

Abstract
IL-17 is the hallmark cytokine of the newly described “Th17” lymphocyte population. The composition, subunit dynamics, and ligand contacts of the IL-17 receptor are poorly defined. We previously demonstrated that the IL-17RA subunit oligomerizes in the membrane without a ligand. In this study, computational modeling identified two fibronectin-III-like (FN) domains in IL-17RA connected by a nonstructured linker, which we predicted to mediate homotypic interactions. In yeast two-hybrid, the membrane-proximal FN domain (FN2), but not the membrane-distal domain (FN1), formed homomeric interactions. The ability of FN2 to drive ligand-independent multimerization was verified by coimmunoprecipitation and fluorescence resonance energy transfer microscopy. Thus, FN2 constitutes a “pre-ligand assembly domain” (PLAD). Further studies indicated that the FN2 linker domain contains the IL-17 binding site, which was never mapped. However, the FN1 domain is also required for high affinity interactions with IL-17. Therefore, although the PLAD is located entirely within FN2, effective ligand binding also involves contributions from the linker and FN1.
The discovery of a novel subset of Th cells termed Th17 has provided new insights regarding the regulation of inflammation. Th17 cells secrete IL-17 (IL-17A), IL-17F, TNF-α, and IL-22 and play crucial roles in host defense and autoimmunity. TGFβ and IL-6 drive the generation of Th17 cells, and expansion of this population requires IL-23 (1). Accumulating evidence supports a causative role for IL-23 and IL-17 in rheumatoid arthritis, colitis, and other autoimmune conditions (2). Accordingly, inhibition of the IL-23/IL-17 axis prevents development of the Th17 lineage or its subsequent inflammatory pathology (3, 4). Thus, blockade of IL-17 is a compelling strategy for treating inflammatory disease (5).
Targeting IL-17 effectively requires a detailed understanding of its receptor. IL-17 signals through IL-17RA, a ubiquitously expressed transmembrane receptor (6). IL-17RA is the founding member of the IL-17R superfamily, members of which share remarkably little homology with other classes of cytokine receptors. Recent evidence indicates that human IL-17 also requires IL-17RC for signal transduction (7). We recently showed that murine IL-17 signals through a complex consisting of at least two IL-17RA subunits, which are preassembled in the membrane in the absence of ligand (8). Notably, ligand-independent oligomerization of receptor subunits is characteristic of other receptor systems, including the erythropoietin receptor and TLRs (9–12). In particular, TNFR superfamily members associate through a “pre-ligand assembly domain” (PLAD),3 located in a cysteine-rich motif that is physically distinct from the ligand-binding domain (9, 13). Preassembly is thought to prevent the formation of nonproductive multimeric complexes. Indeed, dominant interfering mutations in Fas that cause an autoimmune lymphoproliferative syndrome require a functional PLAD to create mixed, nonfunctional trimers (14), and a TNFR homologue in poxviruses prevents the apoptotic function of TNF-α via a conserved viral PLAD (15). Strikingly, a soluble peptide encoding a TNFR PLAD inhibits TNF-α-mediated pathology in murine arthritis models, suggesting that PLAD domains are valid targets for anticytokine therapy (16).
Although IL-17RA bears no sequence homology to the TNFR, our finding that IL-17RA subunits multimerize in the absence of ligand predicted the existence of a functional PLAD in IL-17RA (8). Accordingly, in this study we describe a fibronectin III-like (FN) domain within IL-17RA that mediates homomeric interactions. The IL-17 binding site was also delineated in this study. This is the first structure-function analysis of any IL-17R superfamily domain, and the discovery of an IL-17RA PLAD reveals a potential new avenue of anticytokine therapeutics.
Materials and Methods
Molecular modeling
Using the feature aligner at SWISS-PROT, the extracellular sequence of IL-17RA was submitted to PHYRE (17). Coordinates were manipulated on an SGI Tezro visualization workstation with Accelrys Insight II software.
Plasmids, cell culture, luciferase assays, yeast two-hybrid (Y2H), and immunoprecipitations
IL-17RA constructs fused to YFP and CFP were as described (8), and IL-17RAΔ terminates at residue 526 (8). HEK293 and IL-17RAKO (knockout) cells were maintained in αMEM (Sigma-Aldrich) with 10% FBS and antibiotics. Cells were transfected with FuGENE 6 (Roche) and selected in G418 or hygromycin B. Cytokines were from R&D Systems or Peprotech and used at 100–200 ng/ml (IL-17 and IL-17F) or 2 ng/ml (TNF-α). Luciferase assays were performed with a 24p3-Luc reporter (18). For Y2H, IL-17RA extracellular domain (ECD) constructs were fused to the Gal4 DNA binding domain or activation domain in the pGBKT7 and pGADT7 plasmids (BD Clontech). Saccharomyces cerevisiae was transformed by standard methods. For immunoprecipitation, cell lysates were prepared in 1% Nonidet P-40 buffer as described (19), immunoprecipitated with anti-hemagglutinin (HA) (Roche) or M177 (Amgen) and HA-Sepharose, and Western blots were probed with anti-GFP (Santa Cruz Biotechnology) or anti-HA.
Flow cytometry and fluorescence resonance energy transfer (FRET)
Cells were stained with M750 or M177 anti-IL-17RA and anti-rat PE (BD Pharmingen). IL17.Fc was prepared and used as described (19), and endogenous human IL-17RA was blocked with anti-human IL-17RA (M202). FRET data were obtained from membrane regions from three channel images (Zeiss AIM software) (8, 20). Efficiencies were calculated by normalized FRET (N-FRET) (20, 21).
Results
Identification of the IL-17RA PLAD
To gain insight into the structure of murine IL-17RA we used the PHYRE modeling program, which incorporates multiple structure prediction algorithms (17). Extending a prior report (22), this analysis predicted two FN domains (FN1, residues 69–183; FN2, residues 205–282) joined by a nonstructured linker (residues 184–204) (Fig. 1A). A similar structure was obtained for human IL-17RA (data not shown). To evaluate the functional significance of the FN and linker regions in self-association, we used Y2H (Table I and Fig. 1B). Productive associations were observed between full length (FL) IL-17RA ECD and the FN2 or FN2linker (Table I, rows C and D), but not between FL and FN1 (Table I, row B). Consistently, FN2linker and FN2 were both capable of self-association (Table I, rows I–L) but did not interact with FN1 (Table I, rows F–H). The FN2linker construct interacted with itself or FN2 somewhat more strongly than FN2 alone (Table I, rows I–L), while FN1 was not capable of self-association (Table I, row E). Together, these results suggest that FN2 and not FN1 mediates oligomerization of IL-17RA. Although the inter-FN linker may stabilize interactions, it is not essential for FN2 dimerization.
FIGURE 1.

The FN2 domain of IL-17RA mediates ligand-independent assembly. A, Predicted structure of IL-17RA FN domains. PHYRE predicted two FN domains within the mouse IL-17RA ECD. Yellow, β-Sheets; red, α-helices; green, unstructured loops; blue, turns. Sequences of each domain are shown. B, Schematic diagram of IL-17RA FRET and Y2H constructs. SEFIR is a major signaling domain in IL-17RA (22). AD, Activation domain; BD, binding domain. C, FN2linker drives ligand-independent association in cells. HEK293 cells expressing the indicated combinations of IL-17RA constructs fused to CFP or YFP were assayed for N-FRET in the absence (open bars) or presence of IL-17 (filled bars) or IL-17F (gray bar) for 10 min. Significance was assessed by t test, p < 0.05; n.s., not significant. D, Representative images of IL-17RAΔ/CFP plus IL-17RAΔFN2linker/YFP. CFP, YFP, and FRET emissions are shown. Unstim., Unstimulated. E, IL-17RA can co-IP with IL-17RAΔFN2linker. HEK293 cells expressing IL-17RAΔFN2linker/YFP were transiently transfected with IL-17RA.HA. Cells were lysed and immunoprecipitated (IP) with anti-HA or M177 anti-IL-17RA Abs and immunoblotted with Abs to GFP (which cross-react with YFP) or HA. Note that we always observe slight cross-reactivity of anti-HA Abs with YFP and CFP for unknown reasons. W, Western blot.
Table I.
Homotypic interactions between IL-17RA occur via the FN2 domaina
| pGBKT7 (BD Construct) | pGADT7 (AD Construct) | Growth on Leu/Trp | Growth on Leu/Trp/Ade | |
|---|---|---|---|---|
| A | None | None | − | − |
| B | FL | FN1 | ++++ | − |
| C | FL | FN2linker | ++++ | ++++ |
| D | FL | FN2 | ++++ | ++ |
| E | FN1 | FN1 | ++++ | − |
| F | FN1 | FN2linker | ++++ | − |
| G | FN2 | FN1 | ++++ | − |
| H | FN2linker | FN1 | ++++ | − |
| I | FN2linker | FN2linker | ++++ | ++++ |
| J | FN2linker | FN2 | ++++ | +++ |
| K | FN2 | FN2linker | ++++ | +++ |
| L | FN2 | FN2 | ++++ | ++ |
| M | CARMA1 | CARMA1 | ++++ | ++++ |
FL IL-17RA or the FN1, FN2linker, or FN2 domains were fused to Gal4 DNA binding domain (BD) or activation domain (AD). None of the constructs alone activated transcription in yeast (not shown). S. cerevisiae was transformed with the indicated plasmids, and growth on Leu-Trp-deficient plates or Leu-Trp-Ade-deficient plates was evaluated. CARMA1 served as a positive control (33). −, <5 colonies; ++, 20–50 colonies; +++, 100–200 colonies; ++++, >500 colonies.
To substantiate these data in a cellular setting, we performed FRET and coimmunoprecipitation (co-IP) studies in HEK293 cells. IL-17RA constructs (Fig. 1B) with an intact ECD or containing only the FN2linker were fused to CFP and YFP (for convenience of expression, the cytoplasmic tail was truncated at residue 526, termed “IL-17RAΔ”; in prior studies, FL and IL-17RAΔ showed identical FRET properties) (8). Consistent with our previous findings (8), coexpression of IL-17RAΔ/CFP and IL-17RAΔ/YFP resulted in significant FRET efficiency compared with IL-17RAΔ paired with a noninteracting TNFR subunit. Moreover, FRET between IL-17RAΔ chains was reduced upon the addition of ligand (Fig. 1C) (8). Similarly, pairing IL-17RAΔ/ CFP with IL-17RAΔFN2linker/YFP showed a FRET signal that was reduced after treatment with IL-17 or IL-17F (Fig. 1, C and D), indicating that the loss of a single FN1 domain within the receptor complex does not detectably modify subunit interactions (see Fig. 3). Unexpectedly, cells co expressing IL-17RAΔFN2linker/CFP and IL-17RAΔFN2linker/YFP showed a reproducible increase in FRET after IL-17 treatment (Fig. 1C), suggesting that a major alteration in the relative subunit configuration occurs when both FN1 domains are absent (see Fig. 3). There was no change in FRET in cells expressing IL-17RAΔ/CFP with IL-17RAΔFN2/YFP (which lacks the linker) after ligand treatment, although the baseline FRET was fairly high. Because N-FRET calculations are relative values, these results do not prove absolutely that the FN2 site alone is sufficient to drive receptor assembly in cells but are consistent with this hypothesis. Finally, to test biochemical association, HEK293 cells stably expressing IL-17RAΔFN2linker/YFP were transiently transfected with a full-length IL-17RA construct fused to HA. Using anti-HA or M177 Abs, which recognize FL IL-17RA but not the IL-17RAΔFN2linker, we could demonstrate co-IP of the FL IL-17RA with the IL-17RAΔFN2linker (Fig. 1E). Together, all these approaches suggest that the FN2 domain, probably without the linker, mediates ligand-independent IL-17RA assembly. In addition, the FN1 domain apparently serves to modulate dynamic interactions between IL-17RA subunits.
FIGURE 3.
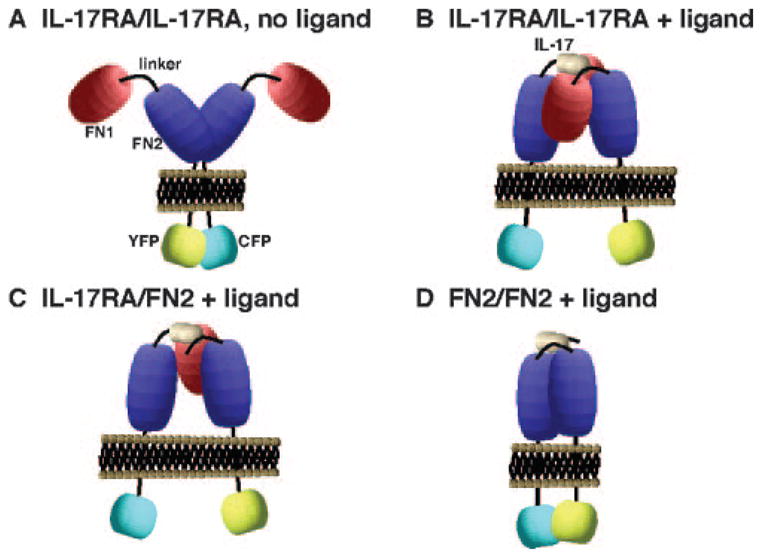
Model of IL-17RA subunit dynamics. Data suggest that the cytoplasmic tails of IL-17RA are held in proximity before ligand binding (A), but separate upon binding IL-17 (B). Based on FRET, at least one FN1 domain is sufficient to mediate ligand-induced subunit reconfiguration (C). However, in the absence of both FN1 domains the cytoplasmic tails show increased association (D). IL-17 requires the FN2 and linker regions to bind the receptor but may also contact FN1 (B–D).
The IL-17 ligand-binding site requires FN2 and the linker domain
The location of the IL-17 binding site on IL-17RA has not been reported. Because IL-17 triggered a change in FRET in cells expressing two IL-17RAΔFN2linker constructs (Fig. 1C), the FN2linker region appeared to encode at least a minimal ligand binding site. To determine requirements for IL-17-mediated signaling, IL-17RAFN2linker and IL-17RAFN1linker constructs (encoding FL cytoplasmic tails; Fig. 1B) were cotransfected into IL-17RA−/− fibroblasts together with an IL-17-responsive luciferase reporter (23). IL-17RAFN2linker, but not IL-17RAFN1linker, mediated IL-17-inducible signaling over a wide dose range (Fig. 2A, left panel and data not shown). Although the signal for FN2linker was reproducibly weaker than that of the wild type, these data nonetheless indicate that the FN1 domain is not essential, nor is the linker sufficient, for signaling. However, the linker appears to be critical for signaling, as IL-17RAFN2 reproducibly failed to mediate IL-17-dependent gene activation (Fig. 2A, right panel).
FIGURE 2.
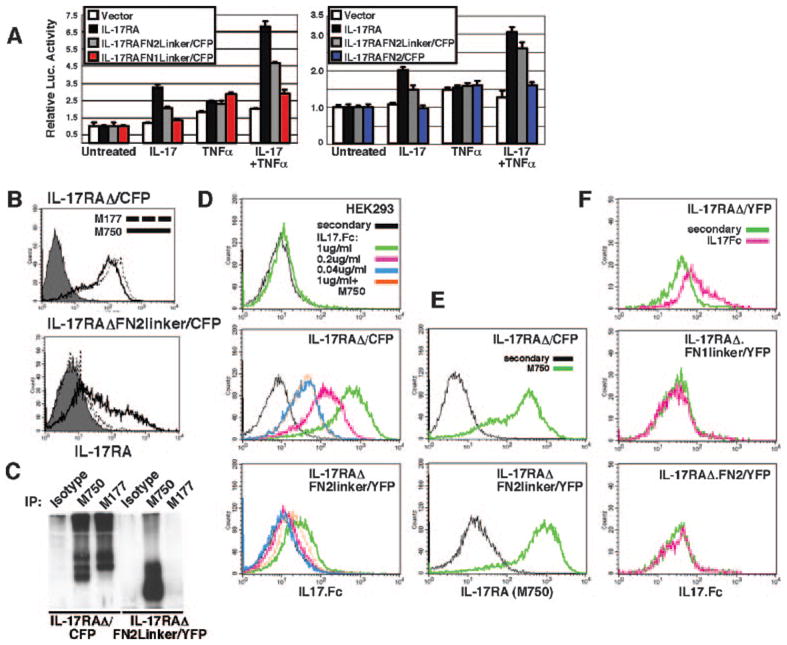
Requirements for IL-17 signaling and binding. A, The FN1 domain is dispensable but the linker is required for IL-17-dependent signaling. IL-17RA−/−fibroblasts were transfected in triplicate with the indicated IL-17RA constructs (with FL cytoplasmic tails) and 24p3-Luc (18). Cells were stimulated with IL-17 (200 ng/ml) and/or TNF-α (2 ng/ml). After 6 h, luciferase activity was determined and normalized to Renilla luciferase. B and C, IL-17RA-neutralizing Abs bind the FN2linker domain. B, HEK293 cells stably transfected with IL-17RΔ/CFP (top) or transiently transfected with IL-17RAΔFN2linker/CFP (bottom) were incubated with a non-neutralizing (clone M177) or neutralizing Ab (clone M750) to murine IL-17RA. Filled histograms are isotype controls. C, Whole cell lysates from HEK293 cells stably expressing IL-17RAΔ/CFP or IL-17RAΔFN2linker/YFP were immunoprecipitated (IP) with M750 or M177 as indicated and blotted with anti-GFP Abs. D and E, FN2linker has a reduced affinity for IL-17. HEK293 cells stably expressing IL-17RAΔ/CFP or IL-17RAΔFN2linker/YFP were stained with various concentrations of IL17.Fc after blocking endogenous human IL-17RA (D) or with M750 (E). F, The linker is required but not sufficient for IL-17 binding. HEK293 cells transiently transfected with IL-17RAΔ/YFP, IL-17RAΔFN1linker/YFP, or IL-17RAΔFN2/YFP were stained with IL17.Fc and gated on YFP-positive cells.
To evaluate the IL-17 interaction site further, cells expressing IL-17RAΔ or IL-17RAΔFN2linker were stained with two Abs to murine IL-17RA. As expected, both Abs bound to IL-17RAΔ, where the entire ECD is preserved (Fig. 2B, top panel). A neutralizing Ab to IL-17RA (clone M750) also stained cells expressing IL-17RAΔFN2linker, whereas a non-neutralizing Ab (clone M177) did not (Fig. 2B, bottom panel), consistent with an IL-17 binding site within the FN2linker region. Consistently, M750 immunoprecipitated both receptor constructs, whereas M177 only pulled down IL-17RAΔ (Fig. 2C; see Fig. 1E). To test the binding directly, cells stably expressing IL-17RAΔ or IL-17RAΔFN2linker were stained with IL-17 fused to Fc (IL17.Fc). Only a weak fluorescent signal was observed in cells expressing IL-17RAΔFN2linker compared with cells with IL-17RAΔ (Fig. 2D) despite an equivalent expression of both constructs (Fig. 2E). Binding was dose dependent and could be inhibited with the neutralizing anti-IL-17RA Ab M750 (Fig. 2D). To assess whether FN1linker or FN2 could bind to IL-17, cells were transiently transfected with these constructs and stained with IL17.Fc. The stable cell lines expressed higher and more uniform levels of IL-17RAΔ than transiently transfected cells (Fig. 2, D vs F); however, there was no detectable binding of IL-17 to cells expressing IL-17RAΔFN1linker or IL-17RAΔFN2 (Fig. 2F) despite similar expression of each construct (not shown). These data indicate a requirement for FN2 and the linker for ligand binding, consistent with the FRET data where the FN2linker, but not FN2 alone, showed ligand-inducible changes in FRET (Fig. 1C). Moreover, FN1 appears to enhance ligand binding substantially but cannot bind IL-17 alone or mediate signaling (Fig. 2, A and F).
In summary, the IL-17RA PLAD appears to be located in the FN2 region of IL-17RA, whereas IL-17 binding also involves the linker and FN1 domain. Despite the ability of IL-17 to signal in the absence of FN1 (as in IL-17RAFN2linker), a complex lacking both FN1 domains is unable to bind IL-17 efficiently or induce the same conformational alteration as an intact IL-17RA ECD. Although a complete picture of IL-17R dynamics will await structural determinations, a model that fits these data is presented in Fig. 3 (8).
Discussion
Anti-cytokine receptor drugs have had a tremendous impact on treatment of autoimmunity, but not all patients respond to these drugs (24). A fundamental understanding of receptor structure can aid in developing novel therapeutics. A case in point is the TNFR, where mutational analysis defined an extra-cellular “pre-ligand assembly domain” or PLAD responsible for subunit trimerization (9). In models of arthritis, a peptide encoding the TNFR PLAD can mitigate disease (16). IL-17RA is considered a compelling target for autoimmunity (5), and the identification of a PLAD in IL-17RA sets the stage for developing analogous inhibitors of IL-17.
Computational analysis of IL-17RA predicted two FN domains joined by a flexible linker. FN domains are characteristic of class I and class II cytokine receptors where they play important roles in oligomerization and ligand binding (25–27). Y2H, co-IP, and FRET microscopy showed that the FN2 region mediates ligand-independent receptor multimerization, whereas the linker and the FN1 region do not seem to be essential in this regard (Table I and Fig. 1). Although FN1 is dispensable for preassembly, it appears to help mediate conformational alterations in the complex that follow ligand binding. Specifically, an increased FRET signal occurred in cells expressing two IL-17RAΔFN2linker constructs after treatment with IL-17, in contrast to the decreased FRET observed in receptor pairings that contain at least one intact IL-17RA ECD (Fig. 1, C and D) (8). Therefore, FN1 may act by reconfiguring the relative positions of IL-17RA subunits, perhaps to permit recruitment of an additional subunit to the complex. There is precedent for ligands to induce large movements in the constituent subunits of cytokine receptors, such as the erythropoietin receptor and TLR9 (12, 28). Interestingly, only one FN1 domain seems to be required for this event, because IL-17RAΔ paired with IL-17RAΔFN2linker showed the same decrease in FRET as two IL-17RAΔ subunits (Fig. 1). Although these studies focused only on IL-17RA, subunit reconfiguration in the IL-17 receptor complex likely involves IL-17RC, a necessary component of the human IL-17R (7). This is a potentially complicated issue, as there are many splice forms of IL-17RC that are poorly characterized in the mouse (29).
Another finding to emerge from these studies was that the FN2linker encodes the IL-17 binding site, which has never been mapped (Fig. 2). Based on other cytokine receptors, the inter-FN linker is likely to contain the actual ligand contact residues (30). Although the FN1linker cannot mediate signaling or bind IL-17 (Fig. 2A, F), the FN1 domain may function as a ligand affinity modulator, analogous to IL-2Rαin the IL-2 receptor (31). Indeed, IL-17-dependent signaling in the absence of FN1 is considerably reduced (Fig. 2A), and IL-17 binding to a receptor lacking FN1 is severely compromised (Fig. 2D). Therefore, it is possible that FN1 interacts with IL-17RC or directly with a ligand.
Interestingly, these data indicate that the PLAD and ligand binding domains are not separable, as they both involve FN2. In contrast, the TNFR PLAD and ligand binding domains are located on different cysteine-rich domains; however, ligand binding is nonetheless dependent on the presence of an intact TNFR PLAD (32). Thus, the PLAD domains in the TNFR and IL-17RA are functionally if not structurally similar.
In summary, this is the first detailed mapping of the IL-17RA extracellular domain, and this work may set the stage for future understanding and therapeutic manipulation of this unique receptor family.
Acknowledgments
We thank F. K. M. Chan for TNFR plasmids and critical reading.
Footnotes
S.L.G. was supported by the National Institutes of Health Grants AR05048 and AI49329. J.M.K. was supported by National Institutes of Health Grant DE14831. J.P.M., was supported by National Institutes of Health Training Grant DE07034 to the Department of Oral Biology. N.I. and T.J. are supported by National Institute of Allergy and Infectious Diseases intramural funding.
Abbreviations used in this paper: PLAD, pre-ligand assembly domain; CFP, cyan fluorescent protein; co-IP, coimmunoprecipitation; ECD, extracellular domain; FL, full length; FN, fibronectin III-like; FRET, fluorescence resonance energy transfer; HA, hemagglutinin; N-FRET, normalized FRET; YFP, yellow fluorescent protein; Y2H, yeast two-hybrid.
Disclosures
The authors have no financial conflict of interest.
References
- 1.Weaver CT, Hatton RD, Mangan PR, Harrington LE. IL-17 family cytokines and the expanding diversity of effector T cell lineages. Annu Rev Immunol. 2007;25:821–852. doi: 10.1146/annurev.immunol.25.022106.141557. [DOI] [PubMed] [Google Scholar]
- 2.Cua DJ, Kastelein RA. TGF-β, a “double agent” in the immune pathology war. Nat Immunol. 2006;7:557–559. doi: 10.1038/ni0606-557. [DOI] [PubMed] [Google Scholar]
- 3.Chen Y, Langrish CL, McKenzie B, Joyce-Shaikh B, Stumhofer JS, McClanahan T, Blumenschein W, Churakovsa T, Low J, Presta L, et al. Anti-IL-23 therapy inhibits multiple inflammatory pathways and ameliorates autoimmune encephalomyelitis. J Clin Invest. 2006;116:1317–1326. doi: 10.1172/JCI25308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lubberts E, Koenders MI, van den Berg WB. The role of T cell interleukin-17 in conducting destructive arthritis: lessons from animal models. Arthritis Res Ther. 2005;7:29–37. doi: 10.1186/ar1478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kikly K, Liu L, Na S, Sedgwick JD. The IL-23/Th(17) axis: therapeutic targets for autoimmune inflammation. Curr Opin Immunol. 2006;18:670–675. doi: 10.1016/j.coi.2006.09.008. [DOI] [PubMed] [Google Scholar]
- 6.Weaver CT, Harrington LE, Mangan PR, Gavrieli M, Murphy KM. Th17: an effector CD4 T cell lineage with regulatory T cell ties. Immunity. 2006;24:677–688. doi: 10.1016/j.immuni.2006.06.002. [DOI] [PubMed] [Google Scholar]
- 7.Toy D, Kugler D, Wolfson M, Vanden Bos T, Gurgel J, Derry J, Tocker J, Peschon J. Cutting edge: interleukin 17 signals through a heteromeric receptor complex. J Immunol. 2006;177:36–39. doi: 10.4049/jimmunol.177.1.36. [DOI] [PubMed] [Google Scholar]
- 8.Kramer JM, Yi L, Shen F, Maitra A, Jiao X, Jin T, Gaffen SL. Cutting edge: evidence for ligand-independent multimerization of the IL-17 receptor. J Immunol. 2006;176:711–715. doi: 10.4049/jimmunol.176.2.711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chan FK, Chun HJ, Zheng L, Siegel RM, Bui KL, Lenardo MJ. A domain in TNF receptors that mediates ligand-independent receptor assembly and signaling. Science. 2000;288:2351–2354. doi: 10.1126/science.288.5475.2351. [DOI] [PubMed] [Google Scholar]
- 10.Constantinescu SN, Keren T, Socolovsky M, Nam H, Henis YI, Lodish HF. Ligand-independent oligomerization of cell-surface erythropoietin receptor is mediated by the transmembrane domain. Proc Natl Acad Sci USA. 2001;98:4379–4384. doi: 10.1073/pnas.081069198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gadd SL, Clevenger CV. Ligand-independent dimerization of the human prolactin receptor isoforms: functional implications. Mol Endocrinol. 2006;20:2734–2746. doi: 10.1210/me.2006-0114. [DOI] [PubMed] [Google Scholar]
- 12.Latz E, Verma A, Visintin A, Gong M, Sirois CM, Klein DC, Monks BG, McKnight CJ, Lamphier MS, Duprex WP, Espevik T, Golenbock DT. Ligand-induced conformational changes allosterically activate Toll-like receptor 9. Nat Immunol. 2007;8:772–779. doi: 10.1038/ni1479. [DOI] [PubMed] [Google Scholar]
- 13.Chan FK. Three is better than one: pre-ligand receptor assembly in the regulation of TNF receptor signaling. Cytokine. 2007;37:101–107. doi: 10.1016/j.cyto.2007.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Siegel RM, Frederiksen JK, Zacharias DA, Chan FK, Johnson M, Lynch D, Tsien RY, Lenardo MJ. Fas preassociation required for apoptosis signaling and dominant inhibition by pathogenic mutations. Science. 2000;288:2354–2357. doi: 10.1126/science.288.5475.2354. [DOI] [PubMed] [Google Scholar]
- 15.Sedger LM, Osvath SR, Xu XM, Li G, Chan FK, Barrett JW, McFadden G. Poxvirus tumor necrosis factor receptor (TNFR)-like T2 proteins contain a conserved preligand assembly domain that inhibits cellular TNFR1-induced cell death. J Virol. 2006;80:9300–9309. doi: 10.1128/JVI.02449-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Deng GM, Zheng L, Chan FK, Lenardo M. Amelioration of inflammatory arthritis by targeting the pre-ligand assembly domain of tumor necrosis factor receptors. Nat Med. 2005;11:1066–1072. doi: 10.1038/nm1304. [DOI] [PubMed] [Google Scholar]
- 17.Jefferys BR, Kelley LA, Sergot MJ, Fox J, Sternberg MJ. Capturing expert knowledge with argumentation: a case study in bioinformatics. Bioinformatics. 2006;22:924–933. doi: 10.1093/bioinformatics/btl018. [DOI] [PubMed] [Google Scholar]
- 18.Shen F, Ruddy MJ, Plamondon P, Gaffen SL. Cytokines link osteo-blasts and inflammation: microarray analysis of interleukin-17- and TNF-α-induced genes in bone cells. J Leukocyte Biol. 2005;77:388–399. doi: 10.1189/jlb.0904490. [DOI] [PubMed] [Google Scholar]
- 19.Maitra A, Shen F, Hanel W, Mossman K, Tocker J, Swart D, Gaffen SL. Distinct functional motifs within the IL-17 receptor regulate signal transduction and target gene expression. Proc Natl Acad Sci USA. 2007;104:7506–7511. doi: 10.1073/pnas.0611589104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Xu X, Meier-Schellersheim M, Jiao X, Nelson LE, Jin T. Quantitative imaging of single live cells reveals spatiotemporal dynamics of multistep signaling events of chemoattractant gradient sensing in Dictyostelium. Mol Biol Cell. 2005;16:676–688. doi: 10.1091/mbc.E04-07-0544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yi L, Fang J, Isik N, Chim J, Jin T. HIV gp120-induced interaction between CD4 and CCR5 requires cholesterol-rich microenvironments revealed by live cell fluorescence resonance energy transfer imaging. J Biol Chem. 2006;281:35446–35453. doi: 10.1074/jbc.M607302200. [DOI] [PubMed] [Google Scholar]
- 22.Novatchkova M, Leibbrandt A, Werzowa J, Neubuser A, Eisenhaber F. The STIR-domain superfamily in signal transduction, development and immunity. Trends Biochem Sci. 2003;28:226–229. doi: 10.1016/S0968-0004(03)00067-7. [DOI] [PubMed] [Google Scholar]
- 23.Shen F, Hu Z, Goswami J, Gaffen SL. Identification of common transcriptional regulatory elements in interleukin-17 target genes. J Biol Chem. 2006;281:24138–24148. doi: 10.1074/jbc.M604597200. [DOI] [PubMed] [Google Scholar]
- 24.Feldmann M, Steinman L. Design of effective immunotherapy for human autoimmunity. Nature. 2005;435:612–619. doi: 10.1038/nature03727. [DOI] [PubMed] [Google Scholar]
- 25.Bazan JF. Structural design and molecular evolution of a cytokine receptor superfamily. Proc Natl Acad Sci USA. 1990;87:6934–6938. doi: 10.1073/pnas.87.18.6934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ruoslahti E. Fibronectin and its receptors. Annu Rev Biochem. 1988;57:375–413. doi: 10.1146/annurev.bi.57.070188.002111. [DOI] [PubMed] [Google Scholar]
- 27.Wijdenes J, Heinrich PC, Muller-Newen G, Roche C, Gu ZJ, Clement C, Klein B. Interleukin-6 signal transducer gp130 has specific binding sites for different cytokines as determined by antagonistic and agonistic anti-gp130 monoclonal antibodies. Eur J Immunol. 1995;25:3474–3481. doi: 10.1002/eji.1830251240. [DOI] [PubMed] [Google Scholar]
- 28.Livnah O, Stura EA, Middleton SA, Johnson DL, Jolliffe LK, Wilson IA. Crystallographic evidence for preformed dimers of erythropoietin receptor before ligand activation. Science. 1999;283:987–990. doi: 10.1126/science.283.5404.987. [DOI] [PubMed] [Google Scholar]
- 29.Haudenschild DR, Curtiss SB, Moseley TA, Reddi AH. Generation of interleukin-17 receptor-like protein (IL-17RL) in prostate by alternative splicing of RNA. Prostate. 2006;66:1268–1274. doi: 10.1002/pros.20422. [DOI] [PubMed] [Google Scholar]
- 30.Murphy JM, I, Young G. IL-3, IL-5, and GM-CSF signaling: crystal structure of the human β-common receptor. Vitam Horm. 2006;74:1–30. doi: 10.1016/S0083-6729(06)74001-8. [DOI] [PubMed] [Google Scholar]
- 31.Gaffen SL. Signaling domains of the interleukin-2 receptor. Cytokine. 2001;14:63–77. doi: 10.1006/cyto.2001.0862. [DOI] [PubMed] [Google Scholar]
- 32.Chan FK. The pre-ligand binding assembly domain: a potential target of inhibition of tumour necrosis factor receptor function. Ann Rheum Dis. 2000;59(Suppl 1):i50–i53. doi: 10.1136/ard.59.suppl_1.i50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Tanner MJ, Hanel W, Gaffen SL, Lin X. CARMA1 Coiled-coil domain is involved in the oligomerization and subcellular localization of CARMA1, and is required for T cell receptor-induced NF-κB activation. J Biol Chem. 2007;282:17141–17147. doi: 10.1074/jbc.M700169200. [DOI] [PubMed] [Google Scholar]