


Abstract
We constructed a dual regulated expression vector cassette (pDuoRex) whereby two heterologous genes can be independently regulated via streptogramin- and tetracycline-responsive promoters. Two different constructs containing growth-promoting and growth-inhibiting genes were stably transfected in recombinant Chinese hamster ovary (CHO) cells that express the streptogramin- and tetracycline-dependent transactivators in a dicistronic configuration. An optimally balanced heterologous growth control scenario was achieved by reciprocal expression of the growth-inhibiting human cyclin-dependent kinase inhibitor p27Kip1 in sense (p27Kip1S) and antisense (p27Kip1AS) orientation. Exclusive expression of p27Kip1S resulted in complete G1-phase-specific growth arrest, while expression of only p27Kip1AS showed significantly increased proliferation compared to control cultures (both antibiotics present), presumably by decreasing host cell p27Kip1 expression. In a second system, a derivative of pDuoRex encoding streptogramin-responsive expression of the growth-promoting SV40 small T antigen (sT) and tetracycline-regulated expression of p27Kip1 was stably transfected into CHO cells. Expression of sT alone resulted in an increase in cell proliferation, but the expression of p27Kip1 failed to provide the expected G1-specific growth arrest despite having demonstrated expression of the protein. This illustrates the difficulty in balancing the complex pathways underlying cell proliferation control through the expression of two functionally distinct genes involved in those pathways, and how a single-gene sense/antisense approach using pDuoRex can overcome this barrier to complete metabolic engineering control.
INTRODUCTION
Future success of cell and tissue engineering for ex vivo expansion will be based on technology managing temporal proliferation control of mammalian cells through well-balanced expression of growth-promoting and growth-suppressing genetic determinants (1). Independent expression control of two different transgenes requires two compatible regulation systems. We have recently adapted the pristinamycin resistance operon of Streptomyces pristinaespiralis for use as a mammalian gene regulation system (PipOFF system) (2). The PipOFF system is responsive to a class of clinically licensed antibiotics, the streptogramins (Pyostacin®, Virginiamycin, Synercid®), and has been shown to be compatible with the widely used tetracycline-repressible expression technology (TetOFF system) (2,3). The combination of the PipOFF and TetOFF systems in a two-vector configuration would enable dual regulation required for proliferation management of mammalian cells and other complex molecular interventions.
Within all mammalian cells, regardless of type and origin, positive and negative proliferation control is an exquisitely balanced mechanism which integrates signals from both external (growth factors, mitogens, cell–cell contact) and internal sources (differentiation, DNA repair, apoptosis) and ultimately generates a molecular signal that impinges on the cell cycle (4). The cyclin-dependent kinase inhibitor p27Kip1 is crucial for balanced regulation between growth and quiescence in many cell types by binding and inhibiting cyclin D-CDK4/6 and cyclin E/CDK2 complexes responsible for G1–S transition (5,6). p27Kip1 expression is induced in contact-inhibited or mitogen-deprived cells which exit from the cell cycle and enter a quiescent state upon accumulation of this cyclin-dependent kinase inhibitor (CKI) (5,7,8). Tetracycline-responsive overexpression of p27Kip1 has been shown to arrest CHO cells in the G1-phase of the cell cycle (9). Furthermore, downregulation of p27Kip1 expression is required for S-phase entry of some cell types, and growth factor-mediated activation of the Ras-Raf1-MEK1-ERK1/2 MAP kinase cascade decreases intracellular p27Kip1 levels (10–12). This virtually ubiquitous p27Kip1-based regulation system is even present during the management of host cell proliferation by an infecting virus. For example, the small T antigen of SV40 (sT) triggers a decline in p27Kip1 levels that enables cell-cycle re-entry of non-dividing target cells (13). Based on these observations in nature, we constructed two different vector-based dual regulation systems: (i) sense and antisense p27Kip1, whereby expression of sense p27Kip1 shuts down host cell proliferation and expression of antisense p27Kip1 alone inhibits endogenous p27Kip1 expression, leading to increased cell proliferation; and (ii) p27Kip1 and sT, whereby expression of p27Kip1 would inhibit host cell proliferation and expression of sT would directly increase proliferation. We tested these two dual regulation systems in CHO cells by stably transfecting the vectors and monitoring growth and cell-cycle profiles of cells following the removal of streptogramin or tetracycline from the cultures. With the former system, it was possible to provide complete proliferation management (growth arrest/growth induction) via mutually exclusive expression of sense or antisense p27Kip1 simply by removing streptogramin or tetracycline from the cultures.
MATERIALS AND METHODS
Cell culture, plasmids, transfection, regulating antibiotics, immunofluorescence and SEAP activity assay
Chinese hamster ovary cells (CHO-K1, ATCC: CCL61) and their derivatives were cultivated in FMX-8 medium (Dr F.Messi, Cell Culture Systems, Switzerland) supplemented with 10% fetal calf serum (FCS; PAA Vienna, Austria) unless stated otherwise. The cell line CHO-TWIN1108 (CHO-K1 stably transfected with pTWIN1) (2) was cultured in medium that also contained 400 µg/ml of G418 (Gibco BRL, Life Technologies). For stable transfection of CHO-TWIN1108 with pDuoRex plasmids, the desired dual regulation plasmids were cotransfected at a ratio of 1:15 with pZeoSV2 (Invitrogen) which confers resistance to 100 µg/ml zeocin. For both transient and stable transfections, 6 µg of DNA was used to transfect 400 000 cells at a transfection efficiency of 35 ± 5% (determined by parallel β-Gal-based assays) using an optimized CaPO4 protocol (9). Mixed stable populations were grown in the presence of pristinamycin (2 µg/ml) and tetracycline (2 µg/ml) to repress the dual regulated expression unit and subsequently cloned using FACS-mediated single cell sorting (FACStarPlus; Beckton Dickinson). Growth characteristics of CHO-233 and CHO-226 cell lines were assessed by seeding 100 000 cells in T25 flasks containing FMX-8 medium supplemented with 10% FCS and the appropriate regulating antibiotics (2 µg/ml) (Figs 2 and 4). The cell number was determined daily as an average taken from three independent cultures using a Casy1® cell counter according to the manufacturer’s protocol (Schärfe System, Germany).
Figure 2.
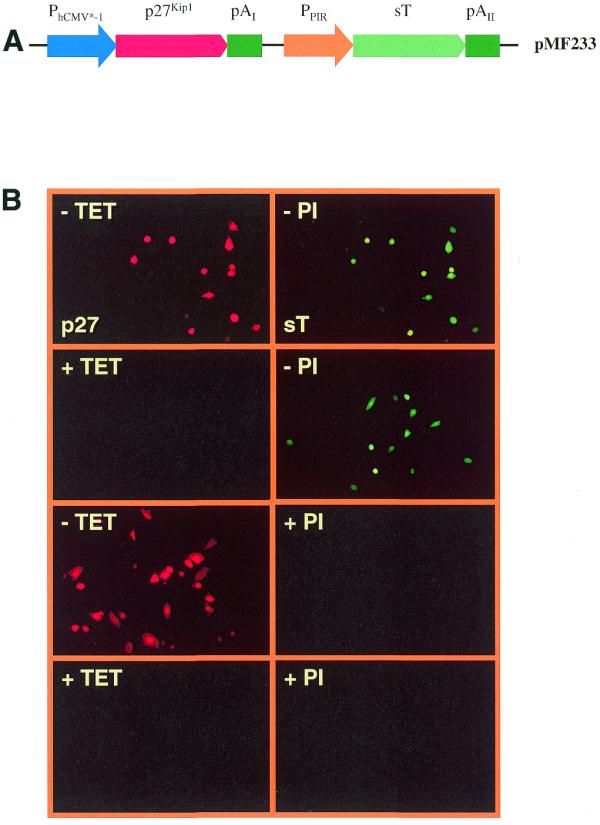
(A) Diagram of the dual regulated expression vector pMF233 encoding the human p27Kip1 cDNA under the control of the tetracycline-dependent promoter (PhCMV*-1) and sT under the control of the pristinamycin-responsive promoter (PPIR). (B) Immunofluorescence specific for p27Kip1 and sT proteins shows independent regulation of p27Kip1 and sT expression in CHO-TWIN1108 (constitutively expressing the pristinamycin- and tetracycline-dependent transactivators) cells transiently transfected with pMF233 in response to addition or withdrawal of tetracycline (+/– TET) or pristinamycin (+/– PI).
Figure 4.

FACS-mediated cell-cycle analysis of CHO-233176 and CHO-22696. Both cell lines were grown for 48 h under conditions allowing for all three expression configurations +Tet/–PI (growth-promoting gene induced, sT for CHO-233176 and p27Kip1AS for CHO-22696), +Tet/+PI (both transgenes repressed) and –Tet/+PI [growth-arresting gene (p27Kip1) induced] and incubated with the DNA-staining chemical propidium iodide for DNA content-based assessment of cell-cycle distribution. The cell-cycle profiles of proliferation-enhanced (+Tet/–PI) conditions and set-ups which repress the dual regulation unit (+Tet/+PI) are compared to the cell-cycle distribution of the parental cell line CHO-TWIN1108 (G1, 29.5%; G2, 10.3%; S, 60.2%). In contrast to CHO-22696 cells which arrest in the G1-phase under –Tet/+PI conditions, CHO-233176 cells do not display any accumulation of this cell-cycle phase (see also Figs 3 and 5).
Plasmid pW2(t+T+) encoding the SV40 small and large T antigens was kindly provided by K.Rundell (13). pMF99 encodes the human p27Kip1 cDNA (14). Vector pTWIN1, encoding the pristinamycin (PIT)- and tetracycline (tTA)-dependent transactivators (PSV40-PIT-IRES-tTA-pA), the dual regulation vector pDuoRex1 (PhCMV*-1-MCSI-IRES-MCSII-pAI-/-PPIR-MCSIII-pAII), the multicistronics expression vector pTRIDENT1 and pTBC-1 have been described previously (2,9). pECFP-C1 and pEYFP-C1 were purchased from Clontech, and pcDNA3.1/V5/His-TOPO was purchased from Invitrogen.
Tetracycline (Sigma) and pristinamycin (Pyostacin®; Aventis Inc.) were used as regulating antibiotics at concentrations of 2 µg/ml. Immunofluorescence was performed as described previously (9) using antibodies specific for p27Kip1 (M-197 or F-8; Santa Cruz) and sT (Pab108; Santa Cruz) along with FITC- or Texas Red-labeled secondary antibodies (Jackson Immuno Research). Immunofluorescence was visualized using a Leica DM-RB fluorescence microscope equipped with appropriate filters. The fluorescence proteins enhanced cyan fluorescent protein (ECFP) and enhanced yellow fluorescent protein (EYFP) were visualized with the same microscope but using different filters XF104 (ECFP) and XF114 (EYFP) (Omega Optical Inc.).
FACS analysis
For FACS-mediated cell-cycle analysis, cells were washed twice with PBS prior to detachment using dissociation solution (Sigma) and centrifugation at 1500 g. The supernatant was removed and the cells were resuspended in PBSAz (0.1% NaAz and 2% FCS in PBS) and centrifuged again. After a second washing step, 106 cells were dissolved in 750 µl of ice-cold PBSAz. While vortexing, 250 µl of 1% PFA in PBS was added and the mixture was incubated at 4°C for 1 h. Following centrifugation the cells were dissolved in 1 ml of 0.2% Tween-20 in PBS and incubated for 15 min at 37°C. Cells were then washed three times with PBSAz before incubation in 500 µl propidium iodide staining medium (10 mg/l propidium iodide and 1 mg/l RNAse A in PBSAz) for 30 min at 37°C in the dark. Analysis was performed on an EPICS ELITE apparatus (Beckmann-Coulter) using the MultiCycle software (Phoenix Flow Systems, Inc.).
Western blot analysis
For preparation of whole cell extracts, cells were washed with ice-cold PBS and scraped in ice-cold NP-40 extraction buffer (50 mM HEPES pH 7.4, 150 mM NaCl, 25 mM β-glycerophosphate, 25 mM NaF, 5 mM EGTA, 1 mM EDTA, 1% NP-40, 10 µg/ml leupeptin, 10 µg/ml aprotinin, 1 mM PMSF, 1 mM sodium orthovanadate). Suspensions were incubated 5 min on ice and the lysates were centrifuged at 23 000 g for 15 min at 4°C. Protein concentrations were determined using a Bradford assay (Bio-Rad). Whole cell extracts were denatured by boiling in sample buffer and subsequent SDS–polyacrylamide gel electrophoresis (15% PAGE). The proteins were blotted onto polyvinylidene difluoride (PVDF) membranes (Millipore). After blocking with 5% non-fat dry milk (Bio-Rad) in TTBS (50 mM Tris pH 7.5, 150 mM NaCl, 0.05% Tween-20), filters were probed with specific antibodies (p27Kip1, F-8; sT, Pab108). Proteins were visualized with HRP-conjugated donkey anti-mouse secondary antibodies using the ECL detection system (Amersham).
Vector construction
pDuoRex3 (pMF196) contains converging pristinamycin- and tetracycline-responsive expression units and was constructed following a multiple step cloning procedure: (i) the gene encoding ECFP was amplified from pECFP-C1 (Clontech) using oligonucleotides OMF96: GATCGAATTCcctcagcaccaggtCATGCTTAAGTCGCGACATATGgatccgctagcgctaccg and OMF89: GATCAAGCTT/GCCCGGGCCACACAAAAAACCAACACACAGATGTAATGAAAATAAAGATATTTTATTTGATCAGGCGCGCCGCGGCCGCATGCttacttgtacagctcgtc, and cloned into pcDNA3.1/V5/His-TOPO (Invitrogen) under the control of PCMV to yield plasmid pSAM228. (ii) The gene encoding the EYFP was amplified from pEYFP-C1 (Clontech) with oligos OMF90: GTACGAATTCGATATCATGCATGGCGCCGTTTAAACGCGTATTTAAATgatccgctagcgctaccg and OMF91: GATCAAGCTTGCGGCCGCGGATCCGCCCGGGCCACACAAAAAACCAACACACAGATGTAATGAAAATAAAGATATTTTATTATCGATACTAGTGCGATCGCTTAATTAATTTAAATttacttgtacagctcgtcc, and cloned into pcDNA3.1/V5/His-TOPO under the control of PCMV to result in plasmid pSAM222. OMF89, OMF90, OMF91 and OMF96 contain a variety of restriction sites which flank ECFP and EYFP and enable straightforward replacement of these fluorescence determinants by other genes of interest. OMF89 and OMF90 contain artificial polyadenylation sites (pA*) in their 3′ extensions (underlined). (iii) The EYFP-pA* containing cassette was excised from pSAM222 by EcoRI/NotI (contained in the oligos; bold) and ligated to the corresponding sites of pMF164 (NotI/EcoRI) thereby replacing GFP of pMF164 and resulting in plasmid pSAM226. (iv) The PPIR-EYFP-pA* cassette was excised with SspI/NotI from pSAM226 and ligated into the corresponding sites of pTRIDENT1 (pMF125) (9), hence replacing PhCMV*-1 and IRES I of pTRIDENT1 to give plasmid pMF190. (v) The ECFP-pA* cassette was excised from pSAM228 by EcoRI and HindIII (contained in the oligos; bold) ligated into the corresponding sites (EcoRI/HindIII) of pTBC-1, resulting in plasmid pSAM227. (vi) The ECFP-pA* containing cassette was released from pSAM227 by digestion with XhoI (contained 5′ of PhCMV*-1 in pTBC-1) and SrfI (contained in OMF89; bold), and was subsequently ligated to pMF190 restricted with SalI (compatible to XhoI) and SrfI to result in plasmid pDuoRex3 (pMF196: PPIR-YFP-pAI-/-pAII-CFP-PhCMV*-1).
For the construction of p27Kip1 sense–antisense expression vector pMF226, the ECFP-EYFP cassette was excised from pMF196 (pDuoRex3) by EcoRI/HindIII and replaced by the EcoRI/HindIII fragment of pMF99 (14) which encodes the cyclin-dependent kinase inhibitor. pMF226 contains a PPIR→p27Kip1←PhCMV*-1 sense–antisense expression unit.
pMF233 was constructed by a two-step cloning procedure: (i) sT was amplified from pW2(t+T+) (13) using oligos OMF104: GATCGATATCACTAGTgcctaggcttttgcaaaaagc and OMF106: GATCAGATCTACGCGTttagagctttaaatctctg and ligated in sense orientation into pcDNA3.1/V5/His-TOPO (Invitrogen) to result in pMF231. (ii) sT was excised from pMF231 with SpeI/BglII and ligated into the corresponding sites of pMF194 (SpeI/BglII) resulting in pMF233 (PhCMV*-1-p27Kip1-pAI-/-PPIR-sT-pAII).
RESULTS
Construction of a dual regulated expression unit
Previous results have shown that the streptogramin- and tetracycline-responsive gene expression systems are compatible and can be used for independent control of two different gene activities (2). Gene therapy and tissue engineering applications require a compact design of expression vectors. The antibiotic-responsive dual regulated expression system consists of PIT and tTA and their responsive promoters PPIR (streptogramin-responsive promoter) and PhCMV*-1 (tetracycline-responsive promoter) (2,3). PIT and tTA were previously cloned in a dicistronic constitutive expression configuration (pTWIN1; PSV40-PIT-IRES-tTA; IRES, picornaviral internal ribosomal entry site) then stably expressed in a CHO cell line, resulting in CHO-TWIN1108 (2). The cloning of the streptogramin- and tetracycline-responsive expression units was more challenging because careful consideration of orientation of the two genes with respect to each other was required. Three orientations were examined: (i) divergent (←/→), (ii) convergent (→/←) and (iii) consecutive (→/→). Divergent configurations showed undesired co-regulation of PPIR- and PhCMV*-1-driven expression units (as exemplified by bidirectional tetracycline-responsive promoters; data not shown) (15). When PPIR-YFP (yellow fluorescent protein) and PhCMV*-1-CFP (cyan fluorescent protein) were oriented in a convergent orientation (pDuoRex3, PPIR-YFP-pAI-/-pAII-CFP-PhCMV*-1) only YFPOFF/CFPOFF, YFPON/CFPOFF and YFPOFF/CFPON expression scenarios were achieved (Fig. 1). The YFPON/CFPON expression configuration was not possible perhaps due to mutual transcriptional silencing (Fig. 1). Only pDuoRex1 derivatives harboring both expression units in consecutive orientation (PhCMV*-1-p27Kip1-pAI-/-PPIR-sT-pAII) enabled all four regulation scenarios (Fig. 2; see below).
Figure 1.
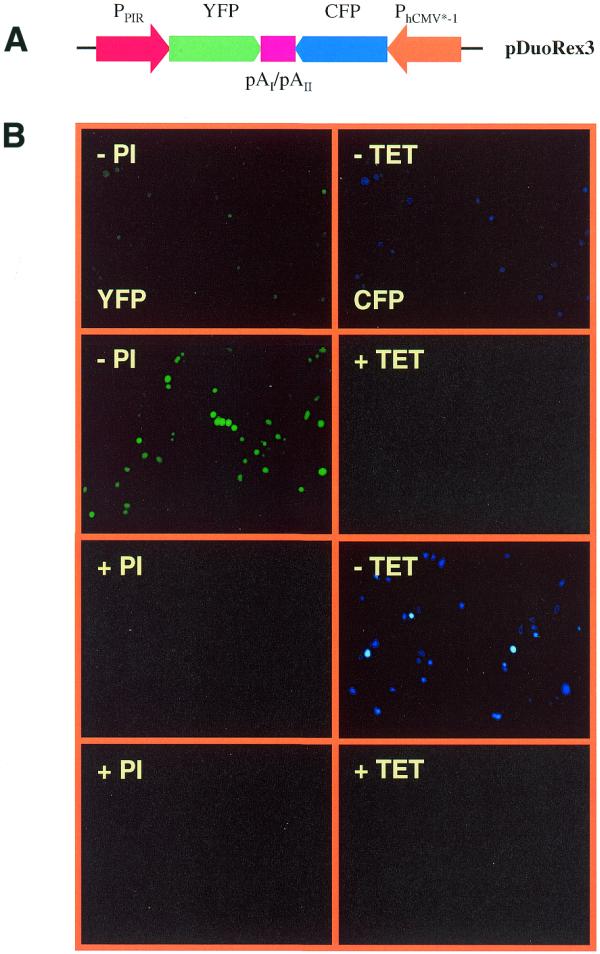
(A) Diagram of the dual regulated expression vector pDuoRex3 containing convergently oriented PPIR-YFP-pAI and PhCMV*-1-CFP-pAII expression units (PPIR-YFP-pAI-/-pAII-CFP-PhCMV*-1). (B) pDuoRex3 was transfected into CHO-TWIN1108, which stably harbors both tTA and PIT-dependent transactivators, and expression of YFP and CFP was assessed 48 h following transfection by fluorescence microscopy in the presence (+) or absence (–) of either tetracycline (TET; 2 µg/ml) or pristinamycin I (PI; 2 µg/ml). In the absence of both antibiotics (–TET/–PI) simultaneous induction of both responsive promoters results in reciprocal transcriptional silencing of the convergently oriented expression units.
Opposite control of p27Kip1 and sT in CHO cells
The human cyclin-dependent kinase inhibitor p27Kip1 and sT are theoretically an ideal combination for opposing proliferation control since overexpression of p27Kip1 has been shown to arrest CHO cells in the G1-phase of the cell cycle (9), and sT exerts positive proliferation control by decreasing p27Kip1 levels (13). We constructed a pDuoRex1 derivative pMF233 which contains a PhCMV*-1-driven p27Kip1 expression unit and a PPIR-driven sT in consecutive orientation (Fig. 2A; PhCMV*-1-p27Kip1-pAI-/-PPIR-sT-pAII). Transient transfection of pMF233 into CHO-TWIN1108 [stably expressing the PSV40-PIT-IRES-tTA expression unit (2)] confirmed independent expression of p27Kip1 and sT in all four expression configurations: p27Kip1ON/sTON (–Tet/–PI); p27Kip1OFF/sTON (+Tet/–PI); p27Kip1ON/sTOFF (–Tet/+PI); p27Kip1OFF/sTOFF (+Tet/+PI) by immunofluorescence without any signs of crossregulation (Fig. 2B).
For construction of stable pMF233-containing CHO-TWIN1108 cell lines both expression units of pMF233 were repressed by addition of pristinamycin (2 µg/ml) and tetracycline (2 µg/ml) during transfection and clonal selection. CHO-23393 and CHO-233176, two randomly chosen clones out of 10 p27Kip1- and sT-expressing clones were used for further analysis. In order to evaluate the potential of pMF233 to provide complete proliferation control of CHO cells, the growth characteristics and cell-cycle profiles of CHO-23393 and CHO-233176 were analyzed under mutually exclusive p27Kip1 and sT expression scenarios [p27Kip1OFF/sTON (+Tet/–PI); p27Kip1ON/sTOFF (–Tet/+PI)] (Figs 3 and 4). Although expression of sT alone (p27Kip1OFF/sTON) increased proliferation of both CHO-233 cell clones, expression of only p27Kip1 (p27Kip1ON/ sTOFF) did not result in desired growth arrest of CHO-23393 or CHO-233176 clones (Fig. 3). Cell-cycle analysis of CHO-233176 confirmed the absence of a G1-specific growth arrest in the p27Kip1ON/sTOFF (–Tet/+PI) configuration compared to the p27Kip1OFF/ sTOFF (+Tet/+PI) control set-up and the isogenic parental cell line CHO-TWIN1108 (Fig. 4). As expected, increased proliferation observed under p27Kip1OFF/sTON conditions did not alter the cell-cycle distribution relative to the CHO-TWIN1108 control (Fig. 4). Since enforced expression of human p27Kip1 has previously been shown to arrest CHO cells in the G1-phase of the cell cycle (9), it is possible that in this p27Kip1ON/sTOFF scenario, induction of p27Kip1 results in leaky expression of the downstream sT expression and/or PPIR shows some basal activity. In both cases, the strong oncogene sT may override p27Kip1-mediated arrest signals or prevent p27Kip1 expression to reach arrest-specific concentrations (13). Indeed, western blot analysis shows that significant leaky expression of sT correlates with induction of p27Kip1 expression (Fig. 5). It is thus clear that the balance between the basal expression of the growth-promoting determinant and p27Kip1 expression are crucial for successful proliferation control independent of the combination of growth-promoting and growth-inhibiting determinants used. To overcome these complex issues, a single gene-based strategy for complete proliferation management of CHO cells was designed around p27Kip1.
Figure 3.
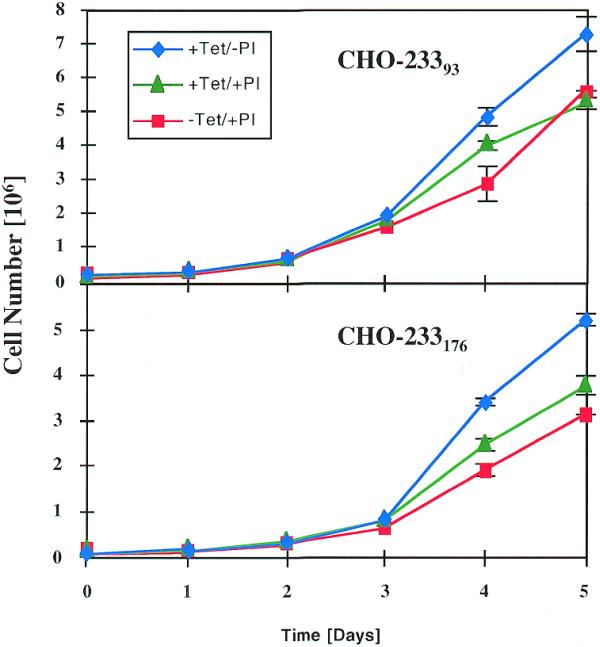
Growth profile of CHO-233 cells expressing p27Kip1 or sT. Cell lines CHO-23393 and CHO-233176 harboring the dual regulation plasmid pMF233 were seeded in triplicate cultures at 105 cells/ml under three different conditions: p27Kip1OFF/sTOFF (+Tet+PI), p27Kip1OFF/sTON (+Tet/–PI) and p27Kip1ON/sTOFF (–Tet/+PI), where +Tet and +PI are cultures containing tetracycline and pristinamycin, respectively (Fig. 1).
Figure 5.
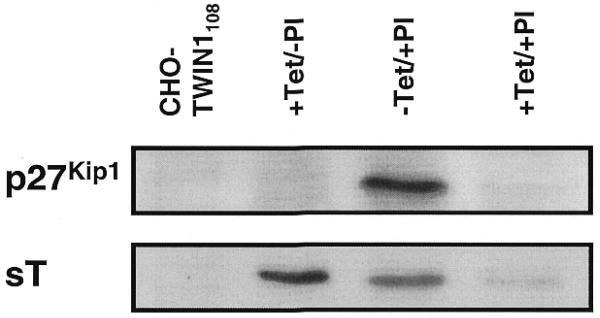
Western blot analysis of CHO-233176. p27Kip1 and sT expression of CHO-233176 was analyzed in the presence (+) and absence (–) of tetracycline (Tet) or pristinamycin I (PI). Owing to the consecutive orientation, induction of p27Kip1 significantly increases leaky expression of sT.
Complete proliferation control of CHO cells by p27Kip1-sense and p27Kip1-antisense expression
Based on the convergent dual regulated expression vector pDuoRex3 (Fig. 1) we constructed pMF226 which encodes human p27Kip1 driven in sense (p27Kip1S) orientation by PhCMV*-1 and in antisense (p27Kip1AS) orientation by PPIR (PhCMV*-1→p27Kip1←PPIR; Fig. 6A). Success of the convergent dual regulated expression configurations would be most desirable because they are inherently less sensitive to the leakiness of antibiotic-responsive promoters than consecutive set-ups because the expressed unit transcriptionally silences its repressed counterpart.
Figure 6.
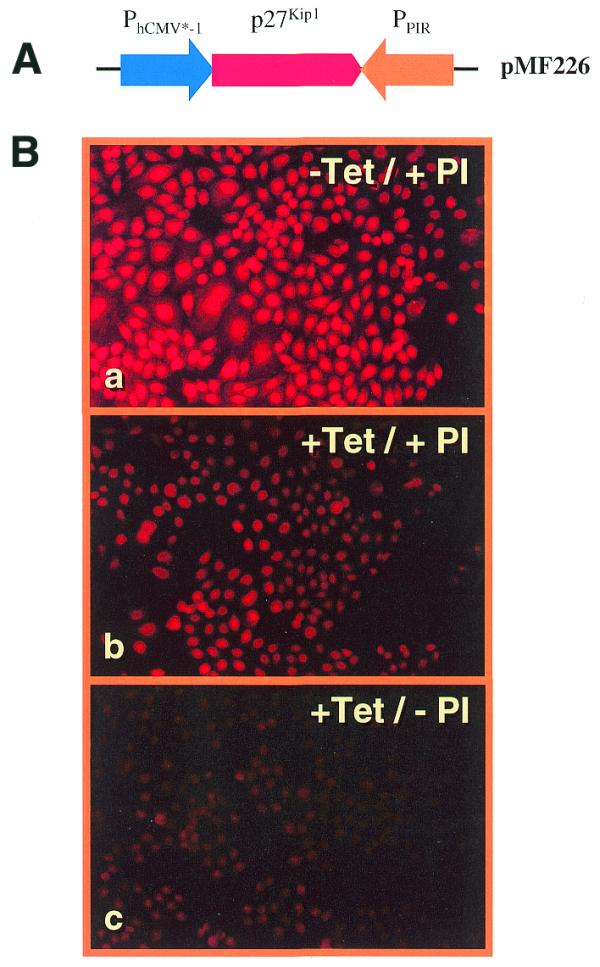
(A) Schematic of the p27Kip1 sense/antisense expression vector pMF226. p27Kip1 sense expression is driven by the tetracycline-responsive promoter (PhCMV*-1) and p27Kip1 antisense expression is controlled by the PI-responsive promoter (PPIR). (B) p27Kip1-based immunofluorescence of the pMF226-harboring cell line CHO-22696 expressing (a) sense p27Kip1 only (–Tet/+PI), (b) neither gene (+Tet/+PI, complete repression) and (c) antisense p27Kip1 only (+Tet/–PI). The images represent a quantitative measure of p27Kip1 accumulation after 48 h of induction.
During the transfection and selection procedure, the dual expression units were repressed by addition of pristinamycin (2 µg/ml) and tetracycline (2 µg/ml). CHO-22696, CHO-226100 and CHO-226108, three randomly chosen clones out of 20 p27Kip1-expressing clones, were used for further studies. The p27Kip1 expression profiles p27Kip1SON/p27Kip1ASOFF (–Tet/+PI), p27Kip1SOFF/p27Kip1ASOFF (+Tet/+PI) and p27Kip1SOFF/p27Kip1ASON (+Tet/–PI) were analyzed by p27Kip1-based immunofluorescence, a quantitative measure of p27Kip1 accumulation 48 h after induction (shown for CHO-22696 in Fig. 6). Significant overexpression of p27Kip1 is observed upon sense expression and intracellular levels of this CKI are reduced below endogenous levels upon exclusive antisense expression of p27Kip1 (Fig. 6B). Analysis of the growth profiles of CHO-22696, CHO-226100 and CHO-226108 demonstrated that p27Kip1S expression successfully arrests proliferation of these engineered CHO derivatives for extended periods (Fig. 7). However, upon exclusive expression of antisense p27Kip1, all three cell clones displayed significantly increased proliferation, reaching almost twice the cell density of CHO-226 clones grown in the presence of pristinamycin and tetracycline (control culture, p27Kip1SOFF/p27Kip1ASOFF) (Fig. 7). FACS-mediated cell-cycle analysis of CHO-22696 clearly confirmed a G1-specific growth arrest under p27Kip1SON/p27Kip1ASOFF conditions while increased proliferation observed with p27Kip1SOFF/p27Kip1ASON configurations did not alter the overall cell-cycle distribution compared to control set-ups (CHO-TWIN1108 and p27Kip1SOFF/p27Kip1ASOFF) (Fig. 4).
Figure 7.
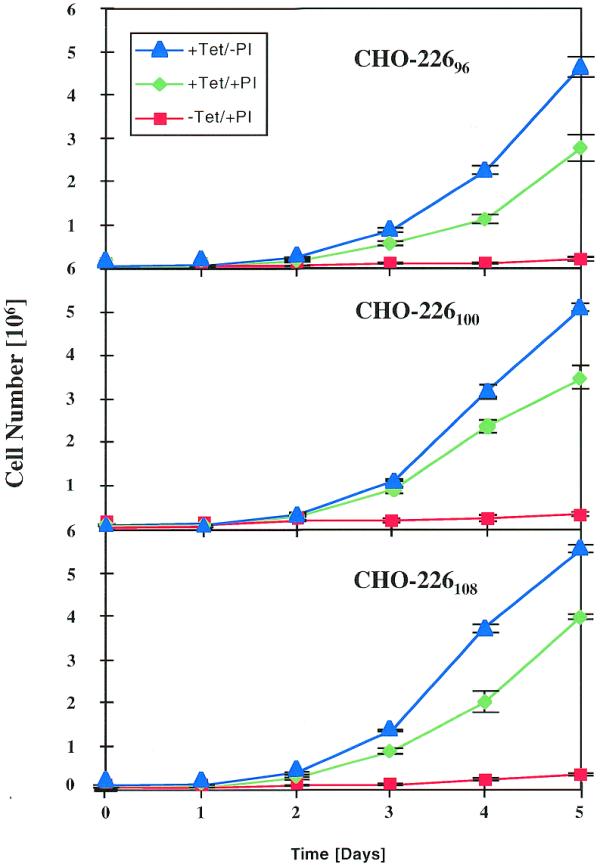
Growth profiles of CHO-226 cells expressing p27Kip1S or antisense p27Kip1AS. Three clones (CHO-22696, CHO-226100, CHO-226108) harboring the plasmid pMF226 (PhCMV*-1-p27Kip1-PPIR) were seeded in triplicate cultures at an initial population of 105 cells/ml under three different conditions: p27Kip1SOFF/p27Kip1ASON (+Tet/–PI), p27Kip1SON/p27Kip1ASOFF (–Tet/+PI), or p27Kip1SOFF/p27Kip1ASOFF (+Tet/+PI, complete repression of both genes).
DISCUSSION
Advances in biomedicine have accentuated the use of gene therapy as an attractive platform for the delivery of a therapeutic protein (16,17). Human-compatible gene regulation technology will become an increasingly important option for pharmacological control of a therapeutic that is delivered by an in vivo vector because it will allow titration of the circulating protein into the desired range, enable the adaptation of a therapy to fluctuating daily dosing regimes and allow reversibility of the therapy as needed (16,17).
Heterologous gene regulation strategies are also used to reprogram mammalian pathways to achieve therapeutic or bioprocess goals (9,18). Cell proliferation control is a particularly important parameter because it dominates many aspects of human disease and therapy including apoptosis (19), differentiation (20), regeneration (21) and cancer (22). In nature, molecular pathways promoting positive and negative proliferation control are carefully balanced by regulation of opposing genes (4). Overexpression of the human CKI p27Kip1 alone arrests mammalian cells in the G1-phase of the cell cycle (9). However, when the same gene was combined with the growth-promoting sT gene in a dual regulated expression configuration, p27Kip1 expression did not induce sustained growth arrest even when sT was repressed by addition of pristinamycin (2 µg/ml). This experiment demonstrates the importance of balanced expression of genes with opposite growth effects because the residual expression of one gene may override the effect of the other. It is a major concern when implementing positive proliferation control engineering strategies in a human therapeutic context that attendant genes can be reliably silenced or their function overcome by induction of growth-arresting genes to prevent carcinogenesis. Based on such considerations, sT or other transforming genes are not ideal candidates to implement positive proliferation control in tissue engineering models. Rather, the reprogramming of quiescent cells to re-enter the cell cycle must mimic natural growth-inducing situations for the optimal integration of metabolic engineering into a complex regulation network. One approach to this was based on chemically induced dimerization [CID (23)] of signalling domains taken from endogenous erythropoietin (24), or thrombopoietin receptors (mpl) (25). Recent in vivo expansion of bone marrow cells engineered for CID-responsive mpl signalling did not result in leukemia in a group of five non-CID-treated control mice monitored over 1 year (26). However, truncated versions of the proto-oncogene c-mpl carried by the myeloproliferative leukemia virus (MPLV; v-mpl) have been reported to immortalize hematopoietic progenitor cells (27). Due to the fine line between successful growth-inducing engineering strategies and the risk of long-term induction of neoplastic growth any therapeutic cell expansion strategy will likely require counteracting growth-arresting molecular interventions prior to approval in human gene therapy.
p27Kip1 may be an ideal candidate for opposite proliferation control because the reduction of p27Kip1 levels is a critical event for S-phase entry during the mammalian cell cycle (11,12,28). Constitutive antisense expression of p27Kip1 has enabled cells to grow for several generations in medium supplemented with insulin and transferrin (12) or in medium containing low concentrations of serum (7). Furthermore, addition of antisense p27Kip1 oligonucleotides (coupled with TGFβ-neutralizing antibodies) induced proliferation of hematopoietic stem cells to an extent that enabled transduction by Moloney-based retroviruses, and targeted disruption of p27Kip1 resulted in enhanced growth of the mice and a striking enlargement of their thymus, pituitary, adrenal and gonadal organs (29–31). We have successfully used streptogramin-regulated full-length antisense expression of human p27Kip1 to increase proliferation of CHO cells even under optimal growth conditions while growth could be completely blocked in the G1-phase by subsequent tetracycline-responsive expression of sense p27Kip1. Dual regulated expression technology may be used as a prototype for next generation tissue engineering strategies which will likely require reprogramming of both growth characteristics and the redirection of differentiation pathways in a desired manner (32,33).
Acknowledgments
ACKNOWLEDGEMENTS
We thank Barbara Lennon and Jim Zanghi for critical comments on the manuscript and Eva Niederer for FACS analysis. This work was supported by the Swiss Priority Program in Biotechnology (SPP BioTech outphasing), the Novartis Foundation and Cistronics Cell Technology GmbH (Einsteinstrasse 1-5, CH-8093 Zurich, Switzerland).
References
- 1.Mulligan R.C. (1993) The basic science of gene therapy. Science, 260, 926–932. [DOI] [PubMed] [Google Scholar]
- 2.Fussenegger M., Morris,R., von Stockar,B., Fux,C., Rimann,M., Thompson,C.J. and Bailey,J.E. (2000) Novel streptogramin-based gene regulation systems for mammalian cells. Nat. Biotechnol., 18, 1203–1208. [DOI] [PubMed] [Google Scholar]
- 3.Gossen M. and Bujard,H. (1992) Tight control of gene expression in mammalian cells by tetracycline-responsive promoters. Proc. Natl Acad. Sci. USA, 89, 5547–5551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fussenegger M. and Bailey,J.E. (1998) Molecular regulation of cell-cycle progression and apoptosis in mammalian cells: implications for biotechnology. Biotechnol. Prog., 14, 807–833. [DOI] [PubMed] [Google Scholar]
- 5.Kato J.-Y., Matsuoka,M., Polyak,K., Massagué,J. and Sherr,C.J. (1994) Cyclin AMP-induced G1 phase arrest mediated by an inhibitor (p27Kip1) of cyclin-dependent kinase 4 activation. Cell, 79, 487–496. [DOI] [PubMed] [Google Scholar]
- 6.Polyak K., Lee,M.-L., Erdjument-Bromage,H., Koff,A., Roberts,J.M., Tempst,P. and Massagué,J. (1994) Cloning of P27Kip1, a cyclin-dependent kinase I inhibitor and a potential mediator of extracellular antimitogenic signals. Cell, 78, 59–66. [DOI] [PubMed] [Google Scholar]
- 7.Coats S., Flanagan,W.M., Nourse,M. and Roberts,J.M. (1996) Requirement of p27Kip1 for restriction point control of the fibroblast cell cycle. Science, 272, 877–880. [DOI] [PubMed] [Google Scholar]
- 8.Polyak K., Kato,J., Solomon,M.J., Sherr,C.J., Massagué,J., Roberts,J.M. and Koff,A. (1994) p27Kip1, a cyclin-Cdk inhibitor, links transforming growth factor-beta and contact inhibition to cell cycle arrest. Genes Dev., 8, 9–22. [DOI] [PubMed] [Google Scholar]
- 9.Fussenegger M., Schlatter,S., Dätwyler,D., Mazur,X. and Bailey,J.E. (1998) Controlled proliferation by multigene metabolic engineering enhances the productivity of Chinese hamster ovary cells. Nat. Biotechnol., 16, 468–472. [DOI] [PubMed] [Google Scholar]
- 10.Greulich H. and Erikson,R.L. (1998) An analysis of Mek1 signaling in cell proliferation and transformation. J. Biol. Chem., 273, 13280–13288. [DOI] [PubMed] [Google Scholar]
- 11.Ladha M.H., Lee,K.Y., Upton,T.M., Reed,M.F. and Ewen,M.E. (1998) Regulation of exit from quiescence by p27 and cyclin D1-CDK4. Mol. Cell Biol., 18, 6605–6615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rivard N., Boucher,M.-J., Asselin,C. and L’Allemain,G. (1999) MAP kinase cascade is required for p27 downregulation and S phase entry in fibroblasts and epithelial cells. Am. J. Physiol., 277, 652–664. [DOI] [PubMed] [Google Scholar]
- 13.Porras A., Gaillard,S. and Rundell,K. (1999) The simian virus 40 small-t and Large-T antigens jointly regulate cell cycle reentry in human fibroblasts. J. Virol., 73, 3102–3107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Fussenegger M., Mazur,X. and Bailey,J.E. (1997) A novel cytostatic process enhances the productivity of Chinese hamster ovary cells. Biotechnol. Bioeng., 55, 927–939. [DOI] [PubMed] [Google Scholar]
- 15.Baron U., Freundlieb,S., Gossen,M. and Bujard,H. (1995) Co-regulation of two gene activities by tetracycline via a bidirectional promoter. Nucleic Acids Res., 23, 3605–3606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Rivera V.M., Wang,X., Wardwell,S., Courage,N.L., Volchuk,A., Keenan,T., Holt,D.A., Gilman,M., Orci,L., Crasoli,F.,Jr, Rothman,J.E. and Clackson,T. (2000) Regulation of protein secretion through controlled aggregation in the endoplasmic reticulum. Science, 287, 826–830. [DOI] [PubMed] [Google Scholar]
- 17.Clackson T. (2000) Regulated gene expression systems. Gene Ther., 7, 120–125. [DOI] [PubMed] [Google Scholar]
- 18.Umaña P., Jean-Mairet,J. and Bailey,J.E. (1999) Engineered glycoforms of an antineuroblastoma IgG1 with optimized antibody-dependent cellular cytotoxic activity. Nat. Biotechnol., 17, 176–180. [DOI] [PubMed] [Google Scholar]
- 19.Fussenegger M., Bailey,J.E. and Varner,J. (2000) A mechanistic model of the cell death machine: caspases. Nat. Biotechnol., 17, 768–718. [DOI] [PubMed] [Google Scholar]
- 20.Timchenko N.A., Wilde,M., Nakanishi,M., Smith,J.R. and Darlington,G.J. (1996) CAAT/enhancer-binding protein α (C/EBPα) inhibits cell proliferation through the p21 (WAF-1/CIP-1/SDI-1) protein. Genes Dev., 10, 804–815. [DOI] [PubMed] [Google Scholar]
- 21.Werner S. (1998) Keratinocyte growth factor: a unique player in epithelial repair processes. Cytokine Growth Factor Rev., 9, 153–165. [DOI] [PubMed] [Google Scholar]
- 22.Kouraklis G. (1999) Progress in cancer gene therapy. Acta Oncol., 38, 675–683. [DOI] [PubMed] [Google Scholar]
- 23.Spencer D.M., Wandless,T.J., Schreiber,S.L. and Crabtree,G.R. (1993) Controlling signal transduction with synthetic ligands. Science, 262, 1019–1034. [DOI] [PubMed] [Google Scholar]
- 24.Blau C.A., Peterson,K.R., Drachman,J.G. and Spencer,D.M. (1997) A proliferation switch for genetically modified cells. Proc. Natl Acad. Sci. USA, 94, 3076–3081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Jin L., Asano,H. and Blau,C.A. (1998) Stimulating cell proliferation through the pharmacologic activation of c-kit. Blood, 91, 890–897. [PubMed] [Google Scholar]
- 26.Jin L., Chien,S., Otto,K.G., Richard,R.E., Emery,D.W. and Blau,C.A. (2000) In vivo selection using a cell-growth switch. Nature Genet., 26, 64–66. [DOI] [PubMed] [Google Scholar]
- 27.Souyri M., Vigon,I., Penciolelli,J.F., Heard,J.M., Tambouring,P. and Wendling,F. (1990) A putative truncated cytokine receptor gene transduced by the myeloproliferative leukemia virus immortalizes hematopoietic progenitors. Cell, 63, 1137–1147. [DOI] [PubMed] [Google Scholar]
- 28.Aktas H., Cai,H. and Cooper,G.M. (1997) Ras links growth factor signalling to the cell cycle machinery via regulation of cyclin D1 and the Cdk inhibitor p27Kip1. Mol. Cell Biol., 17, 3850–3857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fero M.L., Rivkin,M., Tasch,M., Porter,P., Carow,C.E., Firpo,E., Polyak,K., Tsai,L.H., Broudy,V., Perlmutter,R.M., Kaushansky,K. and Roberts,J.M. (1996) A syndrome of multiorgan hyperplasia with features of gigantism, tumorigenesis, and female sterility in p27Kip1-deficient mice. Cell, 85, 733–744. [DOI] [PubMed] [Google Scholar]
- 30.Kiyokawa H., Kineman,R.D., Manova,T.K., Soares,V.C., Hoffman,E.S., Ono,M., Khanam,D., Hayday,A.C., Frohman,L.A. and Koff,A. (1996) Progress in cancer gene therapy. Cell, 85, 721–732. [DOI] [PubMed] [Google Scholar]
- 31.Nakayama K., Ishida,N., Shirane,M., Inomata,A., Inoue,T., Shishido,N., Horii,I., Loh,D.Y. and Nakayama,K. (1999) Mice lacking p27Kip1 display increased body size, multiple organ hyperplasia, retinal dysplasia, and pituitary tumors. Cell, 85, 707–720. [DOI] [PubMed] [Google Scholar]
- 32.Mueller C., Alunni-Babbroni,M., Kowenz-Leutz,E., Mo,X., Tommasino,M. and Leutz,A. (1999) Separation of C/ebpα-mediated proliferation arrest and differentiation pathways. Proc. Natl Acad. Sci. USA, 96, 7276–7281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ross S.E., Hemati,N., Longo,K.A., Bennett,C.N., Lucas,P.C., Erickson,R.L. and MacDougald,O.A. (2000) Inhibition of adipogenesis by wnt signaling. Science, 289, 950–953. [DOI] [PubMed] [Google Scholar]