

Abstract
It is becoming increasingly evident that cancer constitutes a group of diseases involving altered stem-cell maturation/differentiation and the disturbance of regenerative processes. The observed malignant transformation is merely a symptom of normal differentiation processes gone astray rather than the primary event. This review focuses on the role of cancer stem cells (CSCs) in three common but also relatively under-investigated cancers: head and neck, ovarian, and testicular cancer. For didactic purpose, the physiology of stem cells is first introduced using hematopoietic and mesenchymal stem cells as examples. This is followed by a discussion of the (possible) role of CSCs in head and neck, ovarian, and testicular cancer. Aside from basic information about the pathophysiology of these cancers, current research results focused on the discovery of molecular markers specific to these cancers are also discussed. The last part of the review is largely dedicated to signaling pathways active within various normal and CSC types (e.g. Nanog, Nestin, Notch1, Notch2, Oct3 and 4, Wnt). Different elements of these pathways are also discussed in the context of therapeutic opportunities for the development of targeted therapies aimed at CSCs. Finally, alternative targeted anticancer therapies arising from recently identified molecules with cancer--(semi-)selective capabilities (e.g. apoptin, Brevinin-2R) are considered.
Keywords: Nanog, Nestin, Notch1, Notch2, Oct4, Wnt
INTRODUCTION: ADULT STEM-CELL BIOLOGY, HEMATOPOIETIC AND MESENCHYMAL STEM CELLS AS EXAMPLES
Stem cells are increasingly becoming the focus of various areas of biomedical research. They are pluripotent, clonogenic, and self-renewing, with a plasticity to differentiate into the cell types of the particular tissue in which they reside and often to trans-differentiate [51, 73]. The trans-differentiation potential of stem cells and their capacity for tissue renewal and damage repair bring them into the focus of interest of biotechnologists and clinicians [110, 116].
While the field expands rapidly, so far stem cells are still largely defined by their capacity to (re-)colonize a given tissue niche, or initiate tumor growth (cancer stem cells), as only few molecular markers of stemness exist. Among them, the transcription factor Oct 3/4 (POU5F1/Oct 4) is considered as one of the best indicators of stemness capacity [39]. Its expression in nonmalignant cells is restricted to the pluripotent cells in the embryo and germ cells. Oct 3/4 is also a reliable marker in germ cell tumor diagnostics. It is expressed in the lesions that may initiate gonadoblastoma and carcinoma in situ, as well as in invasive embryonal carcinoma and seminomas (see below).
The same qualities that make stem cells valuable in regenerative medicine, tissue engineering, and biotechnology also carry dangers to the host organism. Mutations affecting their differentiation potential combined with the failure to control their unlimited proliferative capacity may transform them into “cancer stem cells” (CSCs). While this review is dedicated to head and neck, ovarian, and testicular CSCs, we provide below, as an introduction, some basic information about selected normal adult stem-cell populations.
Hematopoietic stem cells
Hematopoietic stem cells (HSCs) are the best-studied adult stem cells thus far and they were the first stem cells used in the clinic, thereby serving as an excellent vehicle for an introduction to stem-cell biology. HSCs are mainly located in the bone marrow (BM) and they are a classical example of multipotent adult stem cells, able to generate all cells of the blood and the immune system [2, 144, 153]. A small percentage of HSCs circulates in and can be isolated from the blood [178]. Lineage-specific differentiation of HSCs is regulated by various cytokines and growth factors which activate several signaling pathways (bone morphogenetic protein (BMP)-, Wnt-, and Notch-pathways) [27]. The sialomucin cell-surface marker CD34 is an important marker for HSCs and some other populations of adult stem cells. CD34 also appears to have a significant role in early hematopoiesis [164]. HSCs are hematopoietic lineage marker negative and thus do not express markers present on progenitor cells committed to a specific differentiation pathway. In mice, HSCs are c-Kit+, Thy-1lo, and stem-cell antigen (Sca)-1+ [77, 171], and a single transplanted cell from this cell population can give rise to life-long hematopoiesis [130, 160]. Similarly, human HSCs are CD34+, Thy-1+ and c-Kit+ [18].
An important lineage marker to exclude for HSCs when harvesting is CD45, a type I transmembrane molecule found on the cell surface of all nucleated hematopoietic cells and progenitors [168]. CD45 has important functions during immunological processes and for differentiation into various hematopoietic cell lineages [136]. For therapeutic intervention, HSCs from BM can be mobilized into the blood stream by administration of granulocyte colony-stimulating factor or granulocyte-macrophage colony-stimulating factor [119]. Human HSCs (CD34+/CD133+/VEGFR3+) can also be isolated from umbilical cord blood. They express the 120-kDa 5-transmembrane-domain glycoprotein and stem-cell surface marker CD133 [111, 170, 181] and can differentiate into mature endothelial cells [134].
The c-Kit receptor belongs to subclass III of the tyrosine kinase receptor family with five immunoglobulin-like extracellular domains [142]. Stem cell factor (SCF) is the ligand for the c-Kit receptor [177] and mediates c-Kit receptor dimerization and activation [142]. C-Kit has an essential function for constitutive hematopoiesis and self-renewal of progenitor cells [126]. Within the human hematopoietic system, 70% of CD34+ cells in BM express c-Kit protein, and this includes lineage-restricted hematopoietic progenitor cells [11, 132] and precursor cells capable of establishing long-term in vitro hematopoiesis [154]. The 18-kDa protein Sca-1 (Ly-6A/E) is GPI-anchored to the cell membrane and serves as an early marker for multipotent murine HSCs [161, 172]. Sca-1+ HSCs can be isolated from adult BM, peripheral blood, and spleen [85, 115, 117, 161, 162, 179]. Sca-1 is also present in several non-hematopoietic tissues [173] and has been described in multipotent stem cells in the connective tissue of skeletal muscle [44]. Endothelial precursor cells and HSCs share the markers CD34, Tie-2, and fetal liver kinase (Flk)-1, which suggests a common ancestral stem cell population in the BM [10].
The Flk-1 is a 200- to 230-kDa protein involved in vasculogenesis and, together with CD133, constitutes a marker for endothelial progenitor cells [148]. Murine Flk-1 is also known as vascular endothelial growth factor receptor 2 (VEGFR2), and the kinase insert domain--containing receptor (KDR) is the human homolog. CD34+ BM-derived cells have been demonstrated to differentiate into CD31+ Tie-2 receptor+ endothelial cells, incorporating acetylated LDL and producing nitric oxide in response to VEGF [10, 151]. Human CD34+ KDR+ BM-derived stem cells comprise HSCs [185], endothelial precursors [134], and hemangioblasts [135]. Osteoblasts located in the BM are niche cells mediating quiescence of the HSCs by secreting angiopoietin-1, which binds to the Tie-2 surface receptor on HSCs [9]. The perivascular site serves as another niche for HSCs and reveals local expression of the chemokine CXCL12 [157]. The expression of both the CXCL12 chemokine receptor CXCR4 and CD44 on HSCs may allow the homing of these cells to different niches [2].
Mesenchymal stem cells
Apart from HSCs, BM also harbors mesenchymal stem cells (MSCs). These cells are negative for the HSC markers CD14 and CD34 and for the HSC exclusion marker CD45. MSCs are present in low numbers in the circulation and in tissues [52]. Only 0.01–0.001% of the mononuclear cells isolated from BM have MSC properties, but their multipotency, proliferative capacity in culture, and ability to recruit to sites of organ injury make them attractive therapeutic targets. STRO1, CD105 (endoglin, gp160), CD73, CD166, CD146 (MCAM, MUC18, A32 antigen, S-endo-1), and SSEA-4 are markers for MSCs and, more recently, this list has been extended to include CD145, CD49a, CD106 (VCAM), and CD90 (Thy-1) [14, 23, 62, 139].
STRO1 is a cell surface antigen on stromal BM cells [155] and serves as a valuable marker for the identification, isolation, and functional characterization of clonogenic human BM stromal cell precursors. STRO1+ MSC can differentiate into multiple mesenchymal lineages, including hematopoiesis-supportive stromal cells, pericytes, adipocytes, myofibroblasts, myocytes, cardiomyocytes, osteoblasts, chondrocytes, and neurons [48, 62, 131, 139, 186]. The MSC surface markers CD146 and CD105 are also expressed on endothelial precursor cells, which possess characteristics similar to those of pericytes [17, 169]. In addition to the expression of the Ca2+-independent cell adhesion molecule CD146, BM and dental pulp MSC express α-smooth muscle actin and their location in the perivascular space may suggest that these MSCs are related to pericytes [152]. CD146 is also expressed in human endothelial cells isolated from cord blood and was shown to be involved in endothelial signaling [8]. MSCs may engraft into various organs and, when injected into the bloodstream, settle at sites of injury [150]. MSCs have been used in various therapies, including the treatment of hemophilia, osteogenesis imperfecta, and cartilage lesions and to accelerate hematopoiesis recovery following chemo/radiotherapy, to prevent scarring after myocardial infarction, and to treat neural lesions following stroke or trauma [75, 150, 176]. Because of the promising therapeutic applications of these multipotent adult stem cells, a number of clinical trials involving MSCs are currently under way (see: http://ncrm.peerlis.com/ncrm/content.aspx?id=17&displaypageid=29, http://ora.ra.cwru.edu/stemcellcenter/research/Clinical%20Trials.htm).
ORAL SQUAMOUS CELL CARCINOMA
In maxillofacial surgery, patients with oral squamous cell carcinoma (OSCC) or basal cell cancer (BCC) of the head and neck represent the majority of oncological clinical cases worldwide. Although CSCs are likely to play a role in the cause of these cancers, there have been no research reports proving the successful isolation of specific CSCs from BCC or OSCC lesions to date. Research reports have used immunohistological studies, investigations on the gene transcript level, and micro-morphological data to demonstrate or refute the CSC hypothesis in these carcinomas. Thus we focus below on recent advances and the ongoing discussion regarding the role of CSCs in the development of head and neck cancers.
OSCC is the sixth most common malignant tumor and its incidence is increasing worldwide. Several models have been proposed to elucidate risk factors, such as smoking and alcoholic beverages. Genetic and epidemiological markers are involved in the generation and progress of OSCC [26, 94]. According to the multi-step carcinogenesis concept, OSCC develops in a series of more or less defined intermediate stages through the accumulation of molecular changes and progression from a pre-cancerous leukoplakia to pre-invasive (erythroplakia) and finally to the invasive cancer disease with lymphonodal outspread and severe local destruction [122, 137]. The overall five-year survival rate in patients with OSCC is still lower than 50% [13, 45]. According to histopathological grading, OSCC displays variable metastatic potentials, and highly differentiated OSCC (G1) displays slow growth and destruction to adjacent muscle and the jawbone. By contrast, high invasiveness, early spread to regional and cervical lymph nodes, and poor prognosis are hallmarks of poorly differentiated OSCC. OSCC frequently shows local recurrence after the initial surgical or radiological treatment at the primary site and, unfortunately, even after complete resection (R0-treatment) [13, 45]. Despite all attempts, no significant progress has been achieved by combining surgery and radiotherapy, as “traditional” ways of treatment, with chemotherapy (neo-adjuvant strategies). Furthermore, there are no standardized protocols or common therapeutic strategies currently available, and the identification of relevant cellular and molecular markers primarily involved in oral pre-cancerous lesions and OSCC is imperative.
The treatment strategies of OSCC are based on the idea that oral cancer is generated by a population of cells of equal proliferative and aggressive potential [37]. More recent reports suggest that the CSC hypothesis may likely also apply to OSCC [37, 68]. There are various possible scenarios by which OSCC could originate from stem cells: (a) it develops directly from malignant stem cells or (b) a tumorigenic potential is promoted in the supra-basal compartment when differentiating cells deviate from their predetermined differentiation route and transform into malignant intra-epithelial cells. The latter possibility is supported by the existence of the Tis-stage (carcinoma in situ) with the presence of typical malignant intraepithelial cells and an intact basal membrane. In Tis-lesions, malignant potential seems to develop in the stratum spinosum, but not in the stratum basale of the squamous epithelium. The dedifferentiation cascade in OSCC is typically associated with p53--mutations and loss of DNA-repair mechanisms [25, 175]. In addition, oncogene activation, tumor suppressor gene inactivation (i.e. CD82), and the downregulation of cell-cell contact proteins, such as E-cadherin and β-catenin, were described in OSCC [32, 54, 78, 92, 96, 113, 182]. OSCC is derived from malignant keratinocytes and their corresponding malignant stem cells may be found in the stratum basale of the oral and dermal epithelium, which has been identified in immunohistochemical studies to harbor epithelial stem cells. The highest numbers of stem cells are located adjacent to the dermal papillas and the basal membrane.
Epithelial stem cells express β1-integrin and cytokeratin 15 [102]. Several factors are involved in epidermal cell-cycle control and the regulation of normal differentiation pathways in keratinocyte formation [102]. Notch genes encode large single transmembrane proteins and are crucial for normal epidermal integrity and function. In the skin and oral mucosa, Notch1 and Notch2 signaling pathways induce and control the differentiation of keratinocytes, but their tissue localization suggests specific functions for Notch2, exclusively present in the basal epithelial layer, and Notch1, which is present in all epidermal layers. In skin, Notch1 and Notch2 may act as tumor suppressors and regulate stem-cell maintenance, proliferation, and apoptosis [53, 127, 128].
BCC is the most common form of skin cancer and develops from basal epithelial cells of the skin which become malignant upon prolonged sunlight and/or UV--light exposure. BCC is the result of a long-term process and this carcinoma usually grows much more slowly than squamous cell cancer or malignant melanoma. BCC is believed to be an epidermal stem cell-derived carcinoma of the skin. For BCC, the increase in Sonic hedgehog (SHH) signal transfer and activation of the Wnt pathway were found to be the molecular cause for malignant transformation [128]. This imbalance is strongly connected to Notch deficiency and Notch1 and Notch2 expression are reduced in BCC [128]. Animal experiments showed that deletion of the Notch gene can lead to BCC in mice [123, 140, 167] and UV-light exposure triggered malignant basal cell transformation, especially in Notch1-defient skin areas [128]. BCC has an extremely low metastatic potential and is therefore considered a “semi-malignant” tumor. However, once ulcerated, BCC shows aggressive growth and deep local tissue invasion.
Searching for stem-cell populations
Established OSCC cell lines may contain a small stem-cell fraction with high regenerative potential. When cultured at low densities, OSCC cell lines can generate cell aggregates with morphological and clonal features distinct from those of the rest of the tumor cells. These CD44+ subpopulations vary in their morphological features, display different growth potential for each clonal subtype, and may be considered stem cell-like populations [68]. When cultured at low density in DMEM-F12 plus 10% FCS, fibroblast-shaped BHY--cell lines derived from a highly differentiated OSCC will group into small clusters of cells with an epithelial phenotype and limited mitogenic activity (Mustafa, unpublished data) (Fig. 1). Currently, the source of the CSC population in BHY remains to be determined, this largely due to the lack of stem-cell markers for head and neck cancers. One promising candidate molecule is the EGF-7TM protein CD97. CD97 is highly expressed in BM cells, undifferentiated thyroid carcinoma, and dedifferentiated (G3) OSCC [12, 47, 64, 97, 98, 120]. This surface molecule co-localizes within the basal cell layer of the oral squamous epithelium and its derivatives (Fig. 2). It is currently not known whether CD97+ β1--integrin-positive cells express other known stem-like markers or possess higher proliferative potential, as would be expected from stem/progenitor cells. The identification of specific stem cell-like markers for OSCC is most definitely challenging and requires the combined approach of high-end cell-separation techniques, such as laser dissection of cells, single-cell Q-RT-PCR, and DNA-fingerprinting [79, 149].
Fig. 1.

Possible cancer stem cell populations within Head and Neck cancers. A – cell line BHY, initial holoclone formation with typical polygonal cells and dense intercellular contacts (40×). The cells are immunostained for CD97 and show strong cytoplasmatic DAB-brown staining (monoclonal antibody MEM180, 1:100, ABC-method). B – BHY cells with spider-like shapes cultured at low density fail to form holoclones. CD97 is only detectable in the perinuclear cytoplasma. Short tandem repeat-analysis (DNA-fingerprint) could prove the identical genetic profile as compared with holoclone cells (20×).
Fig. 2.
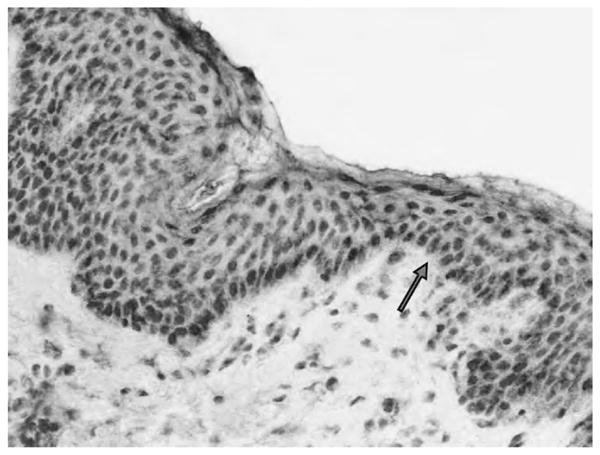
The presence of tissue committed stem cells within human skin. CD97-immunohistochemistry of a non-pathological human skin sample (MEM180 monoclonal antibody, ABC-method, DAB--labeling). The basal epithelial cells express CD97. CD97 is not detectable in the stratum spinosum and the stratum corneum (20×).
OVARIAN CANCER: PLURIPOTENCY AND PUTATIVE CSCs
Somatic stem cells contribute to normal tissue repair and regeneration, and it has been suggested that multi-potent somatic stem cells in the ovary have the potential to regulate surface epithelium repair after ovulatory rupture. Furthermore, preliminary observations indicate the existence of proliferative germ cells sustaining oocyte- and folliculogenesis in the post-natal ovary in the mouse [82], and the surface epithelium of adult women is a source of oocyte and granulosa cells in vitro [87], further indicating that the ovarian tunica albuginea contains stem cells. Ovarian somatic stem cells within this epithelial layer are expected to divide asymmetrically, yielding both a daughter cell that proceeds to terminal differentiation and an undifferentiated self-copy. Repeated asymmetric self-renewal sets the stage for somatic stem cells and their immediate progenitors to accrue mutations over time, which ultimately can lead to their transformation into CSCs and malignant progression. Inflammatory mediators and reactive oxidants generated during the ovulatory process and healing of the ovulatory wound [121] may be important contributors to the carcinogenic dedifferentiation of ovarian surface epithelial cells inducing DNA strand breaks and oxidative base (8-oxoguanine) damage [118]. A genetically altered stem or progenitor cell with damaged DNA, but not committed to death, could therefore give rise to a transformed phenotype that is propagated upon healing of the ovulatory wound.
Epithelial ovarian cancer is an extremely aggressive disease associated with a lack of early symptoms, rapid progression to peritoneal metastases, and poor prognosis. An initial improvement is observed when surgical removal of ovarian cancer tissues is followed by chemotherapy. Drug-resistant tumors, however, will recur in the vast majority of patients, leading to a five--year survival rate of less than 30% [29, 80].
As was recently demonstrated for breast and prostate cancer as well as other malignancies, including ovarian cancer, CSCs may not only survive chemotherapy, but chemotherapy may actively select for chemoresistant, highly aggressive CSCs [183]. Contributing factors are the preferential expression of chemoresistance molecules from the ATP-binding cassette (ABC) transporter family, such as the multidrug-resistance gene 1 and the breast cancer-resistance protein 1, which contribute to chemoresistance by cellular clearance of lipophilic drugs [61]. High expression of ABC-G2 has been observed in the side population (SP), which constitutes a cell population of undifferentiated cells [42, 95] that efficiently exclude Hoechst 33342, representing an enriched source of stem cells [31]. Cancer cells belonging to the SP are immature, poorly differentiated, and highly tumorigenic. Gene expression profiles indicate that cells in the SP-gated fraction express high levels of stem-cell markers and low levels of differentiation markers [66]. Recently, an SP has been identified in two distinct mouse ovarian cancer cell lines as well as in human ovarian cell lines and primary ascites tumor cells [165]. These SP cells show growth characteristics similar to CSCs [71] and are capable of self-renewal and asymmetric division in culture. In vivo, ovarian SP cells have the potential to initiate tumor growth more quickly and at lower cell numbers than non-SP ovarian cancer cells [165]. Moreover, cell clones with sustained clonogenic potential in vitro and tumorgenicity in vivo, even after serial transplantation in mice, have been identified in ovarian cancer [16]. The possible multipotency of such clones was further supported by the expression of three “stemness” markers: Oct4, Nanog, and Nestin [16]. More recently, Ferrandina et al. [50] identified the expression of CD133 antigen, considered to be a marker of undifferentiated cells, in a cellular subpopulation in a large variety of ovarian tissues. CD133+ ovarian tumor cells exhibited higher clonogenic and more extensive proliferative potential than CD133− cells, which is similar to previous reports for other human malignancies [36, 156].
Due to the preferential expression of chemoresistance molecules, it is believed that CSCs may survive after chemotherapy and therefore induce relapse in several patients. It therefore seems intuitive that only the eradication of CSCs can lead to an effective cancer therapy. Recent and preliminary studies have suggested Müllerian inhibiting substance (MIS) as a possible adjuvant to classical ovarian cancer chemotherapeutic regimes in targeting putative CSCs. In the embryonic urogenital ridge, an intact MIS signaling pathway is required for MIS responsiveness [184]. Similarly, a variety of ovarian CSCs, both SP and non-SP, possess functional receptors for MIS. Treatment with MIS is able to inhibit cell proliferation in both SP and non-SP cells, whereas classic chemotherapies inhibit mainly non-SP cells. Thus MIS is able to actively suppress growth of the chemoresistant SP subpopulation [165].
Ovarian cancers come in different forms. Besides surface epithelium-derived ovarian cancer, other categories of ovarian tumors are represented by sex-cord and germ-cell tumors (GCTs). GCTs comprise a heterogeneous group of neoplasms predominantly found along the midline of the body, on the migration route of the primordial germ cells (PGCs) during embryogenesis from the yolk sac to the genital ridge [7]. Different histopathological entities can be distinguished: (a) teratomas and yolk sac tumors, (b) seminomas/dysgerminomas/germinomas of the testis and ovary, mediastinum, and midline of the brain, (c) dermoid cysts of the ovary, and (d) spermatocytic seminomas of elderly testis. Of these, tumors of types a, c, and d are composed of differentiated tissues, whereas type b GCTs demonstrate pluripotent potential. A close resemblance between testicular GCTs and embryonic stem (ES) cells has been shown [4]. Specific expression of Oct 3/4 has been related to the pluripotent capacity of testicular GCTs, suggesting their origin from transformed pluripotent progenitor cells, such as PGCs [100]. Oct 3/4 is critical for the self-renewal of ES cells [124] and, under physiological conditions, Oct 3/4 transcription factor is expressed exclusively in pluripotent ES cells, PGCs, oogonia, and gonocytes [60, 65, 147] and its expression is downregulated during differentiation [138]. Many similarities exist between testicular and ovarian GCTs, including morphological resemblance and a similar pattern of chromosomal alterations [143]. The pathobiology of testicular seminomas is discussed in detail in the next paragraph. Two studies recently demonstrated that ovarian dysgerminomas are characterized by strong positivity for Oct 3/4 [34, 100], which is similar to testicular seminomas. Hoei-Hansen et al. [72] investigated the expression of additional stem cell-related markers of pluripotency in human dysgerminomas. In addition to Oct 3/4, the authors identified the expression of c-Kit, also known as a tyrosine kinase receptor for SCF, Nanog, and AP-2γ. The c-Kit/SCF system has particular importance in the origin of ovarian GCTs (OGCTs) due to its role in PGC proliferation and survival in the developing human gonad [114]. In particular, it has been suggested that the high expression of c-Kit in dysgerminomas is due to spontaneous gene mutations occurring before oocytes enter meiosis, leading to increased survival and proliferation of undifferentiated germ cells which are committed to pluripotency [72].
Malignant OGCTs have a median onset age of 18 years and represent approximately 3% of all ovarian cancers [125]. The fact that OGCTs normally affect women in their reproductive years implies the importance of optimal therapy to maximize the percentage of patients in whom ovarian function can be conserved. The immunohistochemical detection of stem cell-related markers offers a promising tool in the accurate diagnosis and evaluation of OGCTs, facilitating the diagnosis of pre-malignant germ-cell lesions and offering a choice of prompt and targeted therapy.
HUMAN TESTICULAR GCTs
Testicular GCTs (TGCTs) are the most common malignant tumors in the male Caucasian population between the second to fourth decade of life [1, 109]. As first described by Skakkebaek [158, 159], intratubular germ-cell neoplasia unclassified (IGCNU, also known as carcinoma in situ) represents the common precursor lesion of all TGCTs except spermatocytic seminomas. According to the current hypothesis, delayed or compromised maturation of fetal germ cells, i.e. PGCs/gonocytes, leads to the persistence of immature germ cells which subsequently may undergo malignant transformation [129]. The precise nature of transformation of these premature germ cells to IGCNU is not known. Recent studies, however, identified specific differentiation stages of fetal germ cells, which indicates that the process of normal maturation is likely to be interrupted at a distinct period of fetal development prior to neoplastic transformation [133]. Once established, IGCNU will always progress to TGCTs and exhibit a wide spectrum of histological subtypes, including seminoma, embryonal carcinoma, teratoma, choriocarcinoma, and tumors of the yolk sac. Among these, embryonal carcinoma is a malignant pluripotent counterpart of ES cells and is able to differentiate into all three germ lineages.
Previous studies showed a substantial overlap in the expression patterns of embryonal carcinomas and ES cells [5, 163]. Despite differential histology, seminomas and IGCNU share a high overlap of gene expression with ES cells [4, 20, 21]. Pluripotency genes such as Oct 3/4 and Nanog are highly expressed in IGCNU, seminomas, and embryonal carcinomas, but not in other histological types of TGCTs. The entirely pluripotent characteristic of embryonal carcinomas, however, might be established by SOX2, which is highly expressed in embryonal carcinomas but missing in IGCNU and seminomas (Fig. 3). The transcription factor SOX2 is essential for maintaining the pluripotent phenotype in ES cells and is a partner of Oct 3/4 in regulating several ES cell-specific genes [145]. In vitro experiments with ES cell nuclear extracts demonstrated that Oct 3/4 and SOX2 interact specifically and bind to a composite regulatory element. Activation of this element maintains Oct 3/4 and SOX2 expression in pluripotent cells [35]. Thus the SOX2-Oct 3/4 interaction may directly assist in reprogramming or serve as a driving factor leading to the reprogramming of common undifferentiated neoplastic germ cells to become embryonal carcinoma. Several other candidate genes which may contribute to the maintenance of pluripotency have recently been detected by genome-wide arrays and validated by immunohistochemistry, e.g. CENPA and PODXL [20]. Their role in TGCTs should be elucidated by functional studies in the future. In contrast to embryonal carcinomas, seminomas express many genes associated with normal spermatogenesis [21].
Fig. 3.

Stem cell markers expressed in both embryonal carcinomas and seminomas. Pluripotency regulators POU5F1/Oct 3/4 and NANOG are expressed both in embryonal carcinomas and seminomas as shown here by immunohistochemical staining on a seminoma sample (A –POU5F1, B – NANOG). In contrast, pluripotency regulator SOX2 is differentially expressed in embryonal carcinoma only (C – embryonal carcinoma component of a TGCT with a mixed histology expressing SOX2, D – compare a completely negative staining in a seminoma). Bar=100 μm.
ES cells and TGCTs not only exhibit similar gene expressions, but also display similar epigenetic DNA--modifications. Compared with normal somatic cells, ES cells show a very low mutation rate (10−6 in ES cells and 10−4 in somatic cells) [30]. Similarly, TGCTs exhibit a very low mutation rate compared with somatic malignancies [22]. The receptor tyrosine kinase c-Kit represents the only exception from this general finding. Several studies have shown that different gain-of-function c-Kit mutations are often present in bilateral, but not in unilateral GCTs [19, 99]. These differences are likely due to a very early formation of the mutations, probably during embryonal development prior to testis formation. Most TGCTs are highly sensitive to DNA damaging agents, most likely because of the embryonal characteristics of these tumors. ES cells are equally sensitive to irradiation and chemotherapy and the molecular basis for this sensitivity is the cytoplasmic expression of the checkpoint kinase CHK2, which leads to an accelerated transition from G1 to S phase. It is through this mechanism that only intact cells survive, while the damaged cells become apoptotic [74]. Furthermore, all invasive TGCTs show a consistent gain of the short arm of chromosome 12, as found in ES cells upon extensive in vitro culturing [43]. In contrast to many other malignancies, TGCTs have a wild-type p53. A specific cluster of miRNAs which are also expressed in ES cells prevent p53-dependent cellular senescence upon oncogenic stress [174].
While there are no animal models for seminoma to date, two groups have been successful in generating cell lines from seminoma patients [88, 112] which are now being used to study seminomas. These cell lines are JKT-1 [70, 81, 90, 146] and TCam-2 [59, 89]. Gene expression and immunohistochemical profiling have shown that only the TCam-2 cell line is representative for seminoma (Fig. 4) [40, 41, 46]. Both cell lines are essential for future studies on the molecular mechanisms of reprogramming and pluripotency in TGCTs and seminomas.
Fig. 4.

Whole genome expression analysis and comparison between normal testicular tissues, seminoma, embryonal carcinoma, and the relevant cell line models. Whole genome expression analysis of normal testicular tissues (N1-N4), seminoma (S1-S4), embryonal carcinoma (E1-E3) and the cell lines JKT1 and TCam-2 shows that JKT1 clusters with seminoma. The seminoma-like nature was confirmed using reverse transcription/polymerase chain reaction (see main text for details).
TARGETING SIGNALING PATHWAYS ACTIVE IN CSCS DURING CANCER THERAPY
Neoplastic germ cells retain their embryonic features while the normal maturation process of spermatogenesis causes the downregulation of embryonic gene expression and pluripotency in gametes. Therefore, TGCTs represent embryonic cancers found in adults. Both the seminomas and non-seminomas have their specific populations of stem-cell representatives derived from PGCs/gonocytes and from ES cells, respectively. In the future, the potential and functional process of reprogramming the precursor neoplasia IGCNU to different histological types of GCTs will provide potentially new and exciting avenues for the treatment of TGCTs. Recent progress in novel global screening technologies such as genomics, proteomics, and, in the future, metabolomics as well as combinatorial approaches will provide targets and tools for such reprogramming strategies [3, 6, 91].
CSCs have recently become an attractive target for novel anticancer therapies and several biochemical and physiological factors have emerged as potential targets for drug development. Although a multitude of potential therapeutic approaches have been identified, many can be broadly classified as either differentiation or elimination therapy in which CSCs are coaxed to differentiate or are themselves eliminated, respectively [15, 108, 187]. Molecules that have been implicated in playing a role in CSC signaling spanning several tissue types, and may subsequently serve as potential targets for the development of novel therapies, include CD133 [166] and components of the hedgehog/patched (HH/PTCH) [101], Wnt [84], and Notch signaling cascades [84, 108] as well as signal transducer and activator of transcription 3 (STAT3) [58] and telomerase [67]. Regulating various components of the CSC microenvironment and niche, such as the intracellular reactive oxygen species (ROS) levels [166] and tumor vasculature [166, 180], have also been suggested as a potential means of therapy. For a number of cancers, immunotherapeutic approaches, either by targeting antigens over-expressed by cancer cells or, in rare cases, by the induction of the “graft-versus-disease” response, have been successful [24, 83, 93, 141].
Investigators recently demonstrated that CD133+ cells in human brain cancers possess both self-renewal and differentiation properties as well as the ability to initiate tumor growth in vivo, whereas CD133− cells do not. These human CD133+ brain CSCs, isolated from fresh tumor specimens, are capable of escaping the fatal damage induced by ionizing radiation via preferential activation of DNA repair checkpoints through phosphoryla-tion of the cell cycle checkpoint proteins CHK1 and CHK2. Conversely, it was also shown that two pharmacological inhibitors of CHK1 and CHK2 can reduce the radioresistance of CD133+ cells by disrupting the otherwise efficient DNA repair mechanisms. Another study revealed that BMPs, which normally function as soluble factors that induce the differentiation of neural precursor cells into mature astrocytes, can also induce CD133+ brain CSCs to differentiate, which in turn significantly impedes their original tumor-initiating capacity [166]. This implies that forced differentiation of CD133+ brain CSCs via therapeutically induced over-expression of BMPs may serve as an effective means of sensitizing the tumor population to conventional chemotherapy and reducing the rate of recurrence. Alternatively, it is also implied that the cancer-selective inhibition of CD133 in brain CSCs can also sensitize them to radiotherapy and reduce their self-renewal and differentiation capabilities.
Several pathways play a role in the growth and maintenance of normal stem cells in several tissue types, among which is the HH/PTCH pathway. Mammalian HH genes, which include the homologues IHH, DHH, and SHH, are highly expressed in small-cell lung, prostate, breast, gastric, and pancreatic cancer cell lines. In the HH/PTCH pathway, mutational inactivation of PTCH, which serves as the receptor for HH, leads to the constitutive activation of smoothened (SMO), which is a G-protein-coupled receptor family protein that is regulated by PTCH. Deregulation of the HH/PTCH pathway has been proposed to be a precursor for stem cell activation in cancers. Cyclopamine, which is a steroid--like compound that binds to and inactivates SMO, inhibits the growth of cells with activated HH signaling. This was shown in mice treated with cyclopamine in which the PC-3 and DU-145 human prostate cancer cell lines were grown as xenografts and eradicated after 21 days. It was recently shown that vitamin D3 functions as an essential signaling molecule between PTCH and SMO in that it is normally secreted by PTCH and inhibits SMO on adjacent cells and the one from which it was secreted. Cyclopamine may function in a similar manner as it competes with vitamin D3 for binding on SMO. This suggests that vitamin D3 and its steroidal derivatives can be used as anticancer compounds in CSCs with active HH signaling. Cyclopamine treatment also significantly down-regulated the expression of GLI2, GLI3, PTCH, SUFU, and SHH, all of which are components of the HH/PTCH pathway, suggesting that the relative mRNA or protein expression levels of these genes can be utilized as a scale in evaluating the effectiveness of targeted inhibitors of human CSCs [101].
Other signaling cascades involved in the self-renewal and differentiation of normal stem cells are the Notch and canonical Wnt pathways, the former playing an especially important role in neuronal stem cells. Synergy between the canonical Wnt and Notch pathways is involved in inhibiting terminal differentiation of intestinal epithelial cells and both are necessary for the self-renewal of HSCs. In the canonical Wnt pathway, extra-cellular Wnt proteins bind to the receptor of the Frizzled (FZD) family. This event, coupled with the activity of the LRP5 and LRP6 coreceptors, activates a pathway leading to the inhibition of proteolysis and subsequent nuclear accumulation of β-catenin, which in turn activates the transcription of FGF20, DKK1, WISP1, Myc, and CCND1 through an interaction with T-cell factor/lymphoid enhancer factor proteins. It is thought that deregulation of this pathway induces the transformation of normal stem cells into CSCs.
PKF118-310 and ZTM000990 are small-molecule leads targeted to the canonical Wnt signaling pathway, whereas anti-Wnt1 and anti-Wnt2 are monoclonal antibodies that have demonstrated anticancer effects in vitro [84]. Notch ligand binding to Notch family receptors results in the release of the Notch intracellular domain into the cytoplasm via proteolysis. The intracellular domain in turn binds to HLH and induces the transcriptional activation of several downstream target genes [108]. Aberrant activity of the Notch pathway is an early event in a pre-invasive breast cancer, ductal carcinoma in situ. Over-expression of the Notch1 intracellular domain in ductal carcinoma in situ was also predicted to result in a reduced time of recurrence, five years after surgery [49]. Over-expression of the active form of Notch1 in the human lung adenocarcinoma cell line A549 resulted in an inhibition of tumor-initiating and colony-forming properties when implanted in nude mice and grown in methyl-cellulose medium, respectively [38]. This implies that therapies targeting the Notch pathway, such as agents that induce an over-expression of Notch1, may function as a novel means of elimination therapy in human CSCs.
STAT3 plays a role in several processes, including cell survival, proliferation, differentiation, oncogenesis, metastasis, immune invasion, and angiogenesis, under both physiological and pathological conditions [33, 76]. It was recently shown that a small subpopulation of human self-renewing bone sarcoma cells have the ability to form spherical suspended clonal colonies known as “sarcospheres” under serum-starved and anchorage-independent conditions. These sarcospheres expressed activated STAT3 as well as Nanog, Oct3, and Oct4, the latter three of which are markers of pluripotent ES cells. It was shown that as the sarcospheres grew larger and the total number of cells contained therein increased, they adopted an increasingly heterogeneous genotype with a lower percentage of the cells expressing the ES cell markers [58]. The ability of STAT3, in conjunction with Nanog, Oct3, and Oct4, to maintain the “stemness” of the bone sarcoma sarcospheres coupled with its extensively studied over-expression in several human cancers makes it an increasingly attractive and important target for therapy against both bulk tumor cells and CSCs.
A target that may result in increased tumor-selective toxicity to CSCs is telomerase. In most cells, the telomere (chromosome end-sequence) shortens as they divide. The length of the telomeres is preserved by an enzyme called telomerase (RNA template-dependent DNA-synthase), which adds relatively short repeats of nucleotides to chromosome ends. Normal stem cells require telomerase in order to prevent a shortening of their telomeres and replicative senescence (loss of telomere length reserve), although it has been shown that the relatively long telomeres of normal stem cells allows them to temporarily proliferate in the absence of telomerase. It has also been suggested that the presence of telomerase and its subsequent activity in most malignancies is an absolute requirement for growth and maintenance as a means of compensating for the characteristically short telomeres in cancer cells [67, 86]. Taken together, this implies that the inhibition of telomerase may not only represent a potent therapeutic approach against CSCs, but one that offers a reasonable degree of tumor specificity and differential sensitivity due to the differences in telomere length between normal stem cells and CSCs. GRN163L, a direct inhibitor of the RNA template region of human telomerase (hTR), is a 13-mer oligonucleotide lipid conjugate that is currently in clinical trials for patients with chronic lymphocytic leukemia, multiple myeloma, solid tumors, and non-small-cell lung cancer [67]. It is also possible that GRN163L will play a role in eliminating CSCs with active telomerase and reduce the rate of recurrence.
Alternative to the aforementioned biochemical targets for novel therapeutic development against CSCs there are also a number of physiological targets. Among these are the intracellular ROS levels [166] as well as the tumor vasculature [166, 180] that nourishes and supplies CSCs. ROS play a role in tumor initiation in several animal models and humans. Among other factors, superoxide dismutases dependent on bivalent cations play an important role in neutralizing the more active forms of ROS; thus the removal of some bivalent cations kills cancer cells [69]. Although little is known of how ROS status directly relates to CSCs, it is has been suggested that the cellular redox state plays a role in the balance between self-renewal and the differentiation potential of stem cells. It was shown that extracellular growth factors that promote self-renewal and differentiation caused progenitor cells to be more reduced and oxidized, respectively [166]. If more conclusive studies are done that link ROS status with CSCs, this may provide an exploitable target for therapy. Since CSCs exist among a heterogeneous bulk tumor population supplied by a tumor vasculature, it is logical to assume that inhibition of such vessels could also terminate the nutrient supply to CSCs and reduce their resistance to radio- and chemotherapy [180]. Coupling angiogenesis inhibitors with targeted inhibitors of CD133, STAT3, and hTR or the HH/PTCH, Wnt, and Notch pathways may provide a novel and effective means of significantly improving cancer therapy and reducing remission across many malignant tissue types.
Aside from therapeutic approaches that focus on certain known pathways, other novel ways to attack cancer have recently been proposed as well. An interesting example is proteins that either selectively (e.g. apoptin) [28, 106] or semi-selectively (e.g. Brevinin-2R, S100A8/A9) [55, 56, 63] kill cancer cells by mechanism(s) yet to be fully defined. In the case of apoptin, however, it seems that its selectivity is dependent upon the utilization of the very pathways involved in cell proliferation, such as PI3-K/Akt/CDK2, and their targeting (or “hijacking”) to induce cell death, likely via a mitotic catastrophe-related mechanism [104, 105, 107]. Interestingly, opposing processes such as cell survival, cell death, and cell proliferation seem to be tightly interconnected in the cell, and some stimuli may have opposite effects depending on the quantity or duration [57, 103].
Acknowledgments
S. Hombach-Klonisch would like to thank the Manitoba Health Research Council (MHRC) for the generous support. M. Los thankfully acknowledges the support by the Canada Research Chair program (CFI), PCRFC-, CIHR, CIHR-IPM, CCMF-, and MHRC-financed programs.
Abbreviations
- BCC
basal cell cancer
- BM
bone marrow
- BMP
bone morphogenetic protein
- CHK2
checkpoint kinase2
- CSC
cancer stem cell
- CXCL12
chemokine (C-X-C-motif) ligand 12
- ES
embryonic stem
- Flk-1
fetal liver kinase-1
- GCT
germ cell tumor
- GPI
guanyl phosphatidylinositol
- HH/PTCH
hedgehog/patched pathway
- HSC
hematopoietic stem cells
- IGCNU
intratubular germ cell neoplasia unclassified
- KDR
kinase insert domain-containing receptor
- LRP5/6
low-density lipoprotein receptor-related protein 5/6
- MIS
Müllerian inhibiting substance
- MSC
mesenchymal stem cell
- OGCT
ovarian GCT
- OSCC
oral squamous cell carcinoma
- PGC
primordial germ cell
- ROS
reactive oxygen species
- Sca-1
stem cell antigen-1
- SCF-1
stem cell factor-1
- SHH
sonic hedgehog
- SMO
smoothened
- SP
side population
- STAT3
signal transducer and activator of transcription 3
- TGCT
testicular GCTs
- VEGFR2
vascular endothelial growth factor receptor 2
- VCAM
vascular cell adhesion molecule
- FCS
fetal calf serum
References
- 1.Adami HO, Bergstrom R, Mohner M, Zatonski W, Storm H, Ekbom A, Tretli S, Teppo L, Ziegler H, Rahu M, et al. Testicular cancer in nine northern European countries. Int J Cancer. 1994;59:33–38. doi: 10.1002/ijc.2910590108. [DOI] [PubMed] [Google Scholar]
- 2.Adams GB, Scadden DT. The hematopoietic stem cell in its place. Nat Immunol. 2006;7:333–337. doi: 10.1038/ni1331. [DOI] [PubMed] [Google Scholar]
- 3.Alexander HK, Booy EP, Xiao W, Ezzati P, Baust H, Los M. Selected technologies to control genes and their products for experimental and clinical purposes. Arch Immunol Ther Exp. 2007;55:139–149. doi: 10.1007/s00005-007-0025-7. [DOI] [PubMed] [Google Scholar]
- 4.Almstrup K, Hoei-Hansen CE, Wirkner U, Blake J, Schwager C, Ansorge W, Nielsen JE, Skakkebaek NE, Rajpert-De Meyts E, Leffers H. Embryonic stem cell-like features of testicular carcinoma in situ revealed by genome-wide gene expression profiling. Cancer Res. 2004;64:4736–4743. doi: 10.1158/0008-5472.CAN-04-0679. [DOI] [PubMed] [Google Scholar]
- 5.Almstrup K, Ottesen AM, Sonne SB, Hoei-Hansen CE, Leffers H, Rajpert-De Meyts E, Skakkebaek NE. Genomic and gene expression signature of the pre-invasive testicular carcinoma in situ. Cell Tissue Res. 2005;322:159–165. doi: 10.1007/s00441-005-1084-x. [DOI] [PubMed] [Google Scholar]
- 6.Anderson JE, Hansen LL, Mooren FC, Post M, Hug H, Zuse A, Los M. Methods and biomarkers for the diagnosis and prognosis of cancer and other diseases: Towards personalized medicine. Drug Resist Updat. 2006;9:198–210. doi: 10.1016/j.drup.2006.08.001. [DOI] [PubMed] [Google Scholar]
- 7.Anderson R, Copeland TK, Scholer H, Heasman J, Wylie C. The onset of germ cell migration in the mouse embryo. Mech Dev. 2000;91:61–68. doi: 10.1016/s0925-4773(99)00271-3. [DOI] [PubMed] [Google Scholar]
- 8.Anfosso F, Bardin N, Frances V, Vivier E, Camoin-Jau L, Sampol J, Dignat-George F. Activation of human endothelial cells via S-endo-1 antigen (CD146) stimulates the tyrosine phosphorylation of focal adhesion kinase p125(FAK) J Biol Chem. 1998;273:26852–26856. doi: 10.1074/jbc.273.41.26852. [DOI] [PubMed] [Google Scholar]
- 9.Arai F, Hirao A, Ohmura M, Sato H, Matsuoka S, Takubo K, Ito K, Koh GY, Suda T. Tie2/angiopoietin-1 signaling regulates hematopoietic stem cell quiescence in the bone marrow niche. Cell. 2004;118:149–161. doi: 10.1016/j.cell.2004.07.004. [DOI] [PubMed] [Google Scholar]
- 10.Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T, Witzenbichler B, Schatteman G, Isner JM. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997;275:964–967. doi: 10.1126/science.275.5302.964. [DOI] [PubMed] [Google Scholar]
- 11.Ashman LK, Cambareri AC, To LB, Levinsky RJ, Juttner CA. Expression of the YB5. B8 antigen (c-kit proto-oncogene product) in normal human bone marrow. Blood. 1991;78:30–37. [PubMed] [Google Scholar]
- 12.Aust G, Eichler W, Laue S, Lehmann I, Heldin NE, Lotz O, Scherbaum WA, Dralle H, Hoang-Vu C. CD97: a dedifferentiation marker in human thyroid carcinomas. Cancer Res. 1997;57:1798–1806. [PubMed] [Google Scholar]
- 13.Baatenburg de Jong RJ, Hermans J, Molenaar J, Briaire JJ, le Cessie S. Prediction of survival in patients with head and neck cancer. Head Neck. 2001;23:718–724. doi: 10.1002/hed.1102. [DOI] [PubMed] [Google Scholar]
- 14.Baddoo M, Hill K, Wilkinson R, Gaupp D, Hughes C, Kopen GC, Phinney DG. Characterization of mesenchymal stem cells isolated from murine bone marrow by negative selection. J Cell Biochem. 2003;89:1235–1249. doi: 10.1002/jcb.10594. [DOI] [PubMed] [Google Scholar]
- 15.Banerji S, Los M. Important differences between topoisomerase-I and -II targeting agents. Cancer Biol Ther. 2006;5:965–966. doi: 10.4161/cbt.5.8.3274. [DOI] [PubMed] [Google Scholar]
- 16.Bapat SA, Mali AM, Koppikar CB, Kurrey NK. Stem and progenitor-like cells contribute to the aggressive behavior of human epithelial ovarian cancer. Cancer Res. 2005;65:3025–3029. doi: 10.1158/0008-5472.CAN-04-3931. [DOI] [PubMed] [Google Scholar]
- 17.Barresi V, Cerasoli S, Vitarelli E, Tuccari G. Density of microvessels positive for CD105 (endoglin) is related to prognosis in meningiomas. Acta Neuropathol. 2007;114:147–156. doi: 10.1007/s00401-007-0251-4. [DOI] [PubMed] [Google Scholar]
- 18.Baum CM, Weissman IL, Tsukamoto AS, Buckle AM, Peault B. Isolation of a candidate human hematopoietic stem-cell population. Proc Natl Acad Sci USA. 1992;89:2804–2808. doi: 10.1073/pnas.89.7.2804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Biermann K, Goke F, Nettersheim D, Eckert D, Zhou H, Kahl P, Gashaw I, Schorle H, Buttner R. c-KIT is frequently mutated in bilateral germ cell tumours and down-regulated during progression from intratubular germ cell neoplasia to seminoma. J Pathol. 2007;213:311–318. doi: 10.1002/path.2225. [DOI] [PubMed] [Google Scholar]
- 20.Biermann K, Heukamp LC, Steger K, Zhou H, Franke FE, Guetgemann I, Sonnack V, Brehm R, Berg J, Bastian PJ, Muller SC, Wang-Eckert L, Schorle H, Buttner R. Gene expression profiling identifies new biological markers of neoplastic germ cells. Anticancer Res. 2007;27:3091–3100. [PubMed] [Google Scholar]
- 21.Biermann K, Heukamp LC, Steger K, Zhou H, Franke FE, Sonnack V, Brehm R, Berg J, Bastian PJ, Muller SC, Wang-Eckert L, Buettner R. Genome--wide expression profiling reveals new insights into pathogenesis and progression of testicular germ cell tumors. Cancer Genomics Proteomics. 2007;4:359–367. [PubMed] [Google Scholar]
- 22.Bignell G, Smith R, Hunter C, Stephens P, Davies H, Greenman C, Teague J, Butler A, Edkins S, Stevens C, O’Meara S, Parker A, Avis T, Barthorpe S, Brackenbury L, Buck G, Clements J, Cole J, Dicks E, Edwards K, Forbes S, Gorton M, Gray K, Halliday K, Harrison R, Hills K, Hinton J, Jones D, Kosmidou V, Laman R, Lugg R, Menzies A, Perry J, Petty R, Raine K, Shepherd R, Small A, Solomon H, Stephens Y, Tofts C, Varian J, Webb A, West S, Widaa S, Yates A, Gillis AJ, Stoop HJ, van Gurp RJ, Oosterhuis JW, Looijenga LH, Futreal PA, Wooster R, Stratton MR. Sequence analysis of the protein kinase gene family in human testicular germ-cell tumors of adolescents and adults. Genes Chromosomes Cancer. 2006;45:42–46. doi: 10.1002/gcc.20265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Boiret N, Rapatel C, Veyrat-Masson R, Guillouard L, Guerin JJ, Pigeon P, Descamps S, Boisgard S, Berger MG. Characterization of nonexpanded mesenchymal progenitor cells from normal adult human bone marrow. Exp Hematol. 2005;33:219–225. doi: 10.1016/j.exphem.2004.11.001. [DOI] [PubMed] [Google Scholar]
- 24.Booy EP, Johar D, Maddika S, Pirzada H, Sahib MM, Gehrke I, Loewen SD, Louis SD, Kadkhoda K, Mowat M, Los M. Monoclonal and bispecific antibodies as novel therapeutics. Arch Immunol Ther Exp. 2006;54:85–101. doi: 10.1007/s00005-006-0011-5. [DOI] [PubMed] [Google Scholar]
- 25.Boyle JO, Hakim J, Koch W, van der Riet P, Hruban RH, Roa RA, Correo R, Eby YJ, Ruppert JM, Sidransky D. The incidence of p53 mutations increases with progression of head and neck cancer. Cancer Res. 1993;53:4477–4480. [PubMed] [Google Scholar]
- 26.Burek M, Maddika S, Burek CJ, Daniel PT, Schulze--Osthoff K, Los M. Apoptin-induced cell death is modulated by Bcl-2 family members and is Apaf-1 dependent. Oncogene. 2006;25:2213–2222. doi: 10.1038/sj.onc.1209258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bray I, Brennan P, Boffetta P. Projections of alcohol- and tobacco-related cancer mortality in Central Europe. Int J Cancer. 2000;87:122–128. doi: 10.1002/1097-0215(20000701)87:1<122::aid-ijc18>3.0.co;2-w. [DOI] [PubMed] [Google Scholar]
- 28.Bryder D, Rossi DJ, Weissman IL. Hematopoietic stem cells: the paradigmatic tissue-specific stem cell. Am J Pathol. 2006;169:338–346. doi: 10.2353/ajpath.2006.060312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Cannistra SA. Cancer of the ovary. N Engl J Med. 1993;329:1550–1559. doi: 10.1056/NEJM199311183292108. [DOI] [PubMed] [Google Scholar]
- 30.Cervantes RB, Stringer JR, Shao C, Tischfield JA, Stambrook PJ. Embryonic stem cells and somatic cells differ in mutation frequency and type. Proc Natl Acad Sci USA. 2002;99:3586–3590. doi: 10.1073/pnas.062527199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Challen GA, Little MH. A side order of stem cells: the SP phenotype. Stem Cells. 2006;24:3–12. doi: 10.1634/stemcells.2005-0116. [DOI] [PubMed] [Google Scholar]
- 32.Chen Q, Samaranayake LP, Zhen X, Luo G, Nie M, Li B. Up-regulation of Fas ligand and down--regulation of Fas expression in oral carcinogenesis. Oral Oncol. 1999;35:548–553. doi: 10.1016/s1368-8375(99)00029-9. [DOI] [PubMed] [Google Scholar]
- 33.Chen Z, Han ZC. STAT3: A critical transcription activator in angiogenesis. Med Res Rev. 2008;28:185–200. doi: 10.1002/med.20101. [DOI] [PubMed] [Google Scholar]
- 34.Cheng L, Thomas A, Roth LM, Zheng W, Michael H, Karim FW. OCT4: a novel biomarker for dysgerminoma of the ovary. Am J Surg Pathol. 2004;28:1341–1346. doi: 10.1097/01.pas.0000135528.03942.1f. [DOI] [PubMed] [Google Scholar]
- 35.Chew JL, Loh YH, Zhang W, Chen X, Tam WL, Yeap LS, Li P, Ang YS, Lim B, Robson P, Ng HH. Reciprocal transcriptional regulation of Pou5f1 and Sox2 via the Oct4/Sox2 complex in embryonic stem cells. Mol Cell Biol. 2005;25:6031–6046. doi: 10.1128/MCB.25.14.6031-6046.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Collins AT, Berry PA, Hyde C, Stower MJ, Maitland NJ. Prospective identification of tumorigenic prostate cancer stem cells. Cancer Res. 2005;65:10946–10951. doi: 10.1158/0008-5472.CAN-05-2018. [DOI] [PubMed] [Google Scholar]
- 37.Costea DE, Tsinkalovsky O, Vintermyr OK, Johannessen AC, Mackenzie IC. Cancer stem cells – new and potentially important targets for the therapy of oral squamous cell carcinoma. Oral Dis. 2006;12:443–454. doi: 10.1111/j.1601-0825.2006.01264.x. [DOI] [PubMed] [Google Scholar]
- 38.Crisan M, Zheng B, Yap S, Sun B, Giacobino JP, Casteilla L, Buhring H-J, Lazzari L, Huard J, Peault B. The human blood vessel wall as a source of multi-lineage progenitor cells. Conference on “Biology and Clinical Applications of Cord Blood Cells”; 19–21 October 2007; Paris, France. 2007. [Google Scholar]
- 39.de Jong J, Looijenga LH. Stem cell marker OCT3/4 in tumor biology and germ cell tumor diagnostics: history and future. Crit Rev Oncog. 2006;12:171–203. doi: 10.1615/critrevoncog.v12.i3-4.10. [DOI] [PubMed] [Google Scholar]
- 40.de Jong J, Stoop H, Gillis AJ, Hersmus R, van Gurp RJ, van de Geijn GJ, van Drunen E, Beverloo HB, Schneider DT, Sherlock JK, Baeten J, Kitazawa S, van Zoelen EJ, van Roozendaal K, Oosterhuis JW, Looijenga LH. Further characterization of the first seminoma cell line TCam-2. Genes Chromosomes Cancer. 2008;47:185–196. doi: 10.1002/gcc.20520. [DOI] [PubMed] [Google Scholar]
- 41.de Jong J, Stoop H, Gillis AJ, van Gurp RJ, van Drunen E, Beverloo HB, Lau YF, Schneider DT, Sherlock JK, Baeten J, Hatakeyama S, Ohyama C, Oosterhuis JW, Looijenga LH. JKT-1 is not a human seminoma cell line. Int J Androl. 2007;30:350–365. doi: 10.1111/j.1365-2605.2007.00802.x. [DOI] [PubMed] [Google Scholar]
- 42.Decraene C, Benchaouir R, Dillies MA, Israeli D, Bortoli S, Rochon C, Rameau P, Pitaval A, Tronik-Le Roux D, Danos O, Gidrol X, Garcia L, Pietu G. Global transcriptional characterization of SP and MP cells from the myogenic C2C12 cell line: effect of FGF6. Physiol Genomics. 2005;23:132–149. doi: 10.1152/physiolgenomics.00141.2004. [DOI] [PubMed] [Google Scholar]
- 43.Draper JS, Smith K, Gokhale P, Moore HD, Maltby E, Johnson J, Meisner L, Zwaka TP, Thomson JA, Andrews PW. Recurrent gain of chromosomes 17q and 12 in cultured human embryonic stem cells. Nat Biotechnol. 2004;22:53–54. doi: 10.1038/nbt922. [DOI] [PubMed] [Google Scholar]
- 44.Dreyfus PA, Chretien F, Chazaud B, Kirova Y, Caramelle P, Garcia L, Butler-Browne G, Gherardi RK. Adult bone marrow-derived stem cells in muscle connective tissue and satellite cell niches. Am J Pathol. 2004;164:773–779. doi: 10.1016/S0002-9440(10)63165-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Eckardt A, Barth EL, Kokemueller H, Wegener G. Recurrent carcinoma of the head and neck: treatment strategies and survival analysis in a 20-year period. Oral Oncol. 2004;40:427–432. doi: 10.1016/j.oraloncology.2003.09.019. [DOI] [PubMed] [Google Scholar]
- 46.Eckert D, Nettersheim D, Heukamp LC, Kitazawa S, Biermann K, Schorle H. TCam-2 but not JKT-1 cells resemble seminoma in cell culture. Cell Tissue Res. 2008;331:529–538. doi: 10.1007/s00441-007-0527-y. [DOI] [PubMed] [Google Scholar]
- 47.Eichler W, Hamann J, Aust G. Expression characteristics of the human CD97 antigen. Tissue Antigens. 1997;50:429–438. doi: 10.1111/j.1399-0039.1997.tb02897.x. [DOI] [PubMed] [Google Scholar]
- 48.Encina NR, Billotte WG, Hofmann MC. Immunomagnetic isolation of osteoprogenitors from human bone marrow stroma. Lab Invest. 1999;79:449–457. [PubMed] [Google Scholar]
- 49.Farnie G, Clarke RB. Mammary stem cells and breast cancer–role of Notch signalling. Stem Cell Rev. 2007;3:169–175. doi: 10.1007/s12015-007-0023-5. [DOI] [PubMed] [Google Scholar]
- 50.Ferrandina G, Bonanno G, Pierelli L, Perillo A, Procoli A, Mariotti A, Corallo M, Martinelli E, Rutella S, Paglia A, Zannoni G, Mancuso S, Scambia G. Expression of CD133-1 and CD133-2 in ovarian cancer. Int J Gynecol Cancer. 2008;18:506–514. doi: 10.1111/j.1525-1438.2007.01056.x. [DOI] [PubMed] [Google Scholar]
- 51.Filip S, English D, Mokry J. Issues in stem cell plasticity. J Cell Mol Med. 2004;8:572–577. doi: 10.1111/j.1582-4934.2004.tb00483.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Fox JM, Chamberlain G, Ashton BA, Middleton J. Recent advances into the understanding of mesenchymal stem cell trafficking. Br J Haematol. 2007;137:491–502. doi: 10.1111/j.1365-2141.2007.06610.x. [DOI] [PubMed] [Google Scholar]
- 53.Fuchs E. Epidermal differentiation: the bare essentials. J Cell Biol. 1990;111:2807–2814. doi: 10.1083/jcb.111.6.2807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Gastman BR, Atarshi Y, Reichert TE, Saito T, Balkir L, Rabinowich H, Whiteside TL. Fas ligand is expressed on human squamous cell carcinomas of the head and neck, and it promotes apoptosis of T lymphocytes. Cancer Res. 1999;59:5356–5364. [PubMed] [Google Scholar]
- 55.Ghavami S, Asoodeh A, Klonisch T, Halayko AJ, Kadkhoda K, Kroczak TJ, Gibson SB, Booy EP, Naderi-Manesh H, Los M. Brevinin-2R semi--selectively kills cancer cells by a distinct mechanism, which involves the lysosomal-mitochondrial death pathway. J Cell Mol Med. 2008 doi: 10.1111/j.1582-4934.2008.00129.x. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ghavami S, Kerkhoff C, Chazin WJ, Kadkhoda K, Xiao W, Zuse A, Hashemi M, Eshraghi M, Schulze-Osthoff K, Klonisch T, Los M. S100A8/9 induces cell death via a novel, RAGE-independent pathway that involves selective release of Smac/DIABLO and Omi/HtrA2. Biochim Biophys Acta. 2008;1783:297–311. doi: 10.1016/j.bbamcr.2007.10.015. [DOI] [PubMed] [Google Scholar]
- 57.Ghavami S, Rashedi I, Dattilo BM, Eshraghi M, Chazin WJ, Hashemi M, Wesselborg S, Kerkhoff C, Los M. S100A8/A9 at low concentration, promotes tumor cell growth via RAGE ligation and MAP-kinase dependent pathway. J Leukoc Biol. 2008 doi: 10.1189/jlb.0607397. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Gibbs CP, Kukekov VG, Reith JD, Tchigrinova O, Suslov ON, Scott EW, Ghivizzani SC, Ignatova TN, Steindler DA. Stem-like cells in bone sarcomas: implications for tumorigenesis. Neoplasia. 2005;7:967–976. doi: 10.1593/neo.05394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Goddard NC, McIntyre A, Summersgill B, Gilbert D, Kitazawa S, Shipley J. KIT and RAS signalling pathways in testicular germ cell tumours: new data and a review of the literature. Int J Androl. 2007;30:337–348. doi: 10.1111/j.1365-2605.2007.00769.x. [DOI] [PubMed] [Google Scholar]
- 60.Goto T, Adjaye J, Rodeck CH, Monk M. Identification of genes expressed in human primordial germ cells at the time of entry of the female germ line into meiosis. Mol Hum Reprod. 1999;5:851–860. doi: 10.1093/molehr/5.9.851. [DOI] [PubMed] [Google Scholar]
- 61.Gottesman MM, Fojo T, Bates SE. Multidrug resistance in cancer: role of ATP-dependent transporters. Nat Rev Cancer. 2002;2:48–58. doi: 10.1038/nrc706. [DOI] [PubMed] [Google Scholar]
- 62.Gronthos S, Graves SE, Ohta S, Simmons PJ. The STRO-1+ fraction of adult human bone marrow contains the osteogenic precursors. Blood. 1994;84:4164–4173. [PubMed] [Google Scholar]
- 63.Grote J, Konig S, Ackermann D, Sopalla C, Benedyk M, Los M, Kerkhoff C. Identification of poly(ADP-ribose)polymerase-1 and Ku70/Ku80 as transcriptional regulators of S100A9 gene expression. BMC Mol Biol. 2006;7:48. doi: 10.1186/1471-2199-7-48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Hamann J, Eichler W, Hamann D, Kerstens HM, Poddighe PJ, Hoovers JM, Hartmann E, Strauss M, van Lier RA. Expression cloning and chromosomal mapping of the leukocyte activation antigen CD97, a new seven-span transmembrane molecule of the secretion receptor superfamily with an unusual extracellular domain. J Immunol. 1995;155:1942–1950. [PubMed] [Google Scholar]
- 65.Hansis C, Grifo JA, Krey LC. Oct-4 expression in inner cell mass and trophectoderm of human blastocysts. Mol Hum Reprod. 2000;6:999–1004. doi: 10.1093/molehr/6.11.999. [DOI] [PubMed] [Google Scholar]
- 66.Haraguchi N, Utsunomiya T, Inoue H, Tanaka F, Mimori K, Barnard GF, Mori M. Characterization of a side population of cancer cells from human gastrointestinal system. Stem Cells. 2006;24:506–513. doi: 10.1634/stemcells.2005-0282. [DOI] [PubMed] [Google Scholar]
- 67.Harley CB. Telomerase and cancer therapeutics. Nat Rev Cancer. 2008;7:7. doi: 10.1038/nrc2275. [DOI] [PubMed] [Google Scholar]
- 68.Harper LJ, Piper K, Common J, Fortune F, Mackenzie IC. Stem cell patterns in cell lines derived from head and neck squamous cell carcinoma. J Oral Pathol Med. 2007;36:594–603. doi: 10.1111/j.1600-0714.2007.00617.x. [DOI] [PubMed] [Google Scholar]
- 69.Hashemi M, Ghavami S, Eshraghi M, Booy EP, Los M. Cytotoxic effects of intra and extracellular zinc chelation on human breast cancer cells. Eur J Pharmacol. 2007;557:9–19. doi: 10.1016/j.ejphar.2006.11.010. [DOI] [PubMed] [Google Scholar]
- 70.Hatakeyama S, Ohyama C, Minagawa S, Inoue T, Kakinuma H, Kyan A, Arai Y, Suga T, Nakayama J, Kato T, Habuchi T, Fukuda MN. Functional correlation of trophinin expression with the malignancy of testicular germ cell tumor. Cancer Res. 2004;64:4257–4262. doi: 10.1158/0008-5472.CAN-04-0732. [DOI] [PubMed] [Google Scholar]
- 71.Hirschmann-Jax C, Foster AE, Wulf GG, Nuchtern JG, Jax TW, Gobel U, Goodell MA, Brenner MK. A distinct “side population” of cells with high drug efflux capacity in human tumor cells. Proc Natl Acad Sci USA. 2004;101:14228–14233. doi: 10.1073/pnas.0400067101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Hoei-Hansen CE, Kraggerud SM, Abeler VM, Kaern J, Rajpert-De Meyts E, Lothe RA. Ovarian dysgerminomas are characterised by frequent KIT mutations and abundant expression of pluripotency markers. Mol Cancer. 2007;6:12. doi: 10.1186/1476-4598-6-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Hombach-Klonisch S, Panigrahi S, Rashedi I, Seifert A, Alberti E, Pocar P, Kurpisz M, Klonisch T, Schulze--Osthoff K, Mackiewicz A, Los M. Adult stem cells and their trans-differentiation potential – perspectives and therapeutic applications. Curr Med Chem. 2008 doi: 10.1007/s00109-008-0383-6. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Hong Y, Stambrook PJ. Restoration of an absent G1 arrest and protection from apoptosis in embryonic stem cells after ionizing radiation. Proc Natl Acad Sci USA. 2004;101:14443–14448. doi: 10.1073/pnas.0401346101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Horwitz EM, Gordon PL, Koo WK, Marx JC, Neel MD, McNall RY, Muul L, Hofmann T. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: Implications for cell therapy of bone. Proc Natl Acad Sci USA. 2002;99:8932–8937. doi: 10.1073/pnas.132252399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Houghton J. Bone-marrow-derived cells and cancer – an opportunity for improved therapy. Nat Clin Pract Oncol. 2007;4:2–3. doi: 10.1038/ncponc0686. [DOI] [PubMed] [Google Scholar]
- 77.Ikuta K, Weissman IL. Evidence that hematopoietic stem cells express mouse c-kit but do not depend on steel factor for their generation. Proc Natl Acad Sci USA. 1992;89:1502–1506. doi: 10.1073/pnas.89.4.1502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Imai Y, Sasaki T, Shinagawa Y, Akimoto K, Fujibayashi T. Expression of metastasis suppressor gene (KAI1/CD82) in oral squamous cell carcinoma and its clinico-pathological significance. Oral Oncol. 2002;38:557–561. doi: 10.1016/s1368-8375(01)00120-8. [DOI] [PubMed] [Google Scholar]
- 79.Irie T, Aida T, Tachikawa T. Gene expression profiling of oral squamous cell carcinoma using laser microdissection and cDNA microarray. Med Electron Microsc. 2004;37:89–96. doi: 10.1007/s00795-003-0242-z. [DOI] [PubMed] [Google Scholar]
- 80.Jemal A, Murray T, Ward E, Samuels A, Tiwari RC, Ghafoor A, Feuer EJ, Thun MJ. Cancer statistics, 2005. CA Cancer J Clin. 2005;55:10–30. doi: 10.3322/canjclin.55.1.10. [DOI] [PubMed] [Google Scholar]
- 81.Jo Y, Kinugawa K, Matsuki T, Morioka M, Tanaka H. Analysis of the biological properties and use of comparative genomic hybridization to locate chromosomal aberrations in the human testicular seminoma cell line JKT-1 and its highly metastatic cell line JKT-HM. BJU Int. 1999;83:469–475. doi: 10.1046/j.1464-410x.1999.00928.x. [DOI] [PubMed] [Google Scholar]
- 82.Johnson J, Canning J, Kaneko T, Pru JK, Tilly JL. Germline stem cells and follicular renewal in the postnatal mammalian ovary. Nature. 2004;428:145–150. doi: 10.1038/nature02316. [DOI] [PubMed] [Google Scholar]
- 83.Johnston JB, Navaratnam S, Pitz MW, Maniate JM, Wiechec E, Baust H, Gingerich J, Skliris GP, Murphy LC, Los M. Targeting the EGFR pathway for cancer therapy. Curr Med Chem. 2006;13:3483–3492. doi: 10.2174/092986706779026174. [DOI] [PubMed] [Google Scholar]
- 84.Katoh M. WNT signaling pathway and stem cell signaling network. Clin Cancer Res. 2007;13:4042–4045. doi: 10.1158/1078-0432.CCR-06-2316. [DOI] [PubMed] [Google Scholar]
- 85.Kawamoto H, Ohmura K, Katsura Y. Direct evidence for the commitment of hematopoietic stem cells to T, B and myeloid lineages in murine fetal liver. Int Immunol. 1997;9:1011–1019. doi: 10.1093/intimm/9.7.1011. [DOI] [PubMed] [Google Scholar]
- 86.Keith WN, Thomson CM, Howcroft J, Maitland NJ, Shay JW. Seeding drug discovery: integrating telomerase cancer biology and cellular senescence to uncover new therapeutic opportunities in targeting cancer stem cells. Drug Discov Today. 2007;12:611–621. doi: 10.1016/j.drudis.2007.06.009. [DOI] [PubMed] [Google Scholar]
- 87.Kerr JB, Duckett R, Myers M, Britt KL, Mladenovska T, Findlay JK. Quantification of healthy follicles in the neonatal and adult mouse ovary: evidence for maintenance of primordial follicle supply. Reproduction. 2006;132:95–109. doi: 10.1530/rep.1.01128. [DOI] [PubMed] [Google Scholar]
- 88.Kinugawa K, Hyodo F, Matsuki T, Jo Y, Furukawa Y, Ueki A, Tanaka H. Establishment and characterization of a new human testicular seminoma cell line, JKT-1. Int J Urol. 1998;5:282–287. doi: 10.1111/j.1442-2042.1998.tb00604.x. [DOI] [PubMed] [Google Scholar]
- 89.Kitazawa S, Takenaka A, Kondo T, Mizoguchi A, Kitazawa R. Protruding disordered loop of gC1qR is specifically exposed and related to antiapoptotic property in germ cell lineage. Histochem Cell Biol. 2006;126:665–677. doi: 10.1007/s00418-006-0225-y. [DOI] [PubMed] [Google Scholar]
- 90.Kobayashi T, Fujii T, Jo Y, Kinugawa K, Fujisawa M. Possible mechanism responsible for the acquisition of resistance to cis-diamminedichloroplatinum (II) by cultured human testicular seminoma cells. J Urol. 2004;171:1929–1933. doi: 10.1097/01.ju.0000122901.70300.20. [DOI] [PubMed] [Google Scholar]
- 91.Kroczak TJ, Baran J, Pryjma J, Siedlar M, Rashedi I, Hernandez E, Alberti E, Meddika S, Los M. The emerging importance of DNA mapping and other comprehensive screening techniques as tools to identify new drug targets and as a mean of (cancer) therapy personalization. Expert Opin Ther Targets. 2006;10:289–302. doi: 10.1517/14728222.10.2.289. [DOI] [PubMed] [Google Scholar]
- 92.Kropveld A, Rozemuller EH, Leppers FG, Scheidel KC, de Weger RA, Koole R, Hordijk GJ, Slootweg PJ, Tilanus MG. Sequencing analysis of RNA and DNA of exons 1 through 11 shows p53 gene alterations to be present in almost 100% of head and neck squamous cell cancers. Lab Invest. 1999;79:347–453. [PubMed] [Google Scholar]
- 93.Krzemieniecki K, Szpyt E, Rashedi I, Gawron K, Los M. Targeting of solid tumors and blood malignancies by antibody-based therapies. Centr Eur J Biol. 2006;1:167–182. [Google Scholar]
- 94.La Vecchia C, Tavani A, Franceschi S, Levi F, Corrao G, Negri E. Epidemiology and prevention of oral cancer. Oral Oncol. 1997;33:302–312. doi: 10.1016/s1368-8375(97)00029-8. [DOI] [PubMed] [Google Scholar]
- 95.Larderet G, Fortunel NO, Vaigot P, Cegalerba M, Maltere P, Zobiri O, Gidrol X, Waksman G, Martin MT. Human side population keratinocytes exhibit long-term proliferative potential and a specific gene expression profile and can form a pluristratified epidermis. Stem Cells. 2006;24:965–974. doi: 10.1634/stemcells.2005-0196. [DOI] [PubMed] [Google Scholar]
- 96.Levine AJ. The tumor suppressor genes. Annu Rev Biochem. 1993;62:623–651. doi: 10.1146/annurev.bi.62.070193.003203. [DOI] [PubMed] [Google Scholar]
- 97.Liu Y, Chen L, Peng S, Chen Z, Gimm O, Finke R, Hoang-Vu C. The expression of CD97EGF and its ligand CD55 on marginal epithelium is related to higher stage and depth of tumor invasion of gastric carcinomas. Oncol Rep. 2005;14:1413–1420. [PubMed] [Google Scholar]
- 98.Liu Y, Chen L, Peng SY, Chen ZX, Hoang-Vu C. Role of CD97(stalk) and CD55 as molecular markers for prognosis and therapy of gastric carcinoma patients. J Zhejiang Univ Sci B. 2005;6:913–918. doi: 10.1631/jzus.2005.B0913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Looijenga LH, de Leeuw H, van Oorschot M, van Gurp RJ, Stoop H, Gillis AJ, de Gouveia Brazao CA, Weber RF, Kirkels WJ, van Dijk T, von Lindern M, Valk P, Lajos G, Olah E, Nesland JM, Fossa SD, Oosterhuis JW. Stem cell factor receptor (c-KIT) codon 816 mutations predict development of bilateral testicular germ-cell tumors. Cancer Res. 2003;63:7674–7678. [PubMed] [Google Scholar]
- 100.Looijenga LH, Stoop H, de Leeuw HP, de Gouveia Brazao CA, Gillis AJ, van Roozendaal KE, van Zoelen EJ, Weber RF, Wolffenbuttel KP, van Dekken H, Honecker F, Bokemeyer C, Perlman EJ, Schneider DT, Kononen J, Sauter G, Oosterhuis JW. POU5F1 (OCT3/4) identifies cells with pluripotent potential in human germ cell tumors. Cancer Res. 2003;63:2244–2250. [PubMed] [Google Scholar]
- 101.Lou H, Dean M. Targeted therapy for cancer stem cells: the patched pathway and ABC transporters. Oncogene. 2007;26:1357–1360. doi: 10.1038/sj.onc.1210200. [DOI] [PubMed] [Google Scholar]
- 102.Lowell S, Jones P, Le Roux I, Dunne J, Watt FM. Stimulation of human epidermal differentiation by delta-notch signalling at the boundaries of stem-cell clusters. Curr Biol. 2000;10:491–500. doi: 10.1016/s0960-9822(00)00451-6. [DOI] [PubMed] [Google Scholar]
- 103.Maddika S, Ande SR, Panigrahi S, Paranjothy T, Weglarczyk K, Zuse A, Eshraghi M, Manda KD, Wiechec E, Los M. Cell survival, cell death and cell cycle pathways are interconnected: Implications for cancer therapy. Drug Resist Updat. 2007;10:13–29. doi: 10.1016/j.drup.2007.01.003. [DOI] [PubMed] [Google Scholar]
- 104.Maddika S, Ande SR, Wiechec E, Hansen LL, Wesselborg S, Los M. Akt mediated phosphorylation of CDK2 regulates its dual role in cell cycle progression and apoptosis. J Cell Sci. 2008;121:979–988. doi: 10.1242/jcs.009530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Maddika S, Bay GH, Kroczak TJ, Ande SR, Maddika S, Wiechec E, Gibson SB, Los M. Akt is transferred to the nucleus of cells treated with apoptin, and it participates in apoptin-induced cell death. Cell Prolif. 2007;40:835–848. doi: 10.1111/j.1365-2184.2007.00475.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Maddika S, Mendoza FJ, Hauff K, Zamzow CR, Paranjothy T, Los M. Cancer-selective therapy of the future: apoptin and its mechanism of action. Cancer Biol Ther. 2006;5:10–19. doi: 10.4161/cbt.5.1.2400. [DOI] [PubMed] [Google Scholar]
- 107.Maddika S, Wiechec E, Ande SR, Poon IK, Fischer U, Wesselborg S, Jans DA, Schulze-Osthoff K, Los M. Interaction with PI3-kinase contributes to the cytotoxic activity of apoptin. Oncogene. 2008;27:3060–3065. doi: 10.1038/sj.onc.1210958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Massard C, Deutsch E, Soria JC. Tumour stem cell-targeted treatment: elimination or differentiation. Ann Oncol. 2006;17:1620–1624. doi: 10.1093/annonc/mdl074. [DOI] [PubMed] [Google Scholar]
- 109.McKiernan JM, Goluboff ET, Liberson GL, Golden R, Fisch H. Rising risk of testicular cancer by birth cohort in the United States from 1973 to 1995. J Urol. 1999;162:361–363. [PubMed] [Google Scholar]
- 110.Mimeault M, Hauke R, Batra SK. Stem cells: a revolution in therapeutics-recent advances in stem cell biology and their therapeutic applications in regenerative medicine and cancer therapies. Clin Pharmacol Ther. 2007;82:252–264. doi: 10.1038/sj.clpt.6100301. [DOI] [PubMed] [Google Scholar]
- 111.Miraglia S, Godfrey W, Yin AH, Atkins K, Warnke R, Holden JT, Bray RA, Waller EK, Buck DW. A novel five-transmembrane hematopoietic stem cell antigen: isolation, characterization, and molecular cloning. Blood. 1997;90:5013–5021. [PubMed] [Google Scholar]
- 112.Mizuno Y, Gotoh A, Kamidono S, Kitazawa S. Establishment and characterization of a new human testicular germ cell tumor cell line (TCam-2) Nippon Hinyokika Gakkai Zasshi. 1993;84:1211–1218. doi: 10.5980/jpnjurol1989.84.1211. [DOI] [PubMed] [Google Scholar]
- 113.Moles JP, Watt FM. The epidermal stem cell compartment: variation in expression levels of E-cadherin and catenins within the basal layer of human epidermis. J Histochem Cytochem. 1997;45:867–874. doi: 10.1177/002215549704500611. [DOI] [PubMed] [Google Scholar]
- 114.Molyneaux KA, Wang Y, Schaible K, Wylie C. Transcriptional profiling identifies genes differentially expressed during and after migration in murine primordial germ cells. Gene Expr Patterns. 2004;4:167–181. doi: 10.1016/j.modgep.2003.09.002. [DOI] [PubMed] [Google Scholar]
- 115.Morrison SJ, Hemmati HD, Wandycz AM, Weissman IL. The purification and characterization of fetal liver hematopoietic stem cells. Proc Natl Acad Sci USA. 1995;92:10302–10306. doi: 10.1073/pnas.92.22.10302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Morrison SJ, Shah NM, Anderson DJ. Regulatory mechanisms in stem cell biology. Cell. 1997;88:287–298. doi: 10.1016/s0092-8674(00)81867-x. [DOI] [PubMed] [Google Scholar]
- 117.Morrison SJ, Wright DE, Weissman IL. Cyclophosphamide/granulocyte colony-stimulating factor induces hematopoietic stem cells to proliferate prior to mobilization. Proc Natl Acad Sci USA. 1997;94:1908–1913. doi: 10.1073/pnas.94.5.1908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Murdoch WJ, Townsend RS, McDonnel AC. Ovulation-induced DNA damage in ovarian surface epithelial cells of ewes: prospective regulatory mechanisms of repair/survival and apoptosis. Biol Reprod. 2001;65:1417–1424. doi: 10.1095/biolreprod65.5.1417. [DOI] [PubMed] [Google Scholar]
- 119.Murray L, Chen B, Galy A, Chen S, Tushinski R, Uchida N, Negrin R, Tricot G, Jagannath S, Vesole D, et al. Enrichment of human hematopoietic stem cell activity in the CD34+Thy-1+Lin– subpopulation from mobilized peripheral blood. Blood. 1995;85:368–378. [PubMed] [Google Scholar]
- 120.Mustafa T, Eckert A, Klonisch T, Kehlen A, Maurer P, Klintschar M, Erhuma M, Zschoyan R, Gimm O, Dralle H, Schubert J, Hoang-Vu C. Expression of the epidermal growth factor seven-transmembrane member CD97 correlates with grading and staging in human oral squamous cell carcinomas. Cancer Epidemiol Biomarkers Prev. 2005;14:108–119. [PubMed] [Google Scholar]
- 121.Ness RB, Cottreau C. Possible role of ovarian epithelial inflammation in ovarian cancer. J Natl Cancer Inst. 1999;91:1459–1467. doi: 10.1093/jnci/91.17.1459. [DOI] [PubMed] [Google Scholar]
- 122.Neville BW, Day TA. Oral cancer and pre-cancerous lesions. CA Cancer J Clin. 2002;52:195–215. doi: 10.3322/canjclin.52.4.195. [DOI] [PubMed] [Google Scholar]
- 123.Nicolas M, Wolfer A, Raj K, Kummer JA, Mill P, van Noort M, Hui CC, Clevers H, Dotto GP, Radtke F. Notch1 functions as a tumor suppressor in mouse skin. Nat Genet. 2003;33:416–421. doi: 10.1038/ng1099. [DOI] [PubMed] [Google Scholar]
- 124.Niwa H, Miyazaki J, Smith AG. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nat Genet. 2000;24:372–376. doi: 10.1038/74199. [DOI] [PubMed] [Google Scholar]
- 125.Nogales F, Talerman A, Kubik-Huch RA, Tavassoli FA, Devouassoux-Shisheboran M. World Health Organization Classification of Tumours: Pathology of Tumours of the Breast and Female Genital Organs. IARC Press; Lyon: 2003. Germ cell tumours. [Google Scholar]
- 126.Ogawa M, Matsuzaki Y, Nishikawa S, Hayashi S, Kunisada T, Sudo T, Kina T, Nakauchi H, Nishikawa S. Expression and function of c-kit in hemopoietic progenitor cells. J Exp Med. 1991;174:63–71. doi: 10.1084/jem.174.1.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Okuyama R, Nguyen BC, Talora C, Ogawa E, Tommasi di Vignano A, Lioumi M, Chiorino G, Tagami H, Woo M, Dotto GP. High commitment of embryonic keratinocytes to terminal differentiation through a Notch1-caspase 3 regulatory mechanism. Dev Cell. 2004;6:551–562. doi: 10.1016/s1534-5807(04)00098-x. [DOI] [PubMed] [Google Scholar]
- 128.Okuyama R, Tagami H, Aiba S. Notch signaling: Its role in epidermal homeostasis and in the pathogenesis of skin diseases. J Dermatol Sci. 2008;49:187–194. doi: 10.1016/j.jdermsci.2007.05.017. [DOI] [PubMed] [Google Scholar]
- 129.Oosterhuis JW, Looijenga LH. Testicular germ-cell tumours in a broader perspective. Nat Rev Cancer. 2005;5:210–222. doi: 10.1038/nrc1568. [DOI] [PubMed] [Google Scholar]
- 130.Osawa M, Hanada K, Hamada H, Nakauchi H. Long-term lymphohematopoietic reconstitution by a single CD34-low/negative hematopoietic stem cell. Science. 1996;273:242–245. doi: 10.1126/science.273.5272.242. [DOI] [PubMed] [Google Scholar]
- 131.Oyajobi BO, Lomri A, Hott M, Marie PJ. Isolation and characterization of human clonogenic osteoblast progenitors immunoselected from fetal bone marrow stroma using STRO-1 monoclonal antibody. J Bone Miner Res. 1999;14:351–361. doi: 10.1359/jbmr.1999.14.3.351. [DOI] [PubMed] [Google Scholar]
- 132.Papayannopoulou T, Brice M, Broudy VC, Zsebo KM. Isolation of c-kit receptor-expressing cells from bone marrow, peripheral blood, and fetal liver: functional properties and composite antigenic profile. Blood. 1991;78:1403–1412. [PubMed] [Google Scholar]
- 133.Pauls K, Schorle H, Jeske W, Brehm R, Steger K, Wernert N, Buttner R, Zhou H. Spatial expression of germ cell markers during maturation of human fetal male gonads: an immunohistochemical study. Hum Reprod. 2006;21:397–404. doi: 10.1093/humrep/dei325. [DOI] [PubMed] [Google Scholar]
- 134.Peichev M, Naiyer AJ, Pereira D, Zhu Z, Lane WJ, Williams M, Oz MC, Hicklin DJ, Witte L, Moore MA, Rafii S. Expression of VEGFR-2 and AC133 by circulating human CD34(+) cells identifies a population of functional endothelial precursors. Blood. 2000;95:952–958. [PubMed] [Google Scholar]
- 135.Pelosi E, Valtieri M, Coppola S, Botta R, Gabbianelli M, Lulli V, Marziali G, Masella B, Muller R, Sgadari C, Testa U, Bonanno G, Peschle C. Identification of the hemangioblast in postnatal life. Blood. 2002;100:3203–3208. doi: 10.1182/blood-2002-05-1511. [DOI] [PubMed] [Google Scholar]
- 136.Penninger JM, Irie-Sasaki J, Sasaki T, Oliveira-dos--Santos AJ. CD45: new jobs for an old acquaintance. Nat Immunol. 2001;2:389–396. doi: 10.1038/87687. [DOI] [PubMed] [Google Scholar]
- 137.Pereira MC, Oliveira DT, Landman G, Kowalski LP. Histologic subtypes of oral squamous cell carcinoma: prognostic relevance. J Can Dent Assoc. 2007;73:339–344. [PubMed] [Google Scholar]
- 138.Pesce M, Gross MK, Scholer HR. In line with our ancestors: Oct-4 and the mammalian germ. Bioessays. 1998;20:722–732. doi: 10.1002/(SICI)1521-1878(199809)20:9<722::AID-BIES5>3.0.CO;2-I. [DOI] [PubMed] [Google Scholar]
- 139.Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak DR. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143–147. doi: 10.1126/science.284.5411.143. [DOI] [PubMed] [Google Scholar]
- 140.Proweller A, Tu L, Lepore JJ, Cheng L, Lu MM, Seykora J, Millar SE, Pear WS, Parmacek MS. Impaired notch signaling promotes de novo squamous cell carcinoma formation. Cancer Res. 2006;66:7438–7444. doi: 10.1158/0008-5472.CAN-06-0793. [DOI] [PubMed] [Google Scholar]
- 141.Rashedi I, Panigrahi S, Ezzati P, Ghavami S, Los M. Autoimmunity and apoptosis – therapeutic implications. Curr Med Chem. 2007;14:3139–3159. doi: 10.2174/092986707782793952. [DOI] [PubMed] [Google Scholar]
- 142.Reber L, Da Silva CA, Frossard N. Stem cell factor and its receptor c-Kit as targets for inflammatory diseases. Eur J Pharmacol. 2006;533:327–340. doi: 10.1016/j.ejphar.2005.12.067. [DOI] [PubMed] [Google Scholar]
- 143.Riopel MA, Spellerberg A, Griffin CA, Perlman EJ. Genetic analysis of ovarian germ cell tumors by comparative genomic hybridization. Cancer Res. 1998;58:3105–3110. [PubMed] [Google Scholar]
- 144.Robey PG. Stem cells near the century mark. J Clin Invest. 2000;105:1489–1491. doi: 10.1172/JCI10256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Rodda DJ, Chew JL, Lim LH, Loh YH, Wang B, Ng HH, Robson P. Transcriptional regulation of nanog by OCT4 and SOX2. J Biol Chem. 2005;280:24731–24737. doi: 10.1074/jbc.M502573200. [DOI] [PubMed] [Google Scholar]
- 146.Roger C, Lambard S, Bouskine A, Mograbi B, Chevallier D, Nebout M, Pointis G, Carreau S, Fenichel P. Estrogen-induced growth inhibition of human seminoma cells expressing estrogen receptor beta and aromatase. J Mol Endocrinol. 2005;35:191–199. doi: 10.1677/jme.1.01704. [DOI] [PubMed] [Google Scholar]
- 147.Rosner MH, Vigano MA, Ozato K, Timmons PM, Poirier F, Rigby PW, Staudt LM. A POU--domain transcription factor in early stem cells and germ cells of the mammalian embryo. Nature. 1990;345:686–692. doi: 10.1038/345686a0. [DOI] [PubMed] [Google Scholar]
- 148.Rufaihah AJ, Haider HK, Heng BC, Ye L, Toh WS, Tian XF, Lu K, Sim EK, Cao T. Directing endothelial differentiation of human embryonic stem cells via transduction with an adenoviral vector expressing the VEGF(165) gene. J Gene Med. 2007;9:452–461. doi: 10.1002/jgm.1034. [DOI] [PubMed] [Google Scholar]
- 149.Schutze K, Lahr G, Burgemeister R. The force of focused light in different areas of cell and molecular biology. J Mol Med. 2000;78:B21. [PubMed] [Google Scholar]
- 150.Shake JG, Gruber PJ, Baumgartner WA, Senechal G, Meyers J, Redmond JM, Pittenger MF, Martin BJ. Mesenchymal stem cell implantation in a swine myocardial infarct model: engraftment and functional effects. Ann Thorac Surg. 2002;73:1919–1925. doi: 10.1016/s0003-4975(02)03517-8. [DOI] [PubMed] [Google Scholar]
- 151.Shi Q, Rafii S, Wu MH, Wijelath ES, Yu C, Ishida A, Fujita Y, Kothari S, Mohle R, Sauvage LR, Moore MA, Storb RF, Hammond WP. Evidence for circulating bone marrow-derived endothelial cells. Blood. 1998;92:362–367. [PubMed] [Google Scholar]
- 152.Shi S, Gronthos S. Perivascular niche of post-natal mesenchymal stem cells in human bone marrow and dental pulp. J Bone Miner Res. 2003;18:696–704. doi: 10.1359/jbmr.2003.18.4.696. [DOI] [PubMed] [Google Scholar]
- 153.Shizuru JA, Negrin RS, Weissman IL. Hematopoietic stem and progenitor cells: clinical and pre-clinical regeneration of the hematolymphoid system. Annu Rev Med. 2005;56:509–538. doi: 10.1146/annurev.med.54.101601.152334. [DOI] [PubMed] [Google Scholar]
- 154.Simmons PJ, Aylett GW, Niutta S, To LB, Juttner CA, Ashman LK. c-kit is expressed by primitive human hematopoietic cells that give rise to colony-forming cells in stroma-dependent or cytokine-supplemented culture. Exp Hematol. 1994;22:157–165. [PubMed] [Google Scholar]
- 155.Simmons PJ, Torok-Storb B. Identification of stromal cell precursors in human bone marrow by a novel monoclonal antibody, STRO-1. Blood. 1991;78:55–62. [PubMed] [Google Scholar]
- 156.Singh SK, Clarke ID, Terasaki M, Bonn VE, Hawkins C, Squire J, Dirks PB. Identification of a cancer stem cell in human brain tumors. Cancer Res. 2003;63:5821–5828. [PubMed] [Google Scholar]
- 157.Sipkins DA, Wei X, Wu JW, Runnels JM, Cote D, Means TK, Luster AD, Scadden DT, Lin CP. In vivo imaging of specialized bone marrow endothelial microdomains for tumour engraftment. Nature. 2005;435:969–973. doi: 10.1038/nature03703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Skakkebaek NE. Possible carcinoma-in-situ of the testis. Lancet. 1972;2:516–517. doi: 10.1016/s0140-6736(72)91909-5. [DOI] [PubMed] [Google Scholar]
- 159.Skakkebaek NE. Carcinoma in situ of the testis: frequency and relationship to invasive germ cell tumours in infertile men. Histopathology. 1978;2:157–170. doi: 10.1111/j.1365-2559.1978.tb01706.x. [DOI] [PubMed] [Google Scholar]
- 160.Smith LG, Weissman IL, Heimfeld S. Clonal analysis of hematopoietic stem-cell differentiation in vivo. Proc Natl Acad Sci USA. 1991;88:2788–2792. doi: 10.1073/pnas.88.7.2788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Spangrude GJ, Brooks DM. Mouse strain variability in the expression of the hematopoietic stem cell antigen Ly-6A/E by bone marrow cells. Blood. 1993;82:3327–3332. [PubMed] [Google Scholar]
- 162.Spangrude GJ, Heimfeld S, Weissman IL. Purification and characterization of mouse hematopoietic stem cells. Science. 1988;241:58–62. doi: 10.1126/science.2898810. [DOI] [PubMed] [Google Scholar]
- 163.Sperger JM, Chen X, Draper JS, Antosiewicz JE, Chon CH, Jones SB, Brooks JD, Andrews PW, Brown PO, Thomson JA. Gene expression patterns in human embryonic stem cells and human pluripotent germ cell tumors. Proc Natl Acad Sci USA. 2003;100:13350–13355. doi: 10.1073/pnas.2235735100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Sutherland DR, Keating A. The CD34 antigen: structure, biology, and potential clinical applications. J Hematother. 1992;1:115–129. doi: 10.1089/scd.1.1992.1.115. [DOI] [PubMed] [Google Scholar]
- 165.Szotek PP, Pieretti-Vanmarcke R, Masiakos PT, Dinulescu DM, Connolly D, Foster R, Dombkowski D, Preffer F, Maclaughlin DT, Donahoe PK. Ovarian cancer side population defines cells with stem cell-like characteristics and Mullerian Inhibiting Substance responsiveness. Proc Natl Acad Sci USA. 2006;103:11154–11159. doi: 10.1073/pnas.0603672103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Tang C, Ang BT, Pervaiz S. Cancer stem cell: target for anti-cancer therapy. FASEB J. 2007;21:3777–3785. doi: 10.1096/fj.07-8560rev. [DOI] [PubMed] [Google Scholar]
- 167.Thelu J, Rossio P, Favier B. Notch signalling is linked to epidermal cell differentiation level in basal cell carcinoma, psoriasis and wound healing. BMC Dermatol. 2002;2:7. doi: 10.1186/1471-5945-2-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Thomas ML. The leukocyte common antigen family. Annu Rev Immunol. 1989;7:339–369. doi: 10.1146/annurev.iy.07.040189.002011. [DOI] [PubMed] [Google Scholar]
- 169.Tintut Y, Alfonso Z, Saini T, Radcliff K, Watson K, Bostrom K, Demer LL. Multilineage potential of cells from the artery wall. Circulation. 2003;108:2505–2510. doi: 10.1161/01.CIR.0000096485.64373.C5. [DOI] [PubMed] [Google Scholar]
- 170.Uchida N, Buck DW, He D, Reitsma MJ, Masek M, Phan TV, Tsukamoto AS, Gage FH, Weissman IL. Direct isolation of human central nervous system stem cells. Proc Natl Acad Sci USA. 2000;97:14720–14725. doi: 10.1073/pnas.97.26.14720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Uchida N, Tsukamoto A, He D, Friera AM, Scollay R, Weissman IL. High doses of purified stem cells cause early hematopoietic recovery in syngeneic and allogeneic hosts. J Clin Invest. 1998;101:961–966. doi: 10.1172/JCI1681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.van de Rijn M, Heimfeld S, Spangrude GJ, Weissman IL. Mouse hematopoietic stem-cell antigen Sca-1 is a member of the Ly-6 antigen family. Proc Natl Acad Sci USA. 1989;86:4634–4638. doi: 10.1073/pnas.86.12.4634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.van de Rijn M, Heimfeld S, Spangrude GJ, Weissman IL. Mouse hematopoietic stem-cell antigen Sca-1 is a member of the Ly-6 antigen family. Proc Natl Acad Sci USA. 1989;86:4634–4638. doi: 10.1073/pnas.86.12.4634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Voorhoeve PM, le Sage C, Schrier M, Gillis AJ, Stoop H, Nagel R, Liu YP, van Duijse J, Drost J, Griekspoor A, Zlotorynski E, Yabuta N, De Vita G, Nojima H, Looijenga LH, Agami R. A genetic screen implicates miRNA-372 and miRNA-373 as oncogenes in testicular germ cell tumors. Adv Exp Med Biol. 2007;604:17–46. doi: 10.1007/978-0-387-69116-9_2. [DOI] [PubMed] [Google Scholar]
- 175.Wallace-Brodeur RR, Lowe SW. Clinical implications of p53 mutations. Cell Mol Life Sci. 1999;55:64–75. doi: 10.1007/s000180050270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Wang L, Li Y, Chen X, Chen J, Gautam SC, Xu Y, Chopp M. MCP-1, MIP-1, IL-8 and ischemic cerebral tissue enhance human bone marrow stromal cell migration in interface culture. Hematology. 2002;7:113–117. doi: 10.1080/10245330290028588. [DOI] [PubMed] [Google Scholar]
- 177.Witte ON. Steel locus defines new multipotent growth factor. Cell. 1990;63:5–6. doi: 10.1016/0092-8674(90)90280-r. [DOI] [PubMed] [Google Scholar]
- 178.Wright DE, Wagers AJ, Gulati AP, Johnson FL, Weissman IL. Physiological migration of hematopoietic stem and progenitor cells. Science. 2001;294:1933–1936. doi: 10.1126/science.1064081. [DOI] [PubMed] [Google Scholar]
- 179.Yamamoto Y, Yasumizu R, Amou Y, Watanabe N, Nishio N, Toki J, Fukuhara S, Ikehara S. Characterization of peripheral blood stem cells in mice. Blood. 1996;88:445–454. [PubMed] [Google Scholar]
- 180.Yang ZJ, Wechsler-Reya RJ. Hit ’em where they live: targeting the cancer stem cell niche. Cancer Cell. 2007;11:3–5. doi: 10.1016/j.ccr.2006.12.007. [DOI] [PubMed] [Google Scholar]
- 181.Yin AH, Miraglia S, Zanjani ED, Almeida-Porada G, Ogawa M, Leary AG, Olweus J, Kearney J, Buck DW. AC133, a novel marker for human hematopoietic stem and progenitor cells. Blood. 1997;90:5002–1012. [PubMed] [Google Scholar]
- 182.Yokoyama K, Kamata N, Hayashi E, Hoteiya T, Ueda N, Fujimoto R, Nagayama M. Reverse correlation of E-cadherin and snail expression in oral squamous cell carcinoma cells in vitro. Oral Oncol. 2001;37:65–71. doi: 10.1016/s1368-8375(00)00059-2. [DOI] [PubMed] [Google Scholar]
- 183.Yu F, Yao H, Zhu P, Zhang X, Pan Q, Gong C, Huang Y, Hu X, Su F, Lieberman J, Song E. let-7 regulates self renewal and tumorigenicity of breast cancer cells. Cell. 2007;131:1109–1123. doi: 10.1016/j.cell.2007.10.054. [DOI] [PubMed] [Google Scholar]
- 184.Zhan Y, Fujino A, MacLaughlin DT, Manganaro TF, Szotek PP, Arango NA, Teixeira J, Donahoe PK. Mullerian inhibiting substance regulates its receptor/SMAD signaling and causes mesenchymal transition of the coelomic epithelial cells early in Mullerian duct regression. Development. 2006;133:2359–2369. doi: 10.1242/dev.02383. [DOI] [PubMed] [Google Scholar]
- 185.Ziegler BL, Valtieri M, Porada GA, De Maria R, Muller R, Masella B, Gabbianelli M, Casella I, Pelosi E, Bock T, Zanjani ED, Peschle C. KDR receptor: a key marker defining hematopoietic stem cells. Science. 1999;285:1553–1558. doi: 10.1126/science.285.5433.1553. [DOI] [PubMed] [Google Scholar]
- 186.Zuk PA. Stem cell research has only just begun. Science. 2001;293:211–212. [PubMed] [Google Scholar]
- 187.Zuse A, Prinz H, Muller K, Schmidt P, Gunther EG, Schweizer F, Prehn JH, Los M. 9-Benzylidene-naphtho[2,3-b]thiophen-4-ones and benzylidene-9(10H)-anthracenones as novel tubulin interacting agents with high apoptosis-inducing activity. Eur J Pharmacol. 2007;575:34–45. doi: 10.1016/j.ejphar.2007.07.050. [DOI] [PubMed] [Google Scholar]