

Abstract
Objectives
In cyanotic patients undergoing repair of heart defects, chronic hypoxia is thought to lead to greater susceptibility to ischemia and reoxygenation injury. We sought to find an explanation to such a hypothesis by investigating the cardiac gene expression in patients with tetralogy of Fallot undergoing cardiac surgery.
Methods
The myocardial gene profile was investigated in right ventricular biopsy specimens obtained from 20 patients with a diagnosis of cyanotic (n = 11) or acyanotic (n = 9) tetralogy of Fallot undergoing surgical repair. Oligonucleotide microarray analyses were performed on the samples, and the array results were validated with Western blotting and enzyme-linked immunosorbent assay.
Results
Data revealed 795 differentially expressed genes in cyanotic versus acyanotic hearts, with 198 upregulated and 597 downregulated. Growth/morphogenesis, remodeling, and apoptosis emerged as dominant functional themes for the upregulated genes and included the apoptotic gene TRAIL (tumor necrosis factor–related apoptosis-inducing ligand), the remodeling factor OPN (osteopontin), and the mitochondrial function gene COX11 (cytochrome-c oxidase 11). In contrast, transcription, mitogen-activated protein kinase signaling, and contractile machinery were the dominant functional classes for the downregulated genes, which included the calcium-handling gene NCX1 (sodium-calcium exchanger). Protein levels of COX11, NCX1, OPN, and LYZ (lysozyme) in the myocardium followed the same pattern obtained by means of transcriptomics. The TRAIL level did not change in myocardium but increased in circulating blood of cyanotic patients, suggesting the myocardium as a possible source. Additionally, our data showed increased protein expression of apoptosis markers in cyanotic myocardium.
Conclusions
Chronic hypoxia in cyanotic children with tetralogy of Fallot induced the expression of genes associated with apoptosis and remodeling and reduced the expression of genes associated with myocardium contractility and function.
Cyanotic pediatric patients undergoing cardiac surgery are exposed to a chronic hypoxic state that can reduce their antioxidant reserve capacity, leading to a greater susceptibility to the oxidative stress of ischemia and reperfusion at the time of surgical correction.1-3 Our group has shown that cyanotic children have worse myocardial reperfusion injury and clinical outcomes compared with acyanotic children after similar periods of ischemic cardioplegic arrest.4 We have also shown that reintroduction of high oxygen levels to cyanotic patients undergoing cardiopulmonary bypass (CPB) leads to myocardial damage before ischemic cardioplegic arrest.5 Our results suggest that the injury seen after cardioplegic arrest might in part be due to CPB-induced reoxygenation injury, and this has also been demonstrated in previous studies.6
Several recent studies have used microarray technology to examine the global myocardial stress response during cardiac surgery, focusing on the human congenitally malformed hearts.7-10 Kaynak and colleagues7 examined gene expression profiles in patients with tetralogy of Fallot (TOF), ventricular septal defect, and right ventricular hypertrophy (RVH) compared with profiles seen in those with normal hearts. Another investigation examined gene expression changes in patients with cyanotic TOF in comparison with those seen in patients with normal hearts and showed that the upregulation of genes encoding vascular endothelial growth factor (VEGF) and extracellular matrix proteins are the key events contributing to RVH and stunted angiogenesis in patients with TOF.10 Konstantinov and associates8 examined the gene expression profiles in children undergoing cardiac surgery for right heart obstructive lesions, including TOF, and showed that neonatal myocardium has a unique pattern of gene expression dominated by genes with cardioprotective, antihypertrophic, and antiproliferative properties, reflecting a stress-induced protective program.
However, to our knowledge, no previous genome-wide investigation has been made to determine the global gene expression profiles associated with chronic hypoxia in patients with TOF. The aim of this study was therefore to investigate the global ventricular myocardial gene expression profiles of cyanotic and acyanotic patients with a diagnosis of TOF undergoing corrective cardiac surgery.
MATERIALS AND METHODS
Twenty patients with a diagnosis of cyanotic (n = 11) or acyanotic (n = 9) TOF undergoing surgical repair at the Bristol Royal Hospital for Children were studied. All patients were in stable condition without preoperative respiratory or inotropic support. Preoperative characteristics in the 2 groups are summarized in Table 1. Patients with cyanotic TOF presented with several episodes of spells and repeated saturation measurements of less than 90% (mean, 79.6% ± 7.5%), whereas patients with acyanotic TOF did not have a history of cyanotic spells and presented with saturations of greater than 90% (mean, 94.2% ± 3.5%). Preoperative echocardiographic data showed significant gradient across the right ventricular outflow tract in both groups. The study was approved by the hospital research ethics committee, and parental informed consent was gained for all patients. Intraoperative anesthetic and operative techniques were standardized, as previously reported.4 After the operation, all patients were admitted to the pediatric intensive care unit and were managed according to unit protocols.4,5
TABLE 1. Baseline patients’ characteristics and clinical outcomes.
| Cyanotic patients (n = 11) |
Acyanotic patients (n = 9) |
P value |
|
|---|---|---|---|
| Age (mo) | 10.6 ± 5.5 | 9.5 ± 2.3 | .1 |
| Weight (kg) | 8.04 ± 1.7 | 8.01 ± 1.0 | .7 |
| RVOT velocity (m/s) | 4.6 ± 0.4 | 4.4 ± 0.7 | .2 |
| RV wall indexed (mm/m2) | 16.6 ± 3.0 | 17.1 ± 1.9 | .1 |
| VSD size (mm) | 9.9 ± 1.3 | 9.8 ± 1.6 | .1 |
| Use of preoperative β-blockers | 4 | 1 | .05 |
| Preoperative O2 saturation (%) | 79.6 ± 7.5 | 94.2 ± 3.5 | .01 |
| Crossclamp time (min) | 78.0 ± 15.6 | 73.0 ± 20.7 | .1 |
| CPB time (min) | 132.0 ± 29.0 | 90.2 ± 17.7 | .05 |
| In-hospital mortality | 0 | 0 | |
| Inotropic support (minimal/significant) |
1/10 | 7/2 | .01 |
| Inotropic duration (h) | 54 (42–136) | 22 (12–29) | .03 |
| Total μg dopamine/kg (× 103) | 16.3 (12.2–36.3) | 5.2 (1.9–7.3) | .01 |
| Postoperative ventilation time (h) | 42 (24–77) | 18 (5–33) | .01 |
| Postoperative ICU stay (h) | 87 (49–122) | 40 (26–48) | .001 |
| Postoperative hospital stay (d) | 11 (9–12) | 7 (5–9) | .05 |
Data are presented as means ± standard deviations or medians (interquartile ranges). RVOT, Right ventricular outflow tract; RV, right ventricular; VSD, Ventricular septal defect; ICU, intensive care unit.
Postoperative inotropic support was considered to be either minimal (dopamine ≤5 kg−1 · min−1 for ≤24 hours) or significant (dopamine >5 μg · kg−1 · min−1 with or without other inotropic agents, such as adrenaline, nor-adrenaline, or milrinone). Decisions regarding inotropic support and ventilation were based on hemodynamic status (eg, low mixed venous saturation and high lactic acidosis) and clinical judgment.4,5
Cardiac Muscle Biopsy Specimens
Ventricular biopsy specimens (10 mg net weight) were collected from the apex of the right ventricle by using a “Trucut” needle4 immediately after institution of CPB. Each specimen was immediately put in RNA Later solution (Qiagen, Crawley, United Kingdom) and kept overnight at 4°C. The next day, RNA Later solution was removed, and biopsy specimens were kept at −80°C until RNA extraction.
The concentration of total RNA samples was assessed by means of spectrophotometry (Nanodrop, Wilmington, Del) and was further analyzed for integrity with a Bioanalyzer 2100 with RNA 6000 Nano Assay (Agilent Technologies, Stockport, United Kingdom).
Gene Microarrays: Data Analysis
Ventricular total RNAs (1 μg) from individual patients were processed as previously described.11 Raw data (CEL files) were uploaded into ArrayStar software version 2.1 (DNASTAR, Inc, Madison, Wis) for normalization and statistical analysis. The robust multichip analysis algorithm was used for background correction, quantile normalization, and median polish summarization. The statistical analysis was carried out with ArrayStar software. A Student’s t test and Benjamini and Hochberg multiple testing corrections for false discovery rate (FDR) were used. The FDR-adjusted P value was set to less than .05, and transcripts were filtered on the basis of 1.8-fold or greater difference.
Functional Annotation and Network Analysis
Differentially expressed genes were analyzed according to predefined pathways and functional categories annotated by Kyoto Encyclopedia of Genes and Genomes and Gene Ontology by using the Database for Annotation, Visualization, and Integrated Discovery bioinformatics resource.12 For an overrepresented Gene Ontology or Kyoto Encyclopedia of Genes and Genomes pathway, cutoff P values of .01 and .05 have been selected for the downregulated and upregulated genes, respectively. For Tables E1 and E2, all gene annotations were checked by using online tools and databases: Entrez Gene (www.ncbi.nlm.nih.gov/sites/entrez). The software PathwayStudio (Ariadne, Rockville, Md) and the ResNet database were used to explore the networks of interactions in which the physiologically regulated genes are potentially involved.
Immunohistochemistry and Western Blotting
Right ventricular specimens were fixed in 4% paraformaldehyde, washed in PBS, and embedded in paraffin, and 4-μm sections were obtained. Immunohistochemistry was performed with the ABC-Kit from DakoCytomation (Glostrup, Denmark). Photos were taken at 40× magnification.
Western blotting was performed as previously described.13 Statistical analysis (unpaired t test) was carried out with Instat 3 software (GraphPad Software, Inc, La Jolla, Calif), and a P value was calculated for comparison.
Enzyme-Linked Immunosorbent Assay Protein Analysis
Blood was collected from all patients before surgical intervention, and plasma was separated and stored at −80°C. Human TRAIL levels were measured in plasma samples by using the Diaclone enzyme-linked immunosorbent assay (ELISA) kit. Each sample was performed in duplicate. Statistical analysis (unpaired t test) was carried out with Instat 3 software, and a P value was calculated for comparison.
Statistical Analysis
For the clinical data, continuous variables were summarized by using means and standard deviations (or medians and interquartiles range if the distribution was skewed), and categorical data were summarized as numbers and percentages. Differences between groups were compared by using the χ2 test for categorical variables and the t or Wilcoxon rank sum tests, as appropriate, for continuous variables.
RESULTS
Clinical data are summarized in Table 1. The 2 groups were comparable in terms of age, weight, and crossclamp time. There were no deaths and no major morbidity in both groups. One patient in the cyanotic group required a pacemaker but regained sinus rhythm at follow-up. One patient in the acyanotic group required another run of CPB to resect more muscle from the right ventricular outflow tract after the first procedure. Echocardiographic analysis at discharge did not show any significant residual gradient across the right ventricular outflow tract (mean velocity <3 m/s in both groups) or residual ventricular septal defects. CPB time was longer in the cyanotic group, who required more extensive transannular patching and reconstruction of the pulmonary artery compared with the acyanotic patients. Cyanotic patients had longer inotropic support, ventilation time, and intensive care and hospital stays compared with the acyanotic group.
Gene Microarray Analysis
Analysis of differentially expressed genes
We have used the Affymetrix GeneChip Human Genome U133 Plus 2.0 Array in this study because it represents the most comprehensive whole human genome expression array, providing coverage of more than 47,000 well-substantiated human genes (www.affymetrix.com). Of the genes examined, 795 were identified as differentially expressed in cyanotic versus acyanotic heart biopsy specimens (P < .05, t test followed by Benjamini and Hochberg FDR correction), with 198 (25%) upregulated and 597 (75%) downregulated by more than 1.8-fold (Figure 1, A, and Tables E1 and E2). Genes identified as upregulated or downregulated beyond 1.8-fold unmasked overrepresented biologic process in each gene set (Figure 1, B and C). Myocardial contractility and function, as identified by means of cell adhesion, cytoskeletal organization, transcription regulation, guanosine triphosphatase signal transduction, mitogen-activated protein kinase (MAPK) signaling, and intracellular transport, were downregulated in cyanotic hearts (Figure 1, C). Upregulated transcripts functionally overrepresented apoptosis, cell movement, morphogenesis, and development (Figure 1, B). Collectively, these changes indicate genetic remodeling of the cyanotic myocardium in response to chronic hypoxia.
FIGURE 1.
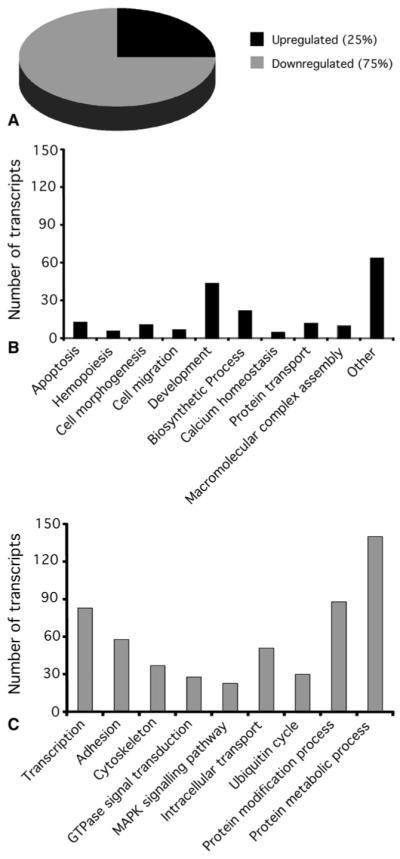
Enrichment analysis of functional groups within the genes differentially expressed in cyanotic compared with acyanotic patients. A, Pie chart of the regulated genes in cyanotic compared with acyanotic patients (≥1.8-fold). Three quarters of the genes were downregulated and a quarter were upregulated. B and C, Selected GO annotations of genes upregulated (B) and downregulated (C) in cyanotic patients. The histograms show the distribution of these annotations.
Pathway analysis
Using Pathway Studio software (Ariadne), we further analyzed the differentially expressed genes (cyanotic vs acyanotic). The analysis looked for the direct pathways connecting genes. For the upregulated genes, the resulting biological association networks are presented in Figure 2. Interestingly, the upregulated genes spanned almost all the cellular organelles. Analysis of downregulated genes associated with the ontological “regulation of transcription” and “MAPK signaling pathway” classes were composed of 83 and 23 downregulated genes, respectively. Of these, 38 and 18 genes, respectively, integrated into direct biological association networks (Figure 3), whereas 45 and 5 genes did not possess interactions.
FIGURE 2.
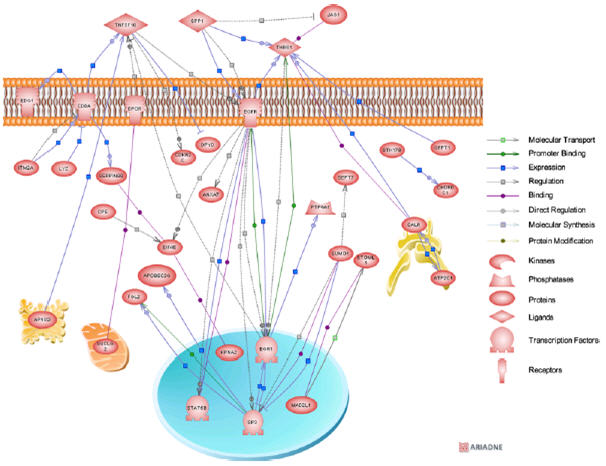
Biological association network of the significantly upregulated genes (>1.8-fold) of cyanotic versus acyanotic patient comparisons. The association between the network entities were based on available PubMed citations. Each node in the network map is linked to an html page showing the protein annotation based on available public databases. Small square boxes connecting 2 nodes are linked to an html page showing the type of effect (positive or negative) and the supporting Medline references.
FIGURE 3.
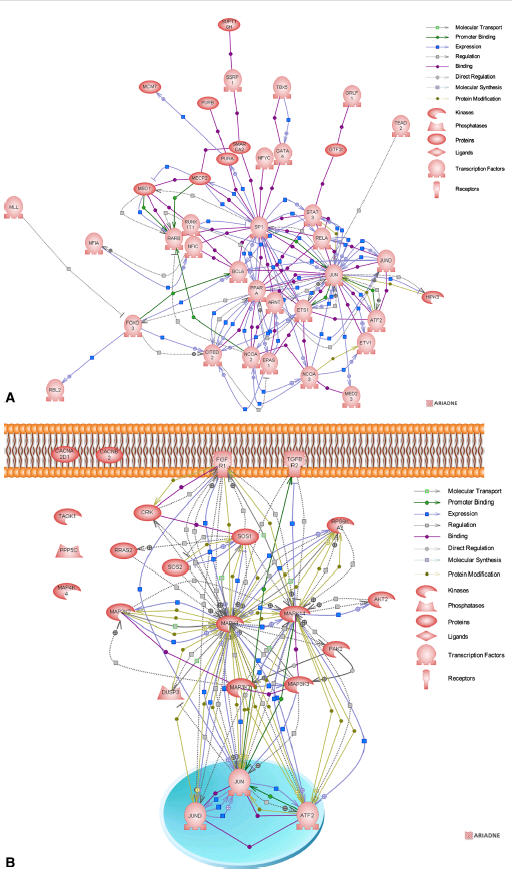
Biological association networks of the significantly downregulated genes associated with the ontological “regulation of transcription” (A) and “MAPK signaling pathway” (B) classes. Each node in network maps is linked to an html page showing the protein annotation based on available public databases. Small square boxes connecting 2 nodes are linked to an html page showing the type of effect (positive or negative) and the supporting Medline references.
Cyanosis increases growth, morphogenesis, and remodeling processes and apoptosis signaling
Increased expression of growth, morphogenesis, and remodeling signaling genes emerged as a dominant functional theme for the upregulated genes. This was evident by the increased expression levels of the growth signaling factors FGF7, TGFBR2, EGFR, and insulin-like growth factor binding protein 7 and the remodeling factors SPP1 (OPN) and ACTL6A (Table E1 and Figure 2).
Another dominant functional theme for the upregulated genes was apoptosis. Indeed, our data showed increased expression levels of genes involved in apoptosis signaling, such as TNFSF10 (TRAIL), B-cell chronic lymphocytic leukemia/lymphoma 10 (BCL10), STAT5B, GULP1 (CED6), and STK17B (DRAK2; Table E1 and Figure 2).
Cyanosis reduces transcription, MAPK signaling, and contractile machinery
One of the dominant functional classes for the downregulated genes was transcription. The transcription factors that showed a reduction of expression levels included JUN, JUND, ATF2, STAT3, GATA4, EPAS1, NCOA2, ARNT, and SP1 (Table E2 and Figure 3, A).
Reduction of MAPK signaling genes emerged as another important functional theme for the downregulated genes. This was evident by the decreased expression levels of MAPK1 (ERK), MAPK14 (SAPK2), MAP3K2, MAP3K3, MAP4K4, AKT2, TAOK1, SOS1, and SOS2 (Table E2 and Figure 3, B). In addition, our data exhibited a downregulation in the contractile machinery (cytoskeleton and cell-adhesion clusters). The downregulated transcripts included troponin T type 2, myosin XVIIIB, ARP2 actin-related protein 2 and nebulin-related anchoring protein (Figure E1).
Validation of Differential Expression of Selected Gene Products
Despite being greatly informative, changes in mRNA levels are insufficient to predict protein expression levels. We therefore went onto assessing the protein levels of the identified genes in the myocardium and blood of cyanotic and acyanotic children using immunohistochemistry, Western blotting, and ELISA. By using immunohistochemistry, we showed COX11, NCX1, OPN, TRAIL, and LYZ protein immunostaining in the myocardium of cyanotic and acyanotic children (Figures E2, C and D; E3, A and B, E4, C and D; and E5, A and B). The strongest signals observed were those of COX11 and LYZ. Subsequently, we semi-quantitatively assessed protein level changes in the myocardium of cyanotic and acyanotic children by using Western blotting. COX11, OPN, and LYZ showed significant increases in cyanotic samples compared with levels seen in acyanotic samples (Figures E2, E and F; E5, C and D; and E6, C and D), which is similar to that observed at the mRNA level. NCX1 protein levels showed a significant decrease caused by cyanosis (Figure E3, C and D), resembling that obtained at the transcriptomic level. In contrast, TRAIL protein levels showed no alteration in the myocardium of the 2 groups of patients (Figure E4, A and B). Because TRAIL is a soluble protein that could be released in blood, we assessed TRAIL plasma levels by means of ELISA. Our data indicated a significant 42% increase of TRAIL plasma levels in cyanotic compared with acyanotic children (Figure E4, E).
Functional Validation
To functionally validate our transcriptomics-derived hypothesis, we examined protein expression levels of markers of hypoxia, apoptosis, and MAPK signaling activation in myocardial tissue. Western blotting of hypoxia-inducible factor 1 alpha showed a significant upregulation in cyanotic compared with acyanotic tissue, confirming the hypoxic state this patient group’s experiences (Figure E7, A). Additionally, we showed a significant increase in cleaved caspase-3 (activated form) and Bcl-2-associated X protein (BAX) protein expression in cyanotic compared with acyanotic myocardium (Figure E7, B and C). Caspase-3 is a critical apoptosis executioner, and BAX is a well-documented apoptotic activator. Increased levels of cleaved caspase-3 and BAX in cyanotic myocardium indicate an increased apoptotic activity in the hearts of these patients. Furthermore, we showed a significant decrease in phospho-SAPK (activated form) expression levels in cyanotic compared with acyanotic myocardium (Figure E7, D). We also detected a tendency toward a decrease in phosphorylated extracellular signal-regulated kinase (ERK) levels in cyanotic myocardium, although it was not significant (data not shown).
DISCUSSION
The results of this microarray-based gene expression profiling study seem to confirm the existence of a reprogramming response that is most evident in the chronically hypoxic cyanotic myocardium. Growth/morphogenesis, remodeling, and apoptosis emerged as dominant functional themes for the up-regulated genes (Figures 1 and 2). In contrast, transcription, MAPK signaling, and contractile machinery (cytoskeleton and adhesion) were the dominant functional classes for the downregulated genes (Figures 1 and 3).
Previous studies have shown RVH in patients with TOF compared with that seen in healthy control subjects.7,10 These studies did not differentiate between cyanotic and acyanotic TOF. Our clinical data did not indicate a higher RVH in cyanotic patients, suggesting that the observed transcriptomic changes are mainly related to cyanosis. In our investigation the expression level changes of extracellular matrix factors (remodeling markers) in cyanotic compared with acyanotic patients went in both directions. Indeed, levels of FBLN1 and SPP1, for example, increased in cyanotic patients, whereas levels of COL3A1, COL4A1, COL4A2, COL6A1, COLA6A2, and COL12A1 decreased (Figure E8). Increased levels of collagen I and III have been shown to be associated with RV remodeling.10 It is tempting to speculate that the right ventricle remodels differently in cyanotic and acyanotic patients, with more injury-related reprogramming occurring in cyanotic patients. However, in the absence of gene expression data from age-matched healthy control subjects, we are unable to validate such a hypothesis.
Our data showed increased gene expression levels of the growth-signaling factors FGF7, TGFBR2, EGFR, and insulin-like growth factor binding protein 7 (Table E1 and Figure 2). These gene expression changes could be attributed to a myocardial survival program in an attempt at protecting itself from hypoxia-related cell damage. It is documented that growth factors mediate the survival response in many cell types,14 and the activation of growth factor signaling is cardioprotective.15 A previous study showed an increased expression level of VEGF in the right ventricle when comparing patients with TOF with healthy control subjects.10 Our findings showed no changes in VEGF expression levels in cyanotic compared with acyanotic patients.
Additionally, our data showed increased apoptosis in cyanotic myocardium and increased expression of apoptosis signaling genes, such as TNFSF10 (TRAIL), B-cell chronic lymphocytic leukemia/lymphoma 10 (BCL10), STAT5B, GULP1 (CED6), and STK17B (DRAK2; (Table E1 and Figure 2). The protein encoded by BCL10 (CARMEN) contains a caspase recruitment domain and has been shown to induce apoptosis and to activate nuclear factor κB.16 The protein encoded by STAT5B, a member of the signal transducer and activator of transcription family of transcription factors, has been shown to be involved in diverse biological processes, including apoptosis.17 GULP1 is an evolutionarily conserved adaptor protein required for efficient engulfment of apoptotic cells by phagocytes.18 DRAK2 is a member of the death-associated protein kinase family capable of inducing apoptosis on overexpression in cell culture.19 The activation of apoptosis signaling illustrates how the chronic hypoxic stress seems to promote an injury program within cyanotic patients’ myocardium. This program might explain the worst reoxygenation injury that cyanotic patients experience during and after corrective heart surgery.4
Transcription, MAPK signaling, and contractile machinery (cytoskeleton and adhesion) were the important functional classes for the downregulated genes (Figures 1 and 3). Three of the identified downregulated transcription factors, JUND, JUN, and ATF2, are players in both transcription and MAPK signaling direct association networks (Figure 3). Other downregulated transcription factors included STA3, GATA4, EPAS1, ARNT, NCOA2, and SP1. The reduction of transcription represents part of the injury-related program initiated by chronic hypoxia. Indeed, fully functional transcription machinery is crucial for myocardial function and survival, and a decrease in transcription factor levels would have a repercussion on cardiac function and survival.
Similarly, the reduction of MAPK signaling is another component of the injury-related program triggered by chronic hypoxia. Indeed, the MAPK pathway is crucial for myocyte survival and function. Decreased expression levels of key players of this pathway (eg, activated SAPK) could have a serious effect on the myocardium. Examples of the MAPK pathway players that showed decreased expression levels are MAPK1 (ERK), MAPK14 (SAPK2), MAP3K2, MAP3K3, MAP4K4, AKT2, TAOK1, SOS1, and SOS2 (Table E2 and Figure 3, B). MAPKs phosphorylate a number of known transcription factors to alter their transactivating activities, thus presumably influencing gene expression to elicit the cellular response.20 In cardiac myocytes members of the ERK MAPK family have been implicated in survival signaling in response to ischemia and reperfusion, oxidative stress, and hypoxia.21 Other studies have also shown that the MAPK/ERK kinase and phosphoinositide 3-kinase–protein kinase Akt/protein kinase B pathways might protect against apoptosis.14
In addition, our study showed a downregulation in the contractile machinery (cytoskeleton and cell adhesion clusters). The downregulated transcripts belonging to this grouping included troponin T type 2, myosin XVIIIB, ARP2 actin-related protein 2, and nebulin-related anchoring protein. The reduction of contractile machinery factors seems to represent another part of the injury-related program initiated by cyanosis. Such alteration of contractility factor levels could explain the susceptibility of cyanotic patients to reoxygenation injury (Figure E1).
COX11
Cytochrome-c oxidase 11 (COX11) showed important upregulation in cyanotic patients at the mRNA and protein levels. The COX11 protein is a constituent of the inner mitochondrial membrane and might be involved in biosynthesis of heme A. The upregulation of COX11 in our study suggests that the cyanosis state might trigger a compensation mechanism through COX11 and copper transport to palliate the lack of oxygen.
NCX1
The sodium-calcium (Na+-Ca2+) exchanger (NCX1, SLC8A1) showed an important downregulation in cyanotic patients at both the mRNA and protein levels. NCX1 is a membrane protein that is considered to play an important role in the Ca2+handling of cardiac myocytes. The observed NCX1 downregulation could decrease myocyte calcium handling capacity, leading to mechanical dysfunction.
TRAIL
TNFSF10 (TRAIL) showed also important upregulation in cyanotic patients. TRAIL is a cytokine that belongs to the tumor necrosis factor ligand family. The binding of this protein to its receptors has been shown to trigger the activation of MAPK8/c-Jun NH2-terminal kinase, caspase-8, and caspase-3, thus inducing apoptosis.22 The observed TRAIL increase in the circulation of cyanotic patients could emanate from the myocardium. TRAIL increases in the circulation of cyanotic patients could induce apoptosis in organs and tissues expressing TRAIL receptors. The activity of this protein might be modulated by binding to decoy receptors that cannot induce apoptosis. These decoy receptors, such as osteoprotegerin, present great potential for use as a therapy to reduce any TRAIL induced-apoptosis.
SPP1
SPP1, also known as osteopontin (OPN), was upregulated by 2-fold in cyanotic patients at the mRNA and protein levels. OPN is expressed in the heart at low levels under normal conditions.23 Although the exact role of OPN in cardiomyocyte function is unknown, there is evidence that cardiomyocytes are a prominent source of OPN in vivo and that induction of OPN expression is strongly associated with ventricular remodeling.24 Additionally, increase of OPN expression in the myocardium coincides with the development of heart failure.25 Our data showed an increase in OPN expression at the mRNA and protein levels in cyanotic children, suggesting a remodeling program initiated by chronic hypoxia in the myocardium.
LYZ
LYZ (lysozyme) showed the highest upregulation (5:1) in cyanotic patients. LYZ, the natural substrate of which is bacterial cell wall peptidoglycan, is one of the antimicrobial agents found in human milk and is also present in the spleen, lung, kidney, white blood cells, plasma, saliva, and tears. Here we show protein expression of the LYZ gene product in the ventricular myocardium and increased levels in cyanotic compared with acyanotic patients. The biological significance of this increase is not known, and further studies would be required to uncover it.
Limitations
Even though there was no difference between the 2 groups in terms of right ventricular wall thickness, one cannot completely exclude the possibility that RVH could be a potential confounder in our findings.
As expected, there was a difference in the 2 groups regarding the preoperative use of β-blockers, and this might be a potential confounding factor because it might influence the course of RVH and remodeling.
We did not study normal or nondiseased right ventricular myocardium; however, our focus was on the gene expression differences between cyanotic and acyanotic patients with TOF.
Finally, our investigation can be interpreted as a descriptive study that generates more hypotheses. However, it is our belief that this will provide a better understanding of the mechanisms associated with chronic hypoxia and help develop interventions aimed at improving the clinical outcome in this high-risk group of patients.
CONCLUSIONS
Overall, the transcriptional profile in the cyanotic group was characterized by increased expression level of genes with literature-validated apoptosis and growth/morphogenesis/remodeling properties. Furthermore, it showed decreased expression levels of genes with cardiac function, cell survival, and cytoprotective properties. The molecular signatures identified suggest a reprogramming response in the cyanotic myocardium activated by the chronic hypoxia imposed by the structural congenital heart disease.
CLINICAL IMPLICATIONS
In this study patients in both groups (cyanotic and acyanotic TOF) exhibited a significant hypertrophy of the right ventricle caused by pressure overload. In cyanotic patients the higher degree of right ventricular obstruction and the smaller size of the pulmonary annulus and main pulmonary artery were responsible for the preoperative chronic hypoxic state. We therefore believe that these 2 groups of patients constitute the best possible model to study the influence of cyanosis on gene expression. Our analysis identified several genes deregulated in the cyanotic heart that might be responsible for the susceptibility of cyanotic children to ischemia and reoxygenation injury during and after surgical intervention. Indeed, our previous data4,5 showed that cyanotic patients have significant perioperative myocardial cell damage compared with acyanotic patients undergoing cardiac surgery. This susceptibility to ischemia and reperfusion damage can be explained by the impairment of factors crucial to cardiac function, induction of apoptotic pathways, and alteration of signal transduction pathways seen in cyanotic patients in this study. This information might have a significant effect in improving surgical strategies in cyanotic patients with TOF undergoing corrective cardiac surgery.
Supplementary Material
Acknowledgments
This work was funded by the British Heart Foundation, the Garfield Weston Trust and the Bristol NIHR BRU in Cardiovascular Medicine. M. T. Ghorbel was supported by an Intermediate Research Fellowship from the British Heart Foundation.
We thank Professors Murphy, Newby, and Suleiman for their valuable comments. We also acknowledge Mr A. Parry and Mrs C. McFadden for his surgical and her secretarial help and Megan Musson for her technical assistance.
Abbreviations and Acronyms
- BAX
Bcl-2-associated X protein
- CPB
cardiopulmonary bypass
- ELISA
enzyme-linked immunosorbent assay
- ERK
extracellular signal-regulated kinase
- FDR
false discovery rate
- MAPK
mitogen-activated protein kinase
- RVH
right ventricular hypertrophy
- TOF
tetralogy of Fallot
- VEGF
vascular endothelial growth factor
Footnotes
Disclosures: None.
Supplemental material is available online.
References
- 1.Ihnken K, Morita K, Buckberg GD, Matheis G, Sherman MP, Allen BS, et al. Studies of hypoxemic/reoxygenation injury: without aortic clamping. II. Evidence for reoxygenation damage. J Thorac Cardiovasc Surg. 1995;110:1171–81. doi: 10.1016/s0022-5223(95)70003-x. [DOI] [PubMed] [Google Scholar]
- 2.Teoh KH, Mickle DA, Weisel RD, Li RK, Tumiati LC, Coles JG, et al. Effect of oxygen tension and cardiovascular operations on the myocardial antioxidant enzyme activities in patients with tetralogy of Fallot and aorta-coronary bypass. J Thorac Cardiovasc Surg. 1992;104:159–64. [PubMed] [Google Scholar]
- 3.Dhaliwal H, Kirshenbaum LA, Randhawa AK, Singal PK. Correlation between antioxidant changes during hypoxia and recovery on reoxygenation. Am J Physiol Heart Circ Physiol. 1991;261:H632–8. doi: 10.1152/ajpheart.1991.261.3.H632. [DOI] [PubMed] [Google Scholar]
- 4.Imura H, Caputo M, Parry A, Pawade A, Angelini GD, Suleiman MS. Age-dependent and hypoxia-related differences in myocardial protection during pediatric open heart surgery. Circulation. 2001;103:1551–6. doi: 10.1161/01.cir.103.11.1551. [DOI] [PubMed] [Google Scholar]
- 5.Modi P, Imura H, Caputo M, Pawade A, Parry A, Angelini GD, et al. Cardiopulmonary bypass-induced myocardial reoxygenation injury in pediatric patients with cyanosis. J Thorac Cardiovasc Surg. 2002;124:1035–6. doi: 10.1067/mtc.2002.122536. [DOI] [PubMed] [Google Scholar]
- 6.del Nido PJ, Mickle DA, Wilson GJ, Benson LN, Coles JG, Trusler GA, et al. Evidence of myocardial free radical injury during elective repair of tetralogy of Fallot. Circulation. 1987;76(suppl):V174–9. [PubMed] [Google Scholar]
- 7.Kaynak B, von Heydebreck A, Mebus S, Seelow D, Hennig S, Vogel J, et al. Genome-wide array analysis of normal and malformed human hearts. Circulation. 2003;107:2467–74. doi: 10.1161/01.CIR.0000066694.21510.E2. [DOI] [PubMed] [Google Scholar]
- 8.Konstantinov IE, Coles JG, Boscarino C, Takahashi M, Goncalves J, Ritter J, et al. Gene expression profiles in children undergoing cardiac surgery for right heart obstructive lesions. J Thorac Cardiovasc Surg. 2004;127:746–54. doi: 10.1016/j.jtcvs.2003.08.056. [DOI] [PubMed] [Google Scholar]
- 9.Arab S, Konstantinov IE, Boscarino C, Cukerman E, Mori A, Li J, et al. Early gene expression profiles during intraoperative myocardial ischemia-reperfusion in cardiac surgery. J Thorac Cardiovasc Surg. 2007;134:74–81. e1–2. doi: 10.1016/j.jtcvs.2007.01.025. [DOI] [PubMed] [Google Scholar]
- 10.Sharma HS, Peters TH, Moorhouse MJ, van der Spek PJ, Bogers AJ. DNA microarray analysis for human congenital heart disease. Cell Biochem Biophys. 2006;44:1–9. doi: 10.1385/CBB:44:1:001. [DOI] [PubMed] [Google Scholar]
- 11.Hindmarch C, Yao S, Beighton G, Paton J, Murphy D. A comprehensive description of the transcriptome of the hypothalamoneurohypophyseal system in euhydrated and dehydrated rats. Proc Natl Acad Sci U S A. 2006;103:1609–14. doi: 10.1073/pnas.0507450103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Dennis G, Jr, Sherman BT, Hosack DA, Yang J, Gao W, Lane HC, et al. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003;4:P3. [PubMed] [Google Scholar]
- 13.Ghorbel MT, Sharman G, Leroux M, Barrett T, Donovan DM, Becker KG, et al. Microarray analysis reveals interleukin-6 as a novel secretory product of the hypothalamo-neurohypophyseal system. J Biol Chem. 2003;278:19280–5. doi: 10.1074/jbc.M209902200. [DOI] [PubMed] [Google Scholar]
- 14.Bishopric NH, Andreka P, Slepak T, Webster KA. Molecular mechanisms of apoptosis in the cardiac myocyte. Curr Opin Pharmacol. 2001;1:141–50. doi: 10.1016/s1471-4892(01)00032-7. [DOI] [PubMed] [Google Scholar]
- 15.Hausenloy DJ, Yellon DM. New directions for protecting the heart against ischaemia-reperfusion injury: targeting the reperfusion injury salvage kinase (RISK)-pathway. Cardiovasc Res. 2004;61:448–60. doi: 10.1016/j.cardiores.2003.09.024. [DOI] [PubMed] [Google Scholar]
- 16.Wang D, You Y, Lin PC, Xue L, Morris SW, Zeng H, et al. Bcl10 plays a critical role in NF-kappaB activation induced by G protein-coupled receptors. Proc Natl Acad Sci U S A. 2007;104:145–50. doi: 10.1073/pnas.0601894104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Joung YH, Lim EJ, Kim MS, Lim SD, Yoon SY, Lim YC, et al. Enhancement of hypoxia-induced apoptosis of human breast cancer cells via STAT5b by momilactone B. Int J Oncol. 2008;33:477–84. [PubMed] [Google Scholar]
- 18.Su HP, Nakada-Tsukui K, Tosello-Trampont AC, Li Y, Bu G, Henson PM, et al. Interaction of CED-6/GULP, an adapter protein involved in engulfment of apoptotic cells with CED-1 and CD91/low density lipoprotein receptor-related protein (LRP) J Biol Chem. 2002;277:11772–9. doi: 10.1074/jbc.M109336200. [DOI] [PubMed] [Google Scholar]
- 19.Kogel D, Prehn JH, Scheidtmann KH. The DAP kinase family of pro-apoptotic proteins: novel players in the apoptotic game. Bioessays. 2001;23:352–8. doi: 10.1002/bies.1050. [DOI] [PubMed] [Google Scholar]
- 20.Yang SH, Sharrocks AD, Whitmarsh AJ. Transcriptional regulation by the MAP kinase signaling cascades. Gene. 2003;320:3–21. doi: 10.1016/s0378-1119(03)00816-3. [DOI] [PubMed] [Google Scholar]
- 21.Baines CP, Molkentin JD. STRESS signaling pathways that modulate cardiac myocyte apoptosis. J Mol Cell Cardiol. 2005;38:47–62. doi: 10.1016/j.yjmcc.2004.11.004. [DOI] [PubMed] [Google Scholar]
- 22.Muhlenbeck F, Haas E, Schwenzer R, Schubert G, Grell M, Smith C, et al. TRAIL/Apo2L activates c-Jun NH2-terminal kinase (JNK) via caspase-dependent and caspase-independent pathways. J Biol Chem. 1998;273:33091–8. doi: 10.1074/jbc.273.49.33091. [DOI] [PubMed] [Google Scholar]
- 23.Subramanian V, Krishnamurthy P, Singh K, Singh M. Lack of osteopontin improves cardiac function in streptozotocin-induced diabetic mice. Am J Physiol Heart Circ Physiol. 2007;292:H673–83. doi: 10.1152/ajpheart.00569.2006. [DOI] [PubMed] [Google Scholar]
- 24.Graf K, Do YS, Ashizawa N, Meehan WP, Giachelli CM, Marboe CC, et al. Myocardial osteopontin expression is associated with left ventricular hypertrophy. Circulation. 1997;96:3063–71. doi: 10.1161/01.cir.96.9.3063. [DOI] [PubMed] [Google Scholar]
- 25.Singh K, Sirokman G, Communal C, Robinson KG, Conrad CH, Brooks WW, et al. Myocardial osteopontin expression coincides with the development of heart failure. Hypertension. 1999;33:663–70. doi: 10.1161/01.hyp.33.2.663. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.