


Abstract
Replication protein A (RPA), the nuclear single-stranded DNA binding protein is involved in DNA replication, nucleotide excision repair (NER) and homologous recombination. It is a stable heterotrimer consisting of subunits with molecular masses of 70, 32 and 14 kDa (p70, p32 and p14, respectively). Gapped DNA structures are common intermediates during DNA replication and NER. To analyze the interaction of RPA and its subunits with gapped DNA we designed structures containing 9 and 30 nucleotide gaps with a photoreactive arylazido group at the 3′-end of the upstream oligonucleotide or at the 5′-end of the downstream oligonucleotide. UV crosslinking and subsequent analysis showed that the p70 subunit mainly interacts with the 5′-end of DNA irrespective of DNA structure, while the subunit orientation towards the 3′-end of DNA in the gap structures strongly depends on the gap size. The results are compared with the data obtained previously with the primer–template systems containing 5′- or 3′-protruding DNA strands. Our results suggest a model of polar RPA binding to the gapped DNA.
INTRODUCTION
The human single-stranded DNA (ssDNA) binding protein, replication protein A (RPA), is a stable heterotrimer consisting of subunits with molecular masses of 70, 32 and 14 kDa (p70, p32 and p14, respectively). This protein has multiple functions in various processes of DNA metabolism (reviewed in 1,2). RPA is required during the initiation and elongation stages of DNA replication and it is involved in both nucleotide excision DNA repair (NER) and homologous recombination processes.
The interaction of RPA with ssDNA has been extensively studied (1,2). Photocrosslinking has been used to analyze the interaction of RPA subunits with the primer of a partial duplex DNA (3–6). In these studies the base-substituted photoreactive dNTP analog, 5-[N-(2-nitro-5-azidobenzoyl)-trans-3-aminopropenyl-1]-2′-deoxyuridine-5′-triphosphate (NAB-4-dUTP; Fig. 1A), was used as substrate for elongation of primer–template by DNA polymerases. Subsequent UV irradiation showed crosslinking of p32 to the nascent DNA. In contrast, only limited crosslinking of p70 and no crosslinking of p14 was observed. The data indicate that the p70 subunit binds predominantly to the single-stranded part of partial duplex DNA whereas the p32 subunit is located near and might even contact the 3′-end of the primer (3). Interactions between the p32 subunit of RPA and nascent DNA in replicating SV40 chromosomes have also been observed indicating that the p32 contacts early intermediates produced by DNA polymerase α-primase but not more advanced products (7). In addition, it has been demonstrated in vitro that the pattern of subunit crosslinking was changed when template extension was shortened (4,6). When photoreagents with the template extensions of 14, 13 and 9 nt instead of 19 and 30 nt were used the p70 subunit of RPA was predominantly photocrosslinked to the 3′-end of the photoreactive primer (6).
Figure 1.

Structural formulae of photoreagents. (A) dUTP analog used for incorporation of photoreactive nucleotide analogs into the 3′-end of the primer. (B) Photoreactive group attached to the 5′-end of oligonucleotide. (C) Photoreactive DNA duplexes used in these studies.
In NER, excision of damage produces an intermediate that contains a 30 nt gap. It is thought that RPA transiently binds to this gap until it is filled in by a PCNA-dependent DNA polymerase (8). Gapped intermediates are thought to exist on the lagging strand during Okazaki strand synthesis (9). Therefore, we were interested in studying RPA binding to gapped DNA structures. In the present work we examined the proximity of RPA subunits to both sides of 9 and 30 nt DNA gaps using reactive DNAs with a photoreactive group localized at the 3′- or 5′-end of the gap. We found that the pattern of RPA subunit crosslinking was strongly dependent on the location of the photoreactive group and on the gap size. The data are discussed in terms of recent observations concerning polarity of RPA binding to ssDNA (10,11).
MATERIALS AND METHODS
Proteins and nucleotides
Recombinant DNA polymerase β was purified according to Beard and Wilson (12). Recombinant RPA was isolated as described (13). Rainbow colored protein molecular mass markers and 3000 Ci/mmol [α-32P]dCTP were from Amersham. [α-32P]ATP was from ICN. Synthetic oligonucleotides were obtained from GENSET. Dephosphorylated primers were 5′-phosphorylated with T4 polynucleotide kinase (New England Biolabs) as described (14). Unreacted [γ-32P]ATP was separated by passing the mixture over a Nensorb-20 column (Du Pont) using the manufacturer’s suggested protocol. The photoreactive nucleotide analog NAB-4-dUTP and 5′-end modified oligonucleotide (ON9) were synthesized and characterized essentially as described by Wlassoff et al. (15) and Levina et al. (16), respectively.
Primer–template
The sequences of oligonucleotides used in the experiments were: ON1 5′-CTGCAGCTGATGCGC-3′, ON2 5′-pAGAACTACGATATCGAA-3′, ON3 5′-TTTTTTGGTTCGATATCGTAGTTCTATCTTCATTAGCGCATCAGCTGCGA-3′, ON4 5′-ATC TTCATTAGCGCATCAGCTGCAG-3′, ON5 5′-TTTTTTGGTTCGATATCGTAGTTCTATCTTCATTTATAGCTCCTACCTTTCTCTCAGCGCATCAGCTGCAG-3′, ON6 5′-ATCTTCATTTATAGCTCCTACCTTTCTCTCAGCGCATCAGCTGCAG-3′, ON7 5′-TTTTTTGGTTCGATATCGTAGTTC TATCTTCATT-3′, ON8 5′-TTTTTTGGTTCGATATCGTAGTTCTATCTTCATTTATAGCTCCTACCTTTCTCTC-3′, ON9 5′-R-pAGAACTACGATA TCGAA-3′. R is the photoreactive 2-nitro-5-azidobenzamido-N-propylamino group (Fig. 1B).
The oligonucleotides were mixed in the following combinations: ON1 + ON2 + ON3; ON1 + ON4; ON1 + ON2 + ON5; ON1 + ON6; ON1 + ON9 + ON3; ON9 + ON7; ON1 + ON9 + ON5; ON8 + ON9 at a molar ratio of 1:1 in 10 mM Tris–HCl pH 7.5, 5 mM EDTA. The mixtures were heated at 90°C for 3 min and then allowed to cool slowly to room temperature.
Photochemical crosslinking
Reaction mixtures (10 µl) contained 50 mM Tris–HCl pH 7.8, 10 mM MgCl2, 50 mM KCl, 1 µM DNA polymerase β, 1 µM each of DNA primer–template complexes and 10 µM NAB-4-dUTP, to obtain photoreactive DNAs I–IV (Fig. 1C), 10 µM [α-32P]dCTP to obtain photoreactive DNAs VI, VIII or 10 µM dTTP, 10 µM [α-32P]dCTP to obtain photoreactive DNAs V and VII (Fig. 1C). The reaction mixtures were incubated at 25°C for 30 min to allow elongation of the primers. RPA (0.86 µM) was then added and incubated for 1 min at 25°C. The mixtures were spotted on Parafilm placed on ice followed by UV irradiation (λ = 320 nm) for 20 min with a Bausch and Lomb monochromator equipped with an HBO W super pressure mercury lamp. Reactions were stopped by adding Laemmli loading buffer and heating for 5 min at 100°C. The photochemically crosslinked protein–DNA samples were separated by 10% SDS–PAGE (17). Dried gels were subjected to autoradiography.
RESULTS
The structure of the base-substituted dUTP analog (NAB-4-dUTP) is shown in Figure 1A. This analog has been successfully used previously to examine the arrangement of RPA subunits near the 3′-end of the primer in the complex of RPA with primer–template structures containing 5′-extended template strand (3–6). Photoreactive DNAs I–IV (Fig. 1C) were synthesized by DNA polymerase β in the presence of NAB-4-dUTP starting from synthetic deoxyoligonucleotide duplexes. Conditions described in previous studies (4–6) allowed us to introduce a single photoreactive moiety into the 3′-end of 32P-labeled primer (data not shown).
To obtain photoreactive DNAs V–VIII (Fig. 1C) a synthetic oligonucleotide with photoreactive group at its 5′-end (ON9, for sequence see Materials and Methods) was synthesized according to the protocol of Levina et al. (16), annealed to correspondent template or template–primers and then was elongated by DNA polymerase β in the presence of [α-32P]dCTP to introduce the radioactive label into the oligonucleotide. Analysis on polyacrylamide gels showed that two residues of [32P]dCMP were incorporated into the 3′-end of the primer according to the template structure (data not shown).
After complete primer elongation RPA (0.86 µM) was added to the reaction mixtures containing photoreactive DNAs and then irradiated with UV light (for conditions see Materials and Methods). In the case of the 9 nt gap, (photoreactive DNA I) the 3′-photoreactive primer crosslinks predominantly to the p70 subunit of RPA with less intensive crosslinking of p32 (Fig. 2, lane 2).The crosslinking experiments with gaps of 7 and 13 nt displayed practically the same pattern of subunit labeling (data not shown). In the case of a 9 nt 5′ template extension (photoreactive DNA II), nearly the same pattern of subunit crosslinking was observed. In the case of both the 30 nt gap (photoreactive DNA III) and primer–template complex with 30 nt template extension (IV) the 3′-end reagent predominantly shows crosslinking of the radioactive primer to p32 subunit of RPA (Fig. 2, lanes 4 and 5). The data suggest that in the case of 30 nt single-stranded part of DNA duplex (template extension or gap) p32 either directly interacts with oligonucleotide 3′-end or is located adjacent to it, whereas with the 9 nt single-stranded portion of the partial DNA duplex the p70 subunit was predominantly crosslinked to the 3′-end of DNA irrespective of whether gap or template extension structures were involved. Thus, one can conclude that the orientation of RPA subunits near the 3′-end of DNA is predetermined by the size of the single-stranded portion that causes the changes in conformation during binding of RPA. All reactive DNAs containing a photoreactive group at the 5′-end were found to predominantly crosslink to p70; only slight crosslinking of p32 was observed (Fig. 2, lanes 7–10). Therefore, arrangement of RPA subunits near the 5′-end is similar in both gapped DNA and 3′-template extensions. These data indicate that p70 is consistently located near the 5′-end of gaps but orientation of the p32 subunit near the 3′-end is strongly dependent on RPA conformation. The conformation of RPA in the 30 nt gap is equivalent to p32 being near the 3′-end of the nascent strand during DNA replication (3,6,7). No crosslinking of p14 was observed with any photoreactive DNAs used (data not shown). This suggests that p14 is either not located near 3′- or 5′-ends in a gapped DNA or is not accessible to the photoreactive groups.
Figure 2.
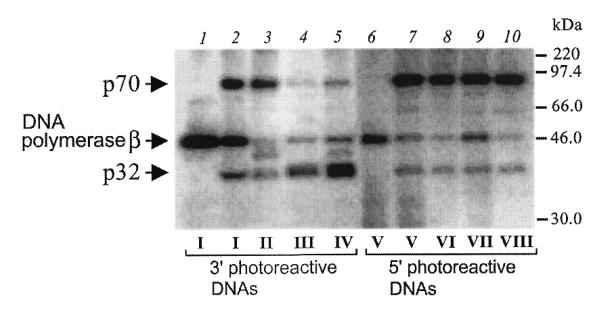
p32 locates near the 3′-end of upstream oligonucleotide whereas p70 locates near the 5′-end of downstream oligonucleotide in DNA gaps. Primer elongation reactions using different template–primer structures (1 µM each) were carried out in the presence of NAB-4-dUTP for reagents I–IV or in the presence [α-32P]dCTP for reagents V–VIII (for conditions see Materials and Methods). After complete primer elongation 0.86 µM RPA was added to the reaction mixtures followed by UV irradiation. The numbers of the photoreagents (I–VIII, lanes 2–5, 7–10) used in each of the reaction mixtures are indicated at the bottom of the figure. Control reactions (lanes 1 and 6) did not contain RPA. The crosslinked protein–DNA complexes were separated by SDS–PAGE and visualized by autoradiography. The positions of crosslinked products and protein markers are indicated on the left and the right margins, respectively. Additional bands in lanes 3 and 5 appear to be products of modification of RPA proteolytic fragments since RPA preparation according to Coomassie staining shows minor bands that appeared during storage (data not shown).
In all experiments, some crosslinking of DNA polymerase β was observed. Addition of RPA reduced the DNA polymerase crosslinking (Fig. 2, compare lane 1 with lanes 2–5, and lane 6 with lanes 7–10). This reduction was less with the 9 nt gap 3′-end reagent (Fig. 2, compare lanes 1, 2 and 3). This finding is consistent with earlier observations that DNA polymerase β has a higher affinity for short gaps than for long gaps (18,19). It has been shown previously that the crosslinking pattern of RPA subunits differed substantially for DNA duplexes with 5′-end protruding template strand when the molar ratio of RPA to partial DNA duplex was higher than 2:1. Under these conditions, intensive crosslinking of both subunits (p70 and p32) was observed independently on the length of DNA template used. This observation was interpreted to indicate that RPA was destabilizing the duplex DNA to produce ssDNA (6). In the present work we examined whether the molar ratio of RPA to DNA gaps could influence RPA subunit crosslinking to photoreactive DNAs. The concentrations of photoreactive DNAs were 1 µM in all experiments. When the RPA concentration increases from 0.21 to 1.72 µM (Fig. 3A and B, lanes 2–5) the intensity of RPA subunit crosslinking is increased for both photoreactive DNAs I and III (Fig. 3A and B, respectively) with change in the relative intensity of p70 and p32 crosslinking in the case of the 30 nt gap. Practically the same tendency was observed in the previous studies with DNA duplexes with the 5′-protruding template strand (31 nt protrusion) (6).
Figure 3.
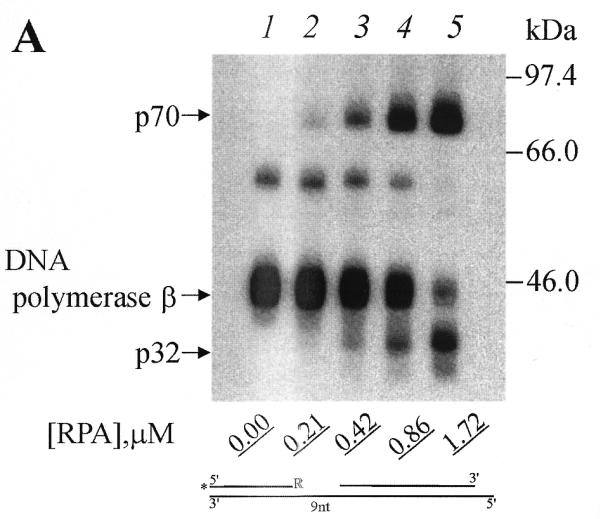
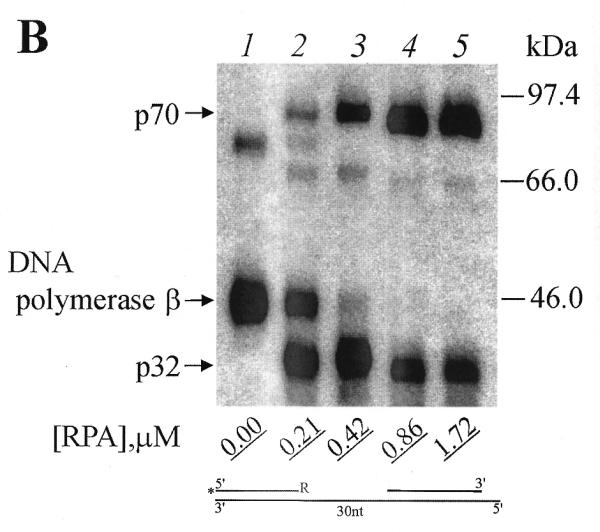
Increasing RPA concentration promotes RPA subunits crosslinking by 3′-photoreagents. (A) Photoreagent I and (B) reagent III were used for crosslinking experiments. After complete photoreagents synthesis (for conditions see Materials and Methods) RPA was added to the reaction mixtures to the indicated (bottom) concentrations followed by irradiation. The UV-crosslinked products were separated by SDS–PAGE and visualized by autoradiography. The positions of crosslinked products and protein markers are indicated in the left and right margins, respectively.
It is interesting to note that the intensity of the primer crosslinking to DNA polymerase β is less influenced by the variation of RPA concentration in the case of the photoreactive DNA I (gap of 9 nt; Fig. 3A, lanes 1–5), whereas crosslinking of DNA polymerase β with photoreactive DNA III is inhibited in the presence of RPA (gap of 30 nt; Fig. 3, compare lanes 1–5). These data are consistent with the previous findings that DNA polymerase β binds with high affinity to short gapped structures (18,19) and that RPA has a lower affinity for short single-stranded oligonucleotides (1,20).
Similar experiments were carried out using photoreagents with 5′-photoreactive gaps. For both photoreactive DNAs V and VII (9 and 30 nt gaps, Fig. 4A and B, respectively) increasing the concentration of RPA from 0.86 to 4.3 µM did not influence RPA subunit crosslinking (Fig. 4A and B, lanes 3–6). A difference was observed only for mixtures with the limited amount of RPA (0.26 µM). In this case, photoreactive DNA V did not crosslink the p70 subunit (Fig. 4A, lane 2), whereas photoreactive DNA VII did crosslink to p70 (Fig. 4B, lane 2). This fact could be attributed to a higher RPA affinity to 30 nt gap structures than to 9 nt gap. These data suggest that different complexes are formed with 9 and 30 nt gaps and that with a 30 nt gap, p70 is in a position which allows more efficient crosslinking to the 5′-end of the adjacent flank oligonucleotide. Similar results were obtained using reagents VI and VIII (data not shown). The crosslinking of DNA polymerase β is again decreased with an increase in RPA concentration (Fig. 4A and B, compare lanes 2–6). However, competition of DNA polymerase β with RPA for the crosslinking with 5′-end photoreactive DNAs in comparison with 3′-end photoreactive DNAs is less effective in the case of short-gap DNA structure (compare Figs 3A and 4A, all lanes). This is probably indicative of a higher affinity RPA interaction followed by its crosslinking to the 5′-end of gapped DNA. Moreover, it is known that DNA polymerase β interacts with the 5′-end phosphate, and this is important for DNA polymerase β affinity to gapped DNA (19). In the case of photoreactive DNAs I–IV, the 5′-end phosphate is not modified, but in the case of DNAs V–VIII, the 5′-end phosphate is modified. This should result in a decrease of DNA polymerase β affinity for gap of the same size but with a modified phosphate. It is interesting to note that as the concentration of RPA increases, the crosslinking of RPA subunits to the 3′ photoreactive group continuously increased, whereas the level of crosslinking to 5′ photoreactive groups remained relatively constant above 0.86 µM RPA (compare Fig. 3A and B, lanes 3–6, with Fig. 4A and B, lanes 3–6). These data are consistent with the earlier observation (11) that RPA contains a strong ssDNA interaction domain at the 5′-side of its DNA binding region and a weak ssDNA-binding domain resides at the 3′-side. Recent observations with fork and overhang DNA substrates demonstrate that RPA initially resides at the 5′-side of a ssDNA substrate and then extends in the 3′-direction to create a stably bound RPA conformation (21).
Figure 4.
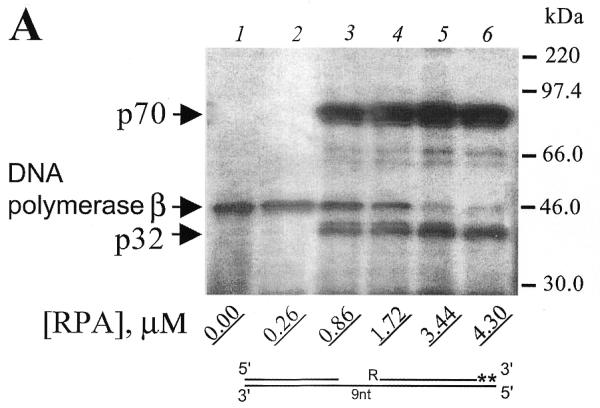
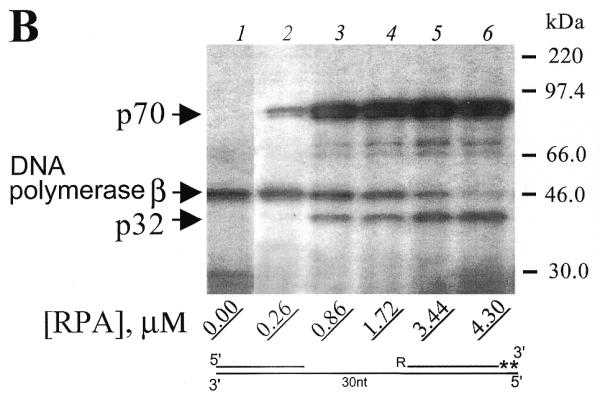
Variation of RPA concentration does not influence RPA crosslinking by 5′-end photoreagents. (A) Photoreagent V and (B) photoreagent VII were used for crosslinking experiments. Reaction mixtures were composed as described in Materials and Methods. After complete primer elongation RPA was added to the reaction mixtures to the indicated (bottom) concentrations followed by UV irradiation. The crosslinked products were separated by SDS–PAGE and visualized by autoradiography. The positions of crosslinked products and protein markers are indicated in the left and right margins, respectively.
DISCUSSION
To unravel the role of RPA in the various processes of DNA metabolism, we have investigated the binding of RPA to DNA structures, which mimic gapped intermediates likely to be found in vivo. We used DNAs containing gaps of 9 and 30 nt with a photoreactive group located either at the 3′-end of the upstream oligonucleotide or 5′-end of the downstream oligonucleotide to identify the subunits of RPA that are located near the two termini.
The major ssDNA binding activity of RPA is associated with p70 (1). The p70 is known to contain three DNA binding domains: two central copies of ssDNA-binding motifs (domains A and B) and a putative (C-X2-C-X15-C-X2-C) zinc finger motif (domain C). The central DNA-binding domains are necessary and sufficient for interactions with ssDNA; however, domain C and part of the N-terminus of the p70 are needed for optimal ssDNA binding activity (22–25). Recently it has been shown that domain C directly interacts with DNA (26,27) and has a preference for binding to damaged DNA (28). The fourth DNA binding domain, domain D, of RPA resides in the p32 subunit (26,29). RPA binds ssDNA with a defined polarity (10,11). In the crystal structure ssDNA binds to the A and B domains in a 5′→3′ direction with the 5′-end associated with domain A and the 3′-end associated with domain B (10). When the full RPA complex binds to DNA there appears to be a strong ssDNA interaction domain of RPA positioned at the 5′-side of its binding region and a weak ssDNA-binding domain resides at the 3′-side (11). It is likely that the strong interactions are caused by domains A and B and that the weak interactions are caused by domains C and possibly D.
The mechanism of ssDNA binding by RPA involves at least two different binding modes, which are best defined by the length of the interacting ssDNA (2,30–34). The first mode, named compact conformation, which is characterized by 8–10 nt occluded binding site, is quasi-stable (31,32). The second mode, named the elongated conformation, which is characterized by 30 nt occluded binding site is stable and seems to be the usual mode of binding DNA. It was proposed that the binding of RPA to ssDNA initially involves an unstable recognition site of 8 nt that subsequently converts to the stable elongated complex. Electron microscopy data have shown that conformational changes of RPA accompanied this process (31,32) When ssDNA binds to domains A and B, the effective concentration of DNA is raised so that the lower affinity sites of domains C and D can bind (2).
In the present work we used two sizes of single-stranded DNA platforms, 9 and 30 nt, to examine RPA subunit arrangements when bound to the gap structures. These interactions were then compared to those found with DNA duplexes with an equivalent extension of the template strand. The ability of RPA subunits to be crosslinked to the 3′ or 5′ photoreactive DNAs indicates their location in the RPA–DNA complex. To explain our results and existing data on RPA binding to DNA, we suggest the model shown in Figure 5. When bound to a 9 nt single-stranded part of template extension or gap using the high affinity A and B DNA binding domains of p70, RPA adopts the compact conformation in which the p32 subunit crosslinking to the 3′-end of upstream oligonucleotide is much less in comparison with crosslinking of p70 subunit. In that case p70 can be crosslinked to the both 3′- and 5′-ends of DNA, therefore suggesting that p70 is located near both ends of gapped DNA (Fig. 5A). The RPA conformation appears to be nearly the same with gap and template extension.
Figure 5.
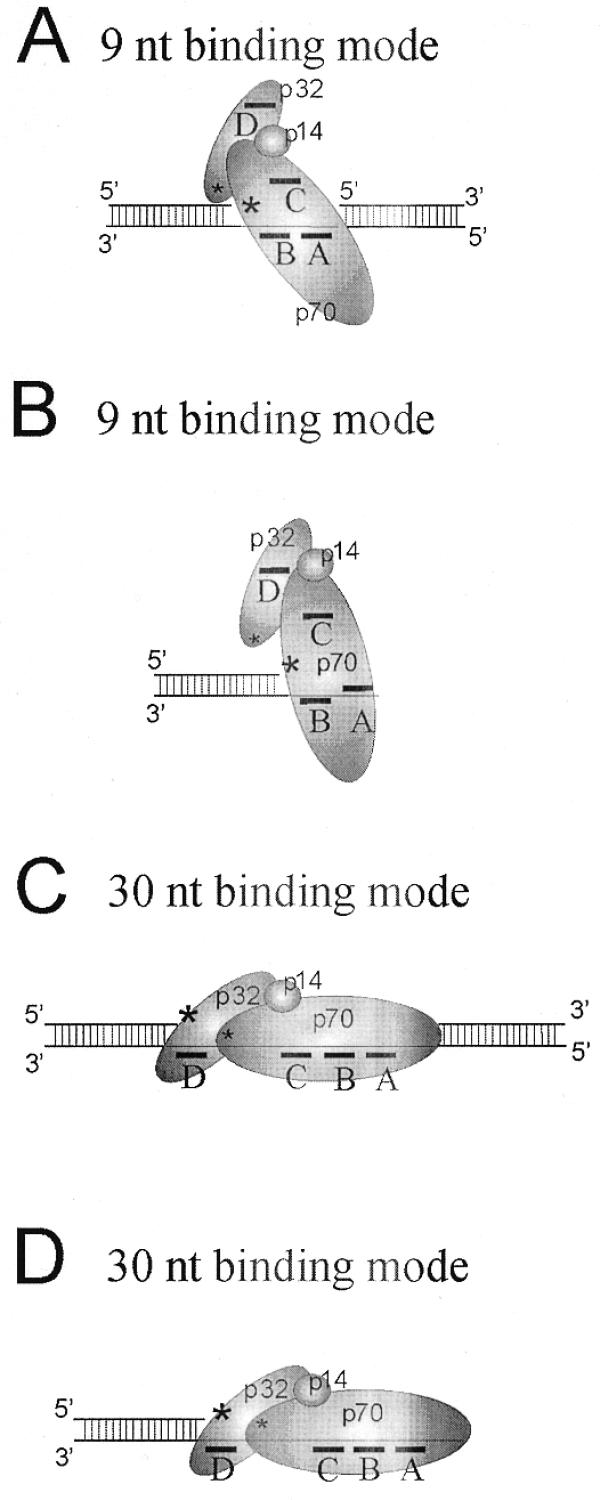
Models of RPA–DNA interactions. Big and small asterisks indicate high and low crosslinking of RPA subunits, respectively. For details see Discussion.
When RPA binds in the stable elongated conformation (Fig. 5C), 30 nt of ssDNA is occluded and p70 is located near the 5′-end of oligonucleotide providing downstream orientation of p32 near the 3′-end of the upstream oligonucleotide. The same subunit orientation can be realized when RPA binds DNA in the stable elongated conformation when 30 nt of ssDNA is occluded by RPA in the case of extended DNA duplex (Fig. 5D). Our data demonstrate that p14 is far from 3′- or 5′-ends of the DNA gap or masked by other subunits. In addition because RPA can be crosslinked to the 3′-end of primer under some conditions these studies suggest that p70 is in close proximity to both ends of the gap, at least transiently.
In summary, the pattern of RPA subunit labeling in DNA structures with gap or template extension is determined by the polar RPA binding to ssDNA (21) when p70 subunit initially resides at the 5′-end of ssDNA platform and then extends in the direction of the 3′-end. Subunit orientation near the 3′-end of DNA structures is predetermined by the size of the single-stranded platform that modulates changes in RPA conformation. These data are in agreement with the earlier observation of RPA interactions with DNA duplexes with the 5′-extended template strands obtained in vitro (6) and in a replicating SV40 chromosome (G.Kaufmann, personal communication).
Polarity of RPA binding to DNA should be very important for RPA function in NER at the stage of damage recognition and following positioning of excision repair nucleases (8). Our observations indicate the orientation of RPA subunits when binding to the gap intermediate formed during the process. It has been suggested by De Laat et al. (11) that the RPA polarity is important for positioning of the excision repair nucleases XPG and EXCC1-XPF on the DNA. With the 3′-oriented side of RPA facing a duplex ssDNA junction, RPA interacts with and stimulates EXCC1-XPF, whereas the 5′-oriented side of RPA at DNA junction allows stable binding of XPG to RPA (11). Based on the proposed model of RPA subunit orientation in DNA gap we could predict interaction of NER nucleases XPG with p70 subunit and ERCC1-XPF with p32 subunit. Therefore, polar orientation of RPA subunits in DNA gap should be critical for the following assembly of NER proteins. The definite mechanism of RPA and its subunits’ interaction with the NER proteins demands further study.
Acknowledgments
ACKNOWLEDGEMENTS
This work was supported in part by an INTAS grant (96-1778), by a grant from the Russian Foundation for Basic Research (98-04-49718, 99-04-04016, 99-04-49277 and 00-04-22002), by grants Convention d’echanges, by grant PICS, a joint international scientific program between CNRS and Russian Academy of Sciences and Ligue Nationale Francaise contre le Cancer to A.F. M.S.W. was supported by grant GM44721 from the National Institutes of Heath General Medicine Institute.
References
- 1.Wold M.S. (1997) Replication protein A: A heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu. Rev. Biochem., 66, 61–62. [DOI] [PubMed] [Google Scholar]
- 2.Iftode C., Daniely,Y. and Borowiec,J.A. (1999) Replication protein A (RPA): the eukaryotic SSB. Crit. Rev. Biochem. Mol. Biol., 34, 141–180. [DOI] [PubMed] [Google Scholar]
- 3.Lavrik O.I., Nasheuer,H.-P., Weisshart,K., Wold,M.S., Prasad,R., Beard,W., Wilson,S.H. and Favre,A. (1998) Subunits of human replication protein A are crosslinked by photoreactive primers synthesized by DNA polymerases. Nucleic Acids Res., 26, 602–607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lavrik O.I., Kolpashchikov,D.M., Nasheuer,H.-P., Weisshart,K. and Favre,A. (1998) Alternative conformations of human replication protein A are detected by crosslinks with primers carrying a photoreactive group at the 3′-end. FEBS Lett., 441, 186–190. [DOI] [PubMed] [Google Scholar]
- 5.Kolpashchikov D.M., Weisshart,K., Nasheuer,H.-P., Khodyreva,S.N., Fanning,E., Favre,A. and Lavrik,O.I. (1999) Interaction of the p70 subunit of RPA with a DNA template directs p32 to the 3′-end of nascent DNA. FEBS Lett., 450, 131–134. [DOI] [PubMed] [Google Scholar]
- 6.Lavrik O.I., Kolpashchikov,D.M., Weisshart,K., Nasheuer,H.-P., Khodyreva,S.N. and Favre,A. (1999) RPA subunit arrangement near the 3′-end of the primer is modulated by the length of the template strand and cooperative protein interactions. Nucleic Acids Res., 27, 4235–4240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mass G., Nethanel,T. and Kaufmann,G. (1998) The middle subunit of replication protein A contacts growing RNA-DNA primers in replicating simian virus 40 chromosomes. Mol. Cell. Biol., 18, 6399–6407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wood R.D. (1996) DNA repair in eukaryotes. Annu. Rev. Biochem., 65, 135–167. [DOI] [PubMed] [Google Scholar]
- 9.Bullock P.A. (1997) The initiation of simian virus 40 DNA replication in vitro. Crit. Rev. Biochem. Mol. Biol., 32, 503–568. [DOI] [PubMed] [Google Scholar]
- 10.Bochkarev A., Pfuetzner,R.A, Edwards,A.M. and Frappier,L. (1997) Structure of the single-stranded-DNA-binding domain of replication protein A bound to DNA. Nature, 385, 176–181. [DOI] [PubMed] [Google Scholar]
- 11.De Laat W.L., Appeldorn,E., Sugasawa,K., Weterings,E., Jaspers,N.G.J. and Hoeijmakers,J.H.J. (1998) DNA-binding polarity of human replication protein A positions nucleases in nucleotide excision repair. Genes Dev., 12, 2598–2609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Beard W.A. and Wilson,S.H. (1995) Purification and domain-mapping of mammalian DNA polymerase β. Methods Enzymol., 262, 98–107. [DOI] [PubMed] [Google Scholar]
- 13.Henricksen L.A., Umbricht,C.B. and Wold,M.S. (1994) Recombinant replication protein A: expression, complex formation, and functional characterization. J. Biol. Chem., 269, 11121–11132. [PubMed] [Google Scholar]
- 14.Sambrook J., Fritsch,E.F. and Maniatis,T. (1989) Molecular Cloning: A Laboratory manual, 2nd Edn. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
- 15.Wlassoff W.A., Dobrikov,M.I., Safronov,I.V., Dudko,R.Y., Bogachev,V.S., Kandaurova,V.V., Shishkin,G.V., Dymshits,G.M. and Lavrik,O.I. (1995). Synthesis and characterization of (d)NTP derivatives substituted with residues of different photoreagents. Bioconjugate Chem., 6, 352–360. [DOI] [PubMed] [Google Scholar]
- 16.Levina A.S., Berezovskii,M.V., Venjaminova,A.G., Dobrikov,M.I., Repkova,M.N. and Zarytova,V.F. (1993) Photomodification of RNA and DNA fragments by oligonucleotide reagents bearing arylazide groups. Biochimie, 32, 249–251. [DOI] [PubMed] [Google Scholar]
- 17.Laemmli U.K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 277, 680–685. [DOI] [PubMed] [Google Scholar]
- 18.Singhal R.K. and Wilson,S.H. (1993) Short gap-filling synthesis by DNA polymerase beta is processive. J. Biol. Chem., 268, 15906–15911. [PubMed] [Google Scholar]
- 19.Prasad R., Beard,W.A. and Wilson,S.H. (1994) Studies of gapped DNA substrate binding by mammalian DNA polymerase beta. Dependence on 5′-phosphate group. J. Biol. Chem., 269, 18096–18101. [PubMed] [Google Scholar]
- 20.Kim C., Paulus,B.F. and Wold,M.S. (1994) Interaction of human replication protein A with oligonucleotides. Biochemistry, 33, 14197–14206. [DOI] [PubMed] [Google Scholar]
- 21.Iftode C. and Borowiec,J.A. (2000) 5′→3′ Molecular polarity of human replication protein A (hRPA) binding to pseudo-origin DNA substrates. Biochemistry, 39, 11970–11981. [DOI] [PubMed] [Google Scholar]
- 22.Walther A.P., Gomes,X.V., Lao,Ye., Lee,C.G. and Wold,M.S. (1999) Replication protein A interactions with DNA: 1. Functions of the DNA-binding and zinc-finger domains of the 70-kDa subunit. Biochemistry, 38, 3963–3973. [DOI] [PubMed] [Google Scholar]
- 23.Gomes X.V. and Wold,M.S. (1995) Structural analysis of human replication protein A. Mapping functional domains of the 70-kDa subunit. J. Biol. Chem., 270, 4534–4543. [DOI] [PubMed] [Google Scholar]
- 24.Gomes X.V. and Wold,M.S. (1996) Functional domains of the 70-kilodalton subunit of human replication protein A. Biochemistry, 35, 10558–10568. [DOI] [PubMed] [Google Scholar]
- 25.Bochkareva E., Frappier,L., Edwards,A.M. and Bochkarev,A. (1998) The RPA32 subunit of human replication protein A contains a single-stranded DNA-binding domain. J. Biol. Chem., 273, 3932–3936. [DOI] [PubMed] [Google Scholar]
- 26.Brill S.J. and Bastin-Shanower,S. (1998) Identification and characterization of the fourth single-stranded-DNA binding domain of replication protein A. Mol. Cell. Biol., 18, 7225–7234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lao Y., Lee,C.G. and Wold,M.S. (1999) Replication protein A interactions with DNA: 2. Characterization of double-stranded DNA binding/helix-destabilization activities and the role of the zinc-finger domain in DNA replication. Biochemistry, 38, 3974–3984. [DOI] [PubMed] [Google Scholar]
- 28.Lao Y., Gomes,X.V., Ren,Y., Taylor,J.S. and Wold,M.S. (2000) Replication protein A interactions with DNA. III. Molecular basis of recognition of damaged DNA. Biochemistry, 39, 850–859. [DOI] [PubMed] [Google Scholar]
- 29.Bochkarev A., Bochkareva,E., Frappier,L. and Edwards,A.M. (1999) The crystal structure of the complex of replication protein A subunits RPA32 and RPA14 reveals a mechanism for single-stranded DNA binding. EMBO J., 18, 4498–4504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Alani E., Thresher,R., Griffithand,J.D. and Kolodner,R.D. (1992) Characterization of DNA-binding and strand-exchange stimulation properties of y-RPA, a yeast single-strand-DNA-binding protein. J. Mol. Biol., 227, 54–71. [DOI] [PubMed] [Google Scholar]
- 31.Blackwell L.J. and Borowiec,J.A. (1994) Human replication protein A binds single-stranded DNA in two distinct complexes. Mol. Cell. Biol., 14, 3993–4001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Blackwell L.J., Borowiec,J.A. and Mastrangelo,I.A. (1996) Single-stranded-DNA binding alerts human replication protein A structure and facilitates interaction with DNA-dependent protein kinase. Mol. Cell. Biol., 16, 4798–4807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sibenaller Z.A., Sorenson,B.R. and Wold,M.S. (1998) The 32- and 14-kilodalton subunits of replication protein A are responsible for species-specific interactions with single-stranded DNA. Biochemistry, 37, 12496–12506. [DOI] [PubMed] [Google Scholar]
- 34.Iftode C. and Borowiec,J.A. (1998) Unwinding of origin-specific structures by human replication protein A occurs in a two-step process. Nucleic Acids Res., 26, 5636–5643. [DOI] [PMC free article] [PubMed] [Google Scholar]