


Abstract
Cbf1p is a Saccharomyces cerevisiae chromatin protein belonging to the basic region helix–loop–helix leucine zipper (bHLHzip) family of DNA binding proteins. Cbf1p binds to a conserved element in the 5′-flanking region of methionine biosynthetic genes and to centromere DNA element I (CDEI) of S.cerevisiae centromeric DNA. We have determined the apparent equilibrium dissociation constants of Cbf1p binding to all 16 CDEI DNAs in gel retardation assays. Binding constants of full-length Cbf1p vary between 1.7 and 3.8 nM. However, the dissociation constants of a Cbf1p deletion variant that has been shown to be fully sufficient for Cbf1p function in vivo vary in a range between 3.2 and 12 nM. In addition, native polyacrylamide gel electrophoresis revealed distinct changes in the 3D structure of the Cbf1p/CEN complexes. We also show that the previously reported DNA binding stimulation activity of the centromere protein p64 functions on both the Cbf1 full-length protein and a deletion variant containing only the bHLHzip domain of Cbf1p. Our results suggest that centromeric DNA outside the consensus CDEI sequence and interaction of Cbf1p with adjacent centromere proteins contribute to the complex formation between Cbf1p and CEN DNA.
INTRODUCTION
Chromosome segregation in mitosis and meiosis depends on a particular chromosomal structure, the centromere. It provides a chromosomal attachment site, the kinetochore, for the spindle microtubules. Once attached, kinetochore proteins resembling molecular motors actively move chromosomes along microtubules. The centromere of Saccharomyces cerevisiae is ideal for a molecular dissection and in vitro reconstitution because its structure is much less complex than that of higher eukaryotes. The DNA involved in building the S.cerevisiae kinetochore, the centromere DNA, is short (125 bp) whereas it is very long and repetitive in higher eukaryotes (for reviews see 1–5).
The centromere DNA (CEN DNA) of S.cerevisiae is organized into three elements, the centromere DNA element I (CDEI), CDEII and CDEIII (3,6). CDEI (8 bp) and CDEIII (26 bp) represent consensus sequences, whereas CDEII (78–86 bp) is an AT-rich sequence that separates CDEI and CDEIII. The CDEI element contains the DNA binding site for the Cbf1 protein. So far, no protein has been reported that specifically binds to the CDEII element although Cse4, a histone H3 homolog (7), has been genetically linked to CDEII (8). CDEII likely contributes to the structure of the centromere with no protein specifically bound to it (9). Deletions or insertions altering the length of the AT-rich CDEII domain decrease mitotic chromosome stability (reviewed in 10). High AT content and specific length rather than nucleotide sequence seem to be critical for CDEII (11). CDEIII is essential for centromere function and specifies the binding site for the multi-subunit protein complex CBF3 (reviewed in 5;12).
Cbf1p is not only involved in kinetochore complex formation at the CDEI site but is also part of a transcription activator complex regulating the biosynthesis of the amino acid l-methionine (13) as well as the transcription activator complexes CYT1 (14), GAL2 and TRP1 (15). Inactivation of Cbf1p leads to a partial loss of centromere function and methionine auxotrophy in S.cerevisiae (13,16). Furthermore, Cbf1p deletion mutants show a 10-fold reduction in chromosome segregation efficiency (15–17). It has been suggested that the major cellular role of Cbf1p is to safeguard the biochemical integrity of the kinetochore (18). Cbf1p consists of 351 amino acids and binds to the CDEI concensus sequence 5′-dRTCACRTG-3′ (R = purine) (19,20) as a homodimer via a basic region helix–loop–helix zipper (bHLHzip) domain at its C-terminal end (15–17). Protein function is mediated by the bHLHzip domain since deletion of the N-terminal 209 amino acids has no effect on chromosome segregation (15). Two α-helical basic regions of bHLH dimers bind into the major grooves of the DNA (for comparison see 21). Cbf1p was shown to bend CDEI upon binding by ∼70° towards the major groove of the CDEI binding site (22).
Polyacrylamide gel electrophoresis (PAGE) revealed different degrees of retarded migration behavior of all 16 CEN DNAs indicating intrinsic curvature (9) and providing further evidence for a model of budding yeast centromeres in which CEN DNA structure is important for the assembly, activity and/or regulation of the centromere protein–DNA complex.
There is increasing genetic and biochemical evidence for a regulated network of protein–protein and protein–DNA interactions that contributes to the structure of the S.cerevisiae kinetochore complex (23). The CBF3 multiprotein complex consists of the four essential subunits: p110, p64, p58 and p23, which form the core element of the centromere (reviewed in 5). In analyses of genetic interactions between centromere protein genes, synthetic lethality was observed between the CEP3 gene encoding p64 and Cbf1. These analyses suggested that Cbf1p may physically interact with p64 in vivo (18,24). This idea was supported by the finding that CBF3 subunits can directly interact with Cbf1p in vitro and that p64 and p23 stimulate the binding of Cbf1p to CEN DNA (25).
In this paper, the apparent affinity constants for the interaction of Cbf1p to all 16 CEN DNAs were determined in saturation binding experiments followed by gel retardation analyses. The data reveal that the affinities of a truncated Cbf1p version carrying only the DNA binding domain (Cbf1ΔN209p) vary in a broad range when compared with full-length Cbf1p. The variations appear to depend on CDEI flanking centromere DNA as well as on the N-terminal part of Cbf1p. The CBF3 subunit p64 stimulates the CEN DNA binding activity of both, Cbf1ΔN209p and the full-length Cbf1p protein indicating that differences in the binding affinities of the two proteins may be overruled through interaction of Cbf1p/CDEI with p64.
MATERIALS AND METHODS
Centromere DNAs
DNA fragments (300 bp) carrying the centromere DNA from each of the 16 yeast chromosomes were cloned recently (9) in pBlueScript II SK+ vectors (Amersham, Freiburg, Germany) yielding the plasmids pCEN1 to pCEN16. The cloned 300 bp fragments comprising CDEI, CDEII and CDEIII are flanked by the centromere-specific 90 or 91 bp of genomic budding yeast DNA (9). The consensus binding site of Cbf1p, 5′-dRTCACRTG-3′, within CDEI is present only once per centromere fragment. The 16 different CEN DNA fragments were amplified by PCR using the primers PH9 (5′-dCGCTCTAGAACTAGTGGATC-3′, 20 bases) and PH10 (5′-TCGAGGTCGACGGTATC-3′, 17 bases) which are complementary to the CEN DNA flanking vector sequences. Thus, the amplified fragments are 337 bp in length. PCR products were purified (QIAquick kit, Qiagen, Hilden, Germany) and analyzed on 4% polyacrylamide gels. DNA purity and concentration were determined in a UV spectrophotometer (Carl Zeiss, Specord M 500, Jena) between 200 and 300 nm wavelength. All fragments were diluted to a concentration of 5 nM and stored at –20°C.
Recombinant proteins
Full-length Cbf1p protein and Cbf1ΔN209p containing only the DNA binding domain of Cbf1p were expressed as histidine-tagged fusion proteins in Escherichia coli BL21(DE3) from plasmids pET-Cbf1p and pET-Cbf1ΔN209, respectively (kindly provided by D.Thomas), and purified according to a protocol detailed in Kuras et al. (26). His-tagged p64 protein was expressed and purified as described previously (27,28). Protein concentrations were determined using the micro BSA protein assay kit (Pierce, USA).
Bandshift assay
The binding constants were determined by bandshift assays in polyacrylamide gels (29). In the reaction mixtures, the amount and concentration of DNA was kept constant while the protein concentration was varied. The binding reactions contained protein at the indicated concentrations, 0.5 nM CEN DNA, 25 mM HEPES buffer pH 7.6, 50% glycerine and 10 mg/ml BSA. Reaction mixtures were kept on ice for 20 min. The reaction mixture was loaded onto 4% native polyacrylamide gels pre-electrophoresed for 1 h. DNA binding stimulation assays were performed as described previously (25). Gels were run in 0.5× TBE (1× TBE contains 100 mM Tris, 83 mM borate, 0.1 mM EDTA pH 8.0) at 20 mA at room temperature and stained in SYBR Gold nucleic acid stain solution (Molecular Probes, Eugene, USA) for 10 min in the dark. The fluorescence of the gel bands was visualized by UV light. Digital pictures were taken with a high resolution CCD camera and stored as tag image files (TIFF). The digital files were analyzed using the gel scan program MacCAM (Cybertec, Berlin, Germany). The intensity of free DNA bands and complex bands in each lane were determined. For calculating the binding constants, the intensities of those bands representing DNA complexes with one or more Cbf1 proteins (multimers) were added. For some CEN DNAs the binding constant of Cbf1p was determined in four completely independent experiments. Deviations of a few percent were observed at concentrations at which most of the DNA is complexed. The deviations of the binding constants from the mean value (for CEN6, mean = 3.69 nM) were <1%.
Data analysis
Free protein Pf binds to free DNA Df to form the complex DP according to:
Df + Pf ↔ DP
For the total concentrations of DNA [Dt] and protein [Pt] holds:
[Dt] = [Df] + [DP] 1
[Pt] = [Pf] + [DP] 2
The dissociation constant Kd is given by:
Kd = [Df][Pf]/[DP] 3
Kd is related to the equilibrium constant KE by:
KE = 1/Kd 4
For Kd we can write:
Kd = ([Dt] – [DP])([Pt] – [DP])/[DP] 5
For equal gel band intensities of free DNA and DNA complexed with protein we have:
[Df] = [DP] 6
Thus:
[DP] = 0.5[Dt] 7
For this protein concentration [Pt,50] we thus have:
Kd = [Pt,50] – 0.5[Dt] 8
For known [Dt] we take [Pt,50] at 50% binding from the binding curve (Fig. 3) and calculate the dissociation constant Kd according to equation 8.
Figure 3.

Binding analysis of Cbf1p and Cbf1ΔN209p to CEN15 DNA. The plot displays the data of gels shown in Figure 2. Triangles, binding values for Cbf1p; squares, binding values for Cbf1ΔN209p.
RESULTS
The centromere binding protein Cbf1p and its deletion variant Cbf1ΔN209p containing only the bHLHzip DNA binding motif (Fig. 1A) were expressed in E.coli and purified. The purity of the two proteins was analysed in denaturing SDS–PAGE indicating a protein purity >95% (Fig. 1B). The binding constants of the full-length protein (His)6-Cbf1p and the deletion mutant (His)6-Cbf1ΔN209 to all 16 CEN DNAs of S.cerevisiae were determined in gel retardation experiments. In binding reactions, 0.5 nM CEN DNA was incubated with increasing amounts of recombinant proteins: 0.625–25 nM (His)6-Cbf1p and 2.82–112.5 nM (His)6-Cbf1ΔN209 (Fig. 2). Binding mixtures were incubated on ice for 20 min and loaded on native polyacrylamide gels, which separate unbound DNA from the protein–DNA complexes. Typical results are displayed for CEN15 DNA in Figure 2. With increasing amounts of protein in the binding mixture, the intensities of the bands representing free DNA decrease and those of the protein–DNA complex bands increase. At high protein concentrations, high molecular weight complexes appear which most likely represent multimere aggregates. The quantitative analyses of the gels in Figure 2 are shown in Figure 3.
Figure 1.
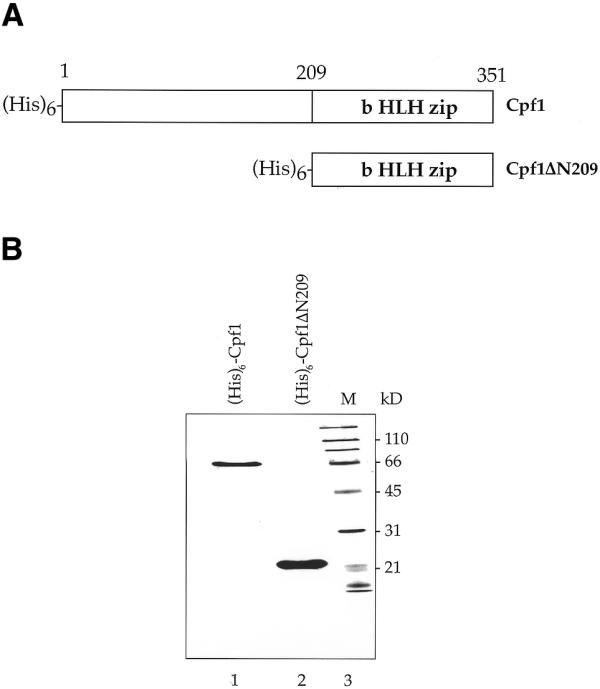
Expression of Cbf1 proteins. (A) Schematic drawing of the two proteins (His)6-Cbf1p and (His)6-Cbf1ΔN209p. The full-length protein is 351 amino acids in length while the deletion variant only contains the C-terminal amino acids 209–351 carrying the bHLHzip DNA binding motif. (B) Denaturing SDS–polyacrylamide gel of (His)6-Cbf1p (lane 1) and (His)6-Cbf1ΔN209p (lane 2). M, marker proteins with molecular weight indicated (kDa).
Figure 2.
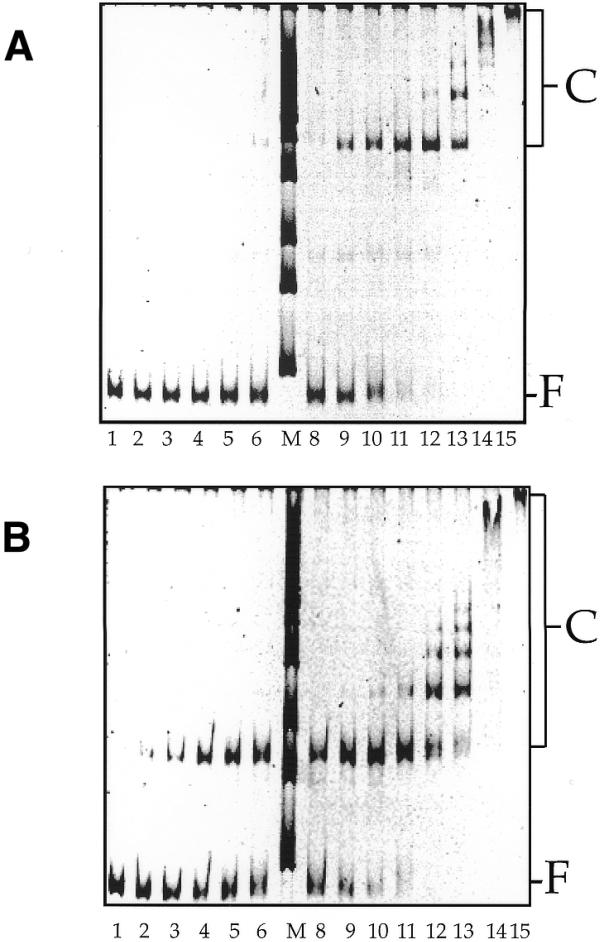
Binding of Cbf1p and Cbf1ΔN209p to CEN DNA. (A) Cbf1p binding. Lanes 1–15, increasing Cbf1p concentrations: lane 1, 0.63 nM; lane 2, 0.89 nM; lane 3, 1.04 nM; lane 4, 1.25 nM; lane 5, 1.39 nM; lane 6, 1.56 nM; lane 8, 1.79 nM; lane 9, 2.08 nM; lane 10, 2.50 nM; lane 11, 3.13 nM; lane 12, 4.17 nM; lane 13, 6.25 nM; lane 14, 12.5 nM; lane 15, 25 nM. (B) Cbf1ΔN209p binding. Lanes 1–15, increasing Cbf1ΔN209p concentrations: lane 1, 1.41 nM; lane 2, 2.01 nM; lane 3, 2.35 nM; lane 4, 2.82 nM; lane 5, 3.13 nM; lane 6, 3.52 nM; lane 8, 4.02 nM; lane 9, 4.69 nM; lane 10, 5.63 nM; lane 11, 7.05 nM; lane 12, 9.37 nM; lane 13, 14.07 nM; lane 14, 28.13 nM; lane 15, 56.25 nM. M, marker DNA in steps of 1 kb; F, free CEN DNA; C, complexed CEN DNA.
The binding constants of the recombinant proteins were deduced from the protein concentration at which 50% DNA was bound, and represent mean values from three to four independent binding curves. Table 1 lists binding constants, DNA binding sequences, relative values of the binding constants for the two proteins and the mean values for the binding constants. The analysis of gel band intensities has particularly small errors for that protein concentration [Pt,50] for which we measure equal distribution of the DNA in the free [Df] and the complex gel band (including multimer complexes, [DP]). For smaller protein concentrations, the band representing the complex becomes faint while at higher protein concentrations we observed multimer complex bands and only a faint band for free DNA. At the protein concentration [Pt,50] we have [Df] = [DP].
Table 1. Binding sequence and dissociation constants of Cbf1p and Cbf1ΔN209p binding to all 16 CEN DNAs of S.cerevisiae.
|
CEN |
CDEI |
Kd ± error Cbf1 [nM] |
Kd ± error Cbf1ΔN209 [nM] |
Kd (Cbf1ΔN209)/Kd (Cbf1) |
| 1 | GTCACATGAC | 1.94 ± 0.16 | 5.45 ± 0.44 | 2.8 |
| 2 | ATCATGTGAC | 3.11 ± 0.25 | 5.02 ± 0.40 | 1.6 |
| 3 | GTCACATGAT | 2.52 ± 0.20 | 3.21 ± 0.26 | 1.3 |
| 4 | GTCACATGCT | 2.43 ± 0.19 | 10.81 ± 0.86 | 4.4 |
| 5 | ATCACGTGCT | 3.35 ± 0.27 | 4.11 ± 0.33 | 1.2 |
| 6 | ATCACGTGCT | 3.69 ± 0.30 | 6.12 ± 0.49 | 1.7 |
| 7 | ATCACGTGTT | 3.27 ± 0.26 | 7.12 ± 0.57 | 2.2 |
| 8 | ATCACATGAC | 3.77 ± 0.30 | 11.48 ± 0.92 | 3.0 |
| 9 | TTCACGTGAA | 2.80 ± 0.22 | 12.11 ± 0.97 | 4.3 |
| 10 | ATCACGTGTT | 1.76 ± 0.14 | 3.78 ± 0.30 | 2.1 |
| 11 | GTCACATGAT | 1.68 ± 0.13 | 5.11 ± 0.41 | 3.0 |
| 12 | ATCACGTGTA | 1.93 ± 0.15 | 3.44 ± 0.28 | 1.8 |
| 13 | ATCACATGAC | 2.43 ± 0.19 | 5.11 ± 0.41 | 2.1 |
| 14 | GTCACGTGCA | 2.85 ± 0.23 | 6.45 ± 0.52 | 2.3 |
| 15 | ATCACGTGAA | 2.43 ± 0.19 | 6.12 ± 0.52 | 2.5 |
| 16 | ATCACATGAT | 2.77 ± 0.22 | 8.80 ± 0.70 | 3.2 |
| Mean value | RTCACRTG | 2.67 | 6.52 | 2.5 |
In addition, the consensus sequence and the mean binding constants of the two proteins to the 16 CEN DNAs are shown. For every CEN DNA the ratio of the two Kds is given.
The binding constants of Cbf1p binding to the 16 CEN DNAs vary between 1.8 nM found for CEN10 and 3.8 nM found for CEN8 (Table 1) with a mean value of 2.7 nM. The binding of the deletion variant Cbf1ΔN209p, however, varies between 3.2 nM found for CEN3 and 12.1 nM found for CEN9 with a mean value of 6.5 nM. This indicates that the N-terminus of Cbf1p contributes to optimal binding to the 16 CDEI elements. While the binding constant of both proteins to CEN3 or CEN5 is similar (Kd difference 20–30%), the binding constant of the two proteins to CEN4 or CEN9 differs by a factor of 4 (Table 1). This result strongly suggests that CDEI flanking CEN DNA contributes to the complex formation between Cbf1p and CDEI.
A previous analysis had shown that the 16 CEN DNAs display different degrees of reduced mobility in native polyacrylamide gels, indicating intrinsic curvature of S.cerevisiae centromeric DNA (9). Since the Cbf1p protein has been shown to bend the CDEI sequence of at least three of the 16 CEN DNAs (22), we tested whether Cbf1p would show any additional effects on the CEN DNA migration behavior (Fig. 4). Bandshift experiments were performed that detected all 16 CEN DNAs and the corresponding Cbf1ΔN209–CEN complex in the same gel (Fig. 4A). The pattern of migration of free CEN DNAs was identical to the one observed previously (9). The Cbf1p–CEN complexes displayed different mobilities. However, the pattern of migration differences of the Cbf1p–CEN complexes was clearly distinct from the pattern of the free CEN DNAs. Pairwise comparison revealed that CEN DNAs that migrated at different positions in their free form were detected at the same position when in complex with Cbf1p (compare CEN4 with CEN5, or CEN14 with CEN15, Fig. 4A), or, vice versa, CEN-DNAs which migrated at similar positions in their free form, were retarded to distinct positions with Cbf1p bound to it (compare CEN8 with CEN9, Fig. 4A). Figure 4B represents an evaluation of the band shift results for each of the CEN DNAs as the ratio between the migration length of the free CEN and the respective Cbf1p–CEN complex. The largest difference (14%) of the complex:CEN migration ratio was observed between CEN5 and CEN12 (Fig. 4B). We obtained similar migration patterns using the full-length Cbf1p derivative indicating that the N-terminal part of Cbf1p does not influence the Cbf1p–CEN structure (data not shown). These results indicate that the altered migration behavior of Cbf1p-complexed CEN may be the result of both Cbf1p-induced bending of CDEI (22) and the intrinsic curvature of the CEN DNA (9).
Figure 4.
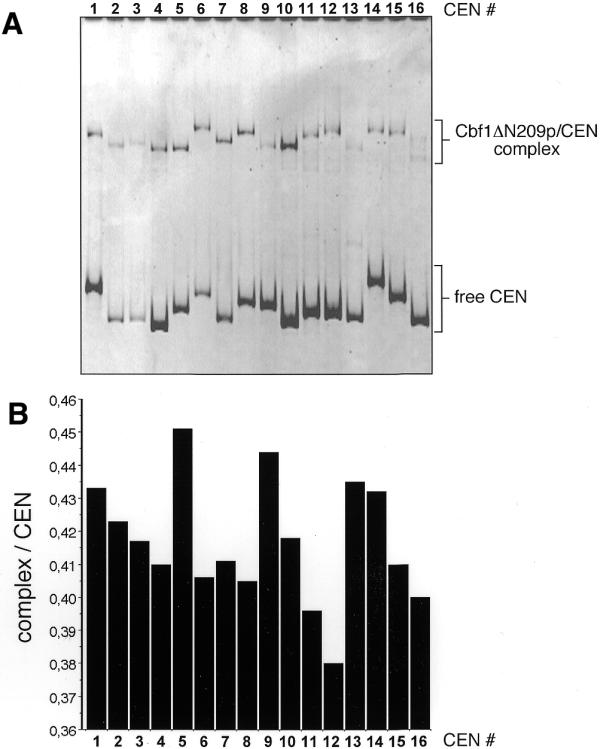
PAGE migration behavior of Cbf1p–CEN complexes. (A) All 16 CEN DNAs were incubated with Cbf1ΔN209p and analysed by native PAGE. (B) The migration length of the free CEN DNA was divided by the migration length of the respective Cbf1ΔN209p–CEN complex and the values are plotted for each CEN DNA. The data were derived from three independent experiments and represent mean values. Standard deviation was <5%.
Components of the essential CBF3 complex can directly interact with Cbf1ΔN209p and increase the affinity of Cbf1ΔN209p for CEN DNA (25). We therefore examined how the addition of p64 would effect the binding of the full-length Cbf1p to CEN DNA. Recombinant p64 was purified from bacteria (Fig. 5A) and showed no intrinsic CEN DNA binding activity (Fig. 5B). We then added p64 to reaction mixtures that contained very low amounts of Cbf1 proteins (Fig. 5C). Although without p64 there was no detectable Cbf1p–CEN9 complex formation (Fig. 5C, lanes 1 and 3), we observed formation of such complexes in the presence of p64 (Fig. 5C, lanes 2 and 4). The p64-induced increase in complex formation between Cbf1ΔN209p and CEN DNA is p64 concentration-dependent (∼40-fold at 20 nm concentration; 25). Figure 5C shows that p64 exhibited a similar DNA binding stimulation activity on Cbf1p (lane 4) indicating that p64 functions on both the Cbf1ΔN209–CEN complex and the Cbf1p–CEN complex with similar activity.
Figure 5.
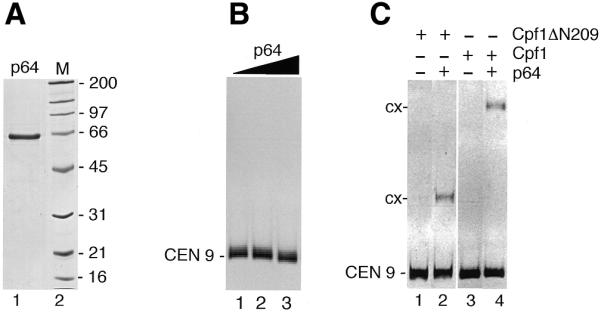
p64-induced stimulation of Cbf1p DNA binding. (A) Coomassie stained SDS–gel of purified, recombinant CBF3 subunit p64 (lane 1). M, molecular weight standard proteins. (B) CEN 9 DNA was incubated with 10, 20 and 40 nM p64, and reaction mixtures analysed by gel shift (lanes 1, 2 and 3, respectively). No complex is observed. (C) CEN 9 DNA was incubated with 0.2 nM of Cbf1p (lanes 1 and 2) or Cbf1ΔN209p (lanes 3 and 4) in the absence (lanes 1 and 3) or presence (lane 2 and 4) of 20 nM p64.
DISCUSSION
We analyzed the binding of the Cbf1p protein and its deletion mutant containing only the DNA binding domain to 16 different centromeric DNA fragments representing a degenerate set of specific binding sites. Each of the 337 bp DNA centromere fragments used in this study contains a single CDEI binding site of the consensus sequence 5′-RTCACRTG-3′ specific for CEN1 to CEN16, respectively. Cbf1p binds to these sites as a homodimer. In addition, the DNA fragments carry the CDEIII region, which represents the binding site of the hetero-oligomeric protein complex Cbf3 (5). The binding constant of Cbf1p to DNA sequences other than the consensus site was determined to be 2 × 10–4 M in competition experiments (26). Assuming 329 independent non-specific binding sites per centromere fragment, one can estimate their contribution to the binding constant as two orders of magnitude off the constants for specific binding (Table 1). Thus, non-specific binding to the fragments can be neglected.
The binding constants of full-length Cbf1p agree with literature values obtained for related proteins. Studies by Baker et al. (30) revealed that the Cbf1p binding equilibrium constant KE to CEN3 DNA is 3 × 108 M–1 (±40%) which corresponds (equation 4) to a Kd of 3.3 nM. Baker et al. (30) had purified the Cbf1p protein from yeast, thus the endogenous protein from yeast exhibits a very similar binding constant when compared to the binding constant of 2.5 nM obtained for the recombinant Cbf1 protein used in this work. The binding constant of the human upstream stimulatory factor USF, also containing a bHLH DNA binding motif, to its binding sequence 5′-dCACGTG-3′ was determined to 1.3 nM (31).
The 16 CEN DNAs offer a variation of binding sequences. It could be speculated that the binding constants are related to the particular CDEI DNA sequence suggesting similar binding constants for those CEN fragments with 5′-dRTCACATG-3′ or with 5′-dRTCACGTG-3′. However, this is not observed. For example, within the CDEI sequence group 5′-dRTCACATG-3′ we find the strongest (CEN11) and the weakest (CEN8) binding by full-length Cbf1p. Very weak (CEN8) and very strong (CEN3) binding within this DNA sequence group is also observed for Cbf1pΔN209p. Thus, DNA sequences next to the consensus binding site seem to influence the DNA binding of Cbf1p and Cbf1ΔN209p. Similar findings have been reported previously by Wilmen et al. (20) who showed that Cbf1p binding in vitro as well as full in vivo centromere function requires a 10 bp recognition sequence including the 8 bp CDEI. Moreover, these studies indicated that Cbf1p interaction with CDEI is different in vitro and in vivo suggesting that the CDEI–Cbf1p complex interacts with other centromere components in the CEN complex (20).
Cbf1p and Cbf1ΔN209p contain the same bHLHzip DNA binding motif. Nevertheless, both proteins bind differently to the 16 DNA sequences indicating that the N-terminal domain of the protein has an influence on binding. The relative value of the binding constants of the two proteins (Table 1) varies from 1.2 (CEN5) to 4.4 (CEN4) showing that the N-terminal part contributes to the binding in a DNA sequence specific way either by interacting with CDEI-surrounding DNA sequences or by modulating the Cbf1 protein structure and thus stabilizing the various interactions between the protein and the 16 different CDEI elements.
Cbf1p and Cbf1ΔN209p bind to the degenerate set of the 16 CEN DNAs with binding constants varying by a factor of 2.2 and 3.8, respectively. Since, in vivo, both the Cbf1p–CDEI and the Cbf1ΔN209p–CDEI complexes exert the same biological function (15), this system gives us an idea of the spectrum of binding strengths of a particular protein–DNA interaction tolerated by the cell. We are not aware of reports on other biological systems with such a DNA sequence and binding strength variation but common function within one cell. It is likely that a variety of protein–protein and protein–DNA interactions are necessary for the formation of fully assembled centromere complexes (23). Thus, the binding strength variations observed in our system represent only one contributing part of the biologically active complex. Obviously, a binding strength variation of a factor of 3.8 appears to be fully tolerated in vivo.
The mobility of DNA molecules in native polyacrylamide gels depends on their 3D structure. All 16 CENDNAs of S.cerevisiae display different degrees of reduced mobility in gels (9). Here we show that all 16 CEN DNA–Cbf1p complexes display a migration pattern that is clearly distinct from the pattern observed for free CEN DNA. It is conceivable that the migration characteristics of the complexes are a result of intrinsic CEN DNA curvature (9) combined with bending of CDEI by Cbf1p (22). The intrinsic curvature of CEN DNAs may also influence the accessibilty of Cbf1 proteins to CDEI, which could also be responsible for the variety of binding constants.
Yeast mutants in which Cbf1 is replaced by Cbf1ΔN209 show no difference in their growth rate when compared to wild-type (32). Obviously, for centromere function the N-terminal protein domain does not seem to be essential and thus for the biological function a considerable difference in the binding constant of Cbf1p to the 16 different DNA sites, as well as differences in the Cbf1p–CEN DNA structure are tolerated in vivo. It was shown previously that binding of Cbf1p to its binding motif in the methionine promoter can be stimulated by the transcription factor Met28 (13). It is possible that similar mechanisms act at centromeres. There are more than 12 established kinetochore proteins contributing to structure and function of the centromeres in budding yeast (23). Based on biochemical and genetic data, the centromere proteins Mif2p, Cse4p, Mcm21p and the Cbf3 subunits p110 and p64 have been proposed as Cbf1p-interacting proteins (18,23,24,33). These proteins may function at the centromere to influence Cbf1p–CDEI complex formation. This has been reported recently for the p64 and p23 CBF3 subunits, which stimulate the DNA binding activity of Cbf1ΔN209p (25). p64 stimulated the CEN DNA binding activity of both Cbf1ΔN209p (25) and full-length Cbf1p with similar efficiencies (Fig. 5). If this interaction does also occur in vivo, we speculate that the observed differences in binding affinities of Cbf1ΔN209p to different CENs are tolerated because p64 shifts the affinity of Cbf1ΔN209p to CEN DNA into a range that makes these differences negligable. Since Cbf1p does not bind to CDEI in the absence of CBF3 in vivo (33), the observed p64 activity may be a critical event that decides whether Cbf1p will bind to CDEI or not.
Taken together the results presented here provide further evidence that the structure of the S.cerevisiae centromere complex is determined by a dynamic network of specific DNA–protein and protein–protein interactions.
Acknowledgments
ACKNOWLEDGEMENTS
The kind gift of the plasmids pET-Cbf1p and pET-Cbf1ΔN209p carrying the genes for the proteins Cbf1p and Cbf1ΔN209p by D.Thomas are gratefully acknowledged. J.Lechner kindly provided plasmid pJL33, which contains the p64 cDNA.
References
- 1.Choo K.H.A. (1997) The Centromere. Oxford University Press, Oxford, UK.
- 2.Lee C., Wevrick,R., Fisher,R.B., Ferguson-Smith,M.A. and Lin,C.C. (1997) Human centromeric DNAs. Hum. Genet., 100, 291–304. [DOI] [PubMed] [Google Scholar]
- 3.Hegemann J.H. and Fleig,U.N. (1993) The centromere of budding yeast. Bioessays, 15, 451–460. [DOI] [PubMed] [Google Scholar]
- 4.Craig J.M., Earnshaw,W.C. and Vagnarelli,P. (1999) Mammalian centromeres: DNA sequence, protein composition and role in cell cycle progression. Exp. Cell Res., 246, 249–262. [DOI] [PubMed] [Google Scholar]
- 5.Lechner J. and Ortiz,J. (1996) The Saccharomyces cerevisiae kinetochore. FEBS Lett., 389, 70–74. [DOI] [PubMed] [Google Scholar]
- 6.Fleig U., Beinhauer,J.D. and Hegemann,J.H. (1995) Functional selection for the centromere DNA from yeast chromosome VIII. Nucleic Acids Res., 23, 922–924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Meluh P.B., Yang,P., Glowczewski,L., Koshland,D. and Smith,M.M. (1998) Cse4p is a component of the core centromere of Saccharomyces cerevisiae. Cell, 94, 607–613. [DOI] [PubMed] [Google Scholar]
- 8.Stoler S., Keith,K.C., Curnick,K.E. and Fitzgerald-Hayes,M. (1995) A mutation in CSE4, an essential gene encoding a novel chromatin-associated protein in yeast, causes chromosome nondisjunction and cell cycle arrest at mitosis. Genes Dev., 9, 573–586. [DOI] [PubMed] [Google Scholar]
- 9.Bechert T., Heck,S., Fleig,U., Diekmann,S. and Hegemann,J.H. (1999) All 16 centromere DNAs from Saccharomyces cerevisiae show DNA curvature. Nucleic Acids Res., 27, 1444–1449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gaudet A. and Fitzgerald-Hayes,M. (1990) In Strauss, P.R. and Wilson,S.H. (eds), The Eukaryotic Nucleus, 2. Telford, NJ.
- 11.Murphy M.R., Fowlkes,D.M. and Fitzgerald-Hayes,M. (1991) Analysis of centromere function in Saccharomyces cerevisiae using synthetic centromere mutants. Chromosoma, 101, 189–197. [DOI] [PubMed] [Google Scholar]
- 12.Lechner J. and Carbon,J. (1991) A 240 kDa multisubunit protein complex, CBF3, is a major component of the budding yeast centromere. Cell, 64, 717–725. [DOI] [PubMed] [Google Scholar]
- 13.Kuras L., Cherest,H., Surdin-Kerjan,Y. and Thomas,D. (1996) A heteromeric complex containing the centromere binding factor 1 and two basic leucine zipper factors, Met4 and Met28, mediates the transcription activation of yeast sulfur metabolism. EMBO J., 15, 2519–2529. [PMC free article] [PubMed] [Google Scholar]
- 14.Oechsner U. and Bandlow,W. (1996) Interactions of the yeast centromere and promoter factor, Cpf1p, with the cytochrome c1 upstream region and functional implications on regulated gene expression. Nucleic Acids Res., 24, 2395–2403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mellor J., Jiang,W., Funk,M., Rathjen,J., Barnes,C.A., Hinz,T., Hegemann,J.H. and Philippsen,P. (1990) CPF1, a yeast protein which functions in centromeres and promoters. EMBO J., 9, 4017–4026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cai M. and Davis,R.W. (1990) Yeast centromere binding protein CBF1, of the helix–loop–helix protein family, is required for chromosome stability and methionine prototrophy. Cell, 61, 437–446. [DOI] [PubMed] [Google Scholar]
- 17.Baker R.E. and Masison,D.C. (1990) Isolation of the gene encoding the Saccharomyces cerevisiae centromere-binding protein CP1. Mol. Cell. Biol., 10, 2458–2467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Baker R.E., Harris,K. and Zhang,K. (1998) Mutations synthetically lethal with cep1 target S.cerevisiae kinetochore components. Genetics, 149, 73–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bram R.J. and Kornberg,R.D. (1987) Isolation of a Saccharomyces cerevisiae centromere DNA-binding protein, its human homolog and its possible role as a transcription factor. Mol. Cell. Biol., 7, 403–409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wilmen A., Pick,H., Niedenthal,R.K., Sen-Gupta,M. and Hegemann,J.H. (1994) The yeast centromere CDEI/Cpf1 complex: differences between in vitro binding and in vivo function. Nucleic Acids Res., 22, 2791–2800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ferre-D’Amare A.R., Prendergast,G.C., Ziff,E.B. and Burley,S.K. (1993) Recognition by Max of its cognate DNA through a dimeric b/HLH/Z domain. Nature, 363, 38–45. [DOI] [PubMed] [Google Scholar]
- 22.Niedenthal R.K., Sen-Gupta,M., Wilmen,A. and Hegemann,J.H. (1993) Cpf1 protein induced bending of yeast centromere DNA element I. Nucleic Acids Res., 21, 4726–4733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ortiz J., Stemmann,O., Rank,S. and Lechner,J. (1999) A putative protein complex consisting of Ctf19, Mcm21 and Okp1 represents a missing link in the budding yeast kinetochore. Genes Dev., 13, 1140–1155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Meluh P.B. and Koshland,D. (1995) Evidence that the MIF2 gene of Saccharomyces cerevisiae encodes a centromere protein with homology to the mammalian centromere protein CENP-C. Mol. Biol. Cell, 6, 793–807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hemmerich P., Stoyan,T., Wieland,G., Koch,M., Lechner,J. and Diekmann,S. (2000) Interaction of yeast kinetochore proteins with centromere-protein/transcription factor cbf1. Proc. Natl Acad. Sci. USA, 97, 12583–12588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kuras L., Barbey,R. and Thomas,D. (1997) Assembly of a bZIP–bHLH transcription activation complex: formation of the yeast Cbf1–Met4–Met28 complex is regulated through Met28 stimulation of Cbf1 DNA binding. EMBO J., 16, 2441–2451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Stemmann O. and Lechner,J. (1996) The Saccharomyces cerevisiae kinetochore contains a cyclin–CDK complexing homologue, as identified by in vitro reconstitution. EMBO J., 15, 3611–3620. [PMC free article] [PubMed] [Google Scholar]
- 28.Lechner J. (1994) A zinc finger protein, essential for chromosome segregation, constitutes a putative DNA binding subunit of the Saccharomyces cerevisiae kinetochore complex, Cbf3. EMBO J., 13, 5203–5211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Neurath M., Ma,X. and Petterson,S. (1997) DNA/Protein-Interaktionen, 1st Edn. Spektrum Akademischer Verlag, Heidelberg.
- 30.Baker R.E., Fitzgerald-Hayes,M. and O’Brien,T.C. (1989) Purification of the yeast centromere binding protein CP1 and a mutational analysis of its binding site. J. Biol. Chem., 264, 10843–10850. [PubMed] [Google Scholar]
- 31.Ferre-D’Amare A.R., Pognonec,P., Roeder,R.G. and Burley,S.K. (1994) Structure and function of the b/HLH/Z domain of USF. EMBO J., 13, 180–189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Masison D.C., O’Connell,K.F. and Baker,R.E. (1993) Mutational analysis of the Saccharomyces cerevisiae general regulatory factor CP1. Nucleic Acids Res., 21, 4133–4141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Meluh P.B. and Koshland,D. (1997) Budding yeast centromere composition and assembly as revealed by in vivo cross-linking. Genes Dev., 11, 3401–3412. [DOI] [PMC free article] [PubMed] [Google Scholar]