

Abstract
Experiments were carried out on blindfolded human subjects to study the contribution of proprioceptive inputs from both arms in a forearm position matching task. Blindfolded matching accuracy was compared with accuracy when the subject could see their indicator (matching) arm, when they used a dummy arm for matching, and when they looked at a mirror image of the matching arm. The position of the mirror had been arranged so that the image of the indicator arm coincided with the position of the reference arm. None of these conditions significantly altered the matching errors. When reference elbow flexors were vibrated at 70–80 Hz, the illusion of extension of the vibrated arm reported by blindfolded subjects was significantly reduced by vision of the mirror image of the indicator arm or when using the dummy arm. It was concluded that visual information about the position of the indicator arm, or the apparent position of the reference arm, could reduce the size of the kinaesthetic illusion from vibration, but not abolish it. In a second experiment, subjects indicated, by tracking with their vibrated arm, the illusion of forearm extension evoked by elbow flexor vibration. It was found that the perceived speed of extension could be reduced by moving the indicator into extension and increased by moving it into flexion. These experiments demonstrate the importance for the matching process of the input provided by the indicator arm. Such a conclusion may help to explain some apparent discrepancies between observations made on position sense using one-arm and two-arm tasks. More broadly, this paper provides support for the idea that aspects of proprioceptive inputs from both arms are processed conjointly, as part of a strategy for use of the two hands as a single instrument in certain skilled tasks.
Introduction
Traditionally proprioception is considered to comprise four senses, the kinaesthetic sense (position and movement), the sense of tension, the sense of effort and the sense of balance. This paper is concerned with the kinaesthetic sense, which is perhaps the most interesting, given its prominence in everyday activities. It has been argued that the two kinaesthetic senses, the sense of position and the sense of movement, are processed independently (McCloskey, 1973; Proske, 2006; Proske & Gandevia, 2009), even though some of the afferents involved, the primary endings of muscle spindles, contribute to both.
The sense of limb position is commonly measured in the arms using a two-arm matching task (Goodwin et al. 1972). The experimenter places the blindfolded subject's reference arm at a particular elbow angle and the subject is asked to match it by placement of their other, indicator arm. In recent experiments concerned with the role of a motor command signal, perceived as a sense of effort, in limb position sense, the two-arm matching task did not reveal any evidence of an effort contribution when one arm was loaded with a weight (Allen et al. 2007). However other experiments, which used only one arm, did suggest that a component of position sense arose from a signal of central origin (Gandevia et al. 2006; Smith et al. 2009). In those experiments subjects were asked to indicate the position of their unseen hand by moving a pointer directly above it. In such a task the subject is presumably restricted in his or her determination of hand position to signals coming from that hand. In the two-arm matching task there is some evidence that signals coming from both arms are used to determine position (White & Proske, 2009). This raised the question of whether it was possible that in a two-arm matching task, position signals of central origin associated with the sense of effort could be masked in some way as a result of the comparison between the two sides.
Previous experiments have shown that when we match the positions of our arms we are most accurate with our arms alongside the body, hands in front (Gooey et al. 2000). That is the posture we normally adopt when we carry out skilled activities involving both hands acting cooperatively. If one or both arms are raised or abducted, matching accuracy falls (Gooey et al. 2000). All of this raises the question, what exactly is the role of the indicator arm in a two-arm matching task? To what extent does the brain rely on incoming information from both arms and how is this used? A better understanding of such issues may help to reconcile the reported differences in outcomes from the one-arm and two-arm experiments, particularly in relation to the important question of the role of motor commands in kinaesthesia.
In the first experiment reported here we have asked the question, in what way is vision able to contribute to matching performance in a two-arm matching task? The experiment was done both with the unseen reference arm held relaxed and during vibration of its elbow flexors. We have studied the effect on matching accuracy of being able to see the indicator arm, its mirror image, or using a dummy arm. The mirror was arranged so that the image of the indicator arm was in a position where its identity could be mistaken for that of the hidden reference arm. In a second experiment we have asked, does moving the indicator arm alter the illusion of arm movement evoked by vibration of elbow flexor muscles in the reference arm? The wider implications of this work concern our understanding of the ability of human subjects to use their arms cooperatively in the execution of skilled movements, and the work provides new insight into the central neural mechanisms underlying proprioception.
Methods
A total of 25 young adult subjects (21 males, 4 females, mean age = 25.6 years) participated in these experiments. Experiment 1 included 16 subjects and Experiment 2 used 18 subjects. Nine males participated in both experiments. In Experiment 2 four subjects, one female and three male, were excluded after an initial screening because of lack of reproducibility in the movement tracking task (female) and the presence of a strong tonic vibration reflex (males). Subjects gave their written, informed consent before undertaking the experiments, which were approved by the Showa University Committee for Human Experimentation. Ethical aspects of the experiments conformed to the Declaration of Helsinki.
Testing apparatus
The experiment involved matching the positions of the forearms in the vertical plane using a custom built device (Fig. 1A). The subject was blindfolded and sat at a table, with each forearm supported by a paddle, hinged at a point close to the elbow joint. Forearms and paddles were placed in a box with a removable lid so that one or both arms were visible. A partition between the two arms consisted of a mirror. The distance between the mirror and the left arm was adjusted so that the mirror image of the arm coincided with the position of the right arm. The mirror could be covered up.
Figure 1. The apparatus and protocol.
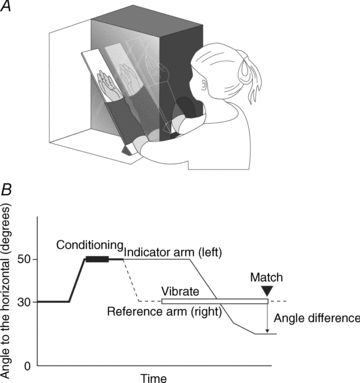
A, subjects sat in front of a two-compartment box. The partition between the compartments was a mirror that showed an image of the left arm. The position of the mirror could be adjusted so that the image of the left arm coincided with the position of the right arm. The mirror could be covered up. Subjects’ arms were taped to paddles which hinged at a point close to the elbow joint. The box had a detachable lid to allow subjects to see one or both arms, as well as permitting the experimenter to hold the arms or place them on supports at the test angle. B, before each trial, both arms were conditioned to put them in a comparable mechanical state. The two arms were placed at an angle of 50 deg to the horizontal and the subject was asked to flex each arm with a contraction of 30–50% of maximum while the arms were held by the experimenter. The right forearm (dashed line) was then placed at the test angle of 30 deg and subjects were asked to match its position with the left arm (continuous line). For the vibration trials the right forearm was placed at 30 deg and vibration of its elbow flexors was commenced. Subjects matched the position of the vibrated arm by placement of their left, indicator arm. When the subjects declared a match, the angle difference between indicator and reference arms was determined. When the indicator arm was placed in a more extended position than the reference arm, the error was expressed as a positive value, if the indicator arm was placed in a more flexed position, the error was expressed as a negative value.
Throughout the experiments the right arm was the reference arm, the left arm the indicator arm. All but one subject were right handed.
Fixed to the side of each paddle was a goniometer (MLTS 700; ADInstruments, Castle Hill, Australia) which provided a continuous signal of the angle of the elbow relative to the horizontal. All data were analysed with Chart software (v. 5.5.6, ADInstruments).
A vibrator was strapped to the belly of biceps of the reference arm by means of elastic straps. The vibrator consisted of a custom-built plastic cylinder which housed a weight mounted eccentrically on the shaft of a small electric motor. The vibrator was 3 cm diameter and 8 cm long. Vibration amplitude was approximately 1 mm and the frequency could be adjusted to lie in the range 70–80 Hz, close to the optimum for engaging responses from muscle spindles (Roll et al. 1989). Because of the small size of the vibrator, its stimulus was relatively localised, acting predominantly on biceps and brachialis. It is not possible to be certain whether some vibration spread to triceps, but the consistency of the illusions it generated and, when present, the direction of any reflex contraction supported the view that the stimulus was largely limited to elbow flexors. In each trial the vibrator was turned on after muscles had been conditioned and the reference arm had been placed at the test angle. Vibration was continued throughout the matching process. The vibrator was turned off as soon as the subject declared that he or she had achieved a satisfactory match. In some subjects a tonic vibration reflex (TVR) developed in elbow flexors during the vibration. This was not a problem in Experiment 1 since the vibrated arm was held at the test angle by the experimenter. In Experiment 2 subjects were asked to focus their attention on the illusion of arm extension produced by the vibration. If they developed a strong TVR they were excluded from further experiment (3 subjects).
Experiment 1
Matching the position illusion from vibration
The procedure used in this experiment is shown diagrammatically in Fig. 1B. Before each trial, both arms were conditioned to place them in a comparable mechanical state (Gregory et al. 1988). To do that, the two arms were placed at an angle of 50 deg to the horizontal and the subject was asked to flex each arm with a contraction of 30–50% of maximum while the arms were held by the experimenter. The right forearm was then placed at the test angle of 30 deg and subjects were asked to match its position with their left, indicator arm. Matching was carried out under four different conditions, each time both with and without vibration of elbow flexors of the right arm, making for a total of eight conditions. For the first condition the subject was blindfolded, so he or she could see neither arm. For the second, the left arm was visible throughout the matching process, and there was no mirror. For the third condition a dummy forearm was placed on the paddle normally occupied by the left arm. After conditioning of both arms, the left arm was placed at 50 deg and screened from view. The reference arm was then placed at the test angle and the subject directed the experimenter to move the dummy to a matching position. For the fourth condition a mirror was inserted between the two arms and the subject watched the mirror image of the left arm as it moved to the matching position. The different conditions were presented in random order and each condition, with and without vibration, was repeated five times, making for a total of 40 trials. In all trials the reference arm remained hidden from view.
Experiment 2
The speed of the movement illusion
The aim of this experiment was to see if movement of the indicator arm in any way disturbed the illusion of movement produced by vibration of the reference arm.
Because this experiment was a little more demanding, subjects underwent a series of initial familiarisation trials. One subject found the task too difficult and was excluded from further measurement. Muscle vibration elicits a sense both of forearm displacement and of movement (Goodwin et al. 1972). In this experiment we studied the illusion of movement generated in the reference arm by its vibration. Here, rather than tracking the illusion by moving the indicator arm, as is done traditionally, we asked subjects to respond to the felt movement by moving the vibrated arm itself, at the speed and in the direction of the illusion.
Both arms were strapped to their supporting paddles and moved to the conditioning angles, and conditioning contractions were carried out as in Experiment 1. The reference arm was always conditioned at 50 deg. The conditioning angle for the left, matching arm, was 50 deg if it was to subsequently be moved in the direction of extension and 10 deg if it was to be moved in the direction of flexion. The smaller conditioning angle before flexion movements allowed the arm to subsequently move through a larger range. Once both arms had been conditioned, the vibrator strapped to the reference arm, which was still sitting at 50 deg, was turned on. Once the illusion had established itself in the reference arm, subjects were asked to move the arm at the speed of the felt movement, in the felt direction. It meant that during the vibration they slowly extended their arm. The actual experiment was begun after the familiarisation trials had been completed and subjects were able to reproducibly track the movement illusion.
During this time the indicator arm was held at the conditioning angle by the experimenter. It was kept stationary until a sufficient segment of record of movement by the reference arm had been obtained. At that point the experimenter began to move the indicator arm in the direction of either flexion (7.1 ± 1.0 deg s−1) or extension (8.9 ± 1.0 deg s−1). Subjects were unable to see what was being done to their left arm and were instructed to continue to signal the illusion generated by vibration in their right arm whether or not the left arm was being moved. Trials with movements into flexion were intermingled in random order with trials for movements into extension and each trial was repeated at least five times.
The movement trace drawn by the vibrated arm was used to calculate arm extension velocity. This was measured with the left arm stationary (Control), with the left arm moving into extension (Extension), or with it moving into flexion (Flexion). Control velocity was measured from the point where the subject started to move his or her reference arm in response to vibration to when the experimenter started to move the indicator arm. Velocity was averaged over this period using Chart software. When the indicator was moved into extension or flexion the change in movement velocity of the reference arm was averaged over the period of movement of the indicator.
Repetitive movements
This experiment, carried out on nine subjects, examined whether continuous, forwards and backwards movements of the indicator arm altered the illusion of movement produced by vibration of the reference arm.
To do this, both arms were strapped to their supporting paddles and moved to the conditioning angles of 30 deg, and conditioning contractions were carried out as in Experiment 1. The reference arm was supported on a block. The vibrator was turned on and once the vibration illusion had manifested itself the indicator was moved up and down 3–4 times at approximately 10–20 deg s−1 over the range 50–10 deg. Subjects were asked to describe what they had felt during the indicator movements.
Statistics
Data were analysed using InStat (GraphPad Software Inc., La Jolla, CA, USA). In Experiment 1 position errors were calculated as angle (reference arm) – angle (indicator arm). Errors were compared between conditions in a one-way ANOVA with the Tukey–Kramer multiple comparisons post hoc test. In Experiment 2 the velocity of movement of the reference arm with the indicator stationary (Control) was compared with the velocity of movement when the indicator was moving into flexion (Flexion) or extension (Extension). Pooled values were compared by means of Student's paired t test. For all analyses, significance was recorded at P < 0.05. All values are given as means ± standard error of the mean (s.e.m.).
Results
Experiment 1
A total of 16 subjects carried out this experiment. One male subject was excluded from the final analysis because averaged angle differences obtained for two of the eight conditions (dummy and mirror) were outliers, defined as values ± 2 standard deviations beyond the group mean. The statistical analyses were therefore carried out on only 15 subjects.
Matching limb position without vibration
The aim of the experiment was to determine the effect of being able to see the indicator arm, a dummy arm, or a mirror image of the indicator arm on the ability to match the position of the other, unseen arm. The working hypothesis was that seeing the matching arm had no effect on matching ability. Data from a typical subject are shown in Fig. 2.
Figure 2. Forearm position matching by a single subject.
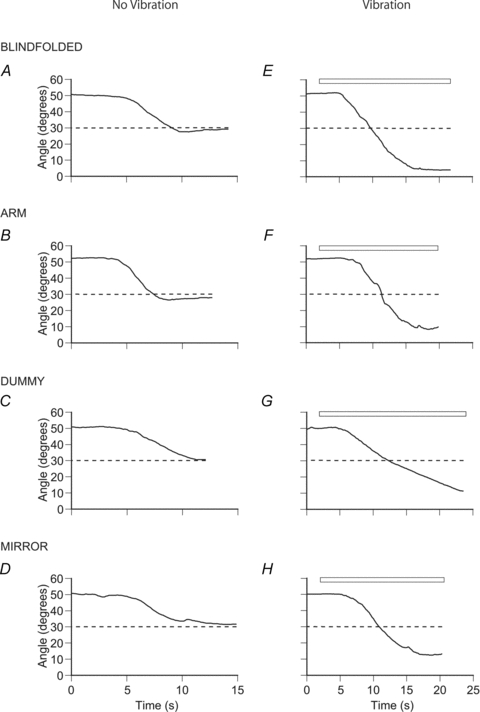
Each panel represents a single matching trial. The reference arm was placed at the test angle and several seconds later the subject was asked to match its position. During each trial the size of the elbow angle has been plotted against time over the 15–25 s duration of the trial. In each panel the dashed line indicates the position of the reference arm, the continuous line the position of the indicator arm. An angle of 0 deg represents the forearm in the horizontal position, an angle of 90 deg the forearm in the vertical position. In A the subject was blindfolded, in B they could see their indicator arm, in C the indicator arm was replaced with a dummy and in D the subject could see a mirror image of their indicator arm. Left-hand panels, matching trials without vibration; right-hand panels, matching trials during vibration of elbow flexors of the reference arm. The period of vibration is shown by the bar at the top of each panel.
The experiment began by asking the blindfolded subject to match the position of their reference arm, which had been placed at an angle of 30 deg to the horizontal after it had undergone a conditioning contraction at 50 deg. Subjects matched the position with reasonable accuracy and for this individual the mean error for five trials was 0.3 deg, that is the matching arm was placed slightly in the direction of extension (Fig. 2A, Fig. 3A). The matching task was then repeated but with the blindfold removed. Now the mean matching error changed to 1.2 deg, that is the indicator arm adopted a position 1.2 deg in the direction of extension relative to the position of the reference arm (Fig. 2B, Fig. 3A). For the condition where matching was done with a dummy arm strapped to the indicator paddle, the mean matching error was 1.3 deg in the direction of extension (Fig. 2C, Fig. 3A). Finally, the mirror was put in place and its position so arranged that the mirror image was coincident with the position of the reference arm. Again the subject was able to match forearm position reasonably accurately (Fig. 2D, Fig. 3A), with a mean matching error of −0.4 deg, that is the matching arm was placed slightly in the direction of flexion.
Figure 3. Position matching errors for a single subject and for the group.
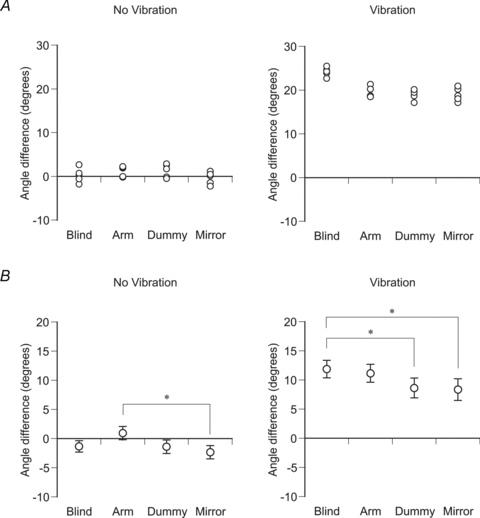
A, single subject; B, group data. Left-hand panels, matching without vibration, right-hand panels with vibration. Matching conditions were with the subject blindfolded (Blind), with their indicator arm visible (Arm), using a dummy arm (Dummy) and with a mirror image of the indicator arm visible (Mirror). Angle difference was calculated as the position of the reference arm minus the position of the indicator arm. 0 deg represented a perfect match. Positive errors were where the indicator arm had adopted a more extended position relative to the reference; negative errors were where the indicator had adopted a more flexed position. In A are shown the angle differences for each of five trials for a particular condition by the one subject. In B are shown the pooled data for the 15 subjects. Values are means (±s.e.m.). Asterisks indicate significant differences (P < 0.05).
The pooled data for the 15 subjects is shown in Fig. 3B (left-hand panel). The mean position error with subjects blindfolded was −1.3 ± 1.0 deg. With the indicator arm in view the error was 0.9 ± 1.1 deg. When the dummy was used, the error was −1.4 ± 1.2 deg. When the mirror was in place it was −2.4 ± 1.1 deg. Statistical analysis showed that position matching errors were significantly different between the four conditions (P < 0.05, one-way ANOVA). Tukey–Kramer multiple comparisons post hoc tests showed that there was a significant difference in matching errors only between the Arm and Mirror conditions (P < 0.05).
Matching limb position during vibration
Throughout the previous matching trials a vibrator had been strapped to biceps of the reference arm but not turned on. When this was turned on, subjects reported a sensation of the arm moving in the direction of extension and they perceived their arm to be more extended than it was. This is the well-known vibration illusion (Goodwin et al. 1972).
During vibration the blindfolded subject illustrated in Figs 2E and 3A believed their arms to be accurately aligned with the indicator arm placed a mean of 24.2 deg in the direction of extension relative to the position of the vibrated reference arm. When the subjects could see their indicator arm the mean error was less, 19.8 deg (Figs 2F and 3A). The error reduced further to 19.0 when the subject matched with a dummy arm (Figs 2G and 3A). When the subject was asked to use a mirror image of the indicator arm during matching the error remained at 19.0 deg (Figs 2H and 3A).
The pooled data showed a similar trend (Fig. 3B, right-hand panel). When subjects were blindfolded, mean position error during vibration was 11.9 ± 1.5 deg. With the indicator arm in view the error was 11.1 ± 1.5 deg. When the subjects used the dummy, the error decreased to 8.6 ± 1.7 deg. When subjects could see a mirror image of their indicator arm the error fell further to 8.4 ± 1.9 deg.
Statistical analysis showed that position matching errors were significantly different between the four conditions (P < 0.05, one-way ANOVA). Tukey–Kramer multiple comparisons post hoc tests showed that when subjects were blindfolded, errors during vibration were significantly larger than when they used the dummy arm (P < 0.05) and when they were looking at the mirror image (P < 0.05), but not when they could see the indicator arm.
Experiment 2
The speed of the movement illusion during vibration
This experiment was concerned with the question of whether moving the indicator arm altered subjects’ perception of motion of the vibrated reference arm. The experiment was attempted with 11 subjects. Of these, three male subjects developed strong TVRs that interfered with their ability to track the extension illusion. They were excluded from further study, as was one female subject who was unable to reproducibly track the perceived movement. That left data from seven subjects. At the start of the experiment the indicator arm was kept stationary. Vibration of the reference arm was turned on and as the vibration illusion developed over the next 2–3 s subjects began to track the apparent movement in the direction of extension. When a sufficient segment of tracking trace had been recorded the experimenter began to move the indicator arm.
Example traces for one subject are shown in Fig. 4. The top panel shows the tracking movement of the reference arm before and during extension movements of the indicator arm. For this particular subject average tracking speed by the reference arm over the period of 3 s where the indicator arm remained stationary was 5.6 deg s−1. In other words, the subject believed their vibrated reference arm was moving into extension at 5.6 deg s−1. If during this time the indicator was not held stationary but moved into extension at 12.1 deg s−1, perceived movement speed of the reference arm slowed to 3.3 deg s−1. In the lower panel of Fig. 4 are shown the tracking movements when the indicator arm was moved into flexion. Here the tracking speed by the reference arm before the indicator was moved was 7.0 deg s−1. During flexion of the indicator at 9.6 deg s−1 movement speed of the reference arm increased to 13.3 deg s−1.
Figure 4. Movement sensations in the vibrated reference arm during movement of the indicator arm.
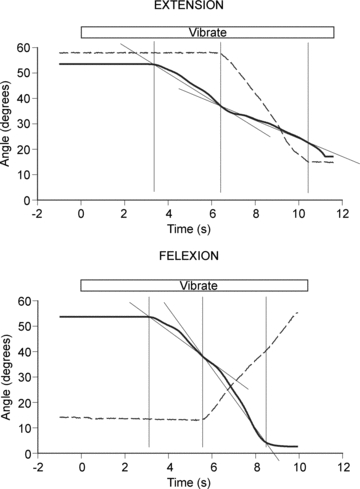
Upper panel, perceived movement of the reference arm with the indicator arm stationary and then moving in the direction of extension. Lower panel, perceived movement of the reference with the indicator stationary and with it moving into flexion. In each panel the dashed line represents the angle of the indicator arm, the continuous line the angle of the reference. The period of vibration is shown at the top. Each trial began when the vibrator on the reference arm was turned on. About 3 s later the illusion of forearm extension set in and the subject began to track it (first vertical line). Up to this point the indicator arm had been kept stationary. Once the subject was tracking successfully, the indicator was moved (second vertical line). Tracking was continued for several more seconds and then stopped (third vertical line). Mean slopes, calculated for movement of the reference arm, have been drawn in.
The pooled data are shown in Fig. 5. Over all trials the average speed of the extensions imposed on the indicator arm by the experimenter was 8.9 ± 1.0 deg s−1 while speed of the imposed flexions was 7.1 ± 1.0 deg s−1.
Figure 5. Pooled data for movement sensations.
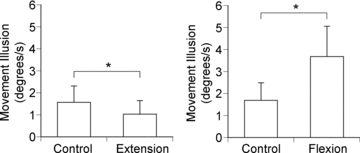
The pooled data from seven subjects are shown. Bars (means ±s.e.m.) show movement illusion speed (deg s−1) when the indicator arm was held stationary (Control), when it was moved in the direction of extension (Extension) and when it was moved in the direction of flexion (Flexion). Asterisks indicate significant differences (both P < 0.05).
In the trials involving indicator extensions (Fig. 5, left-hand panel), mean tracking speed by the vibrated reference arm was 1.6 ± 0.7 deg s−1 while the indicator arm remained stationary and it fell significantly to 1.0 ± 0.6 deg s−1 when the indicator arm was extended (P < 0.05, Student's paired t test). Therefore arm extension slowed the speed of the extension illusion. In the trials with indicator flexions (Fig. 5, right-hand panel), tracking speed with the indicator stationary was 1.7 ± 0.8 deg s−1 and it sped up significantly to 3.7 ± 1.4 deg s−1 during the movement (P < 0.05, Student's paired t test). That is, tracking speed of the reference arm could be sped up by indicator flexion.
Repetitive movements
In an additional experiment on nine male subjects they were asked what they felt when the indicator arm was continuously moved up and down 3–4 times over the range 50–10 deg at about 10–20 deg s−1. All subjects reported a ‘flutter’ sensation of slowing and speeding up of movement of the vibrated arm as the indicator changed its direction of movement. The sensation of slowing was reported to blend smoothly into a sensation of speeding up as the arm moved from extension back into flexion.
Discussion
Experiment 1
Position matching without vibration
The first question we asked was, does vision of the indicator arm, its mirror image, or the image of a dummy arm alter matching accuracy achieved in the blindfolded condition? Looking at Fig. 3B, in the absence of vibration the errors in blindfolded position matching were small, as were the errors during the other three conditions. However, it is worth discussing why some of the effects were not larger. Watching the indicator arm moving into the matching position had only a small effect on matching accuracy (Fig. 3B). In other words, visual confirmation of the proprioceptive signal coming from the indicator during the match did little to improve accuracy. Using a dummy arm again did not significantly alter the distribution of errors. It seems that the proprioceptive signal coming from the indicator does not necessarily play a dominant role since matching accuracy was reasonably well maintained when the matching signal was entirely visual. During this time the subject's left arm was held hidden from view at 50 deg, so its proprioceptive input was likely to be rather different from that coming from the reference arm, held at 30 deg. The discrepancy in proprioceptive signals between the two arms does not appear to have interfered with matching performance. Presumably here the visual information is simply prioritised. Finally, trying to trick the subjects with a mirror into believing that their indicator was really the reference had, not surprisingly, little effect on matching accuracy. If visual confirmation of indicator position had little effect, uncertainty about reliability of the visual signal was not likely to be important either. We conclude that in a two-arm position matching task subjects can use vision and proprioception interchangeably to achieve an accurate match.
Position matching during vibration
The distribution of position errors and the effects of the various conditions changed when matching was carried out during vibration of the reference arm. The vibration produced an illusion of the vibrated arm moving into extension and at the end of vibration the position adopted by the indicator arm during blindfolded matching lay 11.9 deg in the direction of extension beyond the reference arm (Fig. 3B). The overestimate of elbow angle was a little less (11.1 deg) when subjects could see their indicator arm. It was significantly less (8.6 deg) when subjects used a dummy arm to match position. Finally, providing false visual information by means of a mirror also significantly reduced the vibration error (8.4 deg). Nevertheless, despite the significant effects of the dummy and mirror, under all matching conditions errors continued to lie significantly in the direction of extension compared with the errors seen during matching without vibration.
The main effect of vibration on position matching was a strong illusory sense of elbow extension generated in the vibrated arm. None of the manipulations of the indicator arm were able to abolish the illusion and it remained at 70% or more of the value measured with blindfolded matching for all conditions. This suggests that in a task where one arm is asked to match the position of the other, the position signal coming from the reference arm is assigned particular importance. In other words, in an arm matching task, when we ask, ‘Where is that arm?’ we are able to consciously focus our attention on its position signals, at the expense of those coming from the matching arm.
The currently accepted interpretation of the vibration illusion is that the vibration-evoked afferent signal coming from the reference arm is much larger than the signal from the indicator arm, leading to perception of the vibrated muscle as being longer, the arm more extended, than its actual position. That is, the difference signal in proprioceptive input from the indicator and reference is used to express the size of the illusion (Goodwin et al. 1972). The sensation of arm extension implies that this is generated relative to some central reference point, presumably the perceived position of the arm before the vibrator was turned on.
When the subject is blindfolded and has only proprioceptive signals to compare between the two arms, the illusion is at its greatest (Fig. 3). If the subjects can see their indicator arm during matching, they become aware of the discrepancy between its position before and during vibration, leading them to signal a slightly smaller illusion. However when indicator matching is done by vision only, using a dummy arm, the vibration errors generated in the reference arm are significantly reduced. Now, in the absence of any proprioceptive signal from the indicator, the visual signal can exert its full effect. Such an explanation suggests that the visual location of the forearm can be interpreted in terms of an expected proprioceptive signal for that location. Perhaps, as a result of daily experience, we are able to interchangeably use visual and proprioceptive cues in determining limb position.
Somewhat unexpectedly, using the mirror reduced the vibration illusion as much as with a dummy. Perhaps uncertainty about the identity of the seen arm leads the subject to give a greater weighting to the visual signal. So, on the basis that the subject compares the proprioceptive signal from the reference arm with its apparent position signalled visually, it leads to a significant reduction of the vibration illusion.
Finally, it is worth reflecting on the fact that during normal position matching, manipulations of the indicator signals had little effect on matching accuracy, yet they manifested themselves clearly during the vibration illusion. Perhaps it is simply that such effects can only be shown when errors during blindfolded matching are large. Vibration is an unnatural stimulus and it is conceivable that, unconsciously, there is some uncertainty about reliability of this signal. As a result, inputs that throw doubt on the validity of the signal are able to exert their effect.
Throughout the matching process subjects were unaware that they were signalling a smaller vibration illusion when they could see their indicator arm or were looking at a dummy. It implies that the factors involved in position matching were operating at an unconscious level. In that context, it is interesting that muscle vibration at 70 Hz activates predominantly motor areas of the brain (Naito et al. 1999).
There is some psychophysical evidence of changes in vibration illusions by vision. The size of the illusion of wrist flexion from vibration of wrist extensors could be graded by subjects viewing hand movements. This effect was present only when the direction of the illusion was concordant with the direction of the observed movements (Hagura et al. 2009). Other effects of vision on vibration illusions have been reported (Lackner & Taublieb, 1984), as have disturbances of reaching movements when visual and proprioceptive signals are made incongruent by means of mirrors (Holmes & Spence, 2005). All of this is reminiscent of the rubber hand illusion, where the combination of visual and tactile signals is able to override the proprioceptive input in claiming ownership of the hand (Botvinick & Cohen, 1998). Presumably each channel providing input about a particular body part is given a certain weighting and this varies according to the circumstances and based on previous experience. In our experiment the sensory experience evoked by proprioceptive input from vibration is allowed to be attenuated but not abolished by conflicting visual input.
Experiment 2
The simple conclusion from this experiment was that the speed of the illusion of elbow extension evoked by vibration of the reference arm could be altered by movement of the indicator arm. Vibration is known to selectively excite the primary endings of muscle spindles (Roll et al. 1989). The illusion of movement of the arm during vibration is presumably, therefore, generated by the primary endings. The observation that movement of the indicator arm could alter the speed of the illusion suggests that the movement sensation is not derived exclusively from the vibration-induced signal coming from the reference arm, but represents a difference signal calculated from the input coming from both arms. When the indicator arm was moved into extension, this raised the spindle signal in its own elbow flexors and therefore the difference in spindle discharge rate between the two arms became less. That led to perception of a slower vibration illusion (Figs 4 and 5). Similarly, flexing the indicator lowered its flexor spindle signal as the muscles shortened and unloaded their spindles. As a result the difference signal between the two arms increased and that was expressed as an illusion of a faster movement. Interestingly, when the arm was flexed and extended continuously over 40 deg, subjects reported a fluttering sensation of slowing and speeding up of the apparent movement of the reference arm. So the quality of the sensation generated by vibration in one arm can be modified by movement of the other arm. However, just as for the position illusion generated by vibration, the movement illusion can be modified but not abolished by movements of the other arm. Nevertheless this experiment demonstrates unambiguously the existence of an interdependence between the two arms in the proprioceptive sensations to which their movements give rise.
Moving the vibrated arm into extension as it tracked the illusion may have led to changes in the mechanical conditions in elbow muscles and their spindles. The vibration may have been transmitted more effectively to spindles. However, with flexion conditioning of the arms, as used in these experiments, spindle sensitivity to vibration remains high even at short lengths (Proske & Gregory, 1999). It is therefore unlikely that the spindle response itself changed very much during the tracking movements. Certainly, provided the indicator arm remained stationary, constancy of the tracking speed reported by subjects supports that view (Fig. 4).
Another consideration is that position sense may be derived from the position of the limb relative to the gravitational vertical, rather than be based on joint angle information (Worringham et al. 1987). Since position relative to the vertical was changing as the vibrated arm moved into extension, it could have influenced the reported sensation of movement. Finally, since all subjects in this study used their right arm as the reference and all but one were right-handed, it raised the possibility of handedness as being a factor (Goble et al. 2006; Adamo & Martin, 2009). However, our own work on the influence of thixotropy on position sense and the vibration illusion has shown similar effects for both arms (White & Proske, 2009).
It is already known that movement illusions from vibration can be manipulated. Touching a stationary surface during vibration of elbow flexors reduces matching errors (Rabin & Gordon, 2006). Muscle's passive mechanical state can be altered as a result of its thixotropic property (Lakie & Robson, 1988; Proske et al. 1993) and this can lead to errors in an elbow position matching task (Winter et al. 2005). When the muscle is conditioned with a contraction to leave muscle fibres, including the intrafusal fibres of spindles, in a taut state, the speed of the vibration illusion is increased (Gooey et al. 2000; see also Burke & Gandevia, 1995). In a two-arm matching task in the horizontal plane, subjects no longer reliably showed the vibration illusion if the indicator arm was conditioned so that its muscle spindle discharge was likely to be low (White & Proske, 2009). If the vibrated arm was bearing a load, the illusion also disappeared (Goodwin et al. 1972; Ansems et al. 2006). Here we postulated that once the fusimotor supply of spindles had been recruited during motor co-activation (Vallbo, 1974) to support the load, the co-activated spindles no longer contributed to kinaesthesia. When the load was sufficiently heavy to require co-activation of most spindles in the muscle, the vibration illusion was gone.
To the best of our knowledge, this is the first account of manipulations of the movement illusion generated by vibration in one arm by movement of the other arm. In related experiments, Sittig et al. (1987) used movements in the horizontal plane and asked subjects to indicate the magnitude of movement sensations evoked by vibration at different frequencies. Although they did not specifically study the point, they suggested that moving the vibrated arm was likely to increase the size of the illusion. Cordo et al. (1995) asked subjects to indicate the speed of movement of the right arm during its vibration at 40 Hz by moving their left arm. Subjects reported that they felt their arm moving into extension and moving more rapidly than it actually was. When vibration frequency was reduced to 20 Hz there was no effect on dynamic position sense, but a reduction in the perceived velocity of the arm. In a subsequent study, Cordo et al. (2005) reported that the speed of the vibration illusion could be altered by small, slow movements of the vibrated arm. Movements in the same direction as the illusion enhanced it, while movements in the opposite direction reversed it. Interestingly, these effects are the reverse of what was seen in the present study, but are consistent with predicted changes in the mechanical state of the vibrated muscle. Cordo et al. (2005) concluded that their effects might be due to muscle afferents other than spindle primary endings, in addition to other non-muscle, stretch sensitive receptors.
In their original experiments Goodwin et al. (1972) asked subjects to track with their non-vibrated arm the illusion generated by the vibration in the other arm. Our observations suggest that the tracking movement may have acted to reduce the speed of the felt illusion. Similarly, the observations of Cordo et al. (2005) suggest that in our experiments the tracking movements initiated by the vibrated reference arm may have exaggerated the perceived illusion of arm extension.
Broader implications
This study was prompted by the question, why did one-arm and two-arm measurements of position sense lead to different conclusions about the possible contribution of a motor command signal to human position sense? One explanation might be that in a one-arm experiment, afferent input is available only from that arm and a comparison with the other arm is not possible. Because of the limited proprioceptive information available, the brain resorts to accessing motor command signals perceived as a sense of effort and uses these to supplement the peripheral afferent input. In a two-arm matching task, input from both arms is available and it is not necessary to make use of motor command signals. In the matching task, when one arm is loaded, the position signal coming from it may be weaker since its spindles are co-activated during support of the load (see above). Therefore positional information coming from the unloaded arm may be prioritised and any motor command component associated with the loaded arm may not be detectable. The results reported in the present study suggest another explanation. It may be that what appears at first sight to be a one-arm proprioceptive test may in fact include some input from the other arm. Exactly how this might influence the outcome of an experiment on motor command signals and proprioception remains unclear.
However, the implications of the observations in this study extend beyond methodological considerations. In determining arm position in space, in the absence of vision, the brain probably refers to a kinaesthetic map, the body schema (Berlucchi & Aglioti, 2010). The map is likely to be distorted by the numbers of afferents coming from different limb segments and the assignment of particular importance to patterns of input associated with everyday postures. For the arms, the arms-in-front, hands-aligned position is likely to be given such importance (Gooey et al. 2000). In that posture, if subjects are asked to match their arms under visual control they are extremely accurate, with errors typically less than 1 deg. When subjects are blindfolded during matching, accuracy falls a little and typically errors of about 2 deg are observed (Allen et al. 2007). Presumably such levels of accuracy are necessary for normal fine manipulations with the hands.
When we carry out skilled movements with our hands, we rely on tactile, proprioceptive and visual information to place our arms. Indeed, they are the principal modalities determining the body schema (Berlucchi & Aglioti, 2010). Experiments like those of the rubber hand illusion (Botvinick & Cohen, 1998) indicate that the relative importance of each input signal depends on the context within which it is generated. In the absence of vision, proprioception is all important. When we move our arms while blindfolded we are constantly aware of the identity of each arm and its location, the result of independent processing of the sensory information from each limb. The present study has shown that in a position matching or movement tracking task during vibration of elbow flexors of one arm, the sense both of movement and of displaced position of the vibrated arm is influenced to some extent by the information coming from the other arm. This implies a degree of interdependence in the central evaluation of the incoming sensory information from the two arms. For bilateral manual coordination it will be important to preserve the identity of each arm and this will require interhemispheric interactions to allow one hand to know what the other is doing (Koch et al. 2009). A broader implication of the proprioceptive interdependence between the arms shown in the present study is that in some circumstances, in certain respects, the two arms can be considered to be acting as one, a single instrument, in the execution of skilled movements. Yet, at the same time, both retain their unique individual identities.
To conclude, the present studies have demonstrated in an arm matching task the important role played by afferent input coming from both arms. This should be kept in mind when comparing kinaesthetic data from one-arm and two-arm experiments. More generally, the demonstrated interdependence of proprioceptive sensations arising from the two arms hints at a previously unrecognised level of common processing of sensory information coming from the two arms.
Acknowledgments
Financial support from Showa University for U.P. during his stay in Japan is gratefully acknowledged.
Glossary
Abbreviations
- TVR
tonic vibration reflex
Author contributions
M.I., U.P. and I.H. participated in the conception and design of the experiments; all authors performed the experiments; M.I. and U.P. performed data analysis; all authors interpreted the data, contributed to the drafting and revision of the manuscript and approved the final version of the manuscript.
References
- Adamo DE, Martin BJ. Position sense asymmetry. Exp Brain Res. 2009;192:87–95. doi: 10.1007/s00221-008-1560-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen TJ, Ansems GE, Proske U. Effects of muscle conditioning on position sense at the human forearm during loading or fatigue of elbow flexors and the role of the sense of effort. J Physiol. 2007;580:423–434. doi: 10.1113/jphysiol.2006.125161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ansems GE, Allen TJ, Proske U. Position sense at the human forearm in the horizontal plane during loading and vibration of elbow muscles. J Physiol. 2006;576:445–455. doi: 10.1113/jphysiol.2006.115097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berlucchi G, Aglioti S. The body in the brain revisited. Exp Brain Res. 2010;200:25–35. doi: 10.1007/s00221-009-1970-7. [DOI] [PubMed] [Google Scholar]
- Botvinick M, Cohen J. Rubber hands ‘feel’ touch that eyes see. Nature. 1998;391:756. doi: 10.1038/35784. [DOI] [PubMed] [Google Scholar]
- Burke D, Gandevia SC. The human muscle spindle and its fusimotor control. In: Ferrell W, Proske U, editors. Neural Control of Movement. New York: Plenum Press; 1995. pp. 19–25. [Google Scholar]
- Cordo PJ, Gurfinkel VS, Bevan L, Kerr GK. Proprioceptive consequences of tendon vibration during movement. J Neurophysiol. 1995;74:1675–1688. doi: 10.1152/jn.1995.74.4.1675. [DOI] [PubMed] [Google Scholar]
- Cordo PJ, Gurfinkel VS, Brumagne S, Flores-Vieira C. Effect of slow, small movement on the vibration-evoked kinesthetic illusion. Exp Brain Res. 2005;167:324–334. doi: 10.1007/s00221-005-0034-x. [DOI] [PubMed] [Google Scholar]
- Gandevia SC, Smith JL, Crawford M, Proske U, Taylor JL. Motor commands contribute to human position sense. J Physiol. 2006;571:703–710. doi: 10.1113/jphysiol.2005.103093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goble DJ, Lewis CA, Brown SH. Upper limb asymmetries in the utilisation of proprioceptive feedback. Exp Brain Res. 2006;168:307–311. doi: 10.1007/s00221-005-0280-y. [DOI] [PubMed] [Google Scholar]
- Goodwin GM, McCloskey DI, Matthews PB. The contribution of muscle afferents to kinaesthesia shown by vibration induced illusions of movement and by the effects of paralysing joint afferents. Brain. 1972;95:705–748. doi: 10.1093/brain/95.4.705. [DOI] [PubMed] [Google Scholar]
- Gooey K, Bradfield O, Talbot J, Morgan DL, Proske U. Effects of body orientation, load and vibration on sensing position and movement at the human elbow joint. Exp Brain Res. 2000;133:340–348. doi: 10.1007/s002210000380. [DOI] [PubMed] [Google Scholar]
- Gregory JE, Morgan DL, Proske U. Aftereffects in the responses of cat muscle spindles and errors of limb position sense in man. J Neurophysiol. 1988;59:1220–1230. doi: 10.1152/jn.1988.59.4.1220. [DOI] [PubMed] [Google Scholar]
- Hagura N, Oouchida Y, Aramaki Y, Okada T, Matsumura M, Sadato N, Naito E. Visuokinesthetic perception of hand movement is mediated by cerebro-cerebellar interaction between the left cerebellum and right parietal cortex. Cerebral Cortex. 2009;19:176–186. doi: 10.1093/cercor/bhn068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes NP, Spence C. Visual bias of unseen hand position with a mirror: spatial and temporal factors. Exp Brain Res. 2005;166:489–497. doi: 10.1007/s00221-005-2389-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch G, Ruge D, Cheeran B, Fernandez Del Olmo M, Pecchioli C, Marconi B, Versace V, Lo Gerfo E, Torriero S, Oliveri M, Caltagirone C, Rothwell JC. TMS activation of interhemispheric pathways between the posterior parietal cortex and the contralateral motor cortex. J Physiol. 2009;587:4281–4292. doi: 10.1113/jphysiol.2009.174086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lackner JR, Taublieb AB. Influence of vision on vibration-induced illusions of limb movement. Exp Neurol. 1984;85:97–106. doi: 10.1016/0014-4886(84)90164-x. [DOI] [PubMed] [Google Scholar]
- Lakie M, Robson LG. Thixotropy: the effect of stretch size in relaxed frog muscle. Q J Exp Physiol. 1988;73:127–129. doi: 10.1113/expphysiol.1988.sp003110. [DOI] [PubMed] [Google Scholar]
- McCloskey DI. Differences between the senses of movement and position shown by the effects of loading and vibration of muscles in man. Brain Res. 1973;63:119–131. doi: 10.1016/0006-8993(73)90521-0. [DOI] [PubMed] [Google Scholar]
- Naito E, Ehrsson HH, Geyer S, Zilles K, Roland PE. Illusory arm movements activate cortical motor areas: a positron emission tomography study. J Neurosci. 1999;19:6134–6144. doi: 10.1523/JNEUROSCI.19-14-06134.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Proske U. Kinesthesia: the role of muscle receptors. Muscle Nerve. 2006;34:545–558. doi: 10.1002/mus.20627. [DOI] [PubMed] [Google Scholar]
- Proske U, Gregory JE. Vibration sensitivity of cat muscle spindles at short muscle lengths. Exp Brain Res. 1999;124:166–172. doi: 10.1007/s002210050611. [DOI] [PubMed] [Google Scholar]
- Proske U, Morgan DL, Gregory JE. Thixotropy in skeletal muscle and in muscle spindles: a review. Prog Neurobiol. 1993;41:705–721. doi: 10.1016/0301-0082(93)90032-n. [DOI] [PubMed] [Google Scholar]
- Proske U, Gandevia SC. The kinaesthetic senses. J Physiol. 2009;587:4139–4146. doi: 10.1113/jphysiol.2009.175372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rabin E, Gordon AM. Prior experience and current goals affect muscle-spindle and tactile integration. Exp Brain Res. 2006;169:407–416. doi: 10.1007/s00221-005-0154-3. [DOI] [PubMed] [Google Scholar]
- Roll JP, Vedel JP, Ribot E. Alteration of proprioceptive messages induced by tendon vibration in man: a microneurographic study. Exp Brain Res. 1989;76:213–222. doi: 10.1007/BF00253639. [DOI] [PubMed] [Google Scholar]
- Sittig AC, Denier Van DerGon JJ, Gielen CCAM. The contribution of afferent information on position and velocity to the control of slow and fast human forearm movements. Exp Brain Res. 1987;67:33–40. doi: 10.1007/BF00269450. [DOI] [PubMed] [Google Scholar]
- Smith JL, Crawford M, Proske U, Taylor JL, Gandevia SC. Signals of motor command bias joint position sense in the presence of feedback from proprioceptors. J Appl Physiol. 2009;106:950–958. doi: 10.1152/japplphysiol.91365.2008. [DOI] [PubMed] [Google Scholar]
- Vallbo AB. Human muscle spindle discharge during isometric voluntary contractions. Amplitude relations between spindle frequency and torque. Acta Physiol Scand. 1974;90:319–336. doi: 10.1111/j.1748-1716.1974.tb05594.x. [DOI] [PubMed] [Google Scholar]
- White O, Proske U. Illusions of forearm displacement during vibration of elbow muscles in humans. Exp Brain Res. 2009;192:113–120. doi: 10.1007/s00221-008-1561-z. [DOI] [PubMed] [Google Scholar]
- Winter JA, Allen TJ, Proske U. Muscle spindle signals combine with the sense of effort to indicate limb position. J Physiol. 2005;568:1035–1046. doi: 10.1113/jphysiol.2005.092619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Worringham CJ, Stelmach GE, Martin ZE. Limb segment inclination sense in proprioception. Exp Brain Res. 1987;66:653–658. doi: 10.1007/BF00270697. [DOI] [PubMed] [Google Scholar]