduction (PTPLA, PTPN11, TBL1X, TIMM8A). They are all important molecular mechanisms underlying the hearing process, which involve the hair cell capability to transduce the mechanical stimulus into electrical signal, as well as the endolymph production and maintenance.
cycling and pH homeostasis in cochlear fluids (ATP6V0A4, KCNQ1, KCNE1L), essentials for the generation and maintenance of the endocochlear potential; d) signal transduction (PTPLA, PTPN11, TBL1X, TIMM8A). They are all important molecular mechanisms underlying the hearing process, which involve the hair cell capability to transduce the mechanical stimulus into electrical signal, as well as the endolymph production and maintenance.
Stronger evidences come from some of the top-ranked genes which are already linked to different syndromic forms of deafness: USH1G for instance is known to cause Usher syndrome type 1G [31], associated with sensorineural hearing impairment; for this gene a possible role in the development and maintenance of the stereocilia bundles is reported by Weil et al. [31]: it might in fact function as an anchoring/scaffolding protein in hair cells and could be involved in the functional network formed by USH1C, CDH23 and MYO7A that is required for cohesion of the growing hair bundle, making its role in the hearing impairment process quite easily explainable. Similarly, KCNE1L has been associated by Piccini et al. [32] to AMME syndrome (Alport syndrome - mental retardation - midface hypoplasia - elliptocytosis) whose symptoms include, among others, hearing loss, and analogous situations are reported also for TIMM8A, involved in Mohr-Tranebjaerg syndrome [33] and Jensen syndrome [34], and ALMS1, involved in Alström syndrome [35]. It is noteworthy that the association of some top-ranked genes to syndromic deafness forms does not exclude them from being good NSHL candidates, as clearly demonstrated by USH1C involved both in Usher syndrome type 1C [36], and NSHL [37], [38], depending on which mutations it undergoes.
Finally, we produced a graphical bidimensional representation of the 20 top-ranked genes together with the disease genes using Proxscal SPSS, which performs multidimensional scaling of similarity data to find a least squares representation of the objects in a low-dimensional space (Figure 3). The proximity of the two gene sets was in this way highlighted; this allowed identifying different groups of NSHL disease genes (red balls in the figure) – namely myosins, connexins, cadherins, ion channels and so forth – and mapping the best candidates within these groups. The inclusion of the top-scored candidate genes did not enlarge the area occupied by the disease genes and their membership to the relative subgroups was mantained in the graphical representation.
Figure 3. Multidimensional scaling of similarity data to represent the disease and the 20 top-scored candidate genes in a bidimensional space.

Overall, on the basis of these considerations, the majority of them seem to be excellent candidates for subsequent studies on NSHL patients and controls.
Functional characterization of candidate and disease genes using GO
In order to further investigate the obtained ranking and in order to have a more general picture of the molecular functions, biological processes and cellular components more associated to NSHL, as suggested by both the best candidates and disease genes, we designed and implemented two specific statistical tests that allowed to identify the GO terms more representative of NSHL, exploiting the 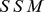 score estimated by our gene scoring system. For the disease genes, we quantified and tested the enrichment of gene-sets defined by functional categories provided by Gene Ontology annotations in disease gene list. In this case the
score estimated by our gene scoring system. For the disease genes, we quantified and tested the enrichment of gene-sets defined by functional categories provided by Gene Ontology annotations in disease gene list. In this case the 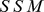 score was used to define the non-disease gene class (see Methods section). For the candidates, we analyzed all GO terms in their annotations, and evaluated the enrichment of the gene set annotated with each of them, by using the
score was used to define the non-disease gene class (see Methods section). For the candidates, we analyzed all GO terms in their annotations, and evaluated the enrichment of the gene set annotated with each of them, by using the 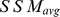 score obtained from our ranking to quantify their association with NSHL (see Materials and Methods section). In this case the
score obtained from our ranking to quantify their association with NSHL (see Materials and Methods section). In this case the 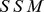 score allowed us, starting from the GO terms associated to all the candidate genes, to identify those GO terms significantly associated with the best candidates, without making any a priori decision on which candidates should be considered as the “best” candidates.
score allowed us, starting from the GO terms associated to all the candidate genes, to identify those GO terms significantly associated with the best candidates, without making any a priori decision on which candidates should be considered as the “best” candidates.
This survey had the purpose to examine the ranking on a larger scale – extending the ranking examination to the whole candidate gene set – to possibly suggest non-obvious pathways to further look into when studying NSHL, hence it was devised as a way to look at the results from a different point of view (i.e. moving from a view of NSHL in terms of genes responsible of the disease to a view of NSHL in terms of biological processes, molecular functions and cellular components distinctive of the disease).
We considered as significantly descriptive of the best candidate and disease genes, those GO terms with a p-value
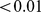 and we ordered them according to their
and we ordered them according to their 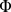 score, function of their p-value and specificity in the corpus of the GO annotations.
score, function of their p-value and specificity in the corpus of the GO annotations.
As for the candidate genes, the enriched terms, divided into biological processes, molecular functions and cellular components (Table 4), include expected concepts such as “auditory receptor cell stereocilium organization” (GO:0060088), “large conductance calcium-activated potassium channel activity” (GO:0060072), “sensory perception of sound” (GO:0007605), “auditory receptor cell stereocilium organization” (GO:0060088), consistent with hearing physiology, as well as less obvious functions or processes such as “regulation of circadian sleep/wake cycle, REM and non-REM sleep” (GO:0042320, GO:0045188), “response to cocaine” (GO:0042220), or “mu-type opioid receptor binding” (GO:0031852), that need further (experimental) investigations. This on the one hand supports again the goodness of the ranking, confirming that the top-scored genes are actually promising candidates for association with NSHL, on the other hand fulfils our initial requirement to suggest new prospective insights in NSHL.
Table 4. Enriched biological processes, cellular components and molecular functions for candidate genes.
| GO term |
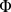 Score Score |
P-value | Definition | Ontology |
| GO:0060082 | 17.0 | 0.002 | eye blink reflex | biological process |
| GO:0014010 | 16.3 | 0.005 | Schwann cell proliferation | biological process |
| GO:0034465 | 16.3 | 0.002 | response to carbon monoxide | biological process |
| GO:0060231 | 16.2 | 0.010 | mesenchymal to epithelial transition | biological process |
| GO:0021771 | 16.1 | 0.001 | lateral geniculate nucleus development | biological process |
| GO:0032344 | 16.0 | 0.002 | regulation of aldosterone metabolic process | biological process |
| GO:0045759 | 16.0 | 0.001 | negative regulation of action potential | biological process |
| GO:0045794 | 15.9 | 0.002 | negative regulation of cell volume | biological process |
| GO:0021562 | 15.7 | 0.001 | vestibulocochlear nerve development | biological process |
| GO:0050975 | 15.6 | 0.005 | sensory perception of touch | biological process |
| GO:0051451 | 15.6 | 0.004 | myoblast migration | biological process |
| GO:0031630 | 15.5 | 0.005 | regulation of synaptic vesicle fusion to presynaptic membrane | biological process |
| GO:0048790 | 15.5 | 0.005 | maintenance of presynaptic active zone structure | biological process |
| GO:0046007 | 15.2 | 0.005 | negative regulation of activated T cell proliferation | biological process |
| GO:0046541 | 15.0 | 0.005 | saliva secretion | biological process |
| GO:0048676 | 14.9 | 0.005 | retinal bipolar neuron differentiation | biological process |
| GO:0045188 | 14.9 | 0.001 | regulation of circadian sleep/wake cycle, non-REM sleep | biological process |
| GO:0050916 | 14.8 | 0.010 | sensory perception of sweet taste | biological process |
| GO:0035022 | 14.8 | 0.009 | positive regulation of Rac protein signal transduction | biological process |
| GO:0042524 | 14.9 | 0.005 | negative regulation of tyrosine phosphorylation of Stat5 protein | biological process |
| GO:0060083 | 14.7 | 0.002 | smooth muscle contraction involved in micturition | biological process |
| GO:0042320 | 14.7 | 0.001 | regulation of circadian sleep/wake cycle, REM sleep | biological process |
| GO:0051496 | 14.6 | 0.005 | positive regulation of stress fiber formation | biological process |
| GO:0030007 | 14.5 | 0.002 | cellular potassium ion homeostasis | biological process |
| GO:0001661 | 14.5 | 0.001 | conditioned taste aversion | biological process |
| GO:0051602 | 14.4 | 0.005 | response to electrical stimulus | biological process |
| GO:0032287 | 14.4 | 0.004 | myelin maintenance in the peripheral nervous system | biological process |
| GO:0050957 | 14.2 | 0.001 | equilibrioception | biological process |
| GO:0045475 | 14.1 | 0.002 | locomotor rhythm | biological process |
| GO:0001895 | 14.1 | 0.005 | retina homeostasis | biological process |
| GO:0060087 | 14.0 | 0.003 | relaxation of vascular smooth muscle | biological process |
| GO:0048484 | 14.0 | 0.007 | enteric nervous system development | biological process |
| GO:0022408 | 14.0 | 0.005 | negative regulation of cell-cell adhesion | biological process |
| GO:0060088 | 13.9 | 0.004 | auditory receptor cell stereocilium organization | biological process |
| GO:0021952 | 13.8 | 0.005 | central nervous system projection neuron axonogenesis | biological process |
| GO:0033081 | 13.8 | 0.004 | regulation of T cell differentiation in the thymus | biological process |
| GO:0051963 | 13.0 | 0.001 | regulation of synaptogenesis | biological process |
| GO:0042220 | 12.9 | 0.001 | response to cocaine | biological process |
| GO:0002262 | 12.9 | 0.004 | myeloid cell homeostasis | biological process |
| GO:0007019 | 12.7 | 0.005 | microtubule depolymerization | biological process |
| GO:0060113 | 12.5 | 0.001 | inner ear receptor cell differentiation | biological process |
| GO:0046620 | 12.4 | 0.004 | regulation of organ growth | biological process |
| GO:0007605 | 11.7 | 0.004 | sensory perception of sound | biological process |
| GO:0045039 | 11.5 | 0.005 | protein import into mitochondrial inner membrane | biological process |
| GO:0031667 | 9.9 | 0.004 | response to nutrient levels | biological process |
| GO:0019725 | 7.2 | 0.004 | cellular homeostasis | biological process |
| GO:0017071 | 15.9 | 0.005 | intracellular cyclic nucleotide activated cation channel complex | cellular component |
| GO:0032588 | 15.7 | 0.005 | trans-Golgi network membrane | cellular component |
| GO:0032839 | 14.1 | 0.004 | dendrite cytoplasm | cellular component |
| GO:0032154 | 13.9 | 0.005 | cleavage furrow | cellular component |
| GO:0016011 | 13.1 | 0.009 | dystroglycan complex | cellular component |
| GO:0042719 | 11.6 | 0.005 | mitochondrial intermembrane space protein transporter complex | cellular component |
| GO:0030660 | 10.3 | 0.005 | Golgi-associated vesicle membrane | cellular component |
| GO:0031852 | 17.0 | 0.005 | mu-type opioid receptor binding | molecular function |
| GO:0043533 | 16.3 | 0.008 | inositol 1.3.4.5 tetrakisphosphate binding | molecular function |
| GO:0060072 | 15.7 | 0.002 | large conductance calcium-activated potassium channel activity | molecular function |
| GO:0015266 | 14.4 | 0.004 | protein channel activity | molecular function |
| GO:0030346 | 14.2 | 0.004 | protein phosphatase 2B binding | molecular function |
| GO:0000822 | 13.8 | 0.008 | inositol hexakisphosphate binding | molecular function |
Candidate gene enriched (p-value
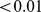 ) GO terms, sorted according to their
) GO terms, sorted according to their 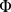 score in each ontology.
score in each ontology. 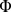 scores take into account the specificity of the terms as described in the text.
scores take into account the specificity of the terms as described in the text.
As for the disease genes, as expected, the enriched terms are all consistent with the hearing physiology (Table 5). To give some examples, among the most relevant enriched biological processes we found “actin filament-based movement” (GO:0030048), “inner ear morphogenesis” (GO:0042472), “regulation of cell shape” (GO:0008360) and a group involving sensory perception (GO:0007605, GO:0007601, GO:0050957). Likewise, among the enriched cellular components, are “stereocilium” (GO:0032420), “myosin complex” (GO:0016459), “cell junction” (GO:0030054), and among the molecular functions, “actin binding” (GO:0003779), “actin filament binding” (GO:0051015), and so forth.
Table 5. Enriched biological processes, cellular components and molecular functions for disease genes.
| GO term |
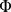 Score Score |
P-value | Definition | Ontology |
| GO:0007605 | 150.4 |
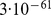
|
sensory perception of sound | biological process |
| GO:0007601 | 22.7 |
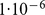
|
visual perception | biological process |
| GO:0050957 | 22.7 | 0.0001 | equilibrioception | biological process |
| GO:0030048 | 21.2 | 0.0001 | actin filament-based movement | biological process |
| GO:0045494 | 19.5 | 0.001 | photoreceptor cell maintenance | biological process |
| GO:0050896 | 18.1 |
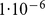
|
response to stimulus | biological process |
| GO:0042472 | 17.8 | 0.001 | inner ear morphogenesis | biological process |
| GO:0008360 | 14.2 | 0.001 | regulation of cell shape | biological process |
| GO:0007155 | 14.0 | 0.001 | cell adhesion | biological process |
| GO:0006355 | 12.0 | 0.0005 | regulation of cellular transcription, DNA-dependent | biological process |
| GO:0006350 | 10.2 | 0.001 | cellular transcription | biological process |
| GO:0006810 | 7.8 | 0.009 | transport | biological process |
| GO:0005886 | 45.5 |
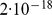
|
plasma membrane | cellular component |
| GO:0016021 | 39.6 |
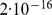
|
integral to membrane | cellular component |
| GO:0005737 | 36.5 |
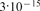
|
cytoplasm | cellular component |
| GO:0032420 | 24.7 |
|
stereocilium | cellular component |
| GO:0016459 | 23.0 |
|
myosin complex | cellular component |
| GO:0005856 | 22.9 |
|
cytoskeleton | cellular component |
| GO:0030054 | 22.2 |
|
cell junction | cellular component |
| GO:0031941 | 21.9 | 0.0001 | filamentous actin | cellular component |
| GO:0005634 | 20.1 |
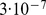
|
nucleus | cellular component |
| GO:0016324 | 19.8 | 0.0001 | apical plasma membrane | cellular component |
| GO:0001726 | 18.4 | 0.001 | ruffle | cellular component |
| GO:0005922 | 18.1 | 0.001 | connexon complex | cellular component |
| GO:0005829 | 16.9 |
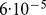
|
cytosol | cellular component |
| GO:0005783 | 16.1 | 0.0001 | endoplasmic reticulum | cellular component |
| GO:0045202 | 15.2 | 0.001 | synapse | cellular component |
| GO:0016020 | 15.8 |
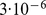
|
membrane | cellular component |
| GO:0042995 | 15.1 | 0.0001 | cell projection | cellular component |
| GO:0005789 | 14.4 | 0.001 | endoplasmic reticulum membrane | cellular component |
| GO:0003779 | 29.4 |
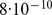
|
actin binding | molecular function |
| GO:0005516 | 28.5 |
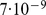
|
calmodulin binding | molecular function |
| GO:0051015 | 20.1 | 0.0001 | actin filament binding | molecular function |
| GO:0043531 | 19.8 | 0.001 | ADP binding | molecular function |
| GO:0003774 | 18.6 |
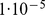
|
motor activity | molecular function |
| GO:0005515 | 16.6 |
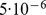
|
protein binding | molecular function |
| GO:0004749 | 16.2 | 0.001 | ribose phosphate diphosphokinase activity | molecular function |
| GO:0042803 | 14.2 | 0.004 | protein dimerization activity | molecular function |
| GO:0005524 | 12.7 | 0.0001 | ATP binding | molecular function |
| GO:0043565 | 12.5 | 0.001 | sequence-specific DNA binding | molecular function |
| GO:0000166 | 11.9 | 0.0001 | nucleotide binding | molecular function |
| GO:0003700 | 11.6 | 0.001 | transcription factor activity | molecular function |
Disease gene enriched (p-value
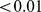 ) GO terms, sorted according to their
) GO terms, sorted according to their 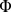 score in each ontology.
score in each ontology. 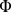 scores take into account the specificity of the terms as described in the text.
scores take into account the specificity of the terms as described in the text.
Interestingly, among all the enriched terms – for both candidate and disease genes – there is a very small amount of overlapping. Only two biological processes are in fact shared between the two gene lists, “sensory perception of sound” (GO:0007605) and “equilibrioception” (GO:0050957), which are neverthless extremely specific terms – a very small number of gene products are annotated with these terms – both deeply linked to the inner ear function. Looking at the GO graph, however, many of the non-shared terms are interconnected with each other on a larger scale, sharing a common parent at different levels of specificity. This is due to the structure of our algorithm that favours the closeness in the graph of the terms in estimating the similarity between genes. It is noteworthy that with this approach we can think of NSHL from a different perspective, exploring portions of the graph that otherwise would have never been explored.
In Figure 4 we reported an elucidative example of this issue: by mapping some enriched disease and candidate biological processes to the GO graph, we observed that the addition of “inner ear receptor cell differentiation” (GO:0060113) to the list of NSHL possible biological processes clearly enlarges the NSHL subgraph covering a new branch of the “inner ear development” (GO:0048839) different from the “inner ear morphogenesis” (GO:0042472), while the addition of “auditory receptor cell stereocilium organization” (GO:0060088) narrows and specializes the concept “inner ear morphogenesis” to one of its components.
Figure 4. GO subgraph of some disease and candidate gene enriched GO terms.

Red circles indicate terms enriched for the disease genes, green circles indicate terms enriched for the candidate genes. Dark blue arrows indicate is a relations, light blue arrows indicate part of relations between the terms.
These findings, as a whole, on the one hand support again the goodness of the ranking, on the other hand they suggest that also some pathways apparently unrelated with NSHL, might deserve future attention by NSHL researchers.
Discussion
In the perspective of discovering new genes potentially involved in NSHL, we built a gene scoring system integrating Gene Ontology (GO), NCBI Gene and Map Viewer databases, which scores the candidate genes for NSHL by comparing them with the 51 NSHL disease genes already known, relying on the assumption that functionally related genes might contribute to the same (disease) phenotype.
We defined a set of candidate genes for NSHL as all the genes contained in the susceptibility loci known so far, and we prioritized them for the association with the disease, without making any a priori selection except that of being annotated with at least one GO term.
We first of all tested whether our metric, 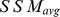 , was able to capture the above assumption, verifying that the disease genes are indeed more similar, according to the metric, than two generic human genes. We also demonstrated that our metric is able to pool the disease genes respect to the other human genes, implying that the former are indeed more closely functionally related than the latter: these results therefore justify a prioritization strategy based on the similarity of the candidate genes respect to the disease gene set.
, was able to capture the above assumption, verifying that the disease genes are indeed more similar, according to the metric, than two generic human genes. We also demonstrated that our metric is able to pool the disease genes respect to the other human genes, implying that the former are indeed more closely functionally related than the latter: these results therefore justify a prioritization strategy based on the similarity of the candidate genes respect to the disease gene set.
Afterwards, we wanted to investigate to what extent our metric is reliable in ranking candidate genes for their potential role in NSHL manifestation. To this purpose we designed a cross-validation procedure and we obtained excellent results also considering the more disadvantageous condition of ranking eight disease genes in the first 8 positions of a list of more than 8700 genes.
Given these preliminary validations, we are extremely confident that the ranking we produced with respect to NSHL is worthy of attention for future NSHL research plan. Indeed, the top-scored candidate genes play all roles compatible with a possible involvement in NSHL phenotype, representing therefore excellent candidates for subsequent studies on NSHL patients and controls.
However two main limitations of this kind of approach should also be taken into account when looking at these data, both concerning the usage of Gene Ontology annotations to build the gene profiles on which the semantic similarity is measured. One is linked to the current knowledge about the human genome and its content in terms of genes. Indeed, the only prerequisite for a gene to be prioritized by our gene scoring system for a given disease is that of being annotated with at least one GO term, but, as clearly evidenced in this study, we are still far from the complete annotation of the entire human genome, as we were forced to exclude almost half of the possible candidates since they completely lacked GO annotations. This limitation obviously biases the results towards the best studied genes; however it will be progressively overcome in the future, due to the daily updates in this field. The other limitation regards the nature of the associations between GO terms and gene products. All the associations in Gene Ontology fall in five general categories indicating the evidences that support the annotation of a gene to a specific term. Four of these categories comprise exclusively manually-curated associations supported by experimental, computational analysis, author statements or curatorial statements. Unfortunately the great majority of GO associations does not fall in any of these manually-curated categories, being inferred from electronic annotation (IEA), which may open a debate on how reliable and precise they are. At present, given the high percentage of IEA associations in GO, it is not conceivable to discard them and consider only those manually-curated. Other solutions must therefore be devised to address this issue. Future developments of our gene scoring system could for instance take into account this problem by down-weighting the IEA associations respect to those manually-curated. However the quantification of the difference in weight between the manual and electronic associations is not trivial and requires an accurate study of the algorithms behind the electronic associations. We reserve in future to enhance our algorithm in this direction.
Final and essential step to confirm the results presented in this study is however the experimental validation. To this end two main aspects should be taken into account: (i) the accurate study and selection among the top-ranked genes of the most intringuing candidates for NSHL; we think for instance that the first one (WDR1) represents a good starting point, due to both what is known about its functions and structure – it is indeed involved in the organization of the actin, fundamental for the auditory process, and small enough to be quite easily sequenced in a large number of subjects; (ii) an equally accurate selection of the appropriate NSHL patients and controls to be screened for causative mutations; it is advisable for instance to screen these genes on a cohort of patients already excluded to carry mutations in GJB2, due to the high incidence of NSHL cases caused by mutations in this gene, and on a control set appropriately matched for their geographic origin, in order to take into account the geographic distribution of the human DNA sequence variation.
Methods
A total of 15727 genes (candidate genes) were prioritized for NSHL in this study. We chose as candidate genes all the genes contained in the NSHL susceptibility loci known so far (Tables S1, S2, S3 respectively for NSHL autosomal dominant, autosomal recessive and X-linked, Y-linked and modifier loci), so that all evidences coming from previous linkage analysis studies were taken into account.
We drew the complete disease gene list starting from the Hereditary Hearing Loss Homepage [9] and a team of experts (geneticists and molecular biologists) further analysed the literature to find additional advances in the field by performing multiple queries on PubMed. To the best of our knowledge, 51 genes belong to this category, as reported in Table 1.
For each disease and candidate gene, we extracted all their GO annotations using the file gene2go downloaded on 29th May 2009 from NCBI Entrez Gene ftp site [39]. One out of fifty-one disease genes – MIRN96 – had no GO annotations, therefore it was not included in this study, consequently narrowing the disease gene list to fifty genes. Likewise, 6987 out of 15727 candidate genes had no GO annotations, therefore the candidate gene list was consequently narrowed to 8740 genes.
Semantic similarity between two genes
As a node-based approach, our metric computes the similarity between two genes by comparing the GO terms describing them, their ancestors, and their descendants in the GO network. It is based on the Information Content (IC), which gives a measure of how specific and informative a term is. The IC of a term  is quantified as the negative log likelihood
is quantified as the negative log likelihood
| (1) |
where 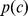 is the probability of occurrence of
is the probability of occurrence of  in a specific corpus, which is normally estimated by the frequency of annotation of the term and its children in the GO structure [24], [40].
in a specific corpus, which is normally estimated by the frequency of annotation of the term and its children in the GO structure [24], [40].
The concept of IC was used by Lin to quantify the semantic similarity between two terms in a tree-structured ontology, measuring the information they share normalized respect to the information contained in their total descriptions. According to Lin's metric [25] the similarity between two terms 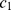 and
and 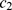 is defined as:
is defined as:
| (2) |
where 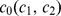 is the most informative common ancestor of the terms
is the most informative common ancestor of the terms 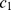 and
and 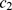 , i.e. the common ancestor with the smallest
, i.e. the common ancestor with the smallest 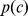 .
.
However, two aspects of this metric limit its application:
it applies only to trees, where a unique most informative common ancestor between two any given concepts exists;
it measures the distance between single terms rather than set of terms.
For the first drawback, it is well known that in the case of a direct acyclic graph (DAG), such as GO, two terms can share parents by multiple paths, as multiple parents for each concept are allowed. Therefore, we chose, as 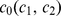 , the minimum subsumer between
, the minimum subsumer between 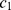 and
and 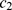 along all their independent paths to the graph root [13].
along all their independent paths to the graph root [13].
To address the second issue, we defined our Semantic Similarity Measure (SSM) by directly extending Lin's idea to quantify the similarity between two concepts to the comparison between two gene products, i.e. two sets of concepts, therefore measuring the IC of the common description of the two gene products, normalized respect to the IC of their global description.
Let be
two gene products, annotated respectively with  and
and  GO terms, that are to be compared. The idea is that each term in
GO terms, that are to be compared. The idea is that each term in 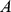 is an independent view of the gene
is an independent view of the gene 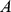 and has to be compared with its counterpart in the
and has to be compared with its counterpart in the 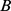 gene annotation, namely the term in
gene annotation, namely the term in 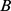 with maximum IC for the common description respect to it. In formulas, for the term
with maximum IC for the common description respect to it. In formulas, for the term 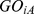 , its counterpart
, its counterpart 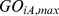 in
in 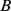 is defined as:
is defined as:
The IC for each GO term c is estimated using its probability of occurrence 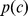 in the corpus of all gene annotations provided by ENGINE database [41], [42]: in details, the probability
in the corpus of all gene annotations provided by ENGINE database [41], [42]: in details, the probability 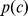 is calculated for every term by counting the number of gene products associated with the term or any of its children, divided by the number of total associations between the GO terms and gene products.
is calculated for every term by counting the number of gene products associated with the term or any of its children, divided by the number of total associations between the GO terms and gene products.
Considering as independent the single views of a gene offered by each of its terms, the semantic similarity of 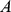 respect to
respect to 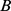 is estimated by the sum of the shared common description ICs between each term in
is estimated by the sum of the shared common description ICs between each term in 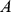 and its counterpart in
and its counterpart in 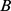 normalized with the IC of their global description:
normalized with the IC of their global description:
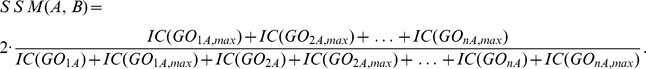 |
The similarity of the gene 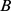 respect to
respect to 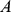 (
(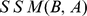 ) is obtained by inverting the roles of
) is obtained by inverting the roles of 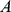 and
and 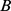 in the above formula. Finally, we defined our Semantic Similarity Measure between
in the above formula. Finally, we defined our Semantic Similarity Measure between 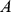 and
and 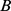 ,
, 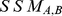 , as the mean between the similarity of
, as the mean between the similarity of 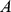 respect to
respect to 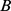 and the similarity of
and the similarity of 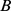 respect to
respect to 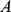 :
:
| (3) |
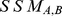 generates normalized similarity values between 0 and 1: it's equal to 0 for genes annotated with terms that share only the root and equal to 1 for genes annotated with the same terms.
generates normalized similarity values between 0 and 1: it's equal to 0 for genes annotated with terms that share only the root and equal to 1 for genes annotated with the same terms.
Validation of the 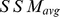 for NSHL gene prioritization
for NSHL gene prioritization
A cross-validation procedure was used to check the reliability of the ranking of candidate genes for their involvement in NSHL. A random set of 8 disease genes was added to the set of candidate genes for 10000 times. Each time the 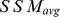 values for this enlarged set of candidates were computed against the remaining disease genes and the number of disease genes
values for this enlarged set of candidates were computed against the remaining disease genes and the number of disease genes 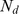 in the first
in the first 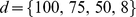 top-ranked positions was counted. The corresponding
top-ranked positions was counted. The corresponding 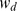 distributions of these countings were then compared with the probabilities of counting
distributions of these countings were then compared with the probabilities of counting  disease genes when a random drawn of 100, 75, 50, 8 genes, respectively, was performed from a set of 8748 genes (8740 candidate genes plus 8 disease genes): in the case of random drawns, the countings are described by a hypergeometric distribution with
disease genes when a random drawn of 100, 75, 50, 8 genes, respectively, was performed from a set of 8748 genes (8740 candidate genes plus 8 disease genes): in the case of random drawns, the countings are described by a hypergeometric distribution with  successes for d draws without replacement.
successes for d draws without replacement.
More in details, we computed the p-value and the power of a statistical test on the hypothesis of equal distributions 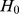 against the hypothesis
against the hypothesis 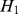 of a greater number of disease genes in the first positions for the
of a greater number of disease genes in the first positions for the 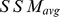 ranked ordering respect to the random ordering. The p-value measures the probability to obtain, by random extraction, a number of disease genes
ranked ordering respect to the random ordering. The p-value measures the probability to obtain, by random extraction, a number of disease genes 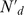 greater than the mean value
greater than the mean value 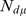 of the number of disease genes found in the
of the number of disease genes found in the 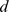 top-ranked positions on the 10000 cross-validations:
top-ranked positions on the 10000 cross-validations:
| (4) |
The p-value is our estimate of the probability of rejecting 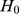 when
when 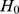 is true: whenever the p-value was less than the significance level
is true: whenever the p-value was less than the significance level 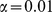 , we maintained that the number of disease genes found in the top-ranked positions was statistically significantly greater than that found in random orderings.
, we maintained that the number of disease genes found in the top-ranked positions was statistically significantly greater than that found in random orderings.
The knowledge of the empirical distribution of 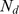 estimated through the cross-validation procedure, allowed us to estimate the power
estimated through the cross-validation procedure, allowed us to estimate the power
 of the test with level
of the test with level  : indicating with
: indicating with 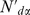 the
the  -quantile of the hypergeometric distribution,
-quantile of the hypergeometric distribution,  is computed as follows:
is computed as follows:
| (5) |
where 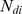 is the number of disease genes found in the first
is the number of disease genes found in the first 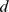 positions for the
positions for the 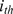 randomization. The larger is the percentage of
randomization. The larger is the percentage of 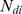 values obtained in cross-validation that are greater than
values obtained in cross-validation that are greater than 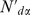 , the more effective is the gene prioritization system.
, the more effective is the gene prioritization system.
Functional characterization of candidate and disease genes for NSHL
Candidate genes
The statistical test used to identify the most representative GO terms associated with the candidate genes was designed as follows: the null hypothesis is that candidate genes annotated with a particular GO category have an average 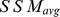 score equal to the average score expected for a random list of candidate genes with the same size, whereas the alternative hypothesis is that the GO category list has a higher average score and, therefore, is supposed to be more associated with the disease than a random candidate gene list. After selecting all the GO terms associated with all the candidate genes, we computed a p-value which scores each GO term according to the following strategy: the higher is the number of its associated candidates which obtained in our ranking a high
score equal to the average score expected for a random list of candidate genes with the same size, whereas the alternative hypothesis is that the GO category list has a higher average score and, therefore, is supposed to be more associated with the disease than a random candidate gene list. After selecting all the GO terms associated with all the candidate genes, we computed a p-value which scores each GO term according to the following strategy: the higher is the number of its associated candidates which obtained in our ranking a high 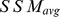 value, the more the GO term is considered enriched in the candidate gene list. This implies that, choosing a significance threshold of 0.01, the GO terms with p-value
value, the more the GO term is considered enriched in the candidate gene list. This implies that, choosing a significance threshold of 0.01, the GO terms with p-value
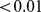 can be considered significantly descriptive of the best candidate genes and consequently significantly associated with the disease. This provides directions for the NSHL researchers about the functions to be more deeply investigated in future laboratory experiments.
can be considered significantly descriptive of the best candidate genes and consequently significantly associated with the disease. This provides directions for the NSHL researchers about the functions to be more deeply investigated in future laboratory experiments.
The p-value for the i-th GO term is computed as follows:
| (6) |
where 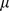 is the average of the
is the average of the  scores resulting for candidate genes annotated with
scores resulting for candidate genes annotated with  ,
,  is the number of candidate genes annotated with the
is the number of candidate genes annotated with the  category,
category, 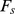 is the empirical cumulative distribution for the
is the empirical cumulative distribution for the 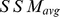 scores, averaged on lists of candidate genes of size
scores, averaged on lists of candidate genes of size  .
. 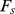 was computed by drawing 10000 random lists of candidate genes of size
was computed by drawing 10000 random lists of candidate genes of size  and averaging the respective gene scores.
and averaging the respective gene scores.
Disease genes
After selecting all the GO terms used to annotate the disease genes, we computed for each GO term a Fisher's exact test p-value which scores the GO category (GO Term C) highly for enrichment if many more disease genes than expected belong to the category. The contingency table (Table 6) is built by counting the disease and non-disease genes associated and not associated with the GO category.
Table 6. Contingency table.
| GO Term C | Disease | Not Disease |
| Genes annotated with C |
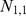
|
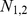
|
| Genes not annotated with C |
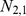
|
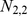
|
C represents the generic GO term in the disease gene GO annotations.
The definition of the non-disease class is not trivial, as it is not possible to know in advance which candidate genes will be discovered as responsible for NSHL in the future – i.e. it is not possible to discriminate disease and non-disease genes among the candidates. To address this issue we decided to use the distribution of SSM scores in the class of candidate genes to define the non-disease class. We considered as non-disease genes the candidate genes with a score less than the 95th percentile of the distribution of candidate gene scores.
The GO terms with a Fisher's test p-value smaller than 0.01 are considered significantly over-represented in the list of the disease genes. These provide indications about the main functions and biological processes involved in the hearing mechanisms, taking into account the 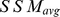 scores computed for our candidate gene list against the NSHL genes at present known.
scores computed for our candidate gene list against the NSHL genes at present known.
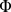 score
score
For both candidate and disease gene lists their over-represented GO terms are weighted taking into account their specificity in the corpus of the GO annotations as follows:
where IC is estimated using the probability of occurrence of the GO terms in the corpus of all gene annotations provided by ENGINE database [41], [42].
Supporting Information
NSHL autosomal dominant loci. Locus names and chromosomal locations have been inferred from literature. References are relative to the articles where the locus association to NSHL was identified.
(0.05 MB PDF)
NSHL autosomal recessive loci. Locus names and chromosomal locations have been inferred from literature. References are relative to the articles where the locus association to NSHL was identified.
(0.05 MB PDF)
NSHL X-linked, Y-linked and modifier loci. Locus names and chromosomal locations have been inferred from literature. References are relative to the articles where the locus association to NSHL was identified.
(0.02 MB PDF)
Acknowledgments
We thank Andreas Gisel and Angelica Tulipano from CNR – ITB Bari for useful discussions and for providing probabilities for IC estimation. We thank Luigi Doronzo from IBM for useful insights and revision of the manuscript. Finally, we thank the members of the research group at Medical Genetics Service, Hospital CSS-IRCCS, San Giovanni Rotondo, Italy, Massimo Carella and Maria Stella Alemanno for useful discussions around the pathology and the gene validation protocol.
Footnotes
Competing Interests: The authors are employed with IBM Italy SpA. They serve as investigators for the research and educational project DM19410 “Laboratorio di Bioinformatica per la Biodiversita' Molecolare” MBLab. The authors confirm their adherence to all the PLoS ONE policies on sharing data and materials.
Funding: The authors acknowledge support of the Ministero dell'Università e della Ricerca (MUR), under the project DM19410 “Laboratorio di Bioinformatica per la Biodiversita' Molecolare” MBLab (www.mblabproject.it). Given the educational spirit of this work and of the entire project, this study was conducted by the authors in total autonomy, and the funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Schrijver I. Hereditary non-syndromic sensorineural hearing loss: transforming silence to sound. J Mol Diagn. 2004;6:275–284. doi: 10.1016/S1525-1578(10)60522-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kelsell DP, Dunlop J, Stevens HP, Lench NJ, Liang JN, et al. Connexin 26 mutations in hereditary non-syndromic sensorineural deafness. Nature. 2009;387:80–83. doi: 10.1038/387080a0. [DOI] [PubMed] [Google Scholar]
- 3.Grifa A, Wagner CA, D'Ambrosio L, Melchionda S, Bernardi F, et al. Mutations in GJB6 cause nonsyndromic autosomal dominant deafness at DFNA3 locus. Nat Genet. 1999;23:16–18. doi: 10.1038/12612. [DOI] [PubMed] [Google Scholar]
- 4.Li XC, Everett LA, Lalwani AK, Desmukh D, Friedman TB, et al. A mutation in PDS causes non-syndromic recessive deafness. Nat Genet. 1998;18:215–217. doi: 10.1038/ng0398-215. [DOI] [PubMed] [Google Scholar]
- 5.Robertson NG, Lu L, Heller S, Merchant SN, Eavey RD, et al. Mutations in a novel cochlear gene cause DFNA9, a human nonsyndromic deafness with vestibular dysfunction. Nat Genet. 1998;20:299–303. doi: 10.1038/3118. [DOI] [PubMed] [Google Scholar]
- 6.De Kok YJ, Van der Maarel SM, Bitner-Glindzicz M, Huber I, Monaco AP, et al. Association between X-linked mixed deafness and mutations in the POU domain gene POU3F4. Science. 1995;267:685–688. doi: 10.1126/science.7839145. [DOI] [PubMed] [Google Scholar]
- 7.Bespalova IN, Van Camp G, Bom SJ, Brown DJ, Cryns K, et al. Mutations in the Wolfram syndrome 1 gene (WFS1) are a common cause of low frequency sensorineural hearing loss. Hum Mol Genet. 2001;10:2501–2508. doi: 10.1093/hmg/10.22.2501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Young TL, Ives E, Lynch E, Person R, Snook S, et al. Non-syndromic progressive hearing loss DFNA38 is caused by heterozygous missense mutation in the Wolfram syndrome gene WFS1. Hum Mol Genet. 2001;10:2509–2514. doi: 10.1093/hmg/10.22.2509. [DOI] [PubMed] [Google Scholar]
- 9.Hereditary Hearing Loss home page. Available: http://herditaryhearingloss.org/. Accessed April 2010.
- 10.Linghu B, Snitkin ES, Hu Z, Xia Y, DeLisi C. Genome-wide prioritization of disease genes and identification of disease-disease associations from an integrated human functional linkage network. Genome Biology. 2009;10:R91. doi: 10.1186/gb-2009-10-9-r91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, et al. Gene Ontology: tool for the unification of biology. the Gene Ontology Consortium. Nat Genet. 2000;25:25–29. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Pesquita C, Faria D, Falcao AO, Lord P, Couto F. Semantic similarity in biomedical ontologies. PLoS Comput Biol. 2009;5:e1000443. doi: 10.1371/journal.pcbi.1000443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lord PW, Stevens RD, Brass A, Goble CA. Semantic similarity measures as tools for exploring the Gene Ontology. Pacific Symposium on Biocomputing. 2003;8:601–612. doi: 10.1142/9789812776303_0056. [DOI] [PubMed] [Google Scholar]
- 14.Wu H, Su Z, Mao F, Olman V, Xu Y. Prediction of functional modules based on comparative genome analysis and gene ontology application. Nucleic Acids Res. 2005;33:2822–2837. doi: 10.1093/nar/gki573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wu X, Zhu L, Guo J, Zhang DY, Lin K. Prediction of yeast protein-protein interaction network: insights from the gene ontology and annotations. Nucleic Acids Res. 2006;34:2137–2150. doi: 10.1093/nar/gkl219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Yu H, Gao L, Tu K, Guo Z. Broadly predicting specific gene functions with expression similarity and taxonomy similarity. Gene. 2005;352:75–81. doi: 10.1016/j.gene.2005.03.033. [DOI] [PubMed] [Google Scholar]
- 17.Cheng J, Cline M, Martin J, Finkelstein D, Awad T, et al. A knowledge-based clustering algorithm driven by gene ontology. Journal of Biopharmaceutical Statistics. 2004;14:687–700. doi: 10.1081/bip-200025659. [DOI] [PubMed] [Google Scholar]
- 18.del Pozo A, Pazos F, Valencia A. Defining functional distances over gene ontology. BMC Bioinformatics. 2008;9:50. doi: 10.1186/1471-2105-9-50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Resnik P. Using information content to evaluate semantic similarity in a taxonomy. Proc of the 14th International Joint Conference on Artificial Intelligence. 1995:448–453. [Google Scholar]
- 20.Couto FM, Silva MJ, Coutinho PM. Semantic similarity over the gene ontology: Family correlation and selecting disjunctive ancestors. 2005. pp. 343–344. In: Proc. of the ACM Conference in Information and Knowledge Management as a short paper. Bremen, Germany.
- 21.Othman R, Deris S, Illias R. A genetic similarity algorithm for searching the gene ontology terms and annotating anonymous protein sequences. J Biomed Inform. 2007;23:529–538. doi: 10.1016/j.jbi.2007.05.010. [DOI] [PubMed] [Google Scholar]
- 22.Jiang J, Conrath D. Semantic similarity based on corpus statistics and lexical taxonomy. 1997. pp. 19–33. In: Proc. of the 10th International Conference on Research on Computational Linguistics. Taiwan.
- 23.Schlicker A, Domingues FS, Rahnenführer J, Lengauer T. A new measure for functional similarity of gene products based on gene ontology. BMC Bioinformatics. 2006;7:302. doi: 10.1186/1471-2105-7-302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bodenreider O, Aubry M, Burgun A. Non-lexical approaches to identifying associative relations in the gene ontology. Pac Symp Biocomput. 2005;7:91–102. [PMC free article] [PubMed] [Google Scholar]
- 25.Lin D. Proc. of the 15th International Conference on Machine Learning. San Francisco, CA: Morgan Kaufmann; 1998. An information-theoretic definition of similarity. pp. 296–304. [Google Scholar]
- 26.Sevilla JL, Segura V, Podhorski A, Guruceaga E, Mato JM, et al. Correlation between gene expression and go semantic similarity. IEEE/ACM Transactions on Computational Biology and Bioinformatics. 2005;2:330–338. doi: 10.1109/TCBB.2005.50. [DOI] [PubMed] [Google Scholar]
- 27.Riensche RM, Baddeley BL, Sanfilippo AP, Posse C, Gopalan B. Xoa: Web-enabled cross-ontological analytics. Services, IEEE Congress. 2007:99–105. [Google Scholar]
- 28.Wang JZZ, Du Z, Payattakool R, Yu PSS, Chen CFF. A new method to measure the semantic similarity of go terms. Bioinformatics. 2007;23:1274–1281. doi: 10.1093/bioinformatics/btm087. [DOI] [PubMed] [Google Scholar]
- 29.Lei Z, Dai Y. Assessing protein similarity with gene ontology and its use in subnuclear localization prediction. BMC Bioinformatics. 2006;7:491. doi: 10.1186/1471-2105-7-491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.R Package Matching. Available: http://sekhon.berkeley.edu/matching.
- 31.Weil D, El-Amraoui A, Masmoudi S, Mustapha M, Kikkawa Y, et al. Usher syndrome type IG (USH1G) is caused by mutations in the gene encoding SANS, a protein that associates with the USH1C protein, harmonin. Hum Molec Genet. 2003;12:463–471. doi: 10.1093/hmg/ddg051. [DOI] [PubMed] [Google Scholar]
- 32.Piccini M, Vitelli F, Seri M, Galietta LJV, Moran O, et al. KCNE1-like gene is deleted in AMME contiguous gene syndrome: Identification and characterization of the human and mouse homologs. Genomics. 1999;60:251–257. doi: 10.1006/geno.1999.5904. [DOI] [PubMed] [Google Scholar]
- 33.Jin H, May M, Tranebjaerg L, Kendall E, Fontan G, et al. A novel X-linked gene, DDP, shows mutations in families with deafness (DFN-1), dystonia, mental deficiency and blindness. Nature Genet. 1996;14:177–180. doi: 10.1038/ng1096-177. [DOI] [PubMed] [Google Scholar]
- 34.Tranebjaerg L, van Ghelue M, Nilssen O, Hodes ME, Dlouhy SR, et al. Jensen syndrome is allelic to Mohr-Tranebjaerg syndrome and both are caused by stop mutations in the DDP gene. (Abstract) Am J Hum Genet. 1997;61(suppl.):A349 only. [Google Scholar]
- 35.Collin GB, Marshall JD, Ikeda A, So WV, Russell-Eggitt I, et al. Mutations in ALMS1 cause obesity, type 2 diabetes and neurosensory degeneration in Alström syndrome. Nature Genet. 2002;31:74–78. doi: 10.1038/ng867. [DOI] [PubMed] [Google Scholar]
- 36.Verpy E, Leibovici M, Zwaenepoel I, Liu XZ, Gal A, et al. A defect in harmonin, a PDZ domain-containing protein expressed in the inner ear sensory hair cells, underlies Usher syndrome type 1C. Nature Genet. 2000;26:51–55. doi: 10.1038/79171. [DOI] [PubMed] [Google Scholar]
- 37.Ouyang XM, Xia XJ, Verpy E, Du LL, Pandya A, et al. Mutations in the alternatively spliced exons of USH1C cause non-syndromic recessive deafness. Hum Genet. 2002;111:26–30. doi: 10.1007/s00439-002-0736-0. [DOI] [PubMed] [Google Scholar]
- 38.Ahmed ZM, Smith TN, Riazuddin S, Makishima T, Ghosh M, et al. Nonsyndromic recessive deafness DFNB18 and Usher syndrome type 1C are allelic mutations of USH1C. Hum Genet. 2002;110:527–531. doi: 10.1007/s00439-002-0732-4. [DOI] [PubMed] [Google Scholar]
- 39.Maglott D, Ostell J, Pruitt KD, Tatusova T. Entrez Gene: gene-centered information at NCBI. Nucleic Acids Res. 2007;35:D26–31. doi: 10.1093/nar/gkl993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tlili A, Männikkö M, Charfedine I, Lahmar I, Benzina Z, et al. A novel autosomal recessive non-syndromic deafness locus, DFNB66, maps to chromosome 6p21.2–22.3 in a large Tunisian consanguineous family. Hum Hered. 2005;60:123–128. doi: 10.1159/000088974. [DOI] [PubMed] [Google Scholar]
- 41.Tulipano A, Donvito G, Licciulli F, Maggi G, Gisel A. Gene analogue finder: a GRID solution for finding functionally analogous gene products. BMC Bioinformatics. 2007;8:329. doi: 10.1186/1471-2105-8-329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.GENe AnaloGue FINdEr ENGINE database: A repository for functional analogous gene products. Available: http://spank.ba.itb.cnr.it/engine/. Accessed June 2009.
- 43.Lynch ED, Lee MK, Morrow JE, Welcsh PL, León PE, et al. Nonsyndromic deafness DFNA1 associated with mutation of a human homolog of the Drosophila gene diaphanous. Science. 1997;278:1315–1318. [PubMed] [Google Scholar]
- 44.Xia JH, Liu CY, Tang BS, Pan Q, Huang L, et al. Mutations in the gene encoding gap junction protein beta-3 associated with autosomal dominant hearing impairment. Nature Genet. 1998;20:370–373. doi: 10.1038/3845. [DOI] [PubMed] [Google Scholar]
- 45.Kubisch C, Schroeder BC, Friedrich T, Lütjohann B, El-Amraoui A, et al. KCNQ4, a novel potassium channel expressed in sensory outer hair cells, is mutated in dominant deafness. Cell. 1999;96:437–446. doi: 10.1016/s0092-8674(00)80556-5. [DOI] [PubMed] [Google Scholar]
- 46.del Castillo I, Villamar M, Moreno-Pelayo MA, del Castillo FJ, Alvarez A, et al. A deletion involving the connexin 30 gene in nonsyndromic hearing impairment. N Engl J Med. 2002;346:243–249. doi: 10.1056/NEJMoa012052. [DOI] [PubMed] [Google Scholar]
- 47.Donaudy F, Snoeckx R, Pfister M, Zenner HP, Blin N, et al. Nonmuscle myosin heavy-chain gene MYH14 is expressed in cochlea and mutated in patients affected by autosomal dominant hearing impairment (DFNA4). Am J of Hum Genet. 2004;74:770–776. doi: 10.1086/383285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Van Laer L, Huizing EH, Verstreken M, van Zuijlen D, Wauters JG, et al. Nonsyndromic hearing impairment is associated with a mutation in DFNA5. Nature Genet. 1998;20:194–197. doi: 10.1038/2503. [DOI] [PubMed] [Google Scholar]
- 49.Verhoeven K, Van Laer L, Kirschhofer K, Legan PK, Hughes DC, et al. Mutations in the human alpha-tectorin gene cause autosomal dominant non-syndromic hearing impairment. Nature Genet. 1998;19:60–62. doi: 10.1038/ng0598-60. [DOI] [PubMed] [Google Scholar]
- 50.Wayne S, Robertson NG, DeClau F, Chen N, Verhoeven K. Mutations in the transcriptional activator EYA4 cause late-onset deafness at the DFNA10 locus. Hum Mol Genet. 2001;10:195–200. doi: 10.1093/hmg/10.3.195. [DOI] [PubMed] [Google Scholar]
- 51.Liu XZ, Walsh J, Mburu P, Kendrick-Jones J, Cope MJ, et al. Mutations in the myosin VIIA gene cause non-syndromic recessive deafness. Nat Genet. 1997;16:188–190. doi: 10.1038/ng0697-188. [DOI] [PubMed] [Google Scholar]
- 52.McGuirt WT, Prasad SD, Griffith AJ, Kunst HP, Green GE, et al. Mutations in COL11A2 cause non-syndromic hearing loss (DFNA13). Hum Mol Genet. 1999;23:413–419. doi: 10.1038/70516. [DOI] [PubMed] [Google Scholar]
- 53.Vahava O, Morell R, Lynch ED, Weiss S, Kagan ME, et al. Mutation in transcription factor POU4F3 associated with inherited progressive hearing loss in humans. Science. 1998;279:1950–1954. doi: 10.1126/science.279.5358.1950. [DOI] [PubMed] [Google Scholar]
- 54.Lalwani AK, Goldstein JA, Kelley MJ, Luxford W, Castelein CM, et al. Human nonsyndromic hereditary deafness DFNA17 is due to a mutation in nonmuscle myosin MYH9. Am J Hum Genet. 2000;67:1121–1128. doi: 10.1016/s0002-9297(07)62942-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Zhu M, Yang T, Wei S, DeWan AT, Morell RJ, et al. Mutations in the gamma-actin gene (ACTG1) are associated with dominant progressive deafness (DFNA20/26). Am J Hum Genet. 2003;73:1082–1091. doi: 10.1086/379286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.van Wijk E, Krieger E, Kemperman MH, De Leenheer EM, Huygen PL, et al. A mutation in the gamma actin 1 (ACTG1) gene causes autosomal dominant hearing loss (DFNA20/26). J Med Genet. 2003;40:879–884. doi: 10.1136/jmg.40.12.879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Melchionda S, Ahituv N, Bisceglia L, Sobe T, Glaser F, et al. MYO6, the human homologue of the gene responsible for deafness in Snell's waltzer mice, is mutated in autosomal dominant nonsyndromic hearing loss. Am J Hum Genet. 2001;69:635–640. doi: 10.1086/323156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Peters LM, Anderson DW, Griffith AJ, Grundfast KM, San Agustin TB, et al. Mutation of a transcription factor, TFCP2L3, causes progressive autosomal dominant hearing loss, DFNA28. Hum Mol Genet. 2002;11:2877–2885. doi: 10.1093/hmg/11.23.2877. [DOI] [PubMed] [Google Scholar]
- 59.Kurima K, Peters LM, Yang Y, Riazuddin S, Ahmed ZM, et al. Dominant and recessive deafness caused by mutations of a novel gene, TMC1, required for cochlear hair-cell function. Nat Genet. 2002;30:277–284. doi: 10.1038/ng842. [DOI] [PubMed] [Google Scholar]
- 60.Abe S, Katagiri T, Saito-Hisaminato A, Usami S, Inoue Y. Identification of CRYM as a candidate responsible for nonsyndromic deafness, through cDNA microarray analysis of human cochlear and vestibular tissues. Am J Hum Genet. 2003;72:73–82. doi: 10.1086/345398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Modamio-Høybjør S, Mencia A, Goodyear R, del Castillo I, Richardson G, et al. A mutation in CCDC50, a gene encoding an effector of epidermal growth factor-mediated cell signaling, causes progressive hearing loss. Am J Hum Genet. 2007;80:1076–1089. doi: 10.1086/518311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Donaudy F, Ferrara A, Esposito L, Hertzano R, Ben-David O, et al. Multiple mutations of MYO1A, a cochlear-expressed gene, in sensorineural hearing loss. Hum Mol Genet. 2003;72:1571–1577. doi: 10.1086/375654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Yang T, Gurrola JGn, Wu H, Chiu SM, Wangemann P, et al. Mutations of KCNJ10 together with mutations of SLC26A4 cause digenic nonsyndromic hearing loss associated with enlarged vestibular aqueduct syndrome. Am J Hum Genet. 2009;84:651–657. doi: 10.1016/j.ajhg.2009.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mencía A, Modamio-Høybjør S, Redshaw N, Morín M, Mayo-Merino F. Mutations in the seed region of human miR-96 are responsible for nonsyndromic progressive hearing loss. Nat Genet. 2009;41:609–613. doi: 10.1038/ng.355. [DOI] [PubMed] [Google Scholar]
- 65.Wang A, Liang Y, Fridell RA, Probst FJ, Wilcox ER, et al. Association of unconventional myosin MYO15 mutations with human nonsyndromic deafness DFNB3. Science. 1998;280:1447–1451. doi: 10.1126/science.280.5368.1447. [DOI] [PubMed] [Google Scholar]
- 66.Naz S, Giguere CM, Kohrman DC, Mitchem KL, Riazuddin S, et al. Mutations in a novel gene, TMIE, are associated with hearing loss linked to the DFNB6 locus. Am J Hum Genet. 2002;71:632–636. doi: 10.1086/342193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Scott HS, Kudoh J, Wattenhofer M, Shibuya K, Berry A, et al. Insertion of beta-satellite repeats identifies a transmembrane protease causing both congenital and childhood onset autosomal recessive deafness. Nat Genet. 2002;27:59–63. doi: 10.1038/83768. [DOI] [PubMed] [Google Scholar]
- 68.Yasunaga S, Grati M, Cohen-Salmon M, El-Amraoui A, Mustapha M, et al. A mutation in OTOF, encoding otoferlin, a FER-1-like protein, causes DFNB9, a nonsyndromic form of deafness. Nat Genet. 1999;21:363–369. doi: 10.1038/7693. [DOI] [PubMed] [Google Scholar]
- 69.Bork JM, Peters LM, Riazuddin S, Bernstein SL, Ahmed ZM, et al. Usher syndrome 1D and nonsyndromic autosomal recessive deafness DFNB12 are caused by allelic mutations of the novel cadherin-like gene CDH23. Am J Hum Genet. 2001;68:26–37. doi: 10.1086/316954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Verpy E, Masmoudi S, Zwaenepoel I, Leibovici M, Hutchin TP, et al. Mutations in a new gene encoding a protein of the hair bundle cause non-syndromic deafness at the DFNB16 locus. Nat Genet. 2001;29:345–349. doi: 10.1038/ng726. [DOI] [PubMed] [Google Scholar]
- 71.Zwaenepoel I, Mustapha M, Leibovici M, Verpy E, Goodyear R, et al. Otoancorin, an inner ear protein restricted to the interface between the apical surface of sensory epithelia and their overlying acellular gels, is defective in autosomal recessive deafness DFNB22. Proc Natl Acad Sci U S A. 2002;99:6240–6245. doi: 10.1073/pnas.082515999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Ahmed ZM, Riazuddin S, Ahmad J, Bernstein SL, Guo Y, et al. PCDH15 is expressed in the neurosensory epithelium of the eye and ear and mutant alleles are responsible for both USH1F and DFNB23. Hum Mol Genet. 2003;12:3215–3223. doi: 10.1093/hmg/ddg358. [DOI] [PubMed] [Google Scholar]
- 73.Khan SY, Ahmed ZM, Shabbir MI, Kitajiri S, Kalsoom S, et al. Mutations of the RDX gene cause nonsyndromic hearing loss at the DFNB24 locus. Hum Mutat. 2007;28:417–423. doi: 10.1002/humu.20469. [DOI] [PubMed] [Google Scholar]
- 74.Shahin H, Walsh T, Sobe T, Abu Sa'ed J, Abu Rayan A, et al. Mutations in a novel isoform of TRIOBP that encodes a filamentous-actin binding protein are responsible for DFNB28 recessive nonsyndromic hearing loss. Am J Hum Genet. 2006;78:144–152. doi: 10.1086/499495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Riazuddin S, Khan SN, Ahmed ZM, Ghosh M, Caution K, et al. Mutations in TRIOBP, which encodes a putative cytoskeletal-organizing protein, are associated with nonsyndromic recessive deafness. Am J Hum Genet. 2006;78:137–143. doi: 10.1086/499164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Wilcox ER, Burton QL, Naz S, Riazuddin S, Smith TN, et al. Mutations in the gene encoding tight junction claudin-14 cause autosomal recessive deafness DFNB29. Cell. 2001;104:165–172. doi: 10.1016/s0092-8674(01)00200-8. [DOI] [PubMed] [Google Scholar]
- 77.Walsh T, Walsh V, Vreugde S, Hertzano R, Shahin H, et al. From flies' eyes to our ears: mutations in a human class iii myosin cause progressive nonsyndromic hearing loss DFNB30. Proc Natl Acad Sci U S A. 2002;99:7518–7523. doi: 10.1073/pnas.102091699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Mburu P, Mustapha M, Varela A, Weil D, El-Amraoui A, et al. Defects in whirlin, a PDZ domain molecule involved in stereocilia elongation, cause deafness in the whirler mouse and families with DFNB31. Nat Genet. 2003;34:421–428. doi: 10.1038/ng1208. [DOI] [PubMed] [Google Scholar]
- 79.Ansar M, Din MA, Arshad M, Sohail M, Faiyaz-Ul-Haque M, et al. A novel autosomal recessive non-syndromic deafness locus (DFNB35) maps to 14q24.1–14q24.3 in large consanguineous kindred from Pakistan. Eur J Hum Genet. 2003;11:77–80. doi: 10.1038/sj.ejhg.5200905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Collin RW, Kalay E, Tariq M, Peters T, van der Zwaag B, et al. Mutations of ESRRB encoding estrogen-related receptor beta cause autosomal-recessive nonsyndromic hearing impairment DFNB35. Am J Hum Genet. 2008;82:125–138. doi: 10.1016/j.ajhg.2007.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Naz S, Griffith AJ, Riazuddin S, Hampton LL, Battey JFJ, et al. Mutations of ESPN cause autosomal recessive deafness and vestibular dysfunction. J Med Genet. 2004;41:591–595. doi: 10.1136/jmg.2004.018523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Schultz JM, Khan SN, Ahmed ZM, Riazuddin S, Waryah AM, et al. Noncoding mutations of HGF are associated with nonsyndromic hearing loss, DFNB39. Am J Hum Genet. 2009;85:25–39. doi: 10.1016/j.ajhg.2009.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Abe S, Usami S, Nakamura Y. Mutations in the gene encoding KIAA1199 protein, an inner-ear protein expressed in deiters' cells and the fibrocytes, as the cause of nonsyndromic hearing loss. J Hum Genet. 2003;48:564–570. doi: 10.1007/s10038-003-0079-2. [DOI] [PubMed] [Google Scholar]
- 84.Riazuddin S, Ahmed ZM, Fanning AS, Lagziel A, Kitajiri S. Tricellulin is a tight-junction protein necessary for hearing. Am J Hum Genet. 2006;79:1040–1051. doi: 10.1086/510022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Delmaghani S, del Castillo FJ, Michel V, Leibovici M, Aghaie A. Mutations in the gene encoding pejvakin, a newly identified protein of the afferent auditory pathway, cause DFNB59 auditory neuropathy. Nat Genet. 2006;38:770–778. doi: 10.1038/ng1829. [DOI] [PubMed] [Google Scholar]
- 86.Liu XZ, Ouyang XM, Xia XJ, Zheng J, Pandya A, et al. Prestin, a cochlear motor protein, is defective in non-syndromic hearing loss. Hum Mol Genet. 2003;12:1155–1162. doi: 10.1093/hmg/ddg127. [DOI] [PubMed] [Google Scholar]
- 87.Ahmed ZM, Masmoudi S, Kalay E, Belyantseva IA, Mosrati MA, et al. Mutations of LRTOMT, a fusion gene with alternative reading frames, cause nonsyndromic deafness in humans. Nat Genet. 2008;40:1335–40. doi: 10.1038/ng.245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Shabbir MI, Ahmed ZM, Khan SY, Riazuddin S, Waryah AM. Mutations of human TMHS cause recessively inherited non-syndromic hearing loss. J Med Genet. 2006;43:634–640. doi: 10.1136/jmg.2005.039834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Kalay E, Li Y, Uzumcu A, Uyguner O, Collin RW. Mutations in the lipoma HMGIC fusion partner-like 5 (LHFPL5) gene cause autosomal recessive nonsyndromic hearing loss. Hum Mutat. 2006;27:633–639. doi: 10.1002/humu.20368. [DOI] [PubMed] [Google Scholar]
- 90.Liu X, Han D, Li J, Han B, Ouyang X. Loss-of-function mutations in the PRPS1 gene cause a type of nonsyndromic X-linked sensorineural deafness, DFN2. Am J Hum Genet. 2010;86:65–71. doi: 10.1016/j.ajhg.2009.11.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Schultz JM, Yang Y, Caride AJ, Filoteo AG, Penheiter AR. Modification of human hearing loss by plasma-membrane calcium pump PMCA2. N Engl J Med. 2005;352:1557–1564. doi: 10.1056/NEJMoa043899. [DOI] [PubMed] [Google Scholar]
- 92.Ficarella R, Di Leva F, Bortolozzi M, Ortolano S, Donaudy F. A functional study of plasma-membrane calcium-pump isoform 2 mutants causing digenic deafness. Proc Natl Acad Sci U S A. 2007;104:1516–1521. doi: 10.1073/pnas.0609775104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Li G, Vega R, Nelms K, Gekakis N, Goodnow C, et al. A role for Alström syndrome protein, alms1, in kidney ciliogenesis and cellular quiescence. PLoS Genet. 2007;3:e8. doi: 10.1371/journal.pgen.0030008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Karamatic Crew V, Burton N, Kagan A, Green CA, Levene C, et al. CD151, the first member of the tetraspanin (TM4) superfamily detected on erythrocytes, is essential for the correct assembly of human basement membranes in kidney and skin. Blood. 2004;104:2217–2223. doi: 10.1182/blood-2004-04-1512. [DOI] [PubMed] [Google Scholar]
- 95.Stover EH, Borthwick KJ, Bavalia C, Eady N, Fritz DM, et al. Novel ATP6V1B1 and ATP6V0A4 mutations in autosomal recessive distal renal tubular acidosis with new evidence for hearing loss. J Med Genet. 2002;39:796–803. doi: 10.1136/jmg.39.11.796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Wangemann P. Supporting sensory transduction: cochlear fluid homeostasis and the endocochlear potential. J Physiol. 2006;576:11–21. doi: 10.1113/jphysiol.2006.112888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Bulfone A, Menguzzato E, Broccoli V, Marchitiello A, Gattuso C, et al. Barhl1, a gene belonging to a new subfamily of mammalian homeobox genes, is expressed in migrating neurons of the CNS. Hum Mol Genet. 2000;9:1443–1452. doi: 10.1093/hmg/9.9.1443. [DOI] [PubMed] [Google Scholar]
- 98.NCBI UniGene: Organized view of the transcriptome. Available: http://www.ncbi.nlm.nih.gov/unigene. Accessed April 2010.
- 99.UniProtKB (Universal Protein resource KnowledgeBase) Available: http://www.uniprot.org/. Accessed April 2010.
- 100.HPRD (Human Protein Reference Database) Available: http://www.hprd.org/. Accessed April 2010.
- 101.The Morton Lab: Cochlear EST database. Available: http://www.brighamandwomens.org/bwh_hearing/default.aspx. Accessed January 2009.
- 102.NCBI GEO: Gene Expression Omnibus Database. Available: http://www.ncbi.nlm.nih.gov/gds. Accessed April 2010.
- 103.IHR (Institute of Hearing Research) Table of gene expression in the developing ear. Available: http://www.ihr.mrc.ac.uk/. Accessed April 2010.
- 104.Bgee (A dataBase for Gene Expression Evolution) Available: http://bgee.unil.ch. Accessed April 2010.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
NSHL autosomal dominant loci. Locus names and chromosomal locations have been inferred from literature. References are relative to the articles where the locus association to NSHL was identified.
(0.05 MB PDF)
NSHL autosomal recessive loci. Locus names and chromosomal locations have been inferred from literature. References are relative to the articles where the locus association to NSHL was identified.
(0.05 MB PDF)
NSHL X-linked, Y-linked and modifier loci. Locus names and chromosomal locations have been inferred from literature. References are relative to the articles where the locus association to NSHL was identified.
(0.02 MB PDF)