


Abstract
We have previously isolated the hpttg proto-oncogene, which is expressed in normal tissues containing proliferating cells and in several kinds of tumors. In fact, expression of hPTTG correlates with cell proliferation in a cell cycle-dependent manner. Recently it was reported that PTTG is a vertebrate analog of the yeast securins Pds1 and Cut2, which are involved in sister chromatid separation. Here we show that hPTTG binds to Ku, the regulatory subunit of the DNA-dependent protein kinase (DNA-PK). hPTTG and Ku associate both in vitro and in vivo and the DNA-PK catalytic subunit phosphorylates hPTTG in vitro. Furthermore, DNA double-strand breaks prevent hPTTG–Ku association and disrupt the hPTTG–Ku complexes, indicating that genome damaging events, which result in the induction of pathways that activate DNA repair mechanisms and halt cell cycle progression, might inhibit hPTTG–Ku interaction in vivo. We propose that hPTTG might connect DNA damage-response pathways with sister chromatid separation, delaying the onset of mitosis while DNA repair occurs.
INTRODUCTION
The pituitary tumor-transforming gene (pttg) was first isolated from clonal rat GH4 pituitary tumor cells by differential display (1), but no homology with known protein sequences or common functional motifs was found. However, overexpression of pttg in NIH 3T3 fibroblasts showed that it induces cell transformation in vitro, and injection of these transfected cells into athymic nude mice resulted in tumor formation in all animals (1). The pttg-transfected NIH 3T3 cells also stimulated expression and secretion of basic fibroblast growth factor, a human pituitary tumor growth-regulating factor (2). Nevertheless, we found that the human counterpart of PTTG, hPTTG (which was firstly isolated by us; 3), is also expressed in other non-pituitary tumors. Samples from patients with leukemia, lymphoma or myelodysplastic diseases (3), as well as adenocarcinomas of mammary and pulmonary origins (4) showed strong hpttg expression. Our further studies demonstrate a correlation between hpttg expression and cell proliferation (5). The hPTTG protein level is up-regulated in rapidly proliferating cells, is down-regulated in response to serum starvation or cell confluence and is regulated in a cell cycle-dependent manner peaking in mitosis. In addition, hPTTG is phosphorylated by CDC2 during mitosis.
hPTTG is a protein of 202 residues partially localized in the nucleus that has a basic N-terminal portion and an acidic C-terminal portion (3). It has several putative SH3-binding sites (3) and a conserved motif localized in the N-terminal portion matches the destruction box (D-box) shared by many anaphase-promoting complex or cyclosome (APC/C) substrates (6). Recently, PTTG has been shown to be involved in sister chromatid separation (7). Precise coordination of multiple cell cycle events is required to ensure proper mitosis. Chromosome cohesion must be maintained until all chromosomes are attached to opposite poles of the mitotic spindle and aligned at the metaphase plate. At the onset of anaphase, the activity of separins contributes to the release of cohesins from chromosomes, allowing for the segregation of sister chromatids to opposite spindle poles. Separin activity is blocked by binding to a class of proteins known as securins, whose destruction at the metaphase-to-anaphase transition is triggered by the APC/C (8–12). PTTG has been identified as a vertebrate securin on the basis of its biochemical analogy to the Pds1 protein of budding yeast and the Cut2 protein of fission yeast (7). The vertebrate securins share extensive sequence similarity with each other but show no sequence similarity to either of their yeast counterparts. Nevertheless, as was previously reported in yeast, expression of a stable Xenopus securin mutant protein blocks sister chromatid separation (7). hPTTG bound to a putative human separin homolog of yeast separins Esp1 and Cut1, and was degraded by proteolysis mediated by APC/C in a D-box-dependent manner. The finding that a vertebrate securin has tumorigenic activity is somewhat anticipated because chromosome missegregation has been predicted to be a major source of genetic instability, with profound consequences for cancer (13). On the basis of its function, the simplest explanation is that tumor formation is the result of aneuploidy caused by defects in the sister chromatid separation (7,14).
To further understand the role of hPTTG in sister chromatid separation and tumorigenesis, we searched for proteins able to bind to hPTTG using the yeast two-hybrid approach (15). This in vivo strategy was employed to provide a physiological environment in which to detect potential interactions involving hPTTG, and in this paper we demonstrate that hPTTG physically associates with Ku-70.
The Ku-70 protein associates with Ku-80 to form a heterodimeric complex (16,17), and together with the ∼470 kDa catalytic subunit, DNA-PKcs, form the DNA-dependent protein kinase (DNA-PK) (18). This enzyme is involved in repairing DNA double-strand breaks (DSBs) caused, for example, by physiological oxidation reactions, V(D)J recombination, ionizing radiation and certain chemotherapeutic drugs. Ku heterodimer binds to DNA ends and other types of discontinuity in double-stranded DNA (19). Ku itself is probably involved in stabilizing broken DNA ends, bringing them together and preparing them for ligation (20). Ku also recruits DNA-PKcs to DNA DSBs, activating its kinase function (21).
Our results demonstrate that hPTTG specifically interacts with Ku-70, which forms a complex with the Ku-70/Ku-80 heterodimer in intact cells, and that DNA-PKcs phosphorylates hPTTG in vitro. In addition, we show that hPTTG–Ku-70 association is prevented by DNA DSBs. The implications of these results, as well as a model of the regulation of hPTTG–Ku-70 interaction, are discussed.
MATERIALS AND METHODS
Plasmids and cloning
Wild-type hpttg (coding for 202 residues) and the subclones hpttg-ES (residues 1–188), hpttg-EA (residues 1–163), hpttg-EXb (residues 1–124) and hpttg-XbXh (residues 123–202), were cloned inframe with gal4-DB (DNA-binding) in the yeast pGBT9 vector (22) to yield pGBT9-hPTTG, pGBT9-hPTTG-ES, pGBT9-hPTTG-EA, pGBT9-hPTTG-EXb and pGBT9-hPTTG-XbXh, respectively. Plasmids pGBT-SNF1 and pGAD-SNF4 (22), carrying unrelated proteins, were used as controls for the two-hydrid screen.
hpttg, hpttg-EA and hpttg-HXh (residues 87–202) were cloned in frame with a 6His-tag in the bacterial pRSET-A vector (Invitrogen). The hPTTG-EXb subclone, previously fused to glutathione-S-transferase (GST) in pGEX4T2 (3), was also used in the in vitro experiments. Plasmids pRSET-S6 (6His-ribosomal protein S6; Romero,F. and Tortolero,M. unpublished results), pRSET-RIIα (23), pRSET-ΔGMAP (residues 618–803) (24) and pGEX (Pharmacia) were used as controls of the in vitro experiments.
Library screening
Saccharomyces cerevisiae strain Hf7c (MATa trp1-901 leu2-3,112 his3-200 ura3-52 lys-801 ade2-101 canr gal4-542 gal80–538 LYS2::GAL1-HIS3 URA3::Gal4 binding site-CYC1-lacZ) was grown at 30°C in YPD medium containing 1% yeast extract, 2% polypeptone and 2% glucose, and sequentially transformed with pGBT9-hPTTG-EA and a Jurkat cell oligo-dT cDNA library (25). Double transformants were plated on yeast drop-out medium lacking Trp, Leu and His (26). They were grown for 5 days at 30°C and then colonies were patched on the same medium and replica-plated on Whatman 40 filters to test for β-galactosidase activity (27). Positive clones were rescued and tested for specificity by retransformation into Hf7c either with pGBT9-hPTTG-EA or with extraneous targets (pGBT-SNF1 or pGBT9) (22). pGAD-Raf (28), pGAD-Sos1 (29), pGAD-hnRNP C (30), pGAD-hnRNP K (Romero,F., Ramos-Morales,F. and Tortolero,M., unpublished results), pGAD-Csk (31), pGAD-Lck (31), pGAD-Sumo (provided by Dr A. Germani, INSERM, Paris, France), pGAD-hPTTG (3) and pGAD-FcγRII(ID) (Romero,F., unpublished results) were also used to test the specificity of hPTTG interactions.
Sequence analysis
Sequences of cDNA inserts from positive clones of the two-hybrid screening were performed on both strands with an automatic sequencer (Pharmacia) using the Sanger dideoxy-termination method (32). Sequence comparisons were done with the BLAST 2.0 program (33).
Cell culture and lysis
HL-60 and Jurkat cells were grown in RPMI 1640 medium and Cos and HeLa cells in Dulbecco’s modified Eagle’s medium (BioWhittaker), supplemented with 10% heat-inactivated fetal calf serum (Gibco BRL), 2 mM l-glutamine, 100 U/ml penicillin and 100 µg/ml streptomycin (BioWhittaker), in a 5% CO2 humidified atmosphere at 37°C. Cell lysis was performed with 5 × 107–108 cells/ml at 4°C in 150 mM NaCl, 10 mM Tris–HCl (pH 7.5), 1% Nonidet P-40 (NP-40), 10% glycerol, 1 mM Na-vanadate, 20 mM Na-pyrophosphate, 5 mM Na-fluoride, 1% aprotinin, 1 mM phenylmethylsulfonyl fluoride (PMSF), 1 µg/ml pepstatin and 1 µg/ml leupeptin for 20 min. The extract was centrifuged at 20 000 g for 20 min and the supernatant frozen in liquid nitrogen and stored at –80°C.
Induction of differentiation in HL-60 cells
HL-60 cells were diluted to 2 × 105 cells per ml and 40 ng/ml phorbol 12-myristate 13-acetate (PMA) or the vehicle dimethyl sulfoxide (DMSO) added. Cells were grown under standard conditions in bacteriological Petri plates for 24–48 h. We took attached cells to be differentiated cells, since attachment is an indicator of macrophage phenotype (34). As control cells we used untreated or DMSO-treated cells.
Electrophoresis and western blot analysis
Proteins were separated by SDS–PAGE and gels were electroblotted onto nitrocellulose membranes and probed with the different antibodies. Peroxidase-coupled anti-rabbit IgG from donkey and anti-mouse IgG from sheep were from Amersham. Immunoreactive bands were visualized using an enhanced chemiluminescence western blotting system (Amersham) according to the manufacturer’s protocol.
Antibodies
Anti-hPTTG polyclonal antibody was produced by us (3), N3H10 (anti-Ku-70), 111 (anti-Ku-80) and 162 (anti-dimer Ku-70/Ku-80) monoclonal antibodies were provided by Dr W.H.Reeves (University of North Carolina) (35), and anti-Gal4-BD was from Clontech.
Affinity chromatography
Expression of the 6His fusion proteins was induced in Escherichia coli BL21 (DE3) by addition of 1 mM isopropyl-β-d-thiogalactoside (IPTG) and the fusion proteins isolated from bacterial lysates with Talon resin (Clontech). Cellular lysates (106–107 cells) were incubated for 2 h with fusion proteins (100–500 ng) bound to Talon resin. Resin was washed six times in lysis buffer and proteins eluted into SDS sample buffer at 95°C for 5 min and subjected to SDS–PAGE.
When indicated, fusion proteins were eluted from resin with 50 mM EDTA and dialyzed against 50 mM NaHCO3 and 150 mM NaCl pH 7.5.
To study the effect of DNA DSBs, lysates from Cos or HL-60 cells were preincubated for 30 min at 4°C with sonicated salmon sperm DNA, pUC18 or pGBT9, supercoiled or digested with EcoRI at 100–200 µg/ml, and used for the in vitro experiments. In some experiments, DNA was added to hPTTG/Ku-70 complexes previously bound to Talon resin.
GST fusion proteins were produced and purified as described (3).
Co-immunoprecipitation experiments
Cellular lysates from HL-60 cells (1.5–3 × 107) were incubated with preimmune serum for 30 min and protein A–Sepharose beads (Pharmacia Biotech Inc.) for 1 h at 4°C. After centrifugation, beads were discarded and supernatants incubated for 3 h with polyclonal anti-hPTTG or preimmune serum, followed by protein A–Sepharose beads for 1 h. Beads were washed six times with lysis buffer and bound proteins were dissolved into SDS sample buffer at 95°C for 5 min and subjected to SDS–PAGE.
In vitro kinase assays
Phosphorylation of hPTTG by DNA-PK. Purified DNA-PKcs–Ku complex (Promega) was incubated with eluted 6His-hPTTG (or derivative subclones) in the presence or absence of sonicated DNA (10 µg/ml) and with [γ-32P]ATP and DNA-PK kinase buffer (12.5 mM HEPES pH 7.5, 25 mM KCl, 6.5 mM MgCl2, 0.2 mM unlabeled ATP, 1 mM spermidine, 10% glycerol, 0.05% NP-40 and 0.5 mM DTT) for 15 min at 30°C. Reactions were terminated by adding 4× SDS sample buffer, and proteins analyzed by SDS–PAGE and autoradiography.
Phosphorylation of hPTTG by anti-dimer Ku-70/Ku-80 immunoprecipitates. HL-60 or HeLa cells were washed once with PBS and frozen. The cell pellets were later thawed and sonicated at 2 × 107 cells/ml in 50 mM NaCl-free Tris–HCl pH 7.5, 2 mM EDTA, 0.3% NP-40, 1 mM Na-vanadate, 20 mM Na-pyrophosphate, 5 mM Na-fluoride, 1% aprotinin, 1 mM PMSF, 1 µg/ml pepstatin and 1 µg/ml leupeptin, and the extracts were cleared twice by centrifugation at 13000 g for 15 min followed by immunoprecipitation with mAb 162. After incubation with protein A–Sepharose beads, the beads were washed three times with NET-NP-40 buffer (150 mM NaCl, 2 mM EDTA, 50 mM Tris–HCl pH 7.5 and 0.3% NP-40) and once with kinase buffer and stored at –80°C. In vitro kinase assays were performed as indicated above.
RESULTS
Isolation of proteins interacting with hPTTG
We recently isolated hpttg cDNA, which codes for a protein highly expressed in proliferating cells and is involved in sister chromatid separation and tumorigenesis (1,3,7). To elucidate the molecular mechanisms that involve hPTTG in these cellular functions, we searched for proteins that interact with hPTTG using the yeast two-hybrid system with Gal4 recognition sites regulating expression of both His3 and LacZ (22). Because full-length hPTTG fused to the DNA-binding domain (BD) of Gal4 in pGBT9 had shown a strong transactivation activity in yeast (3), we subcloned several fragments of hpttg in pGBT9 to find the longest without this transcriptional activity. As shown in Figure 1A, strain Hf7c transformed with pGBT9-hPTTG-EA was not able to grow in selective medium, although this fusion protein was well produced in yeast as detected by western blot with anti-Gal4-BD antibody (data not shown). This strain was retransformed with a Jurkat cell oligo-dT cDNA library constructed in pGAD1318 (25) to find clones able to grow in the absence of histidine. We screened about one million clones and we found a positive clone that specifically interacts with hPTTG-EA. The nucleotide sequence showed that this clone codes for Ku-70 (residues 291–609). This protein forms a heterodimeric complex with Ku-80, which is the DNA-binding component of DNA-PK. This enzyme is involved in DNA repair and V(D)J recombination (36). Furthermore, the specificity of hPTTG/Ku-70 interaction was illustrated by the lack of reporter gene activation when hPTTG-EA association was assayed with cytosolic proteins (Raf or Sos1), protein kinases (Lck or Csk), nuclear proteins (hnRNP C or hnRNP K), the intracellular domain of a receptor (FcγRII), a ubiquitin-related protein (Sumo) or itself (Fig. 1B). Therefore, by using the two-hybrid system, we show that hPTTG interacts with Ku-70.
Figure 1.
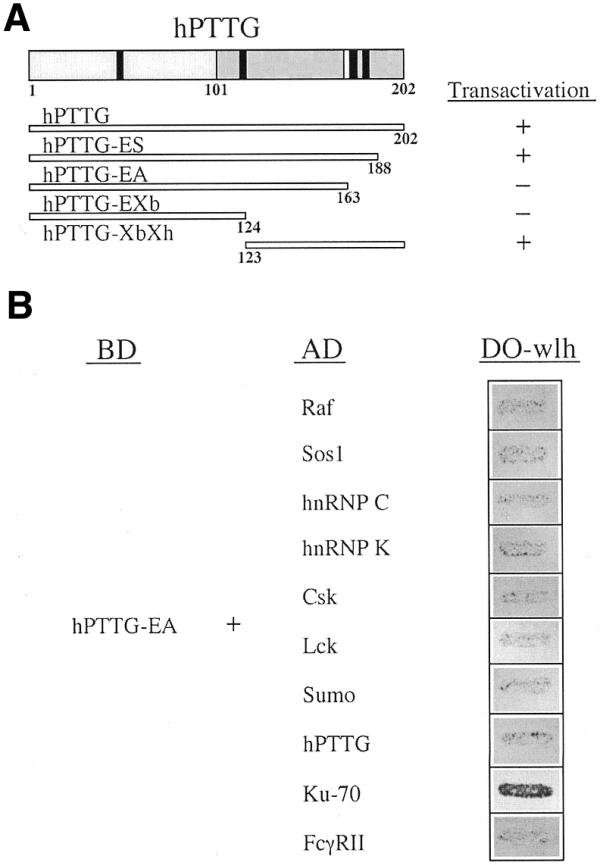
Interaction of Ku-70 with hPTTG using the two-hybrid system. (A) Schematic representation of hPTTG and study of the transcriptional activity of several hPTTG subclones. hPTTG protein can be divided into a basic N-terminal portion (residues 1–101) and an acidic C-terminal portion (residues 102–202). The four Pro-X-X-Pro motifs (black bars), putative binding sites for SH3-containing proteins and the CDC2 phosphorylation site (white bar) are also indicated. Several fragments of hpttg were subcloned in pGBT9 and their transactivation activity tested in Hf7c reporter strain. +, Growth in the absence of histidine and with or without 5 mM of 3-amino-1,2,4-triazole. –, Absence of growth in the same medium. (B) hPTTG associates specifically with Ku-70. Hf7c reporter strain was cotransformed with the indicated plasmids. The interaction between the two-hybrid proteins is indicated by growth in the absence of histidine (DO-wlh) (dark grey patch). BD, fusion with the DNA BD of Gal4. AD, fusion of the activation domain of Gal4.
hPTTG interacts with Ku-70 from cellular extracts
To validate the results obtained with the yeast two-hybrid system, the interaction of hPTTG with Ku-70 was studied by in vitro binding experiments. Full-length hpttg and hpttg-HXh were cloned in frame with a 6His-tag in pRSET-A and purified from bacterial lysates by affinity chromatography on Talon resin. Lysates from HL-60 cells were incubated with the resin bound to 6His fusion proteins, the resin was washed and the eluted proteins resolved on gels and immunoblotted with an anti-Ku-70 monoclonal antibody. Figure 2 clearly shows that the Ku-70 protein from cellular extracts interacts with 6His-hPTTG, whereas there is no interaction with 6His-hPTTG-HXh or with irrelevant fusion proteins (6His-S6, 6His-RIIα or GST). Similar results were obtained with lysates from Jurkat, Cos and HeLa cells (data not shown). These results indicate that hPTTG and Ku-70 also interact in vitro. Furthermore, the finding that 6His-hPTTG-HXh, which lacks the 86 N-terminal residues of hPTTG, no longer interacted with Ku-70 suggests that the N-terminal portion of hPTTG is responsible for the association with Ku-70. In fact, the GST-hPTTG-EXb fusion protein, containing the N-terminal moiety of hPTTG, was able to interact with Ku-70 (Fig. 2).
Figure 2.

hPTTG interacts with Ku-70 in vitro. Expression of the 6His and GST fusion proteins was induced by addition of IPTG. Proteins were purified from bacterial lysates on Talon resin or glutathione–agarose beads. Fusion proteins were incubated with NP-40 extracts from HL-60 cells (5 × 106) and their associations with Ku-70 were determined by immunoblotting with anti-Ku-70 (N3H10) antibody. HL-60, extract from 5 × 105 HL-60 cells. Mw, molecular weights expressed in kDa. These interactions were detected with different amounts of fusion proteins (between 100 and 500 ng).
hPTTG and Ku-70 form a complex in intact cells
hpttg is expressed in tissues that contain highly proliferating cells, and, in fact, we can establish a correlation between hpttg expression and cell proliferation (5). For this reason, we used the HL-60 cell line as an appropriate experimental model to study the in vivo protein complexes involving hPTTG. These cells respond to specific chemical stimuli by acquiring either a granulocyte- (37), monocyte- or macrophage-like phenotype (34). The acquisition of mature phenotypes can be demonstrated by a variety of differentiation markers and leads to growth arrest. As shown in Figure 3A, hPTTG protein is absent in HL-60 cells treated with PMA, which induces a macrophage-like differentiation. Therefore, we have a good model to examine the relevance of the hPTTG interactions.
Figure 3.
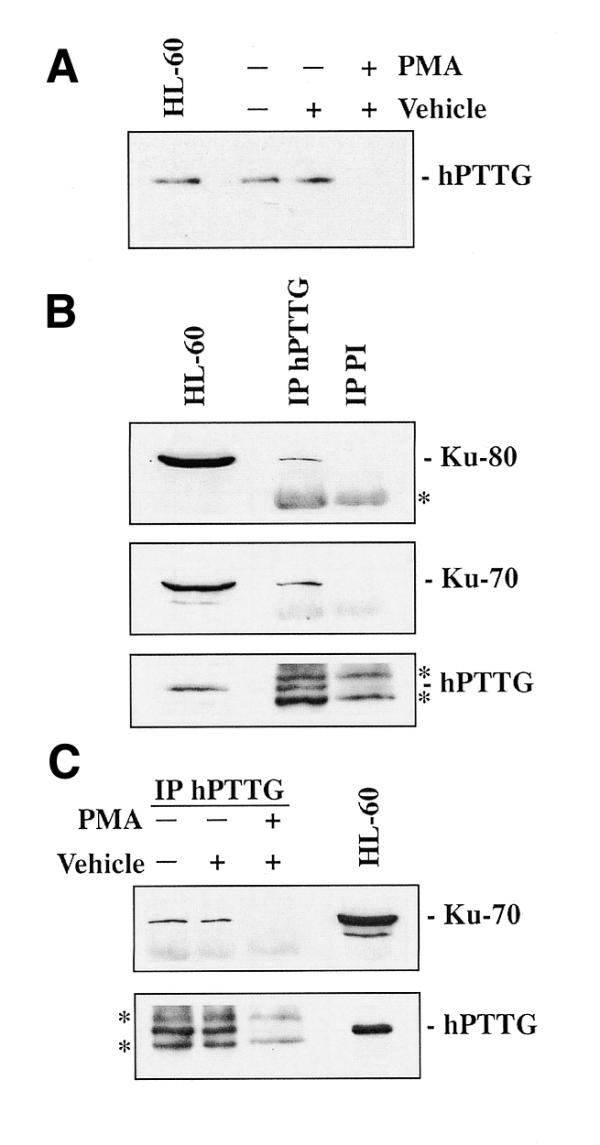
Binding of hPTTG to Ku-70 in intact cells. (A) Expression of hPTTG in differentiated and undifferentiated HL-60 cells. Equal amounts of total extracts from HL-60 cells treated with PMA (40 ng/ml), with the vehicle DMSO or untreated, were separated by SDS–PAGE, transferred to nitrocellulose filters and incubated with anti-hPTTG. HL-60, extract from 106 HL-60 cells. (B) Co-immunoprecipitation of hPTTG with Ku-70 and Ku-80 in undifferentiated HL-60 cells. Anti-hPTTG and preimmune (PI) sera were used to immunoprecipitate from NP-40 extracts of HL-60 cells (3 × 107 cells each). Complexes were resolved by SDS–PAGE, transferred to nitrocellulose filters and incubated with anti-Ku-70, anti-Ku-80 and anti-hPTTG. HL-60, extract from 106 HL-60 cells. (C) Study of the relevance of the hPTTG–Ku-70 interaction. Anti-hPTTG was used to immunoprecipitate from extracts of HL-60 cells (3 × 107) treated with PMA (40 ng/ml), the vehicle DMSO or untreated. Nitrocellulose filters were incubated with anti-Ku-70 and anti-hPTTG. HL-60, extract from 106 HL-60 cells. Asterisks indicate the IgG heavy and light chains.
To know whether hPTTG and Ku-70 formed complexes in intact cells, we performed a series of co-immunoprecipitation experiments. For this purpose, NP-40 lysates from HL-60 cells were incubated with anti-hPTTG polyclonal antibody or with the preimmune serum. The resulting immunocomplexes were blotted with anti-Ku-70 and, as illustrated in Figure 3B, hPTTG co-immunoprecipitated with the endogenous DNA-binding subunit of DNA-PK. Furthermore, Ku-80 was also detected after reblotting the dehybridized nitrocellulose filter with anti-Ku-80 specific monoclonal antibody (mAb 111) (Fig. 3B). Similarly, in vitro experiments with 6His-hPTTG showed that the binding of Ku-80 was detected in addition to the association of hPTTG with Ku-70. However, when anti-hPTTG antibody was previously incubated with 6His-hPTTG bound to resin, and the supernatant used to immunoprecipitate proteins from HL-60 extracts, Ku-70 and Ku-80 were no longer detected (data not shown). These results suggest that binding of hPTTG to Ku-70 does not modify the formation of the Ku-70/Ku-80 heterodimer.
Co-immunoprecipitations using lysates from PMA differentiated HL-60 cells were also performed as controls, and no Ku-70 was detected (Fig. 3C). As indicated above, hPTTG protein was absent in these lysates, while Ku-70 was detected in both differentiated and undifferentiated HL-60 cells. Thus, our findings show that full-length hPTTG and Ku dimer can form a complex in intact cells.
hPTTG is a substrate for DNA-PKcs in vitro
To determine whether the catalytic subunit of the DNA-PK complex phosphorylates hPTTG, we incubated soluble 6His-hPTTG with purified DNA-PKcs and Ku in the presence of sonicated DNA. The results show strong phosphorylation of 6His-hPTTG (Fig. 4A, lane 1). The identification of at least six potential motifs for DNA-PK phosphorylation (Ser/Thr-Gln or Pro-Ser/Thr) in hPTTG might justify this phosphorylation level (38,39). In fact, experiments performed with 6His-hPTTG-EA or 6His-hPTTG-HXh subclones, that only contain three potential phosphorylation sites, revealed lower phosphorylation signals (Fig. 4A, lanes 3 and 5). No phosphorylation was observed in the absence of sonicated DNA or using the control protein 6His-ΔGMAP as substrate for DNA-PK (Fig. 4A, lanes 2, 4 and 6–8).
Figure 4.
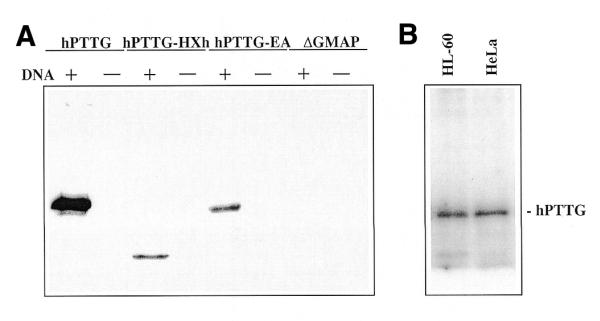
DNA-PKcs phosphorylates hPTTG in vitro. (A) Phosphorylation of hPTTG by DNA-PK. Purified DNA-PKcs–Ku complex was incubated with soluble 6His-fusion proteins in the presence (+) or absence (–) of sonicated DNA (10 µg/ml) and with [γ-32P]ATP and DNA-PK kinase buffer. Reactions were analyzed by SDS–PAGE and autoradiography. 6His-ΔGMAP was used as control. (B) Phosphorylation of hPTTG by anti-dimer Ku-70/Ku-80 immunoprecipitates. Extracts from HL-60 (107) and HeLa (5 × 106) cells were used to immunoprecipitate the DNA-PKcs–Ku complexes with mAb 162 (anti-dimer Ku-70/Ku-80). After incubation with protein A–Sepharose, the beads were washed and used to perform in vitro kinase assays with soluble 6His-hPTTG and sonicated DNA as described.
The phosphorylation of hPTTG by DNA-PK was further studied by incubating the anti-Ku-70/Ku-80 heterodimer immunoprecipitates with 6His-hPTTG. As shown in Figure 4B, the DNA-PKcs immunoprecipitated with anti-Ku-70/Ku-80 also phosphorylates hPTTG in the presence of sonicated DNA.
We next sought to know whether the binding hPTTG/Ku-70 was affected by DNA-PK phosphorylation. To this end, eluted 6His-hPTTG was phosphorylated in vitro by the DNA-PKcs co-immunoprecipitated with anti-Ku-70/Ku-80 heterodimer bound to protein A–Sepharose beads. The supernatant fraction was obtained by sedimentation of the beads. Following washing of the beads with NET-NP-40 buffer, the beads and the supernatant fraction were boiled with SDS sample buffer. The adsorbates and the supernatants were analyzed by immunoblotting with anti-hPTTG and both hPTTG and phosphorylated hPTTG were bound to Ku-70 complexes (data not shown). Therefore, in these experimental conditions, the hPTTG phosphorylation has no detectable effect on hPTTG–Ku-70 association.
DNA DSBs prevent hPTTG–Ku-70 association
Since DNA-PK is involved in DNA DSB repair and V(D)J recombination, we investigated whether the presence of DNA DSBs affects the interaction between hPTTG and Ku-70. NP-40 lysates were preincubated with sonicated salmon sperm DNA or with a linear plasmid (pUC18 digested with EcoRI) at the same concentration prior to incubation with 6His-hPTTG, and western-blot analyses with anti-Ku-70 showed that sonicated DNA largely decreased the hPTTG–Ku-70 interaction and linear plasmid completely abolished this association (Fig. 5A). Similar results were obtained when the yeast linear plasmid pGBT9 was used (data not shown). The observed differences between the sonicated DNA and the linear plasmids may be the result of the number of breaks on the total DNA. The sonicated salmon sperm DNA has a mean size of 7 kb, as visualized in agarose gels, while pUC18 has a size of 2.8 kb. In fact, when we used twice the amount of sonicated DNA, we abolished the hPTTG–Ku-70 interaction (Fig. 5B). In contrast, incubation of NP-40 lysates with supercoiled DNA (pUC18 or pGBT9) did not affect the binding. Furthermore, incubation of sonicated salmon sperm DNA with hPTTG–Ku-70 complexes bound to Talon resin largely disrupted the complexes once formed (Fig. 5C). These data suggest that the presence of DNA breaks prevents hPTTG–Ku-70 interaction and disrupts hPTTG–Ku-70 complexes in vivo.
Figure 5.
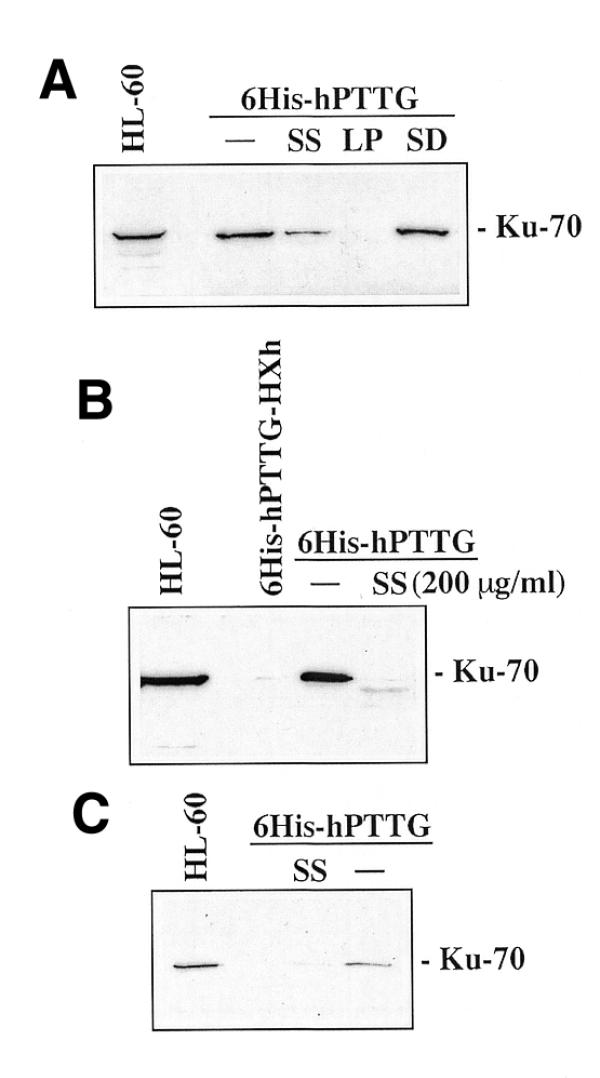
DNA DSBs prevent hPTTG–Ku-70 association. (A) NP-40 lysates from HL-60 cells (5 × 106) were preincubated with sonicated salmon sperm DNA (SS), pUC18 digested with EcoRI (LP) or supercoiled pUC18 DNA (SD) at 100 µg/ml or with the buffer alone (–) prior to incubate with 6His-hPTTG. Western blots were developed with anti-Ku-70. HL-60, extract from 5 × 105 HL-60 cells. (B) Similar experiments were performed but using 200 µg/ml of sonicated salmon sperm DNA (SS). 6His-hPTTG-HXh was used as control. HL-60, extract from 5 × 105 HL-60 cells. (C) NP-40 lysates from HL-60 cells (5 × 106) were incubated with 6His-hPTTG. Resin was washed in lysis buffer and then incubated with sonicated salmon sperm DNA (SS) at 100 µg/ml or with the buffer alone (–). Western blots were developed with anti-Ku-70. HL-60, extract from 5 × 105 HL-60 cells.
DISCUSSION
Mitosis is a complex process that includes nuclear envelope breakdown, chromatin condensation and chromosome segregation. Thus, multiple events must be coordinated in order to ensure the progression of mitosis (40). The onset of mitosis is controlled by the activity of the mitosis-promoting factor (MPF), which induces the formation of a mitotic spindle. After anaphase, the destruction of the mitotic cyclins leads to MPF inactivation, originating spindle disassembly and cytokinesis (41,42). The timing of the metaphase-to-anaphase transition and the exit from mitosis is regulated by the activation of the APC/C, which mediates selective proteolysis of various mitotic regulators. Pds1 from S.cerevisiae and Cut2 from Schizosaccharomyces pombe (the securins) are APC/C sustrates and their degradation is required for chromatid separation (43,44). Pds1 and Cut2 associate with Esp1 and Cut1 (the separins), respectively, and prevent separins from promoting chromatid separation (45). hPTTG has been identified as a human securin through its association with the human separin (7). Recently, it has been proposed that in vertebrates, a cleavage-independent pathway removes cohesin from chromosome arms during prophase, whereas a separin-dependent pathway cleaves centromeric cohesin at the metaphase-to-anaphase transition. In a HeLa cell line stably expressing a tagged version of Scc1 cohesin, it has been shown that Scc1 is cleaved and removed from centromeric regions at the metaphase-to-anaphase transition. Scc1 cleavage depends on the APC/C because securin proteolysis is needed for separin activation (46).
Through a two-hybrid screen in yeast using hPTTG as bait, we isolated a protein of 70 kDa that specifically interacts with hPTTG. This protein is Ku-70, the DNA-binding component of the DNA-dependent protein kinase. DNA-PK is a DNA-activated nuclear serine/threonine protein kinase that consists of a regulatory subunit, the heterodimeric Ku protein (composed of a 70 and 80 kDa subunit), which binds DNA ends and targets the catalytic subunit, DNA-PKcs, to DNA strand breaks. All three gene products are clearly involved in repairing ionizing radiation-induced damage and in V(D)J recombination (47). The Ku-dependent repair process, called illegitimate recombination or non-homologous end joining (NHEJ), appears to be the main DNA DSB repair mechanism in mammalian cells (48). hPTTG interacts both in vitro and in vivo with Ku heterodimer and is a substrate for DNA-PKcs. In budding yeast, DNA damage induces the phosphorylation of the PTTG homolog Pds1, and this phosphorylation is specific for types of DNA damage that elicit a mitotic checkpoint arrest but not of reagents that cause a DNA replication checkpoint arrest (49). The equivalent checkpoint in fission yeast and vertebrates arrests the cell cycle at the G2–M boundary and, although the arrest point differs, the signal transduction pathway involves damage-dependent phosphorylation that is mediated by a conserved element (i.e. Mec1 in budding yeast, Rad3 in fission yeast and probably ATM in mammalian cells) (50). We suggest that in human cells DNA damage inducing G2 arrest not only leads to inhibitory phosphorylation of the kinase CDC2 via an ATM-dependent pathway, but also induces DNA-PK phosphorylation of PTTG.
Our findings that hPTTG forms a complex with Ku-70 and that this interaction is regulated by DNA DSBs allow us to propose a mechanism of regulation of hPTTG–Ku interaction. Ku dimer interacts with the N-terminal portion of hPTTG (Fig. 2). The C-terminal portion of hPTTG would associate with the N-terminal part of human separin, if the human system is similar to the fission yeast system (51). Moreover, we think this is also true in metazoans, because although Pds1, Cut2 and PTTG show no sequence similarity to each other, the C-terminus of the putative human separin homolog (52) has 28% identity with Esp1 and 30% identity with Cut1 (53), and the N-terminus has no similarity. One might presume that this non-homologous region of each separin interacts with the respective securin and the similar C-terminal portions of separins would bind to homolog partners or at least to different partners with similar protein-binding motif. In response to DNA damage, Ku enhances both the strength and stability of the association with DNA and DNA-PKcs becomes activated (54), then hPTTG dissociates from Ku-70 (Fig. 5) and is phosphorylated by DNA-PK (Fig. 4). The phosphorylated form of hPTTG would contribute to block sister chromatid separation.
A recent publication states that telomeric DNA mutants of Tetrahymena thermophila block sister chromatid separation in mitosis (55). On the other hand, it is also known that Ku plays a role in maintaining chromosomal stability protecting the telomeres (56): yeast strains defective in Ku-70 or Ku-80 lose the majority, but not all, of their terminal telomere repeats (57). Thus, through their contribution to efficient telomeric end-capping, NHEJ repair proteins help to preserve the fidelity of genetic inheritance. Therefore, we might suggest that this protective role of Ku could be mediated at least in part by the inhibition of sister chromatid separation. Further experiments will be necessary to investigate this hypothesis.
Stable transfection of NIH 3T3 cells with pttg cDNA causes anchorage-independent transformation in vitro and induces in vivo tumor formation when transfectants are injected into athymic mice (1). This suggests that misregulation of chromatid separation may contribute to generation of malignant tumors (7). In this way, inactivation of the ku-70 gene also leads to a propensity for malignant transformation (58). ku-70–/– mice develop T cell lymphomas and karyotyping analyses on cultured cells derived from primary T cell lymphomas reveal multiple chromosomal abnormalities (monosomies, trisomies, duplications, etc.). This might be explained in vivo, at least in part, in terms of an increased rate of sister chromatid exchange, but we cannot eliminate the possibility that defects in sister chromatid separation also play a role. Therefore, both defective regulation of hPTTG and inactivation of the ku-70 gene facilitate neoplastic growth.
To summarize, our studies show that the human securin/hPTTG is associated with the DNA-dependent protein kinase both in vitro and in vivo and that it is phosphorylated by DNA-PKcs. Our data also demonstrate that DNA DSBs prevent hPTTG–Ku-70 association. Together, these observations suggest that hPTTG might connect the DNA damage–response pathway with sister chromatid separation.
Acknowledgments
ACKNOWLEDGEMENTS
This work was supported by grants from Ministerio de Educación y Cultura (MEC) of Spain (SAF99-0125-C03-01), from DGUI of the Junta de Andalucía and from Rhône-Poulenc Rorer (Paris, France). F.R. was supported by postdoctoral fellowships from the EU and Spanish MEC.
References
- 1.Pei L. and Melmed,S. (1997) Isolation and characterization of a pituitary tumor-transforming gene (PTTG). Mol. Endocrinol., 11, 433–441. [DOI] [PubMed] [Google Scholar]
- 2.Zhang X., Horwitz,G.A., Prezant,T.R., Valentini,A., Nakashima,M., Bronstein,M.D. and Melmed,S. (1999) Structure, expression and function of human pituitary tumor-transforming gene (PTTG). Mol. Endocrinol., 13, 156–166. [DOI] [PubMed] [Google Scholar]
- 3.Dominguez A., Ramos-Morales,F., Romero,F., Rios,R.M., Dreyfus,F., Tortolero,M. and Pintor-Toro,J.A. (1998) hpttg, a human homologue of rat pttg, is overexpressed in hematopoietic neoplasms. Evidence for a transcriptional activation function of hPTTG. Oncogene, 17, 2187–2193. [DOI] [PubMed] [Google Scholar]
- 4.Saez C., Japon,M.A., Ramos-Morales,F., Romero,F., Segura,D.I., Tortolero,M. and Pintor-Toro,J.A. (1999) hpttg is over-expressed in pituitary adenomas and other primary epithelial neoplasias. Oncogene, 18, 5473–5476. [DOI] [PubMed] [Google Scholar]
- 5.Ramos-Morales F., Dominguez,A., Romero,F., Luna,R., Multon,M.C., Pintor-Toro,J.A. and Tortolero,M. (2000) Cell cycle regulated expression and phosphorylation of hpttg proto-oncogene product. Oncogene, 19, 403–409. [DOI] [PubMed] [Google Scholar]
- 6.King R.W., Glotzer,M. and Kirschner,M.W. (1996) Mutagenic analysis of the destruction signal of mitotic cyclins and structural characterization of ubiquitinated intermediates. Mol. Biol. Cell, 7, 1343–1357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zou H., McGarry,T.J., Bernal,T. and Kirschner,M.W. (1999) Identification of a vertebrate sister-chromatid separation inhibitor involved in transformation and tumorigenesis. Science, 285, 418–422. [DOI] [PubMed] [Google Scholar]
- 8.Biggins S. and Murray,A.W. (1999) Sister chromatid cohesion in mitosis. Curr. Opin. Genet. Dev., 9, 230–236. [DOI] [PubMed] [Google Scholar]
- 9.Morgan D.O. (1999) Regulation of the APC and the exit from mitosis. Nature Cell. Biol., 1, E47–E53. [DOI] [PubMed] [Google Scholar]
- 10.Nasmyth K. (1999) Separating sister chromatids. Trends Biochem. Sci., 24, 98–104. [DOI] [PubMed] [Google Scholar]
- 11.Zachariae W. (1999) Progression into and out of mitosis. Curr. Opin. Cell. Biol., 11, 708–716. [DOI] [PubMed] [Google Scholar]
- 12.Zachariae W. and Nasmyth,K. (1999) Whose end is destruction: cell division and the anaphase-promoting complex. Genes Dev., 13, 2039–2058. [DOI] [PubMed] [Google Scholar]
- 13.Lengauer C., Kinzler,K.W. and Vogelstein,B. (1998) Genetic instabilities in human cancers. Nature, 396, 643–649. [DOI] [PubMed] [Google Scholar]
- 14.Orr-Weaver T.L. (1999) The difficulty in separating sisters. Science, 285, 344–345. [DOI] [PubMed] [Google Scholar]
- 15.Fields S. and Song,O. (1989) A novel genetic system to detect protein–protein interactions. Nature, 340, 245–246. [DOI] [PubMed] [Google Scholar]
- 16.Koike M., Awaji,T., Kataoka,M., Tsujimoto,G., Kartasova,T., Koike,A. and Shiomi,T. (1999) Differential subcellular localization of DNA-dependent protein kinase components Ku and DNA-PKcs during mitosis. J. Cell Sci., 112, 4031–4039. [DOI] [PubMed] [Google Scholar]
- 17.Wu X. and Lieber,M.R. (1996) Protein–protein and protein–DNA interaction regions within the DNA end-binding protein Ku70-Ku86. Mol. Cell. Biol., 16, 5186–5193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jin S. and Weaver,D.T. (1997) Double-strand break repair by Ku70 requires heterodimerization with Ku80 and DNA binding functions. EMBO J., 16, 6874–6885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Blier P.R., Griffith,A.J., Craft,J. and Hardin,J.A. (1993) Binding of Ku protein to DNA. Measurement of affinity for ends and demonstration of binding to nicks. J. Biol. Chem., 268, 7594–7601. [PubMed] [Google Scholar]
- 20.Ramsden D.A. and Gellert,M. (1998) Ku protein stimulates DNA end joining by mammalian DNA ligases: a direct role for Ku in repair of DNA double-strand breaks. EMBO J., 17, 609–614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gottlieb T.M. and Jackson,S.P. (1993) The DNA-dependent protein kinase: requirement for DNA ends and association with Ku antigen. Cell, 72, 131–142. [DOI] [PubMed] [Google Scholar]
- 22.Chien C.T., Bartel,P.L., Sternglanz,R. and Field,S. (1991) The two-hybrid system: a method to identify and clone genes for proteins that interact with a protein of interest. Proc. Natl Acad. Sci. USA, 88, 9578–9582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Martin M.E., Hidalgo,J., Vega,F.M. and Velasco,A. (1999) Trimeric G proteins modulate the dynamic interaction of PKAII with the Golgi complex. J. Cell Sci., 112, 3869–3878. [DOI] [PubMed] [Google Scholar]
- 24.Infante C., Ramos-Morales,F., Fedriani,C., Bornens,M. and Rios,R.M. (1999) GMAP-210, A cis-Golgi network-associated protein, is a minus end microtubule-binding protein. J. Cell Biol., 145, 83–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Romero F., Dargemont,C., Pozo,F., Reeves,W.H., Camonis,J., Gisselbrecht,S. and Fischer,S. (1996) p95vav associates with the nuclear protein Ku-70. Mol. Cell. Biol., 16, 37–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sherman F., Fink,G.R. and Hicks,J.B. (1986) Methods in Yeast Genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
- 27.Breeden L. and Nasmyth,K. (1985) Regulation of the yeast HO gene. Quant. Biol., 50, 643–650. [DOI] [PubMed] [Google Scholar]
- 28.van Aelst L., Barr,M., Marcus,S., Polverino,A. and Wigler,M. (1993) Complex formation between RAS and RAF and other protein kinases. Proc. Natl Acad. Sci. USA, 90, 6213–6217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chardin P., Camonis,J.H., Gale,N.W., van Aelst,L., Schlessinger,J., Wigler,M.H. and Bar-Sagi,D. (1993) Human Sos1: a guanine nucleotide exchange factor for Ras that binds to Grb2. Science, 260, 1338–1343. [DOI] [PubMed] [Google Scholar]
- 30.Romero F., Germani,A., Puvion,E., Camonis,J., Varin-Blank,N., Gisselbrecht,S. and Fischer,S. (1998) Vav binding to heterogeneous nuclear ribonucleoprotein (hnRNP) C. Evidence for Vav-hnRNP interactions in an RNA-dependent manner. J. Biol. Chem., 273, 5923–5931. [DOI] [PubMed] [Google Scholar]
- 31.Bougeret C., Delaunay,T., Romero,F., Jullien,P., Sabe,H., Hanafusa,H., Benarous,R. and Fischer,S. (1996) Detection of a physical and functional interaction between Csk and Lck which involves the SH2 domain of Csk and is mediated by autophosphorylation of Lck on tyrosine 394. J. Biol. Chem., 271, 7465–7472. [DOI] [PubMed] [Google Scholar]
- 32.Sanger F., Nicklen,S. and Coulson,A.R. (1977) DNA sequencing with chain-terminating inhibitors. Proc. Natl Acad. Sci. USA, 74, 5463–5467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gish W. and States,D.J. (1993) Identification of protein coding regions by database similarity search. Nature Genet., 3, 266–272. [DOI] [PubMed] [Google Scholar]
- 34.Murao S., Gemmell,M.A., Callaham,M.F., Anderson,N.L. and Huberman,E. (1983) Control of macrophage cell differentiation in human promyelocytic HL-60 leukemia cells by 1,25-dihydroxyvitamin D3 and phorbol-12-myristate-13-acetate. Cancer Res., 43, 4989–4996. [PubMed] [Google Scholar]
- 35.Reeves W.H., Satoh,M., Wang,J. and Ajmani,A.K. (1994) In van Venrooij,W.J. and Maini,R.N. (eds), Manual of Biological Markers of Disease. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp. 1–22.
- 36.Smith G.C. and Jackson,S.P. (1999) The DNA-dependent protein kinase. Genes Dev., 13, 916–934. [DOI] [PubMed] [Google Scholar]
- 37.Collins S.J., Bodner,A., Ting,R. and Gallo,R.C. (1980) Induction of morphological and functional differentiation of human promyelocytic leukemia cells (HL-60) by componuds which induce differentiation of murine leukemia cells. Int. J. Cancer, 25, 213–218. [DOI] [PubMed] [Google Scholar]
- 38.Zernik-Kobak M., Vasunia,K., Connelly,M., Anderson,C.W. and Dixon,K. (1997) Sites of UV-induced phosphorylation of the p34 subunit of replication protein A from HeLa cells. J. Biol. Chem., 272, 23896–23904. [DOI] [PubMed] [Google Scholar]
- 39.Watanabe F., Teraoka,H., Iijima,S., Mimori,T. and Tsukada,K. (1994) Molecular properties, substrate specificity and regulation of DNA-dependent protein kinase from Raji Burkitt’s lymphoma cells. Biochim. Biophys. Acta, 1223, 255–260. [DOI] [PubMed] [Google Scholar]
- 40.Pines J. (1999) Four-dimensional control of the cell cycle. Nature Cell Biol., 1, E73–E79. [DOI] [PubMed] [Google Scholar]
- 41.Holloway S.L., Glotzer,M., King,R.W. and Murray,A.W. (1993) Anaphase is initiated by proteolysis rather than by the inactivation of maturation-promoting factor. Cell, 73, 1393–1402. [DOI] [PubMed] [Google Scholar]
- 42.Nigg E.A., Blangy,A. and Lane,H.A. (1996) Dynamic changes in nuclear architecture during mitosis: on the role of protein phosphorylation in spindle assembly and chromosome segregation. Exp. Cell Res., 229, 174–180. [DOI] [PubMed] [Google Scholar]
- 43.Cohen-Fix O., Peters,J.M., Kirschner,M.W. and Koshland,D. (1996) Anaphase initiation in Saccharomyces cerevisiae is controlled by the APC-dependent degradation of the anaphase inhibitor Pds1p. Genes Dev., 10, 3081–3093. [DOI] [PubMed] [Google Scholar]
- 44.Funabiki H., Yamano,H., Kumada,K., Nagao,K., Hunt,T. and Yanagida,M. (1996) Cut2 proteolysis required for sister-chromatid seperation in fission yeast. Nature, 381, 438–441. [DOI] [PubMed] [Google Scholar]
- 45.Toth A., Ciosk,R., Uhlmann,F., Galova,M., Schleiffer,A. and Nasmyth,K. (1999) Yeast cohesin complex requires a conserved protein, Eco1p(Ctf7), to establish cohesion between sister chromatids during DNA replication. Genes Dev., 13, 320–333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Waizenegger I.C., Hauf,S., Meinke,A. and Peters,J.M. (2000) Two distinct pathways remove mammalian cohesin from chromosome arms in prophase and from centromeres in anaphase. Cell, 103, 399–410. [DOI] [PubMed] [Google Scholar]
- 47.Featherstone C. and Jackson,S.P. (1999) DNA double-strand break repair. Curr. Biol., 9, R759–R761. [DOI] [PubMed] [Google Scholar]
- 48.Lieber M.R. (1999) The biochemistry and biological significance of nonhomologous DNA end joining: an essential repair process in multicellular eukaryotes. Genes Cells, 4, 77–85. [DOI] [PubMed] [Google Scholar]
- 49.Cohen-Fix O. and Koshland,D. (1997) The anaphase inhibitor of Saccharomyces cerevisiae Pds1p is a target of the DNA damage checkpoint pathway. Proc. Natl Acad. Sci. USA, 94, 14361–14366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bentley N.J. and Carr,A.M. (1997) DNA structure-dependent checkpoints in model systems. Biol. Chem., 378, 1267–1274. [PubMed] [Google Scholar]
- 51.Kumada K., Nakamura,T., Nagao,K., Funabiki,H., Nakagawa,T. and Yanagida,M. (1998) Cut1 is loaded onto the spindle by binding to Cut2 and promotes anaphase spindle movement upon Cut2 proteolysis. Curr. Biol., 8, 633–641. [DOI] [PubMed] [Google Scholar]
- 52.Nagase T., Seki,N., Ishikawa,K., Tanaka,A. and Nomura,N. (1996) Prediction of the coding sequences of unidentified human genes. V. The coding sequences of 40 new genes (KIAA0161-KIAA0200) deduced by analysis of cDNA clones from human cell line KG-1. DNA Res., 3, 17–24. [DOI] [PubMed] [Google Scholar]
- 53.Funabiki H., Kumada,K. and Yanagida,M. (1996) Fission yeast Cut1 and Cut2 are essential for sister chromatid separation, concentrate along the metaphase spindle and form large complexes. EMBO J., 15, 6617–6628. [PMC free article] [PubMed] [Google Scholar]
- 54.West R.B., Yaneva,M. and Lieber,M.R. (1998) Productive and nonproductive complexes of Ku and DNA-dependent protein kinase at DNA termini. Mol. Cell. Biol., 18, 5908–5920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kirk K.E., Harmon,B.P., Reichardt,I.K., Sedat,J.W. and Blackburn,E.H. (1997) Block in anaphase chromosome separation caused by a telomerase template mutation. Science, 275, 1478–1481. [DOI] [PubMed] [Google Scholar]
- 56.Bailey S.M., Meyne,J., Chen,D.J., Kurimasa,A., Li,G.C., Lehnert,B.E. and Goodwin,E.H. (1999) DNA double-strand break repair proteins are required to cap the ends of mammalian chromosomes. Proc. Natl Acad. Sci. USA, 96, 14899–14904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Boulton S.J. and Jackson,S.P. (1998) Components of the Ku-dependent non-homologous end-joining pathway are involved in telomeric length maintenance and telomeric silencing. EMBO J., 17, 1819–1828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Li G.C., Ouyang,H., Li,X., Nagasawa,H., Little,J.B., Chen,D.J., Ling,C.C., Fuks,Z. and Cordon-Cardo,C. (1998) Ku70: a candidate tumor suppressor gene for murine T cell lymphoma. Mol. Cell, 2, 1–8. [DOI] [PubMed] [Google Scholar]