

Abstract
The ability of microorganisms to adapt to changing environments challenges the prediction of their history-dependent behavior. Cellular biomarkers that are quantitatively correlated to stress adaptive behavior will facilitate our ability to predict the impact of these adaptive traits. Here, we present a framework for identifying cellular biomarkers for mild stress induced enhanced microbial robustness towards lethal stresses. Several candidate-biomarkers were selected by comparing the genome-wide transcriptome profiles of our model-organism Bacillus cereus upon exposure to four mild stress conditions (mild heat, acid, salt and oxidative stress). These candidate-biomarkers—a transcriptional regulator (activating general stress responses), enzymes (removing reactive oxygen species), and chaperones and proteases (maintaining protein quality)—were quantitatively determined at transcript, protein and/or activity level upon exposure to mild heat, acid, salt and oxidative stress for various time intervals. Both unstressed and mild stress treated cells were also exposed to lethal stress conditions (severe heat, acid and oxidative stress) to quantify the robustness advantage provided by mild stress pretreatment. To evaluate whether the candidate-biomarkers could predict the robustness enhancement towards lethal stress elicited by mild stress pretreatment, the biomarker responses upon mild stress treatment were correlated to mild stress induced robustness towards lethal stress. Both short- and long-term biomarkers could be identified of which their induction levels were correlated to mild stress induced enhanced robustness towards lethal heat, acid and/or oxidative stress, respectively, and are therefore predictive cellular indicators for mild stress induced enhanced robustness. The identified biomarkers are among the most consistently induced cellular components in stress responses and ubiquitous in biology, supporting extrapolation to other microorganisms than B. cereus. Our quantitative, systematic approach provides a framework to search for these biomarkers and to evaluate their predictive quality in order to select promising biomarkers that can serve to early detect and predict adaptive traits.
Introduction
Bacteria are constantly faced with changing environmental conditions and have evolved sophisticated mechanisms to adapt to changing environments. Environmental changes trigger a cascade of cellular events, and regulatory networks of microorganisms serve to prime cells to be prepared for later challenges even before they arise [1], [2]. The adaptive stress response is a crucial survival strategy for a wide spectrum of microorganisms, including food spoilage bacteria, pathogens and organisms used in functional food applications, and prediction of this stress adaptive behavior will allow to control and/or exploit these adaptive traits.
Bacteria are not only exposed to changing environments in their natural habitats but also during industrial processing, and activation of stress adaptation mechanisms can provide cell robustness to harsher stress conditions including stresses other than the one that induced the adaptive stress response. This so-called cross-protection phenomenon challenges the hurdle preservation strategy that targets to guarantee the microbial safety and stability as well as the sensory and nutritional quality of minimally processed foods by simultaneous or successive application of multiple mild preservation factors [3]. While individual hurdles may not be effective in controlling growth of food spoilage bacteria and pathogens, the right combination of hurdles allows to control growth of microorganisms and to minimize organoleptic changes in foods. However, the ability of spoilage and pathogenic microorganisms to adapt to stressing environments could antagonize the benefits of the hurdle preservation strategy. Long- and short-term exposure to mild stress conditions showed to induce (cross-)protection towards otherwise lethal stress conditions [4], [5], [6] and might even affect the virulence of pathogens [7], [8]. The ability of microorganisms to gain cellular robustness upon activation of adaptive stress responses is also beneficial for various industrial applications [9], including the development of reliable starter cultures [10] and selection of robust probiotic strains [11].
Previous studies demonstrated that our model-organism Bacillus cereus gained increased resistance upon preexposure to several mild food preservation stresses [12]–[15], underlining the significance of a better understanding of its stress adaptive behavior. The availability of complete genome sequences of microbes and the development of functional genomics technologies have provided a wealth of data and opportunities to better understand stress adaptation mechanisms. Comparison of genome-wide transcriptome profiles of microorganisms in response to diverse environmental conditions can reveal general stress response features [16]–[19], and could lead to identification of cellular indicators for stress adaptive behavior. Recently, we investigated the genome-wide transcriptome response of B. cereus during exposure to several mild stresses [20]–[23], and these transcriptome analyses gave insight into general and stress-specific adaptive responses, and opened avenues towards identification of potential cellular biomarkers that can predict stress adaptive behavior. Here, we present a framework for the identification of such cellular biomarkers for mild stress induced enhanced robustness towards lethal stresses. We identified both short- and long-term biomarkers of which their induction levels were quantitatively correlated to the mild stress induced enhanced robustness level of the cell. The identification of predictive cellular indicators for stress adaptive behavior will enable the prediction of the impact of adaptive responses during mild stress (processing) conditions on subsequent microbial robustness, and can be exploited to control these stress adaptive traits.
Results
Experimental strategy for identifying biomarkers for mild stress induced enhanced robustness
In search of potential biomarkers for mild stress induced enhanced robustness, the genome-wide transcriptome profiles of our model-organism B. cereus ATCC 14579 grown until the mid-exponential growth phase and then exposed to various mild stress conditions [20]–[23] were compared. This comparison revealed a remarkable limited number of genes (see Table S1 in the supplementary material) that were differentially expressed upon treatment to all those mild stress conditions. This overlap of transcriptome responses included various defense mechanisms and regulatory and metabolic pathways, including cellular defense mechanisms against oxidative stress, factors involved in repair and maintenance of cellular protein quality and energy production, and transcriptional regulators. In order to select the most promising functional categories that could point to candidate-biomarkers, the expression ratios of the differentially expressed genes were evaluated by marking the genes of which the expression ratios were at least five upon exposure to one mild stress condition and at least two upon exposure to two other mild stress conditions. These genes represented three main functional categories: (1) members of the general stress response regulon controlled by transcriptional regulator σB; (2) cellular defense mechanisms against oxidative stress; and (3) repair and maintenance of cellular protein quality. This transcriptional overlap in stress adaptation seemed to be stress-independent, supporting its suitability as a source for identification of potential candidate-biomarkers for stress adaptation. Based on these three categories, candidate-biomarker transcripts for stress adaptive behavior were assigned, namely that of sigB, catA, catE, clpB, clpC and clpP. Conceivably, not only transcripts but also proteins and enzymes could function as biomarker, and therefore, also the proteins SigB, ClpC and ClpP, and catalase activity were included as candidate-biomarkers, resulting in ten candidate-biomarkers (Figure 1a).
Figure 1. Conceptual scheme for measuring and correlating mild stress induced biomarker and robustness responses.

a) Candidate-biomarkers - a transcriptional regulator (activating general stress responses), catalases (removing reactive oxygen species, ROS), and chaperones and proteases (maintenance of protein quality) - were quantitatively measured at transcript, protein and/or activity level upon exposure to four mild stress conditions (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) for various time intervals. Mild stress treated cells were also exposed to three lethal stresses (50°C, pH 3.3, 0.2 mM H2O2) to quantify their robustness towards these lethal stresses following mild stress pretreatment. b) To evaluate whether the candidate-biomarker could predict the robustness level of mild stress pretreated cells, the mild stress induced biomarker and robustness responses were correlated to evaluate the significance of their correlation.
To quantify the stress adaptive behavior of B. cereus to a wide spectrum of stress conditions imposed on bacteria, four mild stress conditions were used to mildly stress the cells for various time intervals, namely, heat stress (43°C), acid-shock (pH 5.5), osmotic-upshift (1.5% NaCl), and oxidative stress (0.1 mM H2O2) (see Figure S1 in the supplementary material for more details about the procedure to select these mild stress conditions for adaptation experiments). Both unstressed and mild stress treated cells were subsequently exposed to three lethal stress inactivation conditions − heat stress (50°C), low pH shock (pH 3.3) and high oxidative stress (0.2 mM H2O2) to determine their specific robustness level towards these lethal stresses. Severe osmotic-upshift (up to 30% sodium chloride [w/v]; reaches maximum saturation of sodium chloride in BHI) did not result in inactivation patterns comparable to those observed with the other three lethal stress conditions (data not shown) and was therefore not used as lethal stress. To quantify the robustness enhancement towards these lethal stresses provided by mild stress pretreatment, the robustness level of mild stress pretreated cells was compared to that of unstressed cells. To evaluate whether one or more of the selected candidate-biomarkers could quantitatively predict the robustness enhancement following mild stress pretreatment, the induction of the candidate-biomarkers was determined upon mild stress treatment and correlated to mild stress induced enhanced robustness towards lethal heat, acid and oxidative stress (Figure 1b). After correlating the mild stress induced biomarker and robustness responses, the Pearson correlation coefficient was calculated to evaluate the significance of the correlations in order to select potential biomarkers for mild stress induced enhanced robustness (Figure 1b).
Induction of candidate-biomarkers and robustness towards lethal stress in response to mild stress treatment
Mild heat stress treatment resulted in induction of almost all candidate-biomarkers, but their induction patterns differed (Figure 2a). The relative transcription levels of sigB and catE, which is a regulon member of σB [23], [24], displayed transient increased expression up to 15 min of mild heat stress treatment and decreased afterwards, whereas the transcription patterns of clpB, clpC and clpP were rather constant over time. Upon 60 min of mild heat stress treatment, the relative transcription levels of sigB and catE increased again, and this is in correspondence with upregulation of SigB-dependent genes in B. subtilis cells entering the transition growth phase [25]. The gene encoding the main vegetative catalase, catA, was just significantly transcribed at one mild heat stress treatment time point, and also, no increased catalase activity was observed in mild heat stress treated cells compared to unstressed cells. This confirmed that the role of catalase CatE is less significant in total cellular catalase activity compared to the main vegetative catalase CatA [26]. Mild heat stress treatment resulted in increased production of the proteins SigB, ClpC and ClpP compared to that of unstressed cells and the production patterns of both Clp proteins were comparable.
Figure 2. Induction of candidate-biomarkers and robustness in response to mild heat stress treatment.

a) Candidate-biomarkers − the transcripts of sigB, catA, catE, clpB, clpC and clpP, the proteins SigB, ClpC and ClpP, and catalase activity − were quantitatively measured in unstressed (uns) cultures (OD600 nm ∼0.4–0.5) and in the stressed cultures (s) after 43°C stress treatment for 2, 5, 10, 15, 20, 30 and 60 min. The columns mark the log10-fold induction after mild heat stress treatment compared to unstressed cells. b) Unstressed and 43°C-treated cells were inactivated by exposure to 50°C for 5 min, pH 3.3 at 30°C for 3.5 min, and 0.2 mM H2O2 at 30°C for 2 min, to determine their robustness towards lethal heat, acid and oxidative stress, respectively. The columns mark the number of microorganisms surviving the lethal stress treatment compared to the initial number of microorganisms (%). Error bars represent standard errors of the reproductions.
Mild heat stress pretreatment provided a robustness advantage towards lethal heat stress treatment, and also conferred cross-protection towards lethal acid stress, whereas no increased resistance towards lethal oxidative stress was observed upon mild heat stress pretreatment (Figure 2b).
Besides mild heat stress treatment, cells were also short- and long-term exposed to mild acid, salt and oxidative stress and the induction patterns of the candidate-biomarkers upon these mild stress treatments are shown in Figure 3a, 4a and 5a, respectively. The induction patterns of the candidate-biomarkers were highly affected by the type of mild stress condition applied. In addition to mild heat stress treatment, also mild acid and salt stress treatment significantly increased transcription of sigB and catE and led to increased SigB protein levels compared to unstressed cells, whereas these responses were not significant in mild oxidative stress treated cells. Mild acid stress treatment for up to 30 min elicited just significantly increased transcription of clpB, clpC, and clpP compared to unstressed cells, but increased production of the ClpC and ClpP proteins could not be demonstrated. Both mild salt and oxidative stress did not significantly increase the transcription of clpB, clpC, and clpP nor did these mild stress conditions lead to higher levels of the ClpC and ClpP proteins. In contrast to mild heat stress treatment, exposure to mild acid, salt and oxidative stress resulted in significantly increased transcription of catA and cellular catalase activity.
Figure 3. Induction of candidate-biomarkers and robustness in response to mild acid stress treatment.

a) Candidate-biomarkers − the transcripts of sigB, catA, catE, clpB, clpC and clpP, the proteins SigB, ClpC and ClpP, and catalase activity − were quantitatively measured in unstressed (uns) cultures (OD600 nm ∼0.4–0.5) and in the stressed cultures (s) after pH 5.5 stress treatment for 2, 5, 10, 15, 20, 30 and 60 min. The columns mark the log10-fold induction after mild acid stress treatment compared to unstressed cells. b) Unstressed and pH 5.5-treated cells were inactivated by exposure to 50°C for 5 min, pH 3.3 at 30°C for 3.5 min, and 0.2 mM H2O2 at 30°C for 2 min, to determine their robustness towards lethal heat, acid and oxidative stress, respectively. The columns mark the number of microorganisms surviving the lethal stress treatment compared to the initial number of microorganisms (%). Error bars represent standard errors of the reproductions.
Figure 4. Induction of candidate-biomarkers and robustness in response to mild salt stress treatment.

a) Candidate-biomarkers − the transcripts of sigB, catA, catE, clpB, clpC and clpP, the proteins SigB, ClpC and ClpP, and catalase activity − were quantitatively measured in unstressed (uns) cultures (OD600 nm ∼0.4–0.5) and in the stressed cultures (s) after 1.5% NaCl stress treatment for 2, 5, 10, 15, 20, 30 and 60 min. The columns mark the log10-fold induction after mild salt stress treatment compared to unstressed cells. b) Unstressed and 1.5% NaCl-treated cells were inactivated by exposure to 50°C for 5 min, pH 3.3 at 30°C for 3.5 min, and 0.2 mM H2O2 at 30°C for 2 min, to determine their robustness towards lethal heat, acid and oxidative stress, respectively. The columns mark the number of microorganisms surviving the lethal stress treatment compared to the initial number of microorganisms (%). Error bars represent standard errors of the reproductions.
Figure 5. Induction of candidate-biomarkers and robustness in response to mild oxidative stress treatment.

a) Candidate-biomarkers − the transcripts of sigB, catA, catE, clpB, clpC and clpP, the proteins SigB, ClpC and ClpP, and catalase activity − were quantitatively measured in unstressed (uns) cultures (OD600 nm ∼0.4–0.5) and in the stressed cultures (s) after 0.1 mM H2O2 stress treatment for 2, 5, 10, 15, 20, 30 and 60 min. The columns mark the log10-fold induction after mild oxidative stress treatment compared to unstressed cells. b) Unstressed and 0.1 mM H2O2-treated cells were inactivated by exposure to 50°C for 5 min, pH 3.3 at 30°C for 3.5 min, and 0.2 mM H2O2 at 30°C for 2 min, to determine their robustness towards lethal heat, acid and oxidative stress, respectively. The columns mark the number of microorganisms surviving the lethal stress treatment compared to the initial number of microorganisms (%). Error bars represent standard errors of the reproductions.
Mild acid stress pretreatment resulted in enhanced robustness towards lethal acid stress and provided also cross-protection towards lethal oxidative stress, whereas no significantly increased resistance towards lethal heat stress was observed (Figure 3b). On the other hand, mild salt stress pretreatment provided cross-protection towards lethal heat and oxidative stress, but did not confer cross-protection towards lethal acid stress (Figure 4b). Mild oxidative stress pretreatment provided cells enhanced robustness towards lethal heat, acid, and also oxidative stress, and therefore, only this mild stress condition conferred enhanced robustness to all three lethal stresses tested (Figure 5b). In Table S2 in the supplementary material, the effects of the different mild stress pretreatments on robustness towards lethal heat, acid and oxidative stress are summarized.
Framework for identifying biomarkers for mild stress induced enhanced robustness
A decision flow chart was designed to evaluate for each candidate-biomarker whether it could indeed predict the enhanced robustness level of mild stress pretreated cells, and therefore functioned as biomarker (Figure 6a). Two types of biomarkers were formulated, namely, long-term biomarkers and short-term biomarkers, and these two types of biomarkers will be discussed below. A step-wise procedure was followed to select those conditions where the candidate-biomarker functioned as biomarker. Firstly, the mild stress induced biomarker and robustness responses upon mild stress treatment for 2, 5, 10, 15, 20, 30 to 60 min (7 treatment time points) compared to that of unstressed cells were correlated for each mild stress and lethal stress pair, and the Pearson correlation coefficient was tested for significance (P<0.05). After testing this correlation, the treatment time point of 60 min was excluded from the analysis, after which the correlation of the remaining treatment time points was tested for significance. The exclusion procedure was followed until the exclusion of treatment time point of 10 min. When the mild stress induced candidate-biomarker and robustness responses were significantly correlated for all mild stress treatment time intervals, then the candidate-biomarker was qualified as long-term biomarker for that mild stress and lethal stress pair. The candidate-biomarker was qualified as short-term biomarker when the mild stress induced biomarker and robustness responses were only significantly correlated for short-term adaptation time intervals.
Figure 6. Framework for identification of biomarkers for mild stress induced enhanced robustness.

a) Decision flow chart for identification of long-term and short-term biomarkers for mild stress induced enhanced robustness. Black box represents a long-term biomarker and gray box represents a short-term biomarker. These biomarkers are mild stress and lethal stress specific. Dashed gray box represents a no response condition. b) Decision flow chart for identification of a convergent-application biomarker that is induced by multiple mild stress conditions and is correlated to a specific lethal stress robustness. c) Decision flow chart for identification of a divergent-application biomarker that is induced by a specific mild stress condition and is correlated to robustness towards multiple lethal stresses.
Several biomarkers functioned as long-term or short-term biomarker for mild stress induced enhanced robustness for more than one specific mild stress and lethal stress pair. Moreover, some mild stress conditions did not significantly induce the biomarker and this corresponded to a lack of enhanced robustness following mild stress pretreatment (i.e. the so-called no response conditions, see Figure 6a). To evaluate whether the biomarker could predict the robustness level towards one specific lethal stress originated from multiple mild stress pretreatments, the mild stress induced biomarker and robustness responses were combined and the correlation was tested for significance. When this correlation remained significant (P<0.05), then the biomarker was defined as convergent-application biomarker (Figure 6b). The biomarker was defined as divergent-application biomarker when it could predict the robustness level towards multiple lethal stresses upon pretreatment to a specific mild stress (Figure 6c).
Identification of biomarkers for mild stress induced enhanced robustness
The step-wise procedure was followed to select those conditions for which the candidate-biomarker could predict the enhanced robustness level of the cell following mild stress pretreatment. The SigB protein functioned as a long-term biomarker for mild heat and mild salt stress induced enhanced robustness towards lethal heat stress (Figure 7). SigB was also significantly correlated to lethal heat stress robustness upon mild acid stress pretreatment, but because mild acid stress pretreatment did not significantly induce a robustness advantage towards lethal heat stress (Figure 3), SigB was not identified as a long-term biomarker for this condition. Since SigB could predict the robustness status towards lethal heat stress of both mild heat and mild salt stress pretreated cells, and remained significantly correlated to lethal heat stress robustness when these mild heat and mild salt stress induced responses were combined, SigB was qualified as convergent-application biomarker for lethal heat stress robustness. In addition to SigB protein, also the proteins ClpC and ClpP were identified as convergent-application biomarkers for lethal heat stress robustness, but their predictive potential was related to mild heat and mild acid stress pretreated cells (see Figure S2 and S3 in the supplementary material). Noteworthy, both ClpC and ClpP proteins could predict the enhanced robustness level towards lethal heat stress provided by mild heat pretreatment only for short adaptation intervals (2 to 30 min). Additionally, SigB was suitable to predict the robustness advantage towards both lethal acid and oxidative stress originating from preexposure to mild acid stress. Since SigB could still predict the mild acid stress induced robustness enhancement towards lethal acid and oxidative stress when these responses were combined, it also functioned as divergent-application biomarker.
Figure 7. Protein SigB as potential biomarker for mild stress induced enhanced robustness.

Induction of protein SigB upon mild stress treatment (43°C, ⋄; pH 5.5, ○; 1.5% NaCl, ▵; 0.1 mM H2O2, □) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of SigB and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which SigB was qualified as biomarker for mild stress induced enhanced robustness: black box represents a long-term biomarker; double black box represents a convergent-application biomarker; double gray box represents a divergent-application biomarker.
Catalase activity could predict the robustness level towards lethal oxidative stress upon pretreatment to more than two mild stress conditions (Figure 8). Catalase activity functioned as long-term biomarker for enhanced robustness towards lethal oxidative stress originating from mild acid and mild oxidative stress pretreatment. Additionally, mild heat stress did not induce catalase activity and this corresponded to a lack of enhanced robustness towards lethal oxidative stress (no response condition, see Figure 6a). This underlined that the predictive potential of catalase activity was not only restricted to mild stress conditions that induced enhanced robustness, but that it could also predict the lack of enhanced robustness following mild stress pretreatment. Because, catalase activity could still predict the robustness status towards lethal oxidative stress when these three mild stress induced responses were combined, it pointed to the potential of catalase activity to act as a vigorous convergent-application biomarker for lethal oxidative stress robustness. Additionally, catalase activity significantly correlated to robustness towards multiple lethal stresses induced upon pretreatment to mild acid or mild oxidative stress, and functioned therefore also as divergent-application biomarker.
Figure 8. Catalase activity as potential biomarker for mild stress induced enhanced robustness.

Induction of catalase activity upon mild stress treatment (43°C, ⋄; pH 5.5, ○; 1.5% NaCl, ▵; 0.1 mM H2O2, □) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of catalase activity and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which catalase activity was qualified as biomarker for mild stress induced enhanced robustness: black box represents a long-term biomarker; dashed gray box represents a no response condition; double black box represents a convergent-application biomarker; double gray box represents a divergent-application biomarker.
The transcripts of both clpC and clpP were qualified as convergent-application biomarkers for lethal acid stress robustness upon mild acid and mild salt stress pretreatment (Figure S4 and S5 in the supplementary material). In addition to protein SigB and catalase activity, clpC and clpP were also suitable to predict the robustness advantage towards multiple lethal stresses, namely lethal acid and oxidative stress, following mild acid stress pretreatment. Furthermore, also the transcript catE (Figure S6 in the supplementary material) showed similar divergent predictive potential for those conditions. Detailed supplementary information about the predictive potential of the proteins ClpC and ClpP, and the transcripts sigB, catA, catE, clpB, clpC and clpP is shown in Figures S2, S3, S4, S5, S6, S7, S8, and S9 in the supplementary material, and summarized for all the candidate-biomarkers in Table S3. Noteworthy, the transcript candidate-biomarkers were for various conditions identified as short-term biomarkers rather than long-term biomarkers.
A concluding overview of the predictive potential of the candidate-biomarkers is presented in Table 1 and 2. The SigB protein was suitable to predict the robustness enhancement towards lethal heat stress upon mild heat and mild salt stress pretreatment and was therefore qualified as convergent-application biomarker for lethal heat stress robustness (Table 1, see also Figure 7). Next to SigB, also the proteins ClpC and ClpP were qualified as convergent-application biomarkers for lethal heat stress robustness, but their potential to predict this stress adaptive behavior was related to mild heat and mild acid stress pretreated cells. The predictive potential of the transcripts clpC and clpP were comparable and both were qualified as convergent-application biomarkers for lethal acid stress robustness following mild acid and mild salt stress preexposure. Catalase activity emerged as a convergent-application biomarker for lethal oxidative stress robustness, which could be elicited by multiple mild stress pretreatments (e.g. mild heat, acid and oxidative stress) (see also Figure 8). Several biomarkers were correlated to robustness towards multiple lethal stresses upon pretreatment to a specific mild stress (Table 2). In addition to SigB, four other biomarkers − catE, clpC, clpP and catalase activity − showed similar predictive potential by emerging as divergent-application biomarker for lethal acid and lethal oxidative stress robustness following mild acid stress pretreatment. Therefore, this study also showed that various biomarkers can have similar predictive potential.
Table 1. Convergent-application biomarkers for mild stress induced enhanced robustness.
| Convergent-application biomarker multiple mild stresses, lethal stress specific | Mild stressa | Lethal stress |
| SigB | heat, salt | heat |
| ClpC | heat, acid | heat |
| ClpP | heat, acid | heat |
| clpC | acid, salt | acid |
| clpP | acid, salt | acid |
| catalase activity | heat, acid, H2O2 | H2O2 |
italic text indicates that mild stress treatment did not significantly induce the biomarker and also did not provide a robustness enhancement towards lethal stress upon pretreatment (no response condition).
Table 2. Divergent-application biomarkers for mild stress induced enhanced robustness.
| Divergent-application biomarker mild stress specific, multiple lethal stresses | Mild stress | Lethal stress |
| SigB | acid | acid, H2O2 |
| catE | acid | acid, H2O2 |
| clpC | acid | acid, H2O2 |
| clpP | acid | acid, H2O2 |
| catalase activity | acid | acid, H2O2 |
| catalase activity | H2O2 | heat, H2O2 |
Discussion
The adaptive stress response of bacteria is a crucial mode of cellular protection and allows bacteria to survive in changing environments. Cellular biomarkers that are quantitatively correlated to stress adaptive behavior will allow to predict the impact of changing environments on bacterial fitness, robustness and survival. The availability of complete genome sequences of a wide variety of bacteria has been instrumental in the development of functional genomics technologies that integrate molecular biology and classical physiology to further understand bacterial stress adaptation mechanisms [27]. These technologies, that make use of holistic and unbiased approaches, can direct our search for crucial cellular components that may function as biomarkers for stress adaptive behavior and subsequent enhanced robustness under challenging conditions. In this study, we identified various transcript biomarkers and also biomarkers at protein and enzyme activity levels, all with predictive quality. We proposed a framework for selecting those cellular biomarkers for mild stress induced enhanced robustness and we systematically evaluated the predictive potential of the candidate-biomarkers for our model-organism B. cereus. Based on a genome-wide comparison of transcriptome profiles of B. cereus cells exposed to four mild stress conditions, several potential candidate-biomarkers were selected. These were the general stress response transcriptional regulator σB, catalases involved in H2O2-scavenging, and chaperones and ATP-dependent Clp proteases involved in protein repair and maintenance. Since several mild stress conditions induced the transcription of genes involved in these stress responses, these findings support their significance in B. cereus general stress response. Activation of cellular mechanisms involved in stress response-signaling and -regulation upon exposure to stress conditions as well as activation of systems involved in oxidative stress defense and maintenance of protein quality might be an essential and general mode of microbial stress adaptation and enhancement of bacterial robustness. Therefore, the canonical induction of these stress responses may extend beyond the species B. cereus, and has been demonstrated for other bacteria including Bacillus subtilis, Listeria monocytogenes and Escherichia coli upon exposure to stress conditions [19], [28], [29], but also in yeast by a wide variety of environmental changes [16], [17]. In the present study, the induction of σB, catalases, chaperones and ATP-dependent Clp proteases were quantitatively determined at transcript, protein and/or activity level upon mild stress treatment and these responses were correlated to mild stress induced enhanced robustness towards lethal stress, aiming to evaluate their predictive biomarker potential at these different functional cell levels. Our study shows that the predictive potential of cellular indicators is highly influenced by the functional cell level at which the indicator is measured. Catalase activity and the SigB protein were identified as long-term biomarkers for various adaptive stress responses. Moreover, the lack of induction of catalase activity in mild heat stress adapted cells corresponded to the lack of enhanced robustness towards lethal oxidative stress, and therefore, it also showed to have predictive potential for this condition where mild stress pretreatment did not provide cross-protection. Both catalase activity and the SigB protein could be employed as convergent- and divergent-application biomarkers as their predictive ability was not restricted to a single mild stress and lethal stress pair. These findings underlined the high predictive quality of these biomarkers and pointed to a promising role in prediction of stress adaptive behavior. In contrast, the catA transcript of the main vegetative catalase was not suitable to predict the robustness status towards lethal stress of mild stress pretreated cells for any of the tested conditions, underpinning the significance to evaluate the predictive potential of cellular indicators at different functional cell levels. The sigB transcript acted as short-term biomarker and predicted the robustness enhancement towards lethal heat stress provided by mild heat stress preexposure for rather short mild heat stress adaptation intervals (up to 15 min). This demonstrated that the predictive potential of biomarkers was manifested across multiple time scales. The better predictive potential of enzyme activity and proteins compared to transcripts might be a reflection of the transient nature of the mRNA levels of some of the selected candidate-biomarkers.
The adaptive traits of bacteria can antagonize food processing strategies that rely on combining mild preservation treatments to control bacterial growth of spoilage and pathogenic bacteria, but are also crucial for various industrial functional food applications. The identification of potential biomarkers for mild stress induced enhanced robustness towards lethal stresses contributes to a better understanding of bacterial stress adaptation mechanisms and can guide our search to control and/or exploit these adaptive traits. The level of robustness enhancement provided by mild stress preexposure was demonstrated to depend on the type and concentration of mild stress applied and the selected lethal stress condition. These different ranges of enhanced (cross-)protection were not taken into consideration in this study when the predictive quality of the candidate-biomarkers was evaluated. The focus of the present study was to design a framework for identification of biomarkers that were able to predict the robustness status of mild stress adapted cells. This can be the basis for further characterizing predictive potential of promising biomarkers by addressing the robustness enhancement ranges provided by mild stress pretreatment. Promising candidate-biomarkers might be species- and genus-specific because the functional conservation of stress-related cellular factors differs among species and genera, and the predictive quality of biomarkers might even be strain specific. The role and regulation of key regulators of general stress responses as well as cellular mechanisms that are crucial for controlling protein quality and defending against oxidative stresses have been shown to differ between closely-related genera. The transcriptional regulator σB functions as central regulator of general stress responses in Gram-positive bacteria with a low GC content including the genera Bacillus, Staphylococcus and Listeria with variations in regulon-members and -size [19], [23], [24], [30]–[35]. This sigma factor is absent in various lactic acid bacteria [10], [36], [37] suggesting that lactic acid bacteria have developed different stress regulatory networks. Clp proteases and chaperones and the main vegetative catalase, predominantly controlled by the CtsR and/or HrcA repressors [38], [39] and by PerR [40]–[43] in low GC Gram-positive bacteria, respectively, are widely conserved in microorganisms [39], [44], [45] and play indispensable roles in cellular repair and defense strategies. Despite apparent variations in their mechanisms of expression control between species and genera, these canonical stress-related components are among the most consistently induced components in microbial stress responses and ubiquitous in biology. We showed that these stress indicators can serve as predictors of stress adaptive behavior in B. cereus and it is conceivable that they may also be employed in other microorganisms. Variations in functional conservations and regulatory circuits necessitates a profound validation of biomarker quality, and our study provides a quantitative approach to systematically search for these biomarkers and to evaluate their predictive potential.
In conclusion, we presented a framework for identifying cellular biomarkers for stress adaptive behavior and to evaluate their predictive potential at transcript, protein and activity level. This quantitative approach opens avenues towards prediction of microbial performance using cellular biomarkers which can serve to early detect and control adaptive behavior that results in enhanced robustness.
Materials and Methods
Bacterial strain and preculturing conditions
Bacillus cereus ATCC 14579 was used as model-organism throughout the study. Stock cultures grown in brain heart infusion (BHI, Becton Dickinson, France) broth were stored at −80°C in 25% (v/v) glycerol. To prepare precultures, 10 ml BHI broth was inoculated with a droplet of the stock culture and incubated overnight at 30°C with shaking at 200 rpm.
Mild stress treatment
The precultures were inoculated in Erlenmeyer flasks containing 50 ml fresh BHI broth and incubated at 30°C with shaking at 200 rpm until the cells were exponentially growing (absorbance value at 600 nm of 0.4 to 0.5; unstressed, reference condition). Upon reaching this optical density, the exponentially growing cells were treated with four different mild stresses for 2, 5, 10, 15, 20, 30 and 60 min with shaking at 200 rpm. The following mild stress conditions were applied: heat stress (43°C); acid-shock (pH 5.5, adjusted with 37% hydrochloric acid, at 30°C); osmotic-upshift (1.5% [w/v] sodium chloride, at 30°C); and oxidative stress (0.1 mM H2O2, at 30°C). Preliminary experiments had demonstrated that preexposure for 15 min to those selected conditions resulted in optimal heat resistance (Figure S1 in the supplementary material).
Determination of lethal stress robustness following mild stress pretreatment
Both unstressed and mild stress treated cells were subsequently exposed to three lethal stress conditions to determine their specific robustness using the inactivation procedure described previously [21]. The following inactivation conditions were chosen: heat stress (50°C); low pH (pH 3.3, adjusted with 37% hydrochloric acid, at 30°C); and oxidative stress (0.2 mM H2O2, at 30°C). Before and after inactivation treatment, samples were taken and decimal dilutions were made in peptone saline solution (1 g neutralized bacteriological peptone [Oxoid, United Kingdom] supplemented with 8.5 g sodium chloride per liter). After acid-inactivation treatment, samples were decimally diluted in BHI broth to ensure no further acid-inactivation during diluting. The appropriate dilutions were surface plated, in duplicate, on BHI agar plates (BHI broth supplemented with 15 g agar [Oxoid, United Kingdom] per liter) using a spiral plater (Eddy Jet; IUL Instruments, Spain) and the plates were incubated at 30°C for 16 to 24 h. The experiments to inactivate unstressed and mild stress pretreated cells were reproduced 2 to 3 times on different days. The robustness of both unstressed and mild stress pretreated cells was determined as the number of microorganism surviving the inactivation treatment, N(t), compared to the initial number of microorganism before the inactivation treatment at t = 0, N(0). t-Tests were performed to compare the log10-robustness of mild stress pretreated cells to that of unstressed cells (with P<0.05 as significance threshold).
Determination of candidate-biomarker induction upon mild stress treatment
Ten candidate-biomarkers were quantitatively measured before and after mild stress treatment for 2 to 60 min, namely, catalase activity, the proteins SigB, ClpC and ClpP, and the transcripts sigB, catA, catE, clpB, clpC, clpP. The experimental procedures followed to measure and to quantify the responses of these candidate-biomarkers upon mild stress treatment are described below.
Catalase activity assay
A previously described procedure was used to determine the catalase activity of unstressed cells and mild stress treated cells [21], [26]. Briefly, cells were washed in phosphate-buffered saline and subsequently exposed to hydrogen peroxide, and the decrease in absorbance at 240 nm was measured over time at 30°C with a spectrophotometer (Spectramax Plus 384; Molecular Devices, USA). One unit of catalase activity was defined as a decrease in absorbance at 240 nm of 1 unit per minute. The rate of decrease for each sample was corrected for the amount of cells used in the assay and standardized to absorbance value of 0.5 at 600 nm. For all mild stress treatment intervals, three biologically independent catalase activity experiments were performed.
The catalase activity was also visualized by catalase activity staining on a native polyacrylamide gel. For that, total proteins were extracted from unstressed and mild stress treated cultures following a similar procedure as previously described [15], [26]. Subsequently, fifty micrograms of protein extracts were separated on a native 10% Tris-HCl polyacrylamide gel (Criterion; Bio-Rad Laboratories, USA). Catalase activity was visualized as described previously [46], which results in yellow catalase bands against a dark-green background.
Western blotting
Total cellular protein was extracted from four biologically independent cultures of unstressed and mild stress treated cells. Forty micrograms of protein extracts were separated by using 15% Tris-HCl gels for SigB and ClpP, and 7.5% Tris-HCl gels for ClpC. Immunoblotting was performed as described previously [47] with anti-SigB antibodies raised against the SigB protein of B. cereus [47], and anti-ClpC and anti-ClpP antibodies raised against these proteins of B. subtilis [15]. Immunocomplexes were visualized using Chemiluminescent Peroxidase Substrate-3 (Sigma-Aldrich, Germany) and scanned with a Chemiluminescence scanner with ChemiDoc XRS software (Bio-Rad Laboratories, USA). The band intensity was quantified using Quantity One software (version 4.6.1; Bio-Rad Laboratories, USA) with background subtraction.
RNA isolation and RT-PCR
RNA was isolated from two biologically independent cultures of unstressed and mild stress treated cells. For that, 10 ml of culture was transferred into a 50-ml Falcon tube, and spun down at 13,000×g for 30 s at 4°C. After the supernatant was decanted, the cell pellets were immediately resuspended in 1 ml of TRI-reagent (Ambion, UK) and snap-frozen in liquid nitrogen. The RNA was further extracted as described previously [47]. Synthesis of cDNA and real time(RT)-PCR was carried out as described previously [48], with 16S-rRNA as reference gene, and sigB, catA, catE, clpB, clpC and clpP as target genes. The primers of the reference and target genes are listed in Table S4 in the supplementary material. The relative expression ratios of the target genes were calculated as previously described [49].
Evaluation of candidate-biomarker induction
The relative induction levels of the candidate-biomarkers in mild stress treated cells compared to unstressed cells were log10-transformed and averaged for the biologically independent reproductions for each mild stress treatment time point. t-Tests were performed to evaluate whether the induction of the candidate-biomarker in mild stress treated cells was statistically significant (with P<0.05 as significance threshold).
Correlation between mild stress induced robustness and candidate-biomarkers responses
The robustness of mild stress pretreated cells (s) was also relatively expressed to that of unstressed cultures (uns) and subsequently log10-transformed, 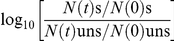 , and averaged for the biological reproductions. Then, for each candidate-biomarker, the mild stress induced candidate-biomarker responses were quantitatively correlated to mild stress induced robustness towards lethal stress per mild stress and lethal stress pair. The Pearson correlation coefficient r was calculated to test the significance of each correlation using PASW software (version 17.0.3).
, and averaged for the biological reproductions. Then, for each candidate-biomarker, the mild stress induced candidate-biomarker responses were quantitatively correlated to mild stress induced robustness towards lethal stress per mild stress and lethal stress pair. The Pearson correlation coefficient r was calculated to test the significance of each correlation using PASW software (version 17.0.3).
Supporting Information
Selection of mild stress condition for mild stress adaptation experiments. Inactivation kinetics of Bacillus cereus ATCC 14579 cells at 50°C following mild stress pretreatment for 15 min. Four mild stress conditions were used for pretreatment, namely, heat stress (a), acid-shock (b), osmotic-upshift (c) and oxidative stress (d), respectively. Pretreatment with 43°C (a), pH 5.5 (b), 1.5% NaCl (c) and 0.1 mM H2O2 (d) resulted in subsequent optimal heat resistance.
(1.00 MB EPS)
Protein ClpC as potential biomarker for mild stress induced enhanced robustness. Induction of protein ClpC upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of ClpC and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which ClpC was qualified as biomarker for mild stress induced enhanced robustness: gray box represents a short-term biomarker; dashed gray box represents a no response condition; double black box represents a convergent-application biomarker. Short-term biomarkers are only correlated to robustness for short-term mild stress adaptation intervals, excluding long-term intervals (black filled symbol).
(1.10 MB EPS)
Protein ClpP as potential biomarker for mild stress induced enhanced robustness. Induction of protein ClpP upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of ClpP and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which ClpP was qualified as biomarker for mild stress induced enhanced robustness: gray box represents a short-term biomarker; dashed gray box represents a no response condition; double black box represents a convergent-application biomarker. Short-term biomarkers are only correlated to robustness for short-term mild stress adaptation intervals, excluding long-term intervals (black filled symbol).
(1.10 MB EPS)
Transcript clpC as potential biomarker for mild stress induced enhanced robustness. Induction of transcript clpC upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of clpC and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which clpC was qualified as biomarker for mild stress induced enhanced robustness: black box represents a long-term biomarker; gray box represents a short-term biomarker; dashed gray box represents a no response condition; double black box represents a convergent-application biomarker; double gray box represents a divergent-application biomarker. Short-term biomarkers are only correlated to robustness for short-term mild stress adaptation intervals, excluding long-term intervals (black filled symbol).
(1.12 MB EPS)
Transcript clpP as potential biomarker for mild stress induced enhanced robustness. Induction of transcript clpP upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of clpP and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which clpP was qualified as biomarker for mild stress induced enhanced robustness: black box represents a long-term biomarker; gray box represents a short-term biomarker; dashed gray box represents a no response condition; double black box represents a convergent-application biomarker; double gray box represents a divergent-application biomarker. Short-term biomarkers are only correlated to robustness for short-term mild stress adaptation intervals, excluding long-term intervals (black filled symbol).
(1.12 MB EPS)
Transcript catE as potential biomarker for mild stress induced enhanced robustness. Induction of transcript catE upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of catE and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which catE was qualified as biomarker for mild stress induced enhanced robustness: black box represents a long-term biomarker; gray box represents a short-term biomarker; double gray box represents a divergent-application biomarker. Short-term biomarkers are only correlated to robustness for short-term mild stress adaptation intervals, excluding long-term intervals (black filled symbols).
(0.98 MB EPS)
Transcript sigB as potential biomarker for mild stress induced enhanced robustness. Induction of transcript sigB upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of sigB and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which sigB was qualified as biomarker for mild stress induced enhanced robustness: black box represents a long-term biomarker; gray box represents a short-term biomarker. Short-term biomarkers are only correlated to robustness for short-term mild stress adaptation intervals, excluding long-term intervals (black filled symbols).
(1.07 MB EPS)
Transcript catA as potential biomarker for mild stress induced enhanced robustness. Induction of transcript catA upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of catA and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. For non of the mild stress and lethal stress pairs, catA was qualified as biomarker.
(1.06 MB EPS)
Transcript clpB as potential biomarker for mild stress induced enhanced robustness. Induction of transcript clpB upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of clpB and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which clpB was qualified as biomarker for mild stress induced enhanced robustness: black box represents a long-term biomarker; dashed gray box represents a no response condition.
(1.06 MB EPS)
Overlap of transcriptome responses upon mild stress treatment
(0.07 MB DOC)
Mild stress induced (cross-)protection towards lethal stress
(0.03 MB DOC)
Biomarkers for mild stress induced enhanced robustness towards lethal stress
(0.06 MB DOC)
RT-PCR primers used in this study
(0.03 MB DOC)
Acknowledgments
The authors would like to thank Michiel Kleerebezem (NIZO Food Research, The Netherlands) for his valuable comments and critical reading of this manuscript.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: The authors have no support or funding to report.
References
- 1.Mitchell A, Romano GH, Groisman B, Yona A, Dekel E, et al. Adaptive prediction of environmental changes by microorganisms. Nature. 2009;460:220–224. doi: 10.1038/nature08112. [DOI] [PubMed] [Google Scholar]
- 2.Wolf DM, Fontaine-Bodin L, Bischofs I, Price G, Keasling J, et al. Memory in microbes: quantifying history-dependent behavior in a bacterium. PLoS ONE. 2008;3:e1700. doi: 10.1371/journal.pone.0001700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Leistner L, Gorris LGM. Food preservation by hurdle technology. Trends Food Sci Technol. 1995;6:41–46. [Google Scholar]
- 4.Abee T, Wouters JA. Microbial stress response in minimal processing. Int J Food Microbiol. 1999;50:65–91. doi: 10.1016/s0168-1605(99)00078-1. [DOI] [PubMed] [Google Scholar]
- 5.Hill C, Cotter PD, Sleator RD, Gahan CGM. Bacterial stress response in Listeria monocytogenes: jumping the hurdles imposed by minimal processing. Int Dairy J. 2002;12:273–283. [Google Scholar]
- 6.Skandamis PN, Yoon Y, Stopforth JD, Kendall PA, Sofos JN. Heat and acid tolerance of Listeria monocytogenes after exposure to single and multiple sublethal stresses. Food Microbiol. 2008;25:294–303. doi: 10.1016/j.fm.2007.10.008. [DOI] [PubMed] [Google Scholar]
- 7.Cotter PD, Hill C. Surviving the acid test: responses of Gram-positive bacteria to low pH. Microbiol Mol Biol Rev. 2003;67:429–453. doi: 10.1128/MMBR.67.3.429-453.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gahan CGM, Hill C. The relationship between acid stress responses and virulence in Salmonella typhimurium and Listeria monocytogenes. Int J Food Microbiol. 1999;50:93–100. doi: 10.1016/s0168-1605(99)00079-3. [DOI] [PubMed] [Google Scholar]
- 9.Serrazanetti DI, Guerzoni ME, Corsetti A, Vogel R. Metabolic impact and potential exploitation of the stress reactions in lactobacilli. Food Microbiol. 2009;26:700–711. doi: 10.1016/j.fm.2009.07.007. [DOI] [PubMed] [Google Scholar]
- 10.Van de Guchte M, Serror P, Chervaux C, Smokvina T, Ehrlich SD, et al. Stress responses in lactic acid bacteria. Antonie Van Leeuwenhoek. 2002;82:187–216. [PubMed] [Google Scholar]
- 11.Culligan E, Hill C, Sleator RD. Probiotics and gastrointestinal disease: successes, problems and future prospects. Gut Pathog. 2009;1:19. doi: 10.1186/1757-4749-1-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Browne N, Dowds BCA. Heat and salt stress in the food pathogen Bacillus cereus. J Appl Microbiol. 2001;91:1085–1094. doi: 10.1046/j.1365-2672.2001.01478.x. [DOI] [PubMed] [Google Scholar]
- 13.Browne N, Dowds BCA. Acid stress in the food pathogen Bacillus cereus. J Appl Microbiol. 2002;92:404–414. doi: 10.1046/j.1365-2672.2002.01541.x. [DOI] [PubMed] [Google Scholar]
- 14.Den Besten HMW, Mataragas M, Moezelaar R, Abee T, Zwietering MH. Quantification of the effects of salt stress and physiological state on thermotolerance of Bacillus cereus ATCC 10987 and ATCC 14579. Appl Environ Microbiol. 2006;72:5884–5894. doi: 10.1128/AEM.00780-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Periago PM, van Schaik W, Abee T, Wouters JA. Identification of proteins involved in the heat stress response of Bacillus cereus ATCC 14579. Appl Environ Microbiol. 2002;68:3486–3495. doi: 10.1128/AEM.68.7.3486-3495.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Causton HC, Ren B, Koh SS, Harbison CT, Kanin E, et al. Remodeling of yeast genome expression in response to environmental changes. Mol Biol Cell. 2001;12:323–337. doi: 10.1091/mbc.12.2.323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gasch AP, Spellman PT, Kao CM, Carmel-Harel O, Eisen MB, et al. Genomic expression programs in the response of yeast cells to environmental changes. Mol Biol Cell. 2000;11:4241–4257. doi: 10.1091/mbc.11.12.4241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gunasekera TS, Csonka LN, Paliy O. Genome-wide transcriptional responses of Escherichia coli K-12 to continuous osmotic and heat stresses. J Bacteriol. 2008;190:3712–3720. doi: 10.1128/JB.01990-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Petersohn A, Brigulla M, Haas S, Hoheisel JD, Völker U, et al. Global analysis of the general stress response of Bacillus subtilis. J Bacteriol. 2001;183:5617–5631. doi: 10.1128/JB.183.19.5617-5631.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ceragioli M, Mols M, Moezelaar R, Ghelardi E, Senesi S, et al. Comparative transcriptome and phenotype analysis of Bacillus cereus in response to disinfectant treatments. Appl Environ Microbiol. 2010;76:3352–3360. doi: 10.1128/AEM.03003-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Den Besten HMW, Mols M, Moezelaar R, Zwietering MH, Abee T. Phenotypic and transcriptomic analyses of mildly and severely salt-stressed Bacillus cereus ATCC 14579 cells. Appl Environ Microbiol. 2009;75:4111–4119. doi: 10.1128/AEM.02891-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mols M, van Kranenburg R, Tempelaars MH, van Schaik W, Moezelaar R, et al. Comparative analysis of transcriptional and physiological responses of Bacillus cereus to organic and inorganic acid shocks. Int J Food Microbiol. 2010;137:13–21. doi: 10.1016/j.ijfoodmicro.2009.09.027. [DOI] [PubMed] [Google Scholar]
- 23.Van Schaik W, van der Voort M, Molenaar D, Moezelaar R, de Vos WM, et al. Identification of the σB regulon of Bacillus cereus and conservation of σB-regulated genes in low-GC-content Gram-positive bacteria. J Bacteriol. 2007;189:4384–4390. doi: 10.1128/JB.00313-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.De Been M, Tempelaars MH, van Schaik W, Moezelaar R, Siezen RJ, et al. A novel hybrid kinase is essential for regulating the σB-mediated stress response of Bacillus cereus. Environ Microbiol. 2010;12:730–745. doi: 10.1111/j.1462-2920.2009.02116.x. [DOI] [PubMed] [Google Scholar]
- 25.Koburger T, Weibezahn J, Bernhardt J, Homuth G, Hecker M. Genome-wide mRNA profiling in glucose starved Bacillus subtilis cells. Mol Genet Genomics. 2005;274:1–12. doi: 10.1007/s00438-005-1119-8. [DOI] [PubMed] [Google Scholar]
- 26.Van Schaik W, Zwietering MH, de Vos WM, Abee T. Deletion of the sigB gene in Bacillus cereus ATCC 14579 leads to hydrogen peroxide hyperresistance. Appl Environ Microbiol. 2005;71:6427–6430. doi: 10.1128/AEM.71.10.6427-6430.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Brul S, Schuren F, Montijn R, Keijser BJF, van der Spek H, et al. The impact of functional genomics on microbiological food quality and safety. Int J Food Microbiol. 2006;112:195–199. doi: 10.1016/j.ijfoodmicro.2006.04.014. [DOI] [PubMed] [Google Scholar]
- 28.Allen KJ, Lepp D, McKellar RC, Griffiths MW. Examination of stress and virulence gene expression in Escherichia coli O157:H7 using targeted microarray analysis. Foodborne Pathog Dis. 2008;5:437–447. doi: 10.1089/fpd.2008.0100. [DOI] [PubMed] [Google Scholar]
- 29.Van der Veen S, Hain T, Wouters JA, Hossain H, de Vos WM, et al. The heat-shock response of Listeria monocytogenes comprises genes involved in heat shock, cell division, cell wall synthesis, and the SOS response. Microbiology. 2007;153:3593–3607. doi: 10.1099/mic.0.2007/006361-0. [DOI] [PubMed] [Google Scholar]
- 30.Bischoff M, Dunman P, Kormanec J, Macapagal D, Murphy E, et al. Microarray-based analysis of the Staphylococcus aureus σB Regulon. J Bacteriol. 2004;186:4085–4099. doi: 10.1128/JB.186.13.4085-4099.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hain T, Hossain H, Chatterjee S, Machata S, Volk U, et al. Temporal transcriptomic analysis of the Listeria monocytogenes EGD-e σB regulon. BMC Microbiol. 2008;8:20. doi: 10.1186/1471-2180-8-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hecker M, Pané-Farré J, Völker U. SigB-dependent general stress response in Bacillus subtilis and related Gram-positive bacteria. Annu Rev Microbiol. 2007;61:215–236. doi: 10.1146/annurev.micro.61.080706.093445. [DOI] [PubMed] [Google Scholar]
- 33.Kazmierczak MJ, Mithoe SC, Boor KJ, Wiedmann M. Listeria monocytogenes σB regulates stress response and virulence functions. J Bacteriol. 2003;185:5722–5734. doi: 10.1128/JB.185.19.5722-5734.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Pané-Farré J, Jonas B, Förstner K, Engelmann S, Hecker M. The σB regulon in Staphylococcus aureus and its regulation. Int J Med Microbiol. 2006;296:237–258. doi: 10.1016/j.ijmm.2005.11.011. [DOI] [PubMed] [Google Scholar]
- 35.Price CW, Fawcett P, Cérémonie H, Su N, Murphy CK, et al. Genome-wide analysis of the general stress response in Bacillus subtilis. Mol Microbiol. 2001;41:757–774. doi: 10.1046/j.1365-2958.2001.02534.x. [DOI] [PubMed] [Google Scholar]
- 36.Hols P, Hancy F, Fontaine L, Grossiord B, Prozzi D, et al. New insights in the molecular biology and physiology of Streptococcus thermophilus revealed by comparative genomics. FEMS Microbiol Rev. 2005;29:435–463. doi: 10.1016/j.femsre.2005.04.008. [DOI] [PubMed] [Google Scholar]
- 37.Van Schaik W, Abee T. The role of σB in the stress response of Gram-positive bacteria - targets for food preservation and safety. Curr Opin Biotechnol. 2005;16:218–224. doi: 10.1016/j.copbio.2005.01.008. [DOI] [PubMed] [Google Scholar]
- 38.Derré I, Rapoport G, Msadek T. CtsR, a novel regulator of stress and heat shock response, controls clp and molecular chaperone gene expression in Gram-positive bacteria. Mol Microbiol. 1999;31:117–131. doi: 10.1046/j.1365-2958.1999.01152.x. [DOI] [PubMed] [Google Scholar]
- 39.Frees D, Savijoki K, Varmanen P, Ingmer H. Clp ATPases and ClpP proteolytic complexes regulate vital biological processes in low GC, Gram-positive bacteria. Mol Microbiol. 2007;63:1285–1295. doi: 10.1111/j.1365-2958.2007.05598.x. [DOI] [PubMed] [Google Scholar]
- 40.Bsat N, Herbig A, Casillas-Martinez L, Setlow P, Helmann JD. Bacillus subtilis contains multiple Fur homologues: identification of the iron uptake (Fur) and peroxide regulon (PerR) repressors. Mol Microbiol. 1998;29:189–198. doi: 10.1046/j.1365-2958.1998.00921.x. [DOI] [PubMed] [Google Scholar]
- 41.Herbig AF, Helmann JD. Roles of metal ions and hydrogen peroxide in modulating the interaction of the Bacillus subtilis PerR peroxide regulon repressor with operator DNA. Mol Microbiol. 2001;41:849–859. doi: 10.1046/j.1365-2958.2001.02543.x. [DOI] [PubMed] [Google Scholar]
- 42.Horsburgh MJ, Clements MO, Crossley H, Ingham E, Foster SJ. PerR controls oxidative stress resistance and iron storage proteins and is required for virulence in Staphylococcus aureus. Infect Immun. 2001;69:3744–3754. doi: 10.1128/IAI.69.6.3744-3754.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Rea R, Hill C, Gahan CGM. Listeria monocytogenes PerR mutants display a small-colony phenotype, increased sensitivity to hydrogen peroxide, and significantly reduced murine virulence. Appl Environ Microbiol. 2005;71:8314–8322. doi: 10.1128/AEM.71.12.8314-8322.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Estruch F. Stress-controlled transcription factors, stress-induced genes and stress tolerance in budding yeast. FEMS Microbiol Rev. 2000;24:469–486. doi: 10.1111/j.1574-6976.2000.tb00551.x. [DOI] [PubMed] [Google Scholar]
- 45.Imlay JA. Cellular defenses against superoxide and hydrogen peroxide. Annu Rev Biochem. 2008;77:755–776. doi: 10.1146/annurev.biochem.77.061606.161055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Woodbury W, Spencer AK, Stahmann MA. An improved procedure using ferricyanide for detecting catalase isozymes. Anal Biochem. 1971;44:301–305. doi: 10.1016/0003-2697(71)90375-7. [DOI] [PubMed] [Google Scholar]
- 47.Van Schaik W, Tempelaars MH, Wouters JA, de Vos WM, Abee T. The alternative sigma factor σB of Bacillus cereus: response to stress and role in heat adaptation. J Bacteriol. 2004;186:316–325. doi: 10.1128/JB.186.2.316-325.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Van Schaik W, Tempelaars MH, Zwietering MH, de Vos WM, Abee T. Analysis of the role of RsbV, RsbW, and RsbY in regulating σB activity in Bacillus cereus. J Bacteriol. 2005;187:5846–5851. doi: 10.1128/JB.187.16.5846-5851.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucl Acids Res. 2001;29:e45. doi: 10.1093/nar/29.9.e45. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Selection of mild stress condition for mild stress adaptation experiments. Inactivation kinetics of Bacillus cereus ATCC 14579 cells at 50°C following mild stress pretreatment for 15 min. Four mild stress conditions were used for pretreatment, namely, heat stress (a), acid-shock (b), osmotic-upshift (c) and oxidative stress (d), respectively. Pretreatment with 43°C (a), pH 5.5 (b), 1.5% NaCl (c) and 0.1 mM H2O2 (d) resulted in subsequent optimal heat resistance.
(1.00 MB EPS)
Protein ClpC as potential biomarker for mild stress induced enhanced robustness. Induction of protein ClpC upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of ClpC and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which ClpC was qualified as biomarker for mild stress induced enhanced robustness: gray box represents a short-term biomarker; dashed gray box represents a no response condition; double black box represents a convergent-application biomarker. Short-term biomarkers are only correlated to robustness for short-term mild stress adaptation intervals, excluding long-term intervals (black filled symbol).
(1.10 MB EPS)
Protein ClpP as potential biomarker for mild stress induced enhanced robustness. Induction of protein ClpP upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of ClpP and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which ClpP was qualified as biomarker for mild stress induced enhanced robustness: gray box represents a short-term biomarker; dashed gray box represents a no response condition; double black box represents a convergent-application biomarker. Short-term biomarkers are only correlated to robustness for short-term mild stress adaptation intervals, excluding long-term intervals (black filled symbol).
(1.10 MB EPS)
Transcript clpC as potential biomarker for mild stress induced enhanced robustness. Induction of transcript clpC upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of clpC and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which clpC was qualified as biomarker for mild stress induced enhanced robustness: black box represents a long-term biomarker; gray box represents a short-term biomarker; dashed gray box represents a no response condition; double black box represents a convergent-application biomarker; double gray box represents a divergent-application biomarker. Short-term biomarkers are only correlated to robustness for short-term mild stress adaptation intervals, excluding long-term intervals (black filled symbol).
(1.12 MB EPS)
Transcript clpP as potential biomarker for mild stress induced enhanced robustness. Induction of transcript clpP upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of clpP and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which clpP was qualified as biomarker for mild stress induced enhanced robustness: black box represents a long-term biomarker; gray box represents a short-term biomarker; dashed gray box represents a no response condition; double black box represents a convergent-application biomarker; double gray box represents a divergent-application biomarker. Short-term biomarkers are only correlated to robustness for short-term mild stress adaptation intervals, excluding long-term intervals (black filled symbol).
(1.12 MB EPS)
Transcript catE as potential biomarker for mild stress induced enhanced robustness. Induction of transcript catE upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of catE and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which catE was qualified as biomarker for mild stress induced enhanced robustness: black box represents a long-term biomarker; gray box represents a short-term biomarker; double gray box represents a divergent-application biomarker. Short-term biomarkers are only correlated to robustness for short-term mild stress adaptation intervals, excluding long-term intervals (black filled symbols).
(0.98 MB EPS)
Transcript sigB as potential biomarker for mild stress induced enhanced robustness. Induction of transcript sigB upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of sigB and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which sigB was qualified as biomarker for mild stress induced enhanced robustness: black box represents a long-term biomarker; gray box represents a short-term biomarker. Short-term biomarkers are only correlated to robustness for short-term mild stress adaptation intervals, excluding long-term intervals (black filled symbols).
(1.07 MB EPS)
Transcript catA as potential biomarker for mild stress induced enhanced robustness. Induction of transcript catA upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of catA and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. For non of the mild stress and lethal stress pairs, catA was qualified as biomarker.
(1.06 MB EPS)
Transcript clpB as potential biomarker for mild stress induced enhanced robustness. Induction of transcript clpB upon mild stress treatment (43°C, pH 5.5, 1.5% NaCl, 0.1 mM H2O2) was correlated to mild stress induced enhanced robustness towards lethal stress (50°C, open symbols; pH 3.3, light gray filled symbols; 0.2 mM H2O2, dark gray filled symbols). Each mild stress was applied for 2, 5, 10, 15, 20, 30 and 60 min and induction of clpB and robustness for mild stress pretreated cells (s) was relatively expressed to that of unstressed cells (uns) (log s/uns). Robustness of mild stress pretreated cells and unstressed cells was determined as the number of bacteria surviving the lethal stress treatment compared to the initial number of bacteria. The graph boxes show the conditions for which clpB was qualified as biomarker for mild stress induced enhanced robustness: black box represents a long-term biomarker; dashed gray box represents a no response condition.
(1.06 MB EPS)
Overlap of transcriptome responses upon mild stress treatment
(0.07 MB DOC)
Mild stress induced (cross-)protection towards lethal stress
(0.03 MB DOC)
Biomarkers for mild stress induced enhanced robustness towards lethal stress
(0.06 MB DOC)
RT-PCR primers used in this study
(0.03 MB DOC)