

Summary
Development of an effective vaccine against hepatitis C virus (HCV) has long been defined as a difficult challenge due to the considerable variability of this RNA virus and the observation that convalescent humans and chimpanzees could be re-infected after re-exposure. On the other hand, progress in the understanding of antiviral immune responses in patients with viral clearance has elucidated key mechanisms playing a role for control of viral infection. Studies investigating prophylactic vaccine approaches in chimpanzees have confirmed that the induction and maintenance of strong helper and cytotoxic T cell immune responses against multiple viral epitopes is necessary for protection against viral clearance and chronic infection. A multispecific B cell response resulting in rapid induction of cross-neutralizing antibodies may assist cellular responses. Therapeutic vaccination formulations currently evaluated in clinical phase are facing the fact that the immune system of chronic carriers is impaired and need the restoration of T cells functions to enhance their efficacy.
Keywords: Animals; Biomedical Research; trends; Hepatitis C; immunology; prevention & control; Hepatitis C Antibodies; immunology; Humans; Pan troglodytes; T-Lymphocytes, Cytotoxic; immunology; Viral Vaccines; immunology
Keywords: antibody, HCV, chimpanzee model, therapeutic vaccine, prophylactic vaccine
Introduction
An estimated 170 million persons are chronically infected with HCV and 3 to 4 million individuals are newly infected each year [1]. Region-specific estimates of HCV prevalence range from < 1.0% in Northern Europe to > 2.9% in Northern Africa. The lowest prevalence (0.01%–0.1%) has been reported from countries in the United Kingdom and Scandinavia; the highest prevalence (up to 15–20%) has been reported from Egypt [2]. About 80% of newly infected subjects progress to chronic infection. Cirrhosis which develops in about 10% to 20% of chronically infected patients is associated with a high risk of liver cancer (1% to 5% of chronically infected persons over a period of 20 to 30 years) [3]. No vaccine is currently available to prevent hepatitis C [4]. Treatment options for chronic hepatitis C [5], are of limited efficacy and too costly for most persons in developing countries to afford [6,7,8]. Thus, the development of a vaccine will be pivotal to decrease the burden of HCV-induced liver diseases including liver cirrhosis and hepatocellular carcinoma world-wide. In this review, we will first summarize the impact of recent progress in molecular virology and antiviral immune responses for the design of novel HCV vaccine strategies. We will then review progress in vaccine development by reviewing the investigation of vaccine candidates in chimpanzees and HCV-infected patients.
Genetic variability – a major challenge for vaccine development
HCV is an enveloped single stranded RNA virus of positive polarity which is the sole member of the genus Hepacivirus within the family Flaviviridae. The HCV RNA genome encodes a unique polyprotein of about 3000 amino acids, and is flanked at its 5′ and 3′ ends by two highly conserved untranslated regions involved in the translation and replication processes of the virus, respectively. The polyprotein encodes at least 10 proteins. The structural proteins, which form the viral particle, include the core protein and the envelope glycoproteins E1 and E2. The non–structural proteins include the p7 ion channel, the NS2-3 protease, the NS3 serine protease and RNA helicase, the NS4A polypeptide, the NS4B and NS5A proteins and the NS5B RNA-dependent RNA polymerase [9,10].
HCV infection in vivo is highly dynamic, with a viral half-life of only a few hours and production and clearance of an estimated 1012 particles per day in a given individual [11]. This high replicative activity, together with the lack of a proof–reading function of the NS5B viral polymerase is at the origin of a high genetic variability of HCV [12]. HCV mutates nearly one nucleotide per replication cycle. Six major HCV genotypes and 100 subtypes have been identified world wide [13]. Furthermore, several distinct but closely related HCV variants coexist within each infected individual referred as quasipecies. The envelope glycoprotein genes display some of the highest levels of genetic heterogeneity with E2 exhibiting greater variability at the quasispecies level than E1 [14]. Analysis of viral evolution has shown that amino terminus of the E2 envelope contains residues that have a very high propensity for adaptive change. This region known as the first hypervariable region (HVR-1) has important functions in viral binding and entry, including CD81 binding and membrane fusion [15] and is targeted by neutralizing antibodies [14]. HCV variability has also been described for cytotoxic T lymphocyte (CTL) epitopes [16,17,18,19].
Progress in the development of model systems for the study of host immune responses
The lack of an efficient in vitro replication and infection model system [20] has long hampered the characterization of neutralizing antibodies and functional studies of viral variants escaping B and T cell responses. In 2003, the development of retroviral particles pseudotyped with HCV envelope glycoproteins (HCVpps) for the first allowed the study of viral entry and antibody-mediated neutralization [21,22]. This model has not only allowed the identification of novel identify HCV entry factors such as claudin-1 [23,24,25] but also the investigation of neutralizing antibodies in HCV-infected chimpanzees and humans [14,26,27,28]. The development of a tissue culture model based on recombinant cell-culture derived HCV (HCVcc) infecting human hepatoma cells in 2005 has been a breakthrough for hepatitis C virus research [29,30,31]. This robust model system based on a unique viral isolate allowing efficient viral replication in the human hepatoma cell line Huh7 for the first time allows the investigation of the entire viral life cycle and virus-host interactions during viral infection. Furthermore, this system has been successfully applied to study the molecular mechanisms of antibody-mediated neutralization [32,33] and mechanisms of viral escape from B and T cell responses [19,26,28].
Challenge for the development of a B cell vaccine: viral escape from neutralizing antibodies
In the last few years, substantial progress has been made in the understanding of the impact of humoral immune responses for control of HCV infection. Recent data obtained from unique patient cohorts with well defined viral isolates provided new insights into the perspectives and challenges for vaccine development.
Recently, longitudinal studies of two cohorts of acute phase patients revealed, a correlation between the rapid induction of circulating neutralizing antibodies and viral clearance [27,34]. High-titer cross-neutralizing antibodies were detected during the acute phase in patients who subsequently cleared viral infection [27]. In contrast, patients progressing into chronic HCV infection were characterized by a delayed induction of neutralizing antibodies [27]. Paradoxically, these antibodies were not able to control HCV infection. An elegant study by von Hahn and colleagues provided insights into the molecular mechanisms of this finding [35]. This latter study demonstrated elegantly that HCV continuously escapes the host neutralizing response, suggesting that the neutralizing antibody response of the host lags behind the rapidly evolving HCV envelope glycoprotein sequences of the quasispecies population [35]. This observation may explain the fact that neutralizing antibodies identified in chronic HCV patients are not able to control chronic HCV infection. This hypothesis was further confirmed by a recent study in liver transplant patients, demonstrating that escape from antibody-mediated neutralization plays a key role for selection of viral variants re-infecting the liver graft (Fafi-Kremer et al., Oral presentation, 43rd Annual Meeting of the European Association for the Study of the Liver, Milan 2008).
Polyclonal and monoclonal antibodies which are capable of neutralizing diverse variants of HCV, at least in vitro recognise discontinuous (conformational) regions of the HCV glycoproteins [27,35]. Most recently, it was shown that HCV neutralizing responses in HCV-infected patients target viral entry after HCV binding most likely related to HCV-CD81 and HCV-SR-BI interaction as well as membrane fusion [36]. It is thus conceivable that the combination of cross-neutralizing antibodies targeting distinct HCV epitopes involved in distinct entry steps might allow to efficiently neutralize HCV in vivo.
In addition to its variability HCV has developed different other means to escape the neutralizing antibody response: (1) the induction of antibodies interfering with neutralizing antibodies [37]; (2) the association of HCV with serum factors such as LDL and very low-density lipoprotein (VLDL) [38]; (3) the interplay of HCV glycoproteins and SR-BI with HDL [39,40,41,42]; (4) the shielding of neutralizing epitopes by glycosylation of defined amino acids of envelope glycoproteins [43,44]; and (5) direct cell-to-cell transfer of the virus [45]. Thus, a combination of different mechanisms (for example, masking of neutralizing epitopes or receptor binding sites by mutational variation or by glycosylation) as described for viruses such as HIV [46,47] may also apply to HCV and provide an explanation for why neutralizing antibodies detected in acute and chronic-phase patients may not be effective in vivo [33].
These studies highlight the importance of the characterization of the molecular mechanisms for escape from neutralizing antibodies for the development of antibody-based therapeutic strategies and the formulation of an efficient B-cell immunogens. The development of a B vaccine is essentially challenged by the identification of epitopes largely conserved among different genotypes and isolates that could induce antibodies capable to neutralize efficiently a heterologous inoculum before it reaches its target cell. Most importantly, the recent report of human monoclonal antibodies (mAbs) that neutralize genetically diverse HCV isolates and protect against heterologous HCV quasispecies challenge in a human liver–chimeric mouse model [48] validates the concept of the use of neutralizing antibodies in prevention of HCV infection. These data also provide evidence that broadly neutralizing antibodies to HCV can protect against heterologous viral infection and that a prophylactic vaccine against HCV may be achievable.
T cell responses and control of viral infection
Studies in humans and chimpanzees have shown that a self-limited course of acute hepatitis C is associated with a vigorous and sustained CD4+ and CD8+ T cell response targeting multiple HCV regions [49,50,51,52,53]. After resolution of infection memory T cells maintained over decades and can mediate protective immunity in spontaneously HCV-recovered chimpanzees following re-challenge with homologous and heterologous HCV [54,55,56]. The rapid control of HCV replication following re-challenge was found to be associated with early anamnestic HCV-specific CD4+ and CD8+ T cell responses, including memory CD4+ T cell responses [57,58,59,60]. However, recent studies in chimpanzees contradict the early studies [61]. Thus, a chimpanzee that had previously demonstrated protective immunity following multiple re-challenges with heterologous viruses became chronically infected when re-exposed to the virus originally inoculated into the animal [61]. These findings indicate that further studies are needed to fully understand the immunological correlates of protective immunity against HCV.
In contrast to acute resolving HCV infection, persisting acute HCV infection is associated with a weak and only monospecific CD4+ T cell responses [62]. Regarding the role of CD8+ T cells during acute HCV infection with persistence, the picture is currently less clear. Recent studies in humans have revealed that even strong CD8+ T cell responses in the acute phase of infection may not be adequate to prevent progression to chronicity [17,63,64]. Urbani et al. showed that at clinical onset, CD8 responses are similarly weak and narrowly focused in both self-limited and chronically evolving infections [63]. At this stage, CD4 responses are deeply impaired in patients with a chronic outcome as they are weak and of narrow specificity, unlike the strong, broad and T helper 1-oriented CD4 responses associated with resolving infections. Only patients able to finally control infection show maturation of CD8 memory sustained by progressive expansion of CD127+ CD8 cells [63]. Thus, a poor CD8 response in the acute stage of infection may enhance the overall probability of chronic viral persistence. The presence of functional CD4 responses represents one of the factors dictating the fate of infection by directly contributing to control of the virus and by promoting maturation of protective memory CD8 responses. Viral escape to CD8+T cell is another important mechanism of T cell response failure in patients developing persistent infection and needs to be taken in account in vaccination setting [17,65,66,67]. Studies in humans and chimpanzees have shown that mutations in HLA class I restricted epitopes targeted by CD8+ T cells, occur early in HCV infection and are associated with persistence [68,69]. The role of HLA alleles in determining the outcome of an HCV infection has been recently studied in an Irish cohort of women accidentally infected with HCV [70]. The HLA class I alleles A3, B27 and Cw*01 were significantly associated with viral clearance whereas B8 was associated with viral persistence indicating that the host genetic background is an important variable that can influence infection outcome [70]. Interestingly stable cytotoxic T cell escape mutations have been linked to maintenance of viral fitness [19]. According to these authors, these observations elucidate potential mechanism by which viral persistence is established. Whereas consequences of stable integration of escape mutations into viral genomes are not clear, it is formally possible that epitopes presented by the most prevalent MHC class I molecules in human population will eventually be lost or become less dominant, an outcome that could have implications for vaccine development.
In the chronic phase, virus specific CD4+ and CD8+ T cell responses are also detectable and mechanisms responsible for their failure to control infection could help to efficiently develop immunotherapeutic vaccine. HCV-specific CD4+ and CD8+ T cells isolated from chronically infected patients usually display functional and maturation defects including reduced cytotoxic potential, reduced secretion of Th1-type cytokines and a reduced proliferative capacity in response to ex vivo antigenic stimulation [53,71,72]. Different mechanisms may be involved in secondary T cell failure of HCV-specific CD4+ and CD8+ T cells in chronic HCV infection, such as down-regulation of virus-specific T cell responses via T regulatory cells (Treg) and signalling through programmed death 1 receptor (PD-1). An increased frequency of Treg cells has been observed in patients with chronic HCV infection compared to individuals who spontaneously resolve HCV infection [73,74,75,76]. However, a recent study in chimpanzees showed no difference in the frequency of Treg cells and the extent of suppression irrespective of the outcome of the infection [77]. Down-regulation of virus-specific T cell responses via signalling through PD-1 on T cells has been linked with virus-specific T cell deficiency during chronic viral infections in a murine model and in humans [78,79]. Several recent studies demonstrated high expression levels of PD1 on HCV specific CD8+ T cells in patients with persistent HCV infection [80]. Moreover, HCV-specific T cells that showed increased expression of PD1 on their surface exhibited impaired IFN-γ production and reduced proliferative potential in response to ex vivo HCV antigen stimulation emphasizing the general state of immunosuppression characterising HCV specific T cells in chronic infection [80,81,82].
Taken together, the outcome of an HCV infection appears to be modulated by a complex interplay of immunological factors (primary and secondary T-cell failure), host genetics (HLA background) and virological factors (viral escape mutations). Thus, the development of prophylactic and therapeutic vaccine approaches for HCV is challenged by: (1) the technical challenge involved in formulating a vaccine that is effective against different HCV genotypes and the multiple circulating viral quasispecies, (2) designing a vaccine targeted at eliciting broad T cell responses in spite of the fact that the quality of a successful T cell response is not completely understood (3) the long term maintenance of long-lived memory CD4+ and CD8+ T cells responses generated by a successful HCV vaccine.
Finally, the development of a therapeutic vaccine is challenged by secondary T cell failure associated with persistent HCV infection. Thus, therapeutic vaccine design has not only to overcome an impaired or suppressed T cell response in patients with persistent HCV infection but also has to ensure that vaccine induced immune enhancement does not result in detrimental immunopathology.
Novel approaches for HCV vaccine development
Although a robust tissue culture system for the production of infectious particles has been established [29,30,31], this system does not yet allow the production of virus in quantities required to design vaccines based on killed or attenuated virus. In addition, a live attenuated approach appears not to be realistic because of the high tendency of the virus to persist in the host. Even recombinant viruses, such as those with deletions of the 3′ untranslated region or HVR1, which appear to cause attenuated acute disease, readily persist in the host [83]. Thus, novel vaccine strategies are pursued. These strategies include among others recombinant subunit proteins, virus-like particles, peptides, DNA, recombinant non-pathogenic live vectors, and prime boost approaches (Table 1) [4,84,85,86]. Recent progress using the HCV tissue culture model, allowed production of noninfectious trans-complemented HCV particles which may represent another promising vaccine approach based on virus-like particles [87].
Table 1. HCV vaccine strategies*.
Currently explored vaccine strategies including potential advantages and limitations are listed.
| Vaccine Strategy | Advantages | Limitations |
|---|---|---|
| Peptides | Induction of humoral and cellular immune responses, low toxicity | Immunogenicity limited to few epitopes |
| Recombinant envelope glycoproteins | Induction of cross-neutralizing neutralizing antibodies, low toxicity, Proof-of-concept as a vaccine for other viruses successful (HBV) | Limited cellular immune response Epitopes restricted to structural proteins |
| Virus-like particles | Induction of humoral and cellular immune responses, Proof-of-concept as a vaccine for other viruses successful (HPV) | Epitopes restricted to structural proteins Challenge of clinical grade purification |
| Plasmid DNA | Humoral and strong cellular immune responses, multigenotype approach | Reduced immunogenicity in humans |
| Live viral vectors | ||
| Gene-deleted adenovirus | Efficient induction of cellular immune responses, experience in clinical trials | Pre-existing immunity to adenovirus may limit immunogenicity in humans |
| Vaccinia virus | Efficient induction of cellular immune responses, experience in clinical trials | Dissemination in patients with immunodeficiency |
| Semliki forest virus | Efficient induction of humoral and cellular immune responses | Limited experience in humans |
Adapted from Barth H., Blum H.E., Liang T.J., Baumert T.F. Novel vaccine strategies. In: “State-of the-Art of Hepatology” (Blum, H.E, Grisham, J.W., Manns, M.P., eds) Kluwer Academic Publishers, Dordrecht, 2004, p. 516–522.
Based on the current knowledge of the mechanisms being involved in control of viral infection, a vaccine should induce a multi-specific and vigorous cellular host response, implicating both CD4+ and CD8+ T cells, and a strong and cross-neutralizing antibody response (Figure 1) [4,84,85]. Since acute HCV infection is usually asymptomatic and not associated with liver failure, protection against chronic HCV infection is the key goal of current vaccine approaches [4].
Figure 1.
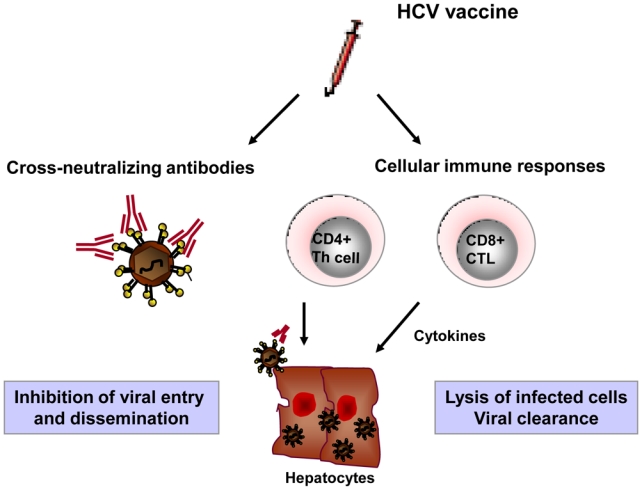
Desired immune responses for an HCV vaccine. Immunization with HCV vaccine should induce broadly cross-neutralizing neutralizing antibodies to inhibit viral spread, and cellular immune response to clear virus and infected hepatocytes via cytolytic and non cytolytic mechanisms.
Evaluation of prophylactic vaccine candidates in the chimpanzee model
In this review we are focussing on vaccine formulations with preclinical evaluation in chimpanzees, the only established in vivo model for the study of HCV infection in an immunocompetent host (Table 2). One of the first vaccine studies in chimpanzees was conducted by Choo and colleagues at Chiron Corp. using recombinant HCV envelope proteins. Derived from mammalian cells, the two glycoproteins associate together to form a non-disulfide linked gpE1-gpE2 heterodimer that is thought to resemble the pre-virion envelope structure. When combined with oil/water based adjuvants and used to vaccinate naïve chimpanzees, this vaccine candidate elicits anti-envelope antibodies as well as CD4+ T cell responses to E1 and E2. Table 2 shows that HCV carrier rate in chimpanzees that have been vaccinated with E1E2 and challenged with heterologous virus was significantly lower than in unimmunised control chimpanzees [4,88]. These results clearly demonstrate the promise of this approach although protection against chronic infection could not be achieved in all animals.
Table 2.
Main prophylactic HCV vaccine candidates tested in chimpanzees
| PROTOCOL | OUTCOME | Ref |
|---|---|---|
| Recombinant gpE1/gpE2 combined with oil/water adjuvents on months 0, 1 and 7 Challenge: 10–100 infectious doses of homologous HCV1 or heterologous HCV-H |
21 vaccinated chimpanzees/24 controls (pooled results)
|
Choo Q.L. et al. and reviewed by Houghton M. et al. [88] |
| Recombinant DNA Plasmid: truncated E2, HCV 1a Challenge: 100 CID50 homologous monoclonal HCV |
2 vaccinated chimpanzees/1 control
|
Forns X. et al. [90] |
| Recombinant DNA plasmid: core-E1–E2 + NS3-5, gHCV1b Booster: recombinant adenovirus NS3-5 Challenge: CID50 close but heterologous HCV1b |
6 vaccinated chimpanzees/2 controls
|
Youn J.W. et al. [91] |
| Recombinant adenovirus NS3 -NS5B, HCV BK 1b Booster: plasmid DNA NS3-5B Challenge: heterologous H77 1a |
5 vaccinated chimpanzees/5 controls
|
Folgori A. et al. [92] |
| HCV-LP: core E1–E2, HCV1b Booster: HCV-LPs Challenge: 100 CID 50 homologous HCV-CG1b |
4 vaccinated chimpanzees/4 controls
|
Elmowalid G.A. et al. [104] |
| Recombinant DNA plasmids: Core E1–E2 and NS3, HCV 1b J4 Booster: MVA core E1–E2 and NS3 Challenge: heterologous HCV1bJ4 |
4 vaccinated chimpanzees/2 controls
|
Rollier C. et al. [105] |
| Recombinant vaccinia virus rVV: HCV 1b Core E1–E2-P7-NS2-NS3 Booster after 52 week: rVV Challenge: 24 CID50 homologous HCV Rechallenge: pool 20 CID50 of each 6 genotypes |
4 vaccinated chimpanzees/2 controls
|
Youn J.W. et al. [107] |
Another small study explored a different glycoprotein E1E2 vaccine formulation (aa192–715; produced using a Sindbis virus expression system) in two chimpanzees: one naive and one recovered from HCV acute infection: the animals became persistently infected despite apparent control of virus replication [89].
Two studies have shown that progression to chronicity appeared be prevented in HCV-infected chimpanzees via DNA-based and/or adenovirus vector-based immunization inducing HCV E1E2 immune responses [90,91] (Table 2). In 2000, Forns et al. [90] showed that DNA vaccine encoding cell surface-expressed E2 did not elicit sterilizing immunity in two vaccinated chimpanzees against challenge with a monoclonal homologous virus. However, both vaccinated chimpanzees resolved the infection early, whereas the control animal became chronically infected. The limitation of this study is the use of a homologous challenge and the small number of animals included.
Interestingly, Youn et al. [91] demonstrated that vaccination by DNA prime recombinant adenovirus boost induces HCV-specific T cell responses and a long lasting E2-specific antibody response that correlates inversely with peak viral loads. They report that one vaccinated chimpanzee that had sterilizing immunity against heterologous virus generated the highest level of E2 specific total and neutralizing antibody responses as well as strong NS3/NS5 specific T cell proliferative responses. The vaccine and challenge strains were both genotype 1b, but HCV-BK challenge strain differed from HCV vaccine strain in about 7% of total amino acids (Table 2). These results suggest that it could be an advantage to combine in the same vaccine formulation immunogens capable to induce both humoral neutralizing and cellular responses of broad spectrums. Furthermore, this study shows the high immunogenicity of a prime-boost protocol capable to decrease the viral load following challenge with heterologous virus. Nevertheless the frequency of chronic infection was very high.
Folgori et al. [92] used the NS proteins as an immunogen. Indeed NS proteins represent about two thirds of the entire HCV polyprotein (about 2000 amino acids) and contain a large number of previously mapped T cell epitopes [93]. They showed that vaccination with adenoviral vectors and electroporated plasmid DNA coding for the HCV non structural region NS3 to NS5B, protected chimpanzees from acute hepatitis induced by challenge with a heterologous virus differing from the vaccine sequence by more than 13% at the amino acid level (Table 2). All vaccinated chimpanzees that developed a cross-reactive T cell response against the challenge virus (four of five) were capable of resolving the infection after a transient viremia of lower level. The average peak viremia was more than 100 times lower in the HCV vaccine group than in the control group. T cell responses were detected also within the liver. CD8 escape mutations were detectable only in the vaccinated that did not control infection. These results suggest that as in natural infection viral escape may contribute to virus–specific CD8+ T cell failure in the vaccination setting. In absence of neutralizing antibodies, none developed a sterilizing immunity. Although the protection rate was not markedly different in vaccinees and controls (4/5 vaccines and 3/5 controls cleared the virus), these data suggest that the immune responses induced by this strategy could control HCV infection.
Another promising approach consists of non-infectious hepatitis C virus-like particles (HCV-LPs) produced in insect cells. HCV-LPs derived from the HCV structural proteins are characterized by similar morphological, antigenic and functional properties as HCV virions [94,95,96]. Similar to virions, HCV-LPs bind and enter human hepatocytes, hepatoma and lymphoma cell lines using heparan sulfate and scavenger receptor B1 as co-receptors [97,98]. HCV-LPs are efficiently taken up, processed and presented to T cells by human dendritic cells [99,100]. Cross-presentation of HCV-LPs to cytotoxic T cells by dendritic cells underlines the functional and structural similarity of HCV-LPs to HCV virions [99,100]. Since HCV-LPs are derived from a partial genome, entry of HCV-LPs does not lead to productive viral infection. In various mouse models including transgenic HLA-A2 and -B7 mice, HCV-LPs induce a strong virus-specific humoral and cellular immune response including HCV-specific cytotoxic T cells [101,102,103]. To evaluate this approach as a vaccine strategy, preclinical studies were performed in 4 chimpanzees [104]. Animals received HCV-LPs with or without ASO1B adjuvant by injection into the gluteal muscle. The chimpanzees were boosted at weeks 4, 8 and 32. All HCV-LP immunized chimpanzees were challenged 3 weeks after the fourth immunization doses by an intravenous administration of 100 CID50 (50% chimpanzee infectious doses) of homologous monoclonal HCV-CG1b virus. HCV-LP immunization induced HCV-specific cellular immune responses that could control HCV viremia in the 4 animals 10 weeks after the challenge. Four naive chimpanzees were infected with the same inoculum, and three developed persistent infection with high viremia. Although the challenging virus was homologous to the vaccine strain, these preclinical data obtained in animal model systems suggest that HCV-LPs represent a promising vaccine candidate for the prevention of chronic HCV infection.
Rollier et al. [105] described an immunization protocol with two DNA plasmids expressing core-E1E2 and NS3 (genotype 1b, J4 strain) and booster immunizations given at weeks 14 and 20 with MVA (modified vaccinia Ankara) encoding core-E1E2 and NS3. Animals were challenged with 25 CID50 of in vivo titrated HCV1bJ4. The vaccine and challenge strains were both genotype 1b, differing in approximately 5% of total amino acids (Table 2). A robust specific immune response capable of reducing acute phase viral load both in plasma and liver was induced in all 4 vaccinees but poor protection against chronicity (3 out of 4 chronic infections). Longitudinal liver biopsies revealed higher PD-1, CTLA-4 (cytotoxic T lymphocyte-associated antigen 4) and IDO (indoleamine 2,3-dioxygenase) expression in the 4 animals (3 vaccinated, 1 control) which became chronically infected in contrast to the lower levels in the 2 animals which cleared infection. Stimulation of IDO activity by CTLA-4 favours development of tolerogenic dendritic cells [106]. This vaccination protocol with heterologous prime-booster DNA confirms the high immunogenicity of this approach. Although the viremia was reduced in vaccinated animals these data bring evidence of a virus–associated tolerogenic–like state which can develop in the liver and brings new insights into the challenges of the induction of effective immunity to HCV.
Most recently [107] Youn et al. hypothesized that a replicating viral vector such as recombinant vaccinia virus (VV) may provide stimulation of long-lasting humoral and cell-mediated immunity after a single injection. Thus 4 chimpanzees were immunized transdermally twice with recombinant VV expressing HCV genotype 1b: capsid, E1, E2, p7, NS2 and NS3 genes. After challenge with 24 CID50 chimpanzee infective doses of homologous HCV, the two control animals that had received only the parental VV developed chronic infection. All 4 immunized animals resolved HCV infection and developed strong cell-mediated immune responses predominantly directed to NS3. Neutralizing antibody responses after the vaccine booster were relatively weak. To investigate cross-genotype protection, the immunized recovered chimpanzees were challenged with a pool of six major HCV genotypes. During the acute phase after the multigenotype challenge, all animals had high titer viremia in which genotype 4 dominated (87%) followed by genotype 5 (13%). However, after fluctuating low-level viremia, the viremia finally turned in one negative or persisted at very low levels in the other one up to 102 weeks, end of follow-up. The use of highly attenuated VV strains deserves to be explored in further studies.
All the preclinical studies described here have been based on the chimpanzee as animal model. Given that many chimpanzees spontaneously resolve acute hepatitis C, definite conclusions await human studies. Interestingly, most vaccine candidates succeeded in inducing an immune response capable of reducing acute phase viral load. However, protection against chronic infection following challenge with heterologous strains was limited. This limited protection highlights the difficulty of developing a vaccine protecting against different isolates and emphasizes the challenge of HCV variability. Nevertheless, chimpanzees studies performed using homologous challenges are useful to provide information of potential relevance for the design of vaccine candidates (Table 2).
Populations for HCV vaccine evaluation and suitability
Vaccines that prevent initial infection should be tested in human cohorts at high risk of infection [108]. Efficacy would be defined as reduction in incidence of acute or chronic infections. Even in human populations with annual incidence rate as high as 1%, enrolment of a very large number of participants would be required to show efficacy of vaccines in preventing infection of viral persistence. Intravenous drug users have a high incidence of HCV infections and could be helpful [1,109]. However, it is to be as certain that efficacy studies in such a population will allow licensure of a prophylactic vaccine in the general population. In rural regions of Egypt prevalence of HCV in older adult populations is often 40–50% and health care providers working in hospitals that care for these patients are at high risk of exposure to HCV infection.
The cost effectiveness of the HCV vaccine will depend on the population. It will be given according to individual’s risk of infection. Intravenous drug users, the population with the highest incidence of infection, and health care providers at risk of needle stick injuries and other exposures to HCV would benefit from a protective vaccine. Other groups that should be considered for vaccination are individuals with potential exposures to blood (i.e. fire-fighters, military personnel), sex workers, household contacts of infected individuals [1,109]. Since people who are coinfected with HIV present a substantial excess morbidity and mortality associated with HCV [110], their protection might be beneficial as long as the efficacy of the vaccine has been established in this immunocompromised population.
However, the design of vaccine trials in human cannot be the same as in chimpanzees: once an immunized human individual develops an acute infection following immunization, anti-viral treatment has to be offered. Thus, it will be a major challenge to evaluate the ability of a vaccine candidate to protect against chronic HCV infection.
Immunoprophylaxis by anti-HCV antibodies
There are two primary areas where a strong and broadly reactive antibody to HCV would have clinical relevance. The first is in post-exposure prophylaxis for accidental needlestick or other percutaneous or mucosal exposure. The second and most relevant application would be in the prevention of recurrent HCV infection in the liver transplant setting. Re-infection of the transplanted liver is universal and 20% to 30% of recurrence results in accelerated progression of fibrosis that can lead to cirrhosis in 5 years. Through a cohort effect, as the large population infected with HCV 25 to 40 years ago now reaches the age when the untoward complications of HCV infection are more common, the demands for liver transplantation will increase considerably. Thus, new therapeutic modalities that enhance transplant and patient survival are sorely needed. Passively administered neutralizing antibody, probably with concomitant antiviral therapy, offers a viable and promising option to suppress/eradicate HCV before it infects the naive transplanted liver. Anti-HCV immune globulin preparations, similar to those used successfully to treat hepatitis B virus infections, might be useful in preventing or controlling HCV infections. Indeed, immune globulin prepared from unscreened donors with chronic HCV infection has prevented hepatitis C in liver recipients when administered before exposure to the virus [111]. In terms of therapeutic antibodies for preventing HCV infection or treating hepatitis C, antibodies with the broadest specificity for E2 from different HCV genotypes are obviously desirable. That patients can produce broadly reactive antibodies in response to monotypic infection has been shown recently for one well-studied patient (Patient H) [112,113]. The significance of this broad reactivity and the identity of the epitopes involved remains to be determined. These data suggest that the epitopes detected by polyclonal antibodies derived from chronically infected individuals late in their disease may be highly conserved and this bodes well for the development of passive immunoprophylaxis against hepatitis C. Unfortunately studies using anti-HCV antibodies in liver transplanted patients have not been successful to date [114]. It has been suggested that this might be due to insufficient antibody doses or frequency of administration. It has also been suggested that the presence of non neutralizing antibodies in some anti-HCV antibody containing immunoglobulin preparations may mask epitopes for neutralizing antibodies, thereby rendering these preparations less efficient [37]. Enrichment of anti-HCV antibodies containing immunoglobulin preparations with efficient neutralizing antibodies by depletion of interfering antibodies might allow improving the efficiency of immunotherapy in HCV-infected patients. Human monoclonal antibodies provide an attractive alternative to polyclonal hyperimmune globulin for immunoprophylaxis and immunotherapy, since monoclonal antibodies can be more readily standardized [14]. Interestingly, human monoclonal antibodies capable to neutralize genetically diverse HCV isolates and protect against heterologous HCV quasispecies challenge in a human liver-chimeric mouse model have been described [48]. However, high concentrations of the monoclonal antibodies were required for protection suggesting that more potent antibody preparations will likely be needed for immunotherapy.
Therapeutic vaccines in clinical trials
Several industrial and academic teams have worked on the development of therapeutic vaccine formulations. The aim of therapeutic vaccination is to induce de novo HCV–specific cellular immune responses or correct pre-existing T cell dysfunction in individuals already infected with the virus resulting in control of viral infection or clearance. Therapeutic vaccination is the only area where multiple human trials have been performed [84,85]. Several formulations of therapeutic vaccines against hepatitis C have reached the clinical phase: In a pilot study of therapeutic vaccination using recombinant envelope protein E1 (adjuvanted on alum) 35 patients with chronic hepatitis C were randomized to receive 20 μg of recombinant HCV E1 or placebo intramuscularly [115,116]. Thirty-four patients then received open-label E1 vaccine. Administration of two courses of 6 doses of E1 was well tolerated and induced strong humoral and cellular immune responses in most patients. Twenty-four patients underwent a biopsy before and after two courses of E1 17 months later. Liver histology was scored by two blinded pathologists according to the Ishak and Metavir systems. Nine of 24 patients (38%) showed an improvement of 2 points or more, 10 (41%) remained unchanged, and 5 (21%) showed worsening in total Ishak score. Nine patients (38%) improved according to the Ishak and Metavir fibrosis score. Plasma HCV-RNA levels however remained unchanged whereas alanine aminotransferase levels showed a trend toward a decrease during treatment [115,116]. However, larger studies showing an impact of this immunogen on viral load or liver disease (e. g. fibrosis) were not published. Another approach used IC41, a synthetic peptide containing 7 relevant HCV T cell epitopes and the T helper cell (Th)1/Tc1 adjuvant poly-L-arginine, as vaccine. Sixty HLA-A2 positive chronic HCV patients not responding to or relapsing from standard therapy were randomized in a double blind phase II study into5 groups to receive 6 vaccinations of IC41 (3 different dose groups), HCV peptides alone or poly-L-arginine alone. IC41 could induce HCV-specific Th1/Tc1 responses in a subset of difficult to treat HCV non responder patients despite persisting viremia. Changes in HCV RNA occurred only in single patients. Since strongest T cell responses correlated with HCV RNA decrease in individual patients [117], this approach could have promise when modified, or used in combination with antiviral therapies. The limitation of these two therapeutic vaccines may lie in several factors, among which the fact that they are based on a single immunogen (E1) [115,116] or on a reduced number of T cell epitopes to face the high HCV variability [117,118]. There use might need a combination with antiviral treatment.
Two open-label, multi-center, dose-escalation studies testing the safety and potential biological activity and immunogenicity of a T-cell inducing vaccine (TG4040) based on the highly attenuated pox virus strain MVA expressing NS3 NS4 and NS5B proteins are ongoing (Fournillier et al., Oral presentation, 15Th International Symposium on HCV and Related Viruses, San Antonio 2008). The first results show that TG4040 is well tolerated, has a good safety profile and appears to have an impact on viral load in individual patients despite ongoing infection.
A phase I clinical study of a personalized peptide vaccination for patients infected with HCV 1b who failed to respond to interferon based therapy has been conducted in Japan [119]. This group assessed the safety and efficacy of 3 doses of personalized HCV 1b–derived peptides in 12 HCV 1b positive patients. Decrease of serum ALT and HCV RNA levels after the 14th vaccination was observed in five and three patients, respectively, in a dose dependent manner. Whether these effects have an impact for chronic HCV liver disease remains to be determined.
The marked diversity of approaches used for immunotherapies for HCV illustrates the need to improve our understanding of the interactions of HCV with the host’s immune system and how viral proteins are able to modulate immune responses in the chronically infected patient [108]. An interesting future approach will be the combination of therapeutic vaccines with interferon-based anti-viral therapies, allowing the combination of complementary antiviral effects of anti-virals and therapeutic vaccines.
Expert commentary
While most vaccine development programs have been based on improving HCV cellular immunity, current knowledge suggests that it is desirable to include, in a same vaccine formulation, immunogens capable to induce both humoral neutralizing and cellular responses of broad spectrums. High viral variability and several immune escape mechanisms render antigen selection difficult and require efforts to identify conserved T cell as well as neutralization epitopes and to decipher neutralization mechanisms. The aim is to discover the optimal target of the HCV life cycle and how to counteract viral escape strategies. At the same time, it will be important to develop antigen presentation systems that are efficient in patients with an impaired antiviral immune response, as observed during chronic hepatitis C. A vaccine preventing chronic hepatitis C will be sufficient to prevent HCV-induced liver disease such as end-stage liver cirrhosis and hepatocellular carcinoma.
Five year view
Recent development of cell culture model systems should help to (i) identify highly conserved epitopes and cross neutralizing antibodies, (ii) better understand the mechanisms associated to chronicity: escape mechanisms, dendritic cells/T cell interaction, tolerogenic state, immune dysfunction, lipid interaction.
Progress in the understanding of virus-host interactions and viral escape will most likely result in new prophylactic vaccine formulations with improved cross protection against diverse genotypes and increase duration of vaccine-mediated protection.
The efficacy of therapeutic vaccines will be investigated in combination with interferon-based antiviral strategies and/or immunomodulators.
The development of human cross-neutralizing monoclonal antibodies will offer a perspective for prevention of liver graft re-infection.
Key issues.
Hepatitis C virus (HCV) is a major cause of chronic hepatitis, liver cirrhosis and hepatocellular carcinoma world-wide
Therapeutic options are still limited, a vaccine is not available.
A major challenge for vaccine development is the high genetic variability of the virus
HCV evades innate and adaptive immune responses.
Resolution of infection is associated with vigorous, broad and sustained HCV specific T cell responses in combination with rapid-induction of cross-neutralizing antibodies.
In chronically infected patients virus-specific T cell responses are impaired and the mechanisms are not clearly understood.
Trials investigating prophylactic vaccines in chimpanzees suggest that protection against chronic infection appears to be possible.
Therapeutic vaccines are facing the impairment of antiviral T cell responses in chronic infected patients and may need a combination with antiviral or immunomodulatory molecules.
Footnotes
Financial & competing interests disclosure
This work was supported by Inserm, France, Université Louis Pasteur de Strasbourg (CONECTUS), France, Hôpitaux Universitaires de Strasbourg, France, the European Union (LSHM-CT-2004-503359), the chair of excellence program of the Agence Nationale de la Recherche (ANR-05-CEXC-008), France, the Agence Nationale de la Recherche sur le SIDA et les Hépatites Virales (ANRS-06221), France, the Else-Kröner-Fresenius Foundation, Bad Homburg (P17/07//A83/06), Germany and the Deutsche Forschungsgemeinschaft (Ba3643/1-1; Ba1417/11-2), Bonn, Germany.
References
- 1.Shepard CW, Finelli L, Alter MJ. Global epidemiology of hepatitis C virus infection. Lancet Infect Dis. 2005;5(9):558–567. doi: 10.1016/S1473-3099(05)70216-4. [DOI] [PubMed] [Google Scholar]
- 2.Alter MJ. Epidemiology of hepatitis C virus infection. World J Gastroenterol. 2007;13(17):2436–2441. doi: 10.3748/wjg.v13.i17.2436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lauer GM, Walker BD. Hepatitis C virus infection. N Engl J Med. 2001;345(1):41–52. doi: 10.1056/NEJM200107053450107. [DOI] [PubMed] [Google Scholar]
- 4*.Houghton M, Abrignani S. Prospects for a vaccine against the hepatitis C virus. Nature. 2005;436(7053):961–966. doi: 10.1038/nature04081. An important review on development of new vaccines. [DOI] [PubMed] [Google Scholar]
- 5.Fried MW, Shiffman ML, Reddy KR, Smith C, Marinos G, Goncales FL, Jr, Haussinger D, Diago M, Carosi G, Dhumeaux D, Craxi A, Lin A, Hoffman J, Yu J. Peginterferon alfa-2a plus ribavirin for chronic hepatitis C virus infection. N Engl J Med. 2002;347(13):975–982. doi: 10.1056/NEJMoa020047. [DOI] [PubMed] [Google Scholar]
- 6.Timm J, Roggendorf M. Sequence diversity of hepatitis C virus: implications for immune control and therapy. World J Gastroenterol. 2007;13(36):4808–4817. doi: 10.3748/wjg.v13.i36.4808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Chevaliez S, Pawlotsky JM. Interferon-based therapy of hepatitis C. Adv Drug Deliv Rev. 2007;59(12):1222–1241. doi: 10.1016/j.addr.2007.07.002. [DOI] [PubMed] [Google Scholar]
- 8.Pawlotsky JM, Chevaliez S, McHutchison JG. The hepatitis C virus life cycle as a target for new antiviral therapies. Gastroenterology. 2007;132(5):1979–1998. doi: 10.1053/j.gastro.2007.03.116. [DOI] [PubMed] [Google Scholar]
- 9*.Bartenschlager R, Frese M, Pietschmann T. Novel insights into hepatitis C virus replication and persistence. Adv Virus Res. 2004;63:71–180. doi: 10.1016/S0065-3527(04)63002-8. [DOI] [PubMed] [Google Scholar]
- 10*.Moradpour D, Penin F, Rice CM. Replication of hepatitis C virus. Nat Rev Microbiol. 2007;5(6):453–463. doi: 10.1038/nrmicro1645. Two articles describing the molecular biology of HCV. [DOI] [PubMed] [Google Scholar]
- 11.Neumann AU, Lam NP, Dahari H, Gretch DR, Wiley TE, Layden TJ, Perelson AS. Hepatitis C viral dynamics in vivo and the antiviral efficacy of interferon-alpha therapy. Science. 1998;282(5386):103–107. doi: 10.1126/science.282.5386.103. [DOI] [PubMed] [Google Scholar]
- 12.Drake JW, Charlesworth B, Charlesworth D, Crow JF. Rates of spontaneous mutation. Genetics. 1998;148(4):1667–1686. doi: 10.1093/genetics/148.4.1667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Simmonds P. Genetic diversity and evolution of hepatitis C virus--15 years on. J Gen Virol. 2004;85(Pt 11):3173–3188. doi: 10.1099/vir.0.80401-0. [DOI] [PubMed] [Google Scholar]
- 14.Keck ZY, Machida K, Lai MM, Ball JK, Patel AH, Foung SK. Therapeutic control of hepatitis C virus: the role of neutralizing monoclonal antibodies. Curr Top Microbiol Immunol. 2008;317:1–38. doi: 10.1007/978-3-540-72146-8_1. [DOI] [PubMed] [Google Scholar]
- 15.Cocquerel L, Voisset C, Dubuisson J. Hepatitis C virus entry: potential receptors and their biological functions. J Gen Virol. 2006;87(Pt 5):1075–1084. doi: 10.1099/vir.0.81646-0. [DOI] [PubMed] [Google Scholar]
- 16.Tester I, Smyk-Pearson S, Wang P, Wertheimer A, Yao E, Lewinsohn DM, Tavis JE, Rosen HR. Immune evasion versus recovery after acute hepatitis C virus infection from a shared source. J Exp Med. 2005;201(11):1725–1731. doi: 10.1084/jem.20042284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cox AL, Mosbruger T, Lauer GM, Pardoll D, Thomas DL, Ray SC. Comprehensive analyses of CD8+ T cell responses during longitudinal study of acute human hepatitis C. Hepatology. 2005;42(1):104–112. doi: 10.1002/hep.20749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cox AL, Mosbruger T, Mao Q, Liu Z, Wang XH, Yang HC, Sidney J, Sette A, Pardoll D, Thomas DL, Ray SC. Cellular immune selection with hepatitis C virus persistence in humans. J Exp Med. 2005;201(11):1741–1752. doi: 10.1084/jem.20050121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Uebelhoer L, Han JH, Callendret B, Mateu G, Shoukry NH, Hanson HL, Rice CM, Walker CM, Grakoui A. Stable cytotoxic T cell escape mutation in hepatitis C virus is linked to maintenance of viral fitness. PLoS Pathog. 2008;4(9):e1000143. doi: 10.1371/journal.ppat.1000143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lohmann V, Korner F, Koch J, Herian U, Theilmann L, Bartenschlager R. Replication of subgenomic hepatitis C virus RNAs in a hepatoma cell line. Science. 1999;285(5424):110–113. doi: 10.1126/science.285.5424.110. [DOI] [PubMed] [Google Scholar]
- 21*.Bartosch B, Dubuisson J, Cosset FL. Infectious hepatitis C virus pseudo-particles containing functional E1–E2 envelope protein complexes. J Exp Med. 2003;197(5):633–642. doi: 10.1084/jem.20021756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22*.Hsu M, Zhang J, Flint M, Logvinoff C, Cheng-Mayer C, Rice CM, McKeating JA. Hepatitis C virus glycoproteins mediate pH-dependent cell entry of pseudotyped retroviral particles. Proc Natl Acad Sci USA. 2003;100(12):7271–7276. doi: 10.1073/pnas.0832180100. Two articles describing the development of retroviral particles pseudotyped with HCV envelope glycoproteins enabling to study HCV entry. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Barth H, Liang TJ, Baumert TF. Hepatitis C virus entry: molecular biology and clinical implications. Hepatology. 2006;44(3):527–535. doi: 10.1002/hep.21321. [DOI] [PubMed] [Google Scholar]
- 24.Von HT, Rice CM. Hepatitis C virus entry. J Biol Chem. 2008;283(7):3689–3693. doi: 10.1074/jbc.R700024200. [DOI] [PubMed] [Google Scholar]
- 25.Evans MJ, Von HT, Tscherne DM, Syder AJ, Panis M, Wolk B, Hatziioannou T, McKeating JA, Bieniasz PD, Rice CM. Claudin-1 is a hepatitis C virus co-receptor required for a late step in entry. Nature. 2007;446(7137):801–805. doi: 10.1038/nature05654. [DOI] [PubMed] [Google Scholar]
- 26.Haberstroh A, Schnober EK, Zeisel MB, Carolla P, Barth H, Blum HE, Cosset FL, Koutsoudakis G, Bartenschlager R, Union A, Depla E, Owsianka A, Patel AH, Schuster C, Stoll-Keller F, Doffoel M, Dreux M, Baumert TF. Neutralizing host responses in hepatitis C virus infection target viral entry at postbinding steps and membrane fusion. Gastroenterology. 2008;135(5):1719–1728. doi: 10.1053/j.gastro.2008.07.018. [DOI] [PubMed] [Google Scholar]
- 27.Pestka JM, Zeisel MB, Blaser E, Schurmann P, Bartosch B, Cosset FL, Patel AH, Meisel H, Baumert J, Viazov S, Rispeter K, Blum HE, Roggendorf M, Baumert TF. Rapid induction of virus-neutralizing antibodies and viral clearance in a single-source outbreak of hepatitis C. Proc Natl Acad Sci USA. 2007;104(14):6025–6030. doi: 10.1073/pnas.0607026104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Meunier JC, Russell RS, Goossens V, Priem S, Walter H, Depla E, Union A, Faulk KN, Bukh J, Emerson SU, Purcell RH. Isolation and characterization of broadly neutralizing human monoclonal antibodies to the e1 glycoprotein of hepatitis C virus. J Virol. 2008;82(2):966–973. doi: 10.1128/JVI.01872-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29*.Wakita T, Pietschmann T, Kato T, Date T, Miyamoto M, Zhao Z, Murthy K, Habermann A, Krausslich HG, Mizokami M, Bartenschlager R, Liang TJ. Production of infectious hepatitis C virus in tissue culture from a cloned viral genome. Nat Med. 2005;11(7):791–796. doi: 10.1038/nm1268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30*.Zhong J, Gastaminza P, Cheng G, Kapadia S, Kato T, Burton DR, Wieland SF, Uprichard SL, Wakita T, Chisari FV. Robust hepatitis C virus infection in vitro. Proc Natl Acad Sci USA. 2005;102(26):9294–9299. doi: 10.1073/pnas.0503596102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31*.Lindenbach BD, Evans MJ, Syder AJ, Wolk B, Tellinghuisen TL, Liu CC, Maruyama T, Hynes RO, Burton DR, McKeating JA, Rice CM. Complete replication of hepatitis C virus in cell culture. Science. 2005;309(5734):623–626. doi: 10.1126/science.1114016. Three articles describing the development of a tissue culture model producing infectious virions. [DOI] [PubMed] [Google Scholar]
- 32.Owsianka AM, Tarr AW, Keck ZY, Li TK, Witteveldt J, Adair R, Foung SK, Ball JK, Patel AH. Broadly neutralizing human monoclonal antibodies to the hepatitis C virus E2 glycoprotein. J Gen Virol. 2008;89(Pt 3):653–659. doi: 10.1099/vir.0.83386-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zeisel MB, Cosset FL, Baumert TF. Host neutralizing responses and pathogenesis of hepatitis C virus infection. Hepatology. 2008;48(1):299–307. doi: 10.1002/hep.22307. [DOI] [PubMed] [Google Scholar]
- 34.Lavillette D, Morice Y, Germanidis G, Donot P, Soulier A, Pagkalos E, Sakellariou G, Intrator L, Bartosch B, Pawlotsky JM, Cosset FL. Human serum facilitates hepatitis C virus infection, and neutralizing responses inversely correlate with viral replication kinetics at the acute phase of hepatitis C virus infection. J Virol. 2005;79(10):6023–6034. doi: 10.1128/JVI.79.10.6023-6034.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35*.Von Hahn T, Yoon JC, Alter H, Rice CM, Rehermann B, Balfe P, McKeating JA. Hepatitis C virus continuously escapes from neutralizing antibody and T-cell responses during chronic infection in vivo. Gastroenterology. 2007;132(2):667–678. doi: 10.1053/j.gastro.2006.12.008. A study describing the role of neutralizing antibodies and T cell response in viral clearance. [DOI] [PubMed] [Google Scholar]
- 36.Haberstroh A. SEKCPea. Neutralizing host reponses in hepatitis C virus infection target viral entry at post-binding steps and membrane fusion. Gastroenterolgy. 2008 doi: 10.1053/j.gastro.2008.07.018. [DOI] [PubMed] [Google Scholar]
- 37.Zhang P, Wu CG, Mihalik K, Virata-Theimer ML, Yu MY, Alter HJ, Feinstone SM. Hepatitis C virus epitope-specific neutralizing antibodies in Igs prepared from human plasma. Proc Natl Acad Sci USA. 2007;104(20):8449–8454. doi: 10.1073/pnas.0703039104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Nielsen SU, Bassendine MF, Burt AD, Martin C, Pumeechockchai W, Toms GL. Association between hepatitis C virus and very-low-density lipoprotein (VLDL)/LDL analyzed in iodixanol density gradients. J Virol. 2006;80(5):2418–2428. doi: 10.1128/JVI.80.5.2418-2428.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bartosch B, Verney G, Dreux M, Donot P, Morice Y, Penin F, Pawlotsky JM, Lavillette D, Cosset FL. An interplay between hypervariable region 1 of the hepatitis C virus E2 glycoprotein, the scavenger receptor BI, and high-density lipoprotein promotes both enhancement of infection and protection against neutralizing antibodies. J Virol. 2005;79(13):8217–8229. doi: 10.1128/JVI.79.13.8217-8229.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Dreux M, Pietschmann T, Granier C, Voisset C, Ricard-Blum S, Mangeot PE, Keck Z, Foung S, Vu-Dac N, Dubuisson J, Bartenschlager R, Lavillette D, Cosset FL. High density lipoprotein inhibits hepatitis C virus-neutralizing antibodies by stimulating cell entry via activation of the scavenger receptor BI. J Biol Chem. 2006;281(27):18285–18295. doi: 10.1074/jbc.M602706200. [DOI] [PubMed] [Google Scholar]
- 41.Dreux M, Cosset FL. The scavenger receptor BI and its ligand, HDL: partners in crime against HCV neutralizing antibodies. J Viral Hepat. 2007;14(Suppl 1):68–76. doi: 10.1111/j.1365-2893.2007.00919.x. [DOI] [PubMed] [Google Scholar]
- 42.Zeisel MB, Koutsoudakis G, Schnober EK, Haberstroh A, Blum HE, Cosset FL, Wakita T, Jaeck D, Doffoel M, Royer C, Soulier E, Schvoerer E, Schuster C, Stoll-Keller F, Bartenschlager R, Pietschmann T, Barth H, Baumert TF. Scavenger receptor class B type I is a key host factor for hepatitis C virus infection required for an entry step closely linked to CD81. Hepatology. 2007;46(6):1722–1731. doi: 10.1002/hep.21994. [DOI] [PubMed] [Google Scholar]
- 43.Falkowska E, Kajumo F, Garcia E, Reinus J, Dragic T. Hepatitis C virus envelope glycoprotein E2 glycans modulate entry, CD81 binding, and neutralization. J Virol. 2007;81(15):8072–8079. doi: 10.1128/JVI.00459-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Helle F, Goffard A, Morel V, Duverlie G, McKeating J, Keck ZY, Foung S, Penin F, Dubuisson J, Voisset C. The neutralizing activity of anti-hepatitis C virus antibodies is modulated by specific glycans on the E2 envelope protein. J Virol. 2007;81(15):8101–8111. doi: 10.1128/JVI.00127-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Timpe JM, Stamataki Z, Jennings A, Hu K, Farquhar MJ, Harris HJ, Schwarz A, Desombere I, Roels GL, Balfe P, McKeating JA. Hepatitis C virus cell-cell transmission in hepatoma cells in the presence of neutralizing antibodies. Hepatology. 2008;47(1):17–24. doi: 10.1002/hep.21959. [DOI] [PubMed] [Google Scholar]
- 46.Wei X, Decker JM, Wang S, Hui H, Kappes JC, Wu X, Salazar-Gonzalez JF, Salazar MG, Kilby JM, Saag MS, Komarova NL, Nowak MA, Hahn BH, Kwong PD, Shaw GM. Antibody neutralization and escape by HIV-1. Nature. 2003;422(6929):307–312. doi: 10.1038/nature01470. [DOI] [PubMed] [Google Scholar]
- 47.Kwong PD, Doyle ML, Casper DJ, Cicala C, Leavitt SA, Majeed S, Steenbeke TD, Venturi M, Chaiken I, Fung M, Katinger H, Parren PW, Robinson J, Van RD, Wang L, Burton DR, Freire E, Wyatt R, Sodroski J, Hendrickson WA, Arthos J. HIV-1 evades antibody-mediated neutralization through conformational masking of receptor-binding sites. Nature. 2002;420(6916):678–682. doi: 10.1038/nature01188. [DOI] [PubMed] [Google Scholar]
- 48.Law M, Maruyama T, Lewis J, Giang E, Tarr AW, Stamataki Z, Gastaminza P, Chisari FV, Jones IM, Fox RI, Ball JK, McKeating JA, Kneteman NM, Burton DR. Broadly neutralizing antibodies protect against hepatitis C virus quasispecies challenge. Nat Med. 2008;14(1):25–27. doi: 10.1038/nm1698. [DOI] [PubMed] [Google Scholar]
- 49*.Bowen DG, Walker CM. Adaptive immune responses in acute and chronic hepatitis C virus infection. Nature. 2005;436(7053):946–952. doi: 10.1038/nature04079. An important review on determinants of viral clearance or persistence. [DOI] [PubMed] [Google Scholar]
- 50.Lechner F, Wong DK, Dunbar PR, Chapman R, Chung RT, Dohrenwend P, Robbins G, Phillips R, Klenerman P, Walker BD. Analysis of successful immune responses in persons infected with hepatitis C virus. J Exp Med. 2000;191(9):1499–1512. doi: 10.1084/jem.191.9.1499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Thimme R, Oldach D, Chang KM, Steiger C, Ray SC, Chisari FV. Determinants of viral clearance and persistence during acute hepatitis C virus infection. J Exp Med. 2001;194(10):1395–1406. doi: 10.1084/jem.194.10.1395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Thimme R, Bukh J, Spangenberg HC, Wieland S, Pemberton J, Steiger C, Govindarajan S, Purcell RH, Chisari FV. Viral and immunological determinants of hepatitis C virus clearance, persistence, and disease. Proc Natl Acad Sci USA. 2002;99(24):15661–15668. doi: 10.1073/pnas.202608299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Wedemeyer H, He XS, Nascimbeni M, Davis AR, Greenberg HB, Hoofnagle JH, Liang TJ, Alter H, Rehermann B. Impaired effector function of hepatitis C virus-specific CD8+ T cells in chronic hepatitis C virus infection. J Immunol. 2002;169(6):3447–3458. doi: 10.4049/jimmunol.169.6.3447. [DOI] [PubMed] [Google Scholar]
- 54.Weiner AJ, Paliard X, Selby MJ, Medina-Selby A, Coit D, Nguyen S, Kansopon J, Arian CL, Ng P, Tucker J, Lee CT, Polakos NK, Han J, Wong S, Lu HH, Rosenberg S, Brasky KM, Chien D, Kuo G, Houghton M. Intrahepatic genetic inoculation of hepatitis C virus RNA confers cross-protective immunity. J Virol. 2001;75(15):7142–7148. doi: 10.1128/JVI.75.15.7142-7148.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Bassett SE, Guerra B, Brasky K, Miskovsky E, Houghton M, Klimpel GR, Lanford RE. Protective immune response to hepatitis C virus in chimpanzees rechallenged following clearance of primary infection. Hepatology. 2001;33(6):1479–1487. doi: 10.1053/jhep.2001.24371. [DOI] [PubMed] [Google Scholar]
- 56.Major ME, Mihalik K, Puig M, Rehermann B, Nascimbeni M, Rice CM, Feinstone SM. Previously infected and recovered chimpanzees exhibit rapid responses that control hepatitis C virus replication upon rechallenge. J Virol. 2002;76(13):6586–6595. doi: 10.1128/JVI.76.13.6586-6595.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Shoukry NH, Grakoui A, Houghton M, Chien DY, Ghrayeb J, Reimann KA, Walker CM. Memory CD8+ T cells are required for protection from persistent hepatitis C virus infection. J Exp Med. 2003;197(12):1645–1655. doi: 10.1084/jem.20030239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Nascimbeni M, Mizukoshi E, Bosmann M, Major ME, Mihalik K, Rice CM, Feinstone SM, Rehermann B. Kinetics of CD4+ and CD8+ memory T-cell responses during hepatitis C virus rechallenge of previously recovered chimpanzees. J Virol. 2003;77(8):4781–4793. doi: 10.1128/JVI.77.8.4781-4793.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Grakoui A, Shoukry NH, Woollard DJ, Han JH, Hanson HL, Ghrayeb J, Murthy KK, Rice CM, Walker CM. HCV persistence and immune evasion in the absence of memory T cell help. Science. 2003;302(5645):659–662. doi: 10.1126/science.1088774. [DOI] [PubMed] [Google Scholar]
- 60.Lanford RE, Guerra B, Chavez D, Bigger C, Brasky KM, Wang XH, Ray SC, Thomas DL. Cross-genotype immunity to hepatitis C virus. J Virol. 2004;78(3):1575–1581. doi: 10.1128/JVI.78.3.1575-1581.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Bukh J, Thimme R, Meunier JC, Faulk K, Spangenberg HC, Chang KM, Satterfield W, Chisari FV, Purcell RH. Previously infected chimpanzees are not consistently protected against reinfection or persistent infection after reexposure to the identical hepatitis C virus strain. J Virol. 2008;82(16):8183–8195. doi: 10.1128/JVI.00142-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Thimme R, Lohmann V, Weber F. A target on the move: innate and adaptive immune escape strategies of hepatitis C virus. Antiviral Res. 2006;69(3):129–141. doi: 10.1016/j.antiviral.2005.12.001. [DOI] [PubMed] [Google Scholar]
- 63*.Urbani S, Amadei B, Fisicaro P, Tola D, Orlandini A, Sacchelli L, Mori C, Missale G, Ferrari C. Outcome of acute hepatitis C is related to virus-specific CD4 function and maturation of antiviral memory CD8 responses. Hepatology. 2006;44(1):126–139. doi: 10.1002/hep.21242. [DOI] [PubMed] [Google Scholar]
- 64*.Kaplan DE, Sugimoto K, Newton K, Valiga ME, Ikeda F, Aytaman A, Nunes FA, Lucey MR, Vance BA, Vonderheide RH, Reddy KR, McKeating JA, Chang KM. Discordant role of CD4 T-cell response relative to neutralizing antibody and CD8 T-cell responses in acute hepatitis C. Gastroenterology. 2007;132(2):654–666. doi: 10.1053/j.gastro.2006.11.044. Two important studies on the role of CD8 T cell responses. [DOI] [PubMed] [Google Scholar]
- 65.Chang KM, Rehermann B, McHutchison JG, Pasquinelli C, Southwood S, Sette A, Chisari FV. Immunological significance of cytotoxic T lymphocyte epitope variants in patients chronically infected by the hepatitis C virus. J Clin Invest. 1997;100(9):2376–2385. doi: 10.1172/JCI119778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Weiner A, Erickson AL, Kansopon J, Crawford K, Muchmore E, Hughes AL, Houghton M, Walker CM. Persistent hepatitis C virus infection in a chimpanzee is associated with emergence of a cytotoxic T lymphocyte escape variant. Proc Natl Acad Sci USA. 1995;92(7):2755–2759. doi: 10.1073/pnas.92.7.2755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Erickson AL, Kimura Y, Igarashi S, Eichelberger J, Houghton M, Sidney J, McKinney D, Sette A, Hughes AL, Walker CM. The outcome of hepatitis C virus infection is predicted by escape mutations in epitopes targeted by cytotoxic T lymphocytes. Immunity. 2001;15(6):883–895. doi: 10.1016/s1074-7613(01)00245-x. [DOI] [PubMed] [Google Scholar]
- 68.Timm J, Lauer GM, Kavanagh DG, Sheridan I, Kim AY, Lucas M, Pillay T, Ouchi K, Reyor LL, Schulze zur WJ, Gandhi RT, Chung RT, Bhardwaj N, Klenerman P, Walker BD, Allen TM. CD8 epitope escape and reversion in acute HCV infection. J Exp Med. 2004;200(12):1593–1604. doi: 10.1084/jem.20041006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Ray SC, Fanning L, Wang XH, Netski DM, Kenny-Walsh E, Thomas DL. Divergent and convergent evolution after a common-source outbreak of hepatitis C virus. J Exp Med. 2005;201(11):1753–1759. doi: 10.1084/jem.20050122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Neumann-Haefelin C, McKiernan S, Ward S, Viazov S, Spangenberg HC, Killinger T, Baumert TF, Nazarova N, Sheridan I, Pybus O, von WF, Roggendorf M, Kelleher D, Klenerman P, Blum HE, Thimme R. Dominant influence of an HLA-B27 restricted CD8+ T cell response in mediating HCV clearance and evolution. Hepatology. 2006;43(3):563–572. doi: 10.1002/hep.21049. [DOI] [PubMed] [Google Scholar]
- 71.Urbani S, Boni C, Missale G, Elia G, Cavallo C, Massari M, Raimondo G, Ferrari C. Virus-specific CD8+ lymphocytes share the same effector-memory phenotype but exhibit functional differences in acute hepatitis B and C. J Virol. 2002;76(24):12423–12434. doi: 10.1128/JVI.76.24.12423-12434.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Spangenberg HC, Viazov S, Kersting N, Neumann-Haefelin C, McKinney D, Roggendorf M, von WF, Blum HE, Thimme R. Intrahepatic CD8+ T-cell failure during chronic hepatitis C virus infection. Hepatology. 2005;42(4):828–837. doi: 10.1002/hep.20856. [DOI] [PubMed] [Google Scholar]
- 73.Sugimoto K, Ikeda F, Stadanlick J, Nunes FA, Alter HJ, Chang KM. Suppression of HCV-specific T cells without differential hierarchy demonstrated ex vivo in persistent HCV infection. Hepatology. 2003;38(6):1437–1448. doi: 10.1016/j.hep.2003.09.026. [DOI] [PubMed] [Google Scholar]
- 74.Cabrera R, Tu Z, Xu Y, Firpi RJ, Rosen HR, Liu C, Nelson DR. An immunomodulatory role for CD4(+)CD25(+) regulatory T lymphocytes in hepatitis C virus infection. Hepatology. 2004;40(5):1062–1071. doi: 10.1002/hep.20454. [DOI] [PubMed] [Google Scholar]
- 75.Boettler T, Spangenberg HC, Neumann-Haefelin C, Panther E, Urbani S, Ferrari C, Blum HE, von WF, Thimme R. T cells with a CD4+CD25+ regulatory phenotype suppress in vitro proliferation of virus-specific CD8+ T cells during chronic hepatitis C virus infection. J Virol. 2005;79(12):7860–7867. doi: 10.1128/JVI.79.12.7860-7867.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Rushbrook SM, Ward SM, Unitt E, Vowler SL, Lucas M, Klenerman P, Alexander GJ. Regulatory T cells suppress in vitro proliferation of virus-specific CD8+ T cells during persistent hepatitis C virus infection. J Virol. 2005;79(12):7852–7859. doi: 10.1128/JVI.79.12.7852-7859.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Manigold T, Shin EC, Mizukoshi E, Mihalik K, Murthy KK, Rice CM, Piccirillo CA, Rehermann B. Foxp3+CD4+CD25+ T cells control virus-specific memory T cells in chimpanzees that recovered from hepatitis C. Blood. 2006;107(11):4424–4432. doi: 10.1182/blood-2005-09-3903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Barber DL, Wherry EJ, Masopust D, Zhu B, Allison JP, Sharpe AH, Freeman GJ, Ahmed R. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature. 2006;439(7077):682–687. doi: 10.1038/nature04444. [DOI] [PubMed] [Google Scholar]
- 79.Day CL, Kaufmann DE, Kiepiela P, Brown JA, Moodley ES, Reddy S, Mackey EW, Miller JD, Leslie AJ, DePierres C, Mncube Z, Duraiswamy J, Zhu B, Eichbaum Q, Altfeld M, Wherry EJ, Coovadia HM, Goulder PJ, Klenerman P, Ahmed R, Freeman GJ, Walker BD. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature. 2006;443(7109):350–354. doi: 10.1038/nature05115. [DOI] [PubMed] [Google Scholar]
- 80.Urbani S, Amadei B, Tola D, Massari M, Schivazappa S, Missale G, Ferrari C. PD-1 expression in acute hepatitis C virus (HCV) infection is associated with HCV-specific CD8 exhaustion. J Virol. 2006;80(22):11398–11403. doi: 10.1128/JVI.01177-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Radziewicz H, Ibegbu CC, Fernandez ML, Workowski KA, Obideen K, Wehbi M, Hanson HL, Steinberg JP, Masopust D, Wherry EJ, Altman JD, Rouse BT, Freeman GJ, Ahmed R, Grakoui A. Liver-infiltrating lymphocytes in chronic human hepatitis C virus infection display an exhausted phenotype with high levels of PD-1 and low levels of CD127 expression. J Virol. 2007;81(6):2545–2553. doi: 10.1128/JVI.02021-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Bowen DG, Shoukry NH, Grakoui A, Fuller MJ, Cawthon AG, Dong C, Hasselschwert DL, Brasky KM, Freeman GJ, Seth NP, Wucherpfennig KW, Houghton M, Walker CM. Variable patterns of programmed death-1 expression on fully functional memory T cells after spontaneous resolution of hepatitis C virus infection. J Virol. 2008;82(10):5109–5114. doi: 10.1128/JVI.00060-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Forns X, Bukh J, Purcell RH. The challenge of developing a vaccine against hepatitis C virus. J Hepatol. 2002;37(5):684–695. doi: 10.1016/s0168-8278(02)00308-2. [DOI] [PubMed] [Google Scholar]
- 84*.Lang K, Weiner DB. Immunotherapy for HCV infection: next steps. Expert Rev Vaccines. 2008;7(7):915–923. doi: 10.1586/14760584.7.7.915. [DOI] [PubMed] [Google Scholar]
- 85*.Inchauspe G, Michel ML. Vaccines and immunotherapies against hepatitis B and hepatitis C viruses. J Viral Hepat. 2007;14(Suppl 1):97–103. doi: 10.1111/j.1365-2893.2007.00922.x. Two recent reviews on HCV vaccines. [DOI] [PubMed] [Google Scholar]
- 86.Shiina M, Rehermann B. Hepatitis C vaccines: Inducing and challenging memory T cells. Hepatology. 2006;43(6):1395–1398. doi: 10.1002/hep.21210. [DOI] [PubMed] [Google Scholar]
- 87.Steinmann E, Brohm C, Kallis S, Bartenschlager R, Pietschmann T. Efficient trans-encapsidation of hepatitis C virus RNAs into infectious virus-like particles. J Virol. 2008;82(14):7034–7046. doi: 10.1128/JVI.00118-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Choo QL, Kuo G, Ralston R, Weiner A, Chien D, Van NG, Han J, Berger K, Thudium K, Kuo C. Vaccination of chimpanzees against infection by the hepatitis C virus. Proc Natl Acad Sci USA. 1994;91(4):1294–1298. doi: 10.1073/pnas.91.4.1294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Puig M, Major ME, Mihalik K, Feinstone SM. Immunization of chimpanzees with an envelope protein-based vaccine enhances specific humoral and cellular immune responses that delay hepatitis C virus infection. Vaccine. 2004;22(8):991–1000. doi: 10.1016/j.vaccine.2003.09.010. [DOI] [PubMed] [Google Scholar]
- 90.Forns X, Payette PJ, Ma X, Satterfield W, Eder G, Mushahwar IK, Govindarajan S, Davis HL, Emerson SU, Purcell RH, Bukh J. Vaccination of chimpanzees with plasmid DNA encoding the hepatitis C virus (HCV) envelope E2 protein modified the infection after challenge with homologous monoclonal HCV. Hepatology. 2000;32(3):618–625. doi: 10.1053/jhep.2000.9877. [DOI] [PubMed] [Google Scholar]
- 91.Youn JW, Park SH, Lavillette D, Cosset FL, Yang SH, Lee CG, Jin HT, Kim CM, Shata MT, Lee DH, Pfahler W, Prince AM, Sung YC. Sustained E2 antibody response correlates with reduced peak viremia after hepatitis C virus infection in the chimpanzee. Hepatology. 2005;42(6):1429–1436. doi: 10.1002/hep.20934. [DOI] [PubMed] [Google Scholar]
- 92.Folgori A, Capone S, Ruggeri L, Meola A, Sporeno E, Ercole BB, Pezzanera M, Tafi R, Arcuri M, Fattori E, Lahm A, Luzzago A, Vitelli A, Colloca S, Cortese R, Nicosia A. A T-cell HCV vaccine eliciting effective immunity against heterologous virus challenge in chimpanzees. Nat Med. 2006;12(2):190–197. doi: 10.1038/nm1353. [DOI] [PubMed] [Google Scholar]
- 93.Kuiken C, Yusim K, Boykin L, Richardson R. The Los Alamos hepatitis C sequence database. Bioinformatics. 2005;21(3):379–384. doi: 10.1093/bioinformatics/bth485. [DOI] [PubMed] [Google Scholar]
- 94.Baumert TF, Ito S, Wong DT, Liang TJ. Hepatitis C virus structural proteins assemble into viruslike particles in insect cells. J Virol. 1998;72(5):3827–3836. doi: 10.1128/jvi.72.5.3827-3836.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Baumert TF, Vergalla J, Satoi J, Thomson M, Lechmann M, Herion D, Greenberg HB, Ito S, Liang TJ. Hepatitis C virus-like particles synthesized in insect cells as a potential vaccine candidate. Gastroenterology. 1999;117(6):1397–1407. doi: 10.1016/s0016-5085(99)70290-8. [DOI] [PubMed] [Google Scholar]
- 96.Wellnitz S, Klumpp B, Barth H, Ito S, Depla E, Dubuisson J, Blum HE, Baumert TF. Binding of hepatitis C virus-like particles derived from infectious clone H77C to defined human cell lines. J Virol. 2002;76(3):1181–1193. doi: 10.1128/JVI.76.3.1181-1193.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Barth H, Schafer C, Adah MI, Zhang F, Linhardt RJ, Toyoda H, Kinoshita-Toyoda A, Toida T, Van Kuppevelt TH, Depla E, Von Weizsacker F, Blum HE, Baumert TF. Cellular binding of hepatitis C virus envelope glycoprotein E2 requires cell surface heparan sulfate. J Biol Chem. 2003;278(42):41003–41012. doi: 10.1074/jbc.M302267200. [DOI] [PubMed] [Google Scholar]
- 98.Barth H, Cerino R, Arcuri M, Hoffmann M, Schurmann P, Adah MI, Gissler B, Zhao X, Ghisetti V, Lavezzo B, Blum HE, von WF, Vitelli A, Scarselli E, Baumert TF. Scavenger receptor class B type I and hepatitis C virus infection of primary tupaia hepatocytes. J Virol. 2005;79(9):5774–5785. doi: 10.1128/JVI.79.9.5774-5785.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Barth H, Ulsenheimer A, Pape GR, Diepolder HM, Hoffmann M, Neumann-Haefelin C, Thimme R, Henneke P, Klein R, Paranhos-Baccala G, Depla E, Liang TJ, Blum HE, Baumert TF. Uptake and presentation of hepatitis C virus-like particles by human dendritic cells. Blood. 2005;105(9):3605–3614. doi: 10.1182/blood-2004-05-1952. [DOI] [PubMed] [Google Scholar]
- 100.Barth H, Schnober EK, Neumann-Haefelin C, Thumann C, Zeisel MB, Diepolder HM, Hu Z, Liang TJ, Blum HE, Thimme R, Lambotin M, Baumert TF. Scavenger receptor class B is required for hepatitis C virus uptake and cross-presentation by human dendritic cells. J Virol. 2008;82(7):3466–3479. doi: 10.1128/JVI.02478-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Lechmann M, Murata K, Satoi J, Vergalla J, Baumert TF, Liang TJ. Hepatitis C virus-like particles induce virus-specific humoral and cellular immune responses in mice. Hepatology. 2001;34(2):417–423. doi: 10.1053/jhep.2001.26523. [DOI] [PubMed] [Google Scholar]
- 102.Murata K, Lechmann M, Qiao M, Gunji T, Alter HJ, Liang TJ. Immunization with hepatitis C virus-like particles protects mice from recombinant hepatitis C virus-vaccinia infection. Proc Natl Acad Sci USA. 2003;100(11):6753–6758. doi: 10.1073/pnas.1131929100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Qiao M, Murata K, Davis AR, Jeong SH, Liang TJ. Hepatitis C virus-like particles combined with novel adjuvant systems enhance virus-specific immune responses. Hepatology. 2003;37(1):52–59. doi: 10.1053/jhep.2003.50000. [DOI] [PubMed] [Google Scholar]
- 104.Elmowalid GA, Qiao M, Jeong SH, Borg BB, Baumert TF, Sapp RK, Hu Z, Murthy K, Liang TJ. Immunization with hepatitis C virus-like particles results in control of hepatitis C virus infection in chimpanzees. Proc Natl Acad Sci USA. 2007;104(20):8427–8432. doi: 10.1073/pnas.0702162104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Rollier CS, Paranhos-Baccala G, Verschoor EJ, Verstrepen BE, Drexhage JA, Fagrouch Z, Berland JL, Komurian-Pradel F, Duverger B, Himoudi N, Staib C, Meyr M, Whelan M, Whelan JA, Adams VC, Larrea E, Riezu JI, Lasarte JJ, Bartosch B, Cosset FL, Spaan WJ, Diepolder HM, Pape GR, Sutter G, Inchauspe G, Heeney JL. Vaccine-induced early control of hepatitis C virus infection in chimpanzees fails to impact on hepatic PD-1 and chronicity. Hepatology. 2007;45(3):602–613. doi: 10.1002/hep.21573. [DOI] [PubMed] [Google Scholar]
- 106.Mellor AL, Munn DH. IDO expression by dendritic cells: tolerance and tryptophan catabolism. Nat Rev Immunol. 2004;4(10):762–774. doi: 10.1038/nri1457. [DOI] [PubMed] [Google Scholar]
- 107.Youn JW, Hu YW, Tricoche N, Pfahler W, Shata MT, Dreux M, Cosset FL, Folgori A, Lee DH, Brotman B, Prince AM. Evidence for protection against chronic hepatitis C virus infection in chimpanzees by immunization with replicating recombinant vaccinia virus. J Virol. 2008;82(21):10896–10905. doi: 10.1128/JVI.01179-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Strickland GT, El-Kamary SS, Klenerman P, Nicosia A. Hepatitis C vaccine: supply and demand. Lancet Infect Dis. 2008;8(6):379–386. doi: 10.1016/S1473-3099(08)70126-9. [DOI] [PubMed] [Google Scholar]
- 109.Esteban JI, Sauleda S, Quer J. The changing epidemiology of hepatitis C virus infection in Europe. J Hepatol. 2008;48(1):148–162. doi: 10.1016/j.jhep.2007.07.033. [DOI] [PubMed] [Google Scholar]
- 110.Blackard JT, Sherman KE. HCV/HIV co-infection: time to re-evaluate the role of HIV in the liver? J Viral Hepat. 2008;15(5):323–330. doi: 10.1111/j.1365-2893.2008.00970.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Feray C, Gigou M, Samuel D, Ducot B, Maisonneuve P, Reynes M, Bismuth A, Bismuth H. Incidence of hepatitis C in patients receiving different preparations of hepatitis B immunoglobulins after liver transplantation. Ann Intern Med. 1998;128(10):810–816. doi: 10.7326/0003-4819-128-10-199805150-00003. [DOI] [PubMed] [Google Scholar]
- 112.Logvinoff C, Major ME, Oldach D, Heyward S, Talal A, Balfe P, Feinstone SM, Alter H, Rice CM, McKeating JA. Neutralizing antibody response during acute and chronic hepatitis C virus infection. Proc Natl Acad Sci USA. 2004;101(27):10149–10154. doi: 10.1073/pnas.0403519101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Meunier JC, Engle RE, Faulk K, Zhao M, Bartosch B, Alter H, Emerson SU, Cosset FL, Purcell RH, Bukh J. Evidence for cross-genotype neutralization of hepatitis C virus pseudo-particles and enhancement of infectivity by apolipoprotein C1. Proc Natl Acad Sci USA. 2005;102(12):4560–4565. doi: 10.1073/pnas.0501275102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Davis GL. Hepatitis C immune globulin to prevent HCV recurrence after liver transplantation: chasing windmills? Liver Transpl. 2006;12(9):1317–1319. doi: 10.1002/lt.20889. [DOI] [PubMed] [Google Scholar]
- 115.Leroux-Roels G, Depla E, Hulstaert F, Tobback L, Dincq S, Desmet J, Desombere I, Maertens G. A candidate vaccine based on the hepatitis C E1 protein: tolerability and immunogenicity in healthy volunteers. Vaccine. 2004;22(23–24):3080–3086. doi: 10.1016/j.vaccine.2004.02.002. [DOI] [PubMed] [Google Scholar]
- 116.Nevens F, Roskams T, Van VH, Horsmans Y, Sprengers D, Elewaut A, Desmet V, Leroux-Roels G, Quinaux E, Depla E, Dincq S, Vander SC, Maertens G, Hulstaert F. A pilot study of therapeutic vaccination with envelope protein E1 in 35 patients with chronic hepatitis C. Hepatology. 2003;38(5):1289–1296. doi: 10.1053/jhep.2003.50474. [DOI] [PubMed] [Google Scholar]
- 117.Klade CS, Wedemeyer H, Berg T, Hinrichsen H, Cholewinska G, Zeuzem S, Blum H, Buschle M, Jelovcan S, Buerger V, Tauber E, Frisch J, Manns MP. Therapeutic vaccination of chronic hepatitis C nonresponder patients with the peptide vaccine IC41. Gastroenterology. 2008;134(5):1385–1395. doi: 10.1053/j.gastro.2008.02.058. [DOI] [PubMed] [Google Scholar]
- 118.Firbas C, Jilma B, Tauber E, Buerger V, Jelovcan S, Lingnau K, Buschle M, Frisch J, Klade CS. Immunogenicity and safety of a novel therapeutic hepatitis C virus (HCV) peptide vaccine: a randomized, placebo controlled trial for dose optimization in 128 healthy subjects. Vaccine. 2006;24(20):4343–4353. doi: 10.1016/j.vaccine.2006.03.009. [DOI] [PubMed] [Google Scholar]
- 119.Yutani S, Yamada A, Yoshida K, Takao Y, Tamura M, Komatsu N, Ide T, Tanaka M, Sata M, Itoh K. Phase I clinical study of a personalized peptide vaccination for patients infected with hepatitis C virus (HCV) 1b who failed to respond to interferon-based therapy. Vaccine. 2007;25(42):7429–7435. doi: 10.1016/j.vaccine.2007.08.005. [DOI] [PubMed] [Google Scholar]