

Abstract
Two experiments tested the assumption of Urcuioli’s (2008) theory of pigeons’ equivalence-class formation that consistent non-reinforcement of certain stimulus combinations in successive matching juxtaposed with consistent reinforcement of other combinations generates stimulus classes containing the elements of the reinforced combinations. In Experiment 1, pigeons were concurrently trained on symbolic (AB) and two identity (AA and BB) successive tasks in which half of all identity trials ended in non-reinforcement but all AB trials were reinforced, contingent upon either responding or not-responding to the comparisons. Subsequent symmetry (BA) probe trials showed evidence of symmetry in one of four pigeons. In Experiment 2, pigeons learned three pair-comparison tasks in which left versus right spatial choices were reinforced after the various sample-comparison combinations comprising AB, AA, and BB conditional discriminations. Nondifferentially-reinforced BA probe trials following acquisition showed some indication of symmetrical choice responding. The overall results contradict the theoretical predictions derived from Urcuioli (2008) and those from Experiment 2 challenge other stimulus class analyses as well.
Keywords: associative symmetry, stimulus classes, successive matching, non-reinforcement, pair-comparison task, pigeons
Associative symmetry is the finding that after training symbolic (AB) matching in which subjects learn to respond to B comparisons after A samples, they are now able to do the reverse of what they learned by appropriately responding to A comparisons after B samples. In other words, new or untrained BA performances can emerge from the explicitly reinforced AB baseline relations. Associative symmetry, or symmetry for short, is one of three emergent relations that define stimulus equivalence (Sidman & Tailby, 1982), the other two being reflexivity (the untrained ability to match each stimulus to itself) and transitivity (exhibiting untrained AC performances after explicit training on AB and BC relations). Each of these phenomena reflects the fact that stimuli related by equivalence are substitutable or interchangeable for one another.
Until recently, associative symmetry was rarely observed in studies using non-human animals (e.g., D’Amato, Salmon, Loukas, & Tomie, 1985; Dugdale & Lowe, 2000; Lionello-DeNolf & Urcuioli, 2002; Richards, 1988; Sidman, Rauzin, Lazar, Cunningham, Tailby, & Carrigan, 1982; Tomonaga, Matsuzawa, Fujita, & Yamamoto, 1991). Virtually all of these studies trained and tested subjects using n-alternative matching-to-sample like those routinely used in studies of human equivalence-class formation (e.g., Cowley, Green, & Braunling-McMorrow, 1992; Fields, Adams,Verhave, & Newman, 1990; Saunders, Drake, & Spradlin, 1999; Sidman & Tailby, 1982). Unfortunately, the different spatial locations at which the samples and comparison alternatives are presented in such tasks likely compromised these symmetry tests because the functional matching stimuli probably differed from what researchers had supposed they were. Specifically, reinforcing the selection of, say, a triangle comparison on a left or right side key after presentation of a red center-key sample during training does not necessarily mean that subjects learn to match triangle to red. Instead, subjects may learn to match triangle-on-the-left (right)-key to red-on-the-center-key (Lionello & Urcuioli, 1998; Lionello-DeNolf & Urcuioli, 2002; Urcuioli, 2007). If the functional stimuli include stimulus location, simply swapping the nominal stimuli will be insufficient to generate a viable symmetry test because, by definition, triangle-on-the-center-key (a sample for the symmetry test) is not the same stimulus as the triangle-on-the-left (right)-key (a comparison in training) and, likewise, red-on- the-left (right)-key (a comparison for the symmetry test) is not the same as the red-on-the-center-key (a sample in training).
A viable test requires that the stimuli appearing in the tested relations are functionally identical to those appearing in baseline training. One way this might be accomplished, or at least better approximated, is through training designed to negate the impact of stimulus location – e.g., by explicit training in which stimulus location varies in a manner that emphasizes its irrelevant nature. However, even when pigeons learn through such training to ignore location, they still do not exhibit symmetry in n-alternative matching (Lionello-DeNolf & Urcuioli, 2002; see also Lionello-DeNolf & Urcuioli, 2000).
Recently, though, Frank and Wasserman (2005) and Urcuioli (2008, Experiment 3) reported replicable evidence for symmetry in pigeons trained and tested in successive or go/no-go matching (Wasserman, 1976; see also Nelson & Wasserman, 1978). In successive matching task, samples and comparisons are presented individually and sequentially at a single spatial location such as the center key of a three-key apparatus. (Because location never changes, this stimulus feature is inconsequential for a viable symmetry test.) On half of the sample-comparison sequences, the first response to the comparison after some period of time (e.g., 5 or 10 s) is reinforced (“go” trials); on the other half, responding to the comparison ends without reinforcement (“no-go” trials). With training, pigeons confine most of their comparison responding to the reinforced sample-comparison combinations, with only infrequent comparison responding on the non-reinforced trials. In both of the aforementioned studies, pigeons were trained concurrently on three successive matching tasks: symbolic (AB) matching and two identity tasks (AA and BB) involving the stimuli from symbolic matching. Later, non-reinforced BA (symmetry) probe trials were inserted among the various baseline relations. Both pigeons trained in this fashion by Frank and Wasserman (2005, Experiment 1), and four of seven pigeons in Urcuioli (2008, Experiment 3) showed evidence for symmetry: They responded more to the comparisons on probe trials that reversed the reinforced symbolic baseline relations than on probe trials that reversed the non-reinforced symbolic baseline relations. For example, if pecking a triangle comparison was reinforced after a red sample in training, but pecking a horizontal-lines comparison after a red sample was not, then pigeons pecked more to a red comparison following a triangle sample than following a horizontal sample in the symmetry test.
Given these results and the failure to find symmetry in corresponding n-alternative matching versions of training (e.g., Urcuioli, 2008, Experiments 1A, 1B, and 2), Urcuioli (2008) proposed that the continual juxtaposition of non-reinforced sample-comparison combinations with the reinforced combinations throughout successive matching training generates stimulus classes containing the elements of the reinforced combinations. Thus, if a red sample – triangle comparison combination is reinforced in training, but a red sample – horizontal comparison combination is not, a stimulus class consisting of the red sample and triangle comparison will result, as depicted in the top left circle of Figure 1 where R1 designates the red sample (red in the first ordinal position within a matching trial) and T2 designates the triangle comparison (triangle in the second ordinal position). Likewise, if a green sample – horizontal comparison combination is reinforced in training, but a green sample – triangle comparison combination is not, a separate class consisting of the green sample and horizontal comparison (G1 and H2) will result, as depicted in the top right circle in Figure 1. The triangle comparison is in a different class than the green sample because this baseline combination is always non-reinforced, and likewise for the horizontal comparison and red sample. This segregation of matching stimuli into distinct classes presumably does not (or is much less likely to) occur in n-alternative matching because the frequency of non-reinforcement drops considerably as training progresses; in the limit (viz., when matching accuracy is 100% correct), it drops to zero.
Figure 1.

Top panel: The stimulus classes hypothesized to develop from the reinforced sample-comparison successive matching sequences in hue-form symbolic matching, hue identity matching, and form identity matching. Ellipses highlight common class members. Bottom panel: Two 4-member stimulus classes hypothesized to arise from the merger of the stimulus classes shown in the top panel via their common elements. Arrows indicate sample-comparison sequences to which pigeons should preferentially respond in a symmetry test assuming reinforced red sample – triangle comparison and green sample – horizontal sequences in training. R = red, G = green, T = triangle, H = horizontal, 1 = first ordinal position within a matching trial, 2 = second ordinal position within a matching trial.
Of course, a stimulus class containing the red sample and triangle comparison alone does not mean that pigeons will later respond more to a red comparison following a triangle sample. For that to occur, the latter stimuli (T1 and R2) must also be members of the same stimulus class. Urcuioli (2008) proposed that concurrently training identity matching makes this possible with an additional assumption of class merger via common elements. Specifically, hue-hue matching should yield a class containing the red sample (R1) and red comparison (R2) because this is one of the reinforced hue identity combinations. Likewise, form-form matching should yield a class containing the triangle sample (T1) and triangle comparison (T2) because this is one of the reinforced form identity combinations. These two classes (see top left half of Figure 1) plus the symbolic class share certain elements, as highlighted by the ellipses. If common elements cause their respective classes to merge, the result is a 4-member class (see bottom left of Figure 1), which contains both the reinforced hue sample – triangle comparison (R1-T2) combination from training and its predicted symmetrical counterpart (T1-R2), as denoted by the arrow.
Crucial to this prediction is the assumption (Urcuioli, 2008) that distinct stimulus classes form because half of the class-relevant sample-comparison combinations are non-reinforced throughout successive matching training. The purpose of the two experiments reported here was to test this assumption. Experiment 1 involved successive matching training like that used by Urcuioli (2008, Experiment 3) except that all symbolic (AB) trials ended in reinforcement. This was accomplished by reinforcing responding to the comparisons on half of the symbolic trials and reinforcing not-responding to the comparisons on the other half of the trials. Experiment 2 involved pair-comparison training (Pontecorvo, 1985; Shimp & Moffitt, 1977; Urcuioli & DeMarse, 1997), a hybrid of successive and n-alternative procedures. On each pair-comparison trial, the sample and comparison are presented individually on the center key (just as in successive matching) followed by a spatial (left- vs. right-key) choice (similar to n-alternative matching). One choice is always reinforced after certain sample-comparison combinations and the other is always reinforced after the remaining combinations. As this task is learned, the frequency of non-reinforcement drops just as in n-alternative matching. As with training in Experiment 1, this should preclude the development of the stimulus classes needed for symmetry.
Experiment 1
The pigeons for this experiment originally participated as one of two groups in Urcuioli (2008, Experiment 3). Each had been concurrently, but unsuccessfully, trained on symbolic (AB) and identity (AA and BB) successive matching tasks in which the first comparison peck after 5 s was reinforced on one half of the matching trials whereas not-pecking the comparison for 5 s was reinforced on the remaining matching trials. Most pigeons simply learned to wait 5 s after comparison-stimulus onset and, if food was not presented, to peck that comparison. Although an effective strategy, this behavior was not conducive to differential control over comparison responding by the sample-comparison combinations comprising each task.
To try to establish such control, these pigeons were re-trained in the present experiment on the same three successive matching tasks but with the DRO contingencies in effect for only one of them (viz., for the symbolic or AB task). For the identity tasks, non-matching sample-comparison sequences now ended after 5 s without reinforcement, whereas the matching sequences continued to be reinforced on the FI schedule as before. According to Urcuioli (2008), this altered set of baseline contingencies should generate the identity stimulus classes [R1, R2], [G1, G2], [T1, T2], and [H1, H2] shown in the top portion of Figure 1 but not the symbolic classes [R1, T2] and [G1, H2]. Without the latter, there can be no class merger because the identity classes by themselves do not share any common members, thus precluding symmetry (i.e., having T1 and R2 in the same class, for instance). In other words, the theoretical prediction is that pigeons should not respond more in testing to the reverse of the symbolic baseline relations for which comparison responding was reinforced than to the reverse of the symbolic baseline relations for which not-responding to the comparisons was reinforced.
Method
Subjects
Eight pigeons (retired breeders) obtained from the Palmetto Pigeon Plant (Sumter, SC) participated in the experiment. As just mentioned, they had originally been run as part of Urcuioli (2008, Experiment 3) but were discontinued from that study (and their data not reported) because they failed to meet the acquisition criterion despite extensive training. Their free-feeding weights had been initially established upon arrival to the lab by providing daily unlimited access to Purina Pro-Grains over a 5-10 day period. Running weights were 80% of the free-feeding values and were reached by restricted feeding over a 2-3 week period. Grit and water were available at all times in their stainless-steel, wire-mesh home cages that were located in a colony room with a 14h-10h light-dark cycle (lights on at 07:00).
Apparatus
Pigeons were run in one of two identically configured experimental chambers, each consisting of a 3-key response panel (Model PIP-016; BRS/LVE, Laurel, MD) inside a Model SEC-002 enclosure. The 2.5-cm-diameter keys were horizontally aligned in a row approximately 7.5 cm from the top of the panel and were spaced 5.7 cm apart, center-to-center. An inline projector (BRS/LVE Model IC-901-IDD) was mounted behind each key, although only the center-key projector was used in this experiment. It was equipped to display red, green, and white homogeneous fields, an inverted white triangle on a black background, and a set of three horizontal white lines also on a black background (BRS/LVE Pattern No. 692). A rear-mounted food hopper was positioned behind a 5.8-cm-square opening located 13 cm below the center key. When raised, a small miniature bulb (ESB-28) was lit inside the metal housing surrounding the food hopper so that pigeons could readily see the available grain. A constantly running blower fan (Dayton Electric Mfg. Model 4C441A) attached to the outside of each enclosure provided ventilation and masking noise. General chamber illumination was provided by a GE #1829 miniature bulb positioned 7.6 cm above the center key. Its light was directed toward the ceiling of the chamber by the opening in a small metal cover surrounding it. An IBM-386 computer controlled and recorded all experimental events.
Procedure
Preliminary training
Each pigeon was first trained to eat quickly and reliably out of the food hopper when raised for short periods of time and, afterwards, to peck white on the center key via the method of successive approximations. After pecking to white was established, each pigeon learned to peck red and green and, in separate 60-trial sessions, triangle and horizontal on the center key. Each center-key stimulus appeared 30 times per session in pseudo-random order with a single peck to each reinforced with food. The house light was on continuously throughout these initial sessions.
Next, pigeons received eight 60-trial training sessions, four with red and green and four with triangle and horizontal, during which pecking each center-key stimulus was reinforced on fixed-interval (FI) schedules. The FI parameter was initially set at 2 s, then at 3 s, and finally at 5 s for the last two sessions with each set of stimuli. Successive stimulus presentations were separated by a 15-s intertrial interval (ITI). The house light was off during the first 14 s of the ITI. It came on for the last 1 s of the ITI and remained on until the end of the reinforcement cycle. Reinforcement duration was constant within a session but could vary between 2 and 6 s across sessions as needed to maintain each pigeon’s body weight at the 80% value.
Successive matching acquisition
Concurrent training on hue-form (AB) and hue-hue (AA) and form-form (BB) successive matching began immediately after the completion of preliminary training. Table 1 shows the final training contingencies to which pigeons were exposed. The four sample-comparison combinations comprising each task are shown with sample stimuli appearing to the left of the arrows and the comparison stimuli to the right of the arrows followed by the comparison-response schedules. Each 96-trial training session was divided equally among the 12 possible trial types (4 sample-comparison combinations per task × 3 tasks) which were presented in pseudo-random order with the constraint that none occur more than twice in a row.
Table 1.
Successive Matching Training Contingencies in Experiment 1
| R → T • FI 5” | Symbolic (AB) |
| R → H • DRO 5” | |
| G → T • DRO 5” | |
| G → H • FI 5” | |
| R → R • FI 5” | Hue identity (AA) |
| R → G • EXT | |
| G → R • EXT | |
| G → G • FI 5” | |
| T → T • FI 5” | Form identity (BB) |
| T → H • EXT | |
| H → T • EXT | |
| H → H • FI 5” |
Note. R = red, G = green, T = triangle, H = horizontal, FI = fixed interval schedule, DRO = differential-reinforcement-of-other-behavior schedule, EXT = extinction. A and B denote hue and form stimulus sets, respectively. Samples are shown to the left of the arrows; comparisons are shown to the right of the arrows. Sample-response schedules and counterbalancing of the symbolic contingencies are omitted for clarity.
Each successive matching trial began with a sample stimulus appearing on the center key. The first peck to the sample initiated a 5-s interval ending with sample offset, a blank 500-ms interval, and then onset of a single comparison stimulus on the same (center) key. For both identity matching tasks, the first comparison peck to a matching comparison stimulus after 5 s turned off that stimulus and produced food (a FI 5-s schedule). On trials with a non-matching comparison stimulus, the comparison simply went off automatically after 5 s and no food was presented (EXT). For symbolic matching, the first comparison peck after 5 s on one half of the trials with each sample turned off the comparison and produced food (FI 5 s). On the remaining symbolic matching trials, not-pecking the comparison for 5 s produced food (a differential-reinforcement-of-other-behavior or DRO 5-s schedule). Any peck within 5 s of comparison onset or of a prior comparison peck reset the 5-s interval on the DRO trials. For one half of the pigeons, pecking the triangle comparison after the red sample and the horizontal comparison after the green sample was reinforced, whereas not-pecking horizontal after red and triangle after green was reinforced. For the other half of the pigeons, the opposite contingencies were in effect (not shown in Table 1).
Each successive matching trial was separated from the next by a 15-s ITI, the first 14 s of which was spent in darkness. The house light was turned on for the last 1 s of the ITI and remained on until the end of the reinforcement cycle (on reinforced trials) or comparison offset (on non-reinforced trials). Reinforcement durations were again constant within a session but varied from 2 – 6 s across sessions to maintain body weights at 80% of free-feeding.
A discrimination ratio (DR) was computed for each task by dividing the total number of comparison pecks within 5 s of comparison onset on trials on which comparison responding was reinforced by the total number of comparison pecks within 5 s of comparison onset over all trials. DRs in the neighborhood of 0.50 indicate little or no discrimination between reinforced and non-reinforced trial types (identity tasks) or between pecking-reinforced versus not-pecking-reinforced trials (symbolic task). Discrimination learning is evident as DRs increase toward the maximum value of 1.00. Each pigeon was trained in the acquisition phase until it achieved a DR of 0.80 or greater (“criterion”) on all three successive matching tasks for 5 of 6 consecutive sessions. A minimum of 10 overtraining sessions followed the last session at criterion to ensure stable performances. Overtraining ended when the DRs for all three tasks were at criterion for 5 of 6 consecutive sessions.
Again, Table 1 depicts the final successive matching training contingencies to which these pigeons were exposed. Prior to this experiment, each pigeon was trained on the same 3 sets of sample-comparison sequences but with FI versus DRO comparison-response schedules for all three tasks. Specifically, besides the symbolic matching FI versus DRO contingencies shown in Table 1, the FI 5-s comparison-response schedule was in effect for the matching sample-comparison combinations in each identity task and the DRO 5-s schedule was in effect for the non-matching combinations in each identity task. None of the 8 pigeons was able to reach criterion levels of performance with this set of contingencies even after extensive training. Consequently, all were shifted to the present set of contingencies.
Symmetry testing
After overtraining, symmetry testing began. A total of 8 test sessions were run in blocks of two, with each block separated from the next by 5 baseline sessions with performances on each successive matching task at criterion. A symmetry test session consisted of 104 trials, 96 trials divided equally among the symbolic and identity baseline tasks and 8 non-reinforced test trials in which the samples and comparisons of symbolic matching were reversed in their order of appearance. In other words, test trials consisted of either the triangle or horizontal sample followed by either the red or green comparison. Each of the four possible form sample – hue comparison combinations occurred twice in a test session. The sample-response requirement on the test trials was the same as on the baseline trials, but the comparison stimulus (and house light) went off response-independently and without food 5 s after comparison onset. The first probe trial in each symmetry-test session occurred only after each of the 12 baseline trials was presented at least once, and successive test trials were always separated from one another by at least 6 baseline trials. All remaining procedural details were identical to those described for successive matching acquisition.
Statistical analyses
Most of the data in this experiment and the one to follow were analyzed using analyses of variance (ANOVAs) and post-hoc contrasts with the critical F values reported by Rodger (1975). These values control Type I error rate (set here at .05) on a per-decision basis.
Results
Baseline performances
Prior to this experiment, all pigeons had had 92 – 116 sessions of training with FI versus DRO contingencies for all three successive matching tasks, and none had met the acquisition criteria by then. The average DRs for symbolic, hue-identity, and form-identity matching over the last 5 training sessions were 0.68, 0.78, and 0.60, respectively. Changing to the contingencies used in the present experiment (cf. Table 1) did not yield any substantive improvement in performance for 4 pigeons even after 50-60 sessions of additional training: Average DRs for symbolic, hue-identity, and form-identity matching over the last 5 training sessions for these pigeons were 0.69, 0.74, and 0.60, respectively. Consequently, their experimental participation was discontinued.
The remaining 4 pigeons were able to achieve criterion levels of performance after 61 – 70 training sessions. The average DRs for symbolic, hue-identity, and form-identity matching over their last 5 baseline sessions preceding testing were 0.89, 0.96, and 0.86, respectively. A randomized-blocks ANOVA showed an overall between-task difference in DRs, F(2, 6) = 7.98. Post-hoc contrasts revealed better discriminative performance on hue-identity matching than on symbolic and form-identity matching, F(2, 6) = 7.12, which did not differ from one another F(2, 6) = 0.85. For the most part, these pigeons maintained criterion levels of discriminative performance on the 3 baseline tasks during the test sessions themselves. The main exceptions were two birds whose form-identity DRs for fell noticeably (i.e., to 0.63 and 0.73, respectively) on their first two symmetry test sessions. In subsequent test sessions, however, their form-identity DRs were always above 0.80.
Symmetry testing
Figure 2 shows the symmetry test data (filled circles) and the symbolic matching baseline data (open circles) for the latter pigeons averaged over all test sessions. The baseline results show that each pigeon continued to respond discriminatively during testing: They pecked at much higher comparison-response rates on positive (reinforcement for comparison responding) than on negative (reinforcement for not-responding) baseline trials. The symmetry probe results show that three pigeons (DRO2, DRO3, and DRO4) responded at roughly the same rate to the comparisons on positive and negative probe trials, which individual ANOVAs confirmed, Fs(1, 62) = 0.40, 0.54, and 3.57, respectively1. Overall comparison-response rates for these pigeons on the symmetry probes were considerably lower than on the baseline symbolic trials, and this difference was apparent virtually from the beginning of testing (not shown)
Figure 2.
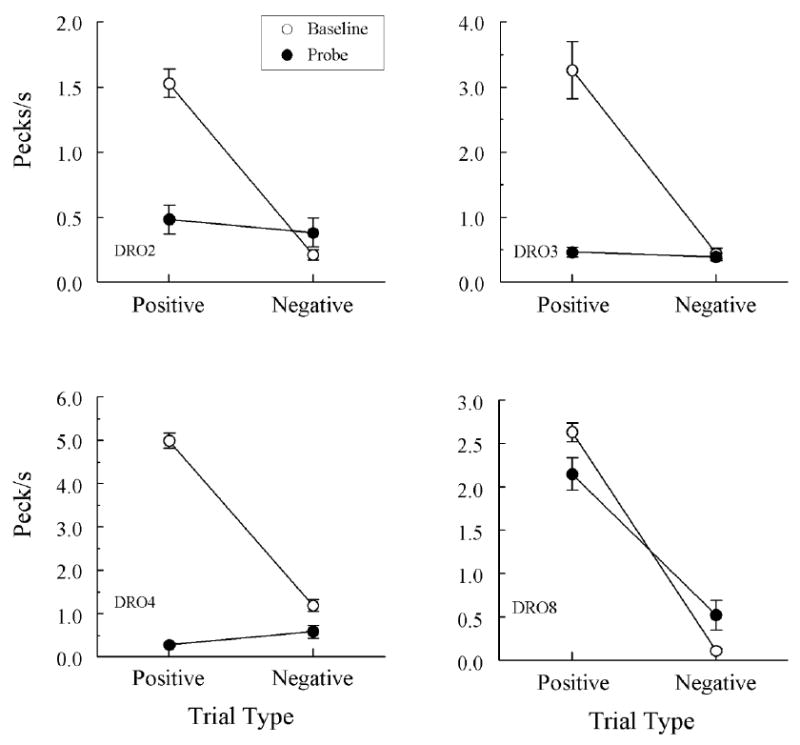
Comparison-response rates (pecks/s ± 1 SEM) on symbolic matching baseline trials (open circles) and non-reinforced symmetry probe trials (filled circles) averaged over the eight test sessions for each pigeon in Experiment 1 that met the acquisition criterion. Positive = symbolic baseline relations for which responding to the comparisons was reinforced and their symmetrical test relations. Negative = symbolic baseline relations for which not-responding to the comparisons was reinforced and their symmetrical test relations. Note the different ranges of comparison-response rates across pigeons that are identified individually in the bottom left corner of each panel.
By contrast, Pigeon DRO8 showed clear evidence of symmetry: Its comparison-response rates were considerably higher on probe trials that reversed the symbolic baseline relations on which comparison responding was reinforced (positive trials) than on probe trials that reversed the symbolic baseline relations on which not-responding was reinforced (negative trials), F(1, 62) = 39.94. Furthermore, this difference was apparent from the outset of testing: On the first two test sessions, for example, the comparison-response rate on the positive probe trials was 2.82 pecks/s versus 0.95 pecks/s on the negative probe trials, F(1, 14) = 12.67.
Discussion
This experiment tested the assumption (Urcuioli, 2008) that successive matching promotes stimulus-class formation by continually juxtaposing non-reinforced sample-comparison combinations with reinforced combinations throughout training. Thus, if responding to a triangle comparison (T2) after a red sample (R1) and to a horizontal comparison (H2) after a green sample (G1) are reinforced, but responding to a horizontal comparison after the red sample and to a triangle comparison after the green sample are not, [R1, T2] and [G1, H2] stimulus classes result. According to this hypothesis, non-reinforced training trials are essential to segregate these 4 stimuli into the two classes.
Stated otherwise, distinct stimulus classes cannot develop absent the non-reinforcement versus reinforcement contrast between the various sample-comparison combinations comprising a particular successive matching task. Thus, the symbolic matching contingencies used in the present experiment (cf. Table 1) should not yield [R1, T2] and [G1, H2] classes because reinforcement occurred after every symbolic sample-comparison combination. Indeed, if stimulus classes contain the elements of the reinforced combinations, one class (not two) should result from the FI versus DRO contingencies: [R1, T2, G1, H2]. Ignoring for the moment that such a class could not support differential responding to the baseline symbolic relations, a common-elements merger of this class with the classes arising from the two identity tasks would yield a single 8-member class containing every sample and every comparison from training. Obviously, symmetry is not uniquely predicted in this situation.
The three pigeons in testing that responded non-differentially on the symmetry probe trials performed in line with predictions. However, one pigeon (DRO8) responded differentially and, moreover, did so in a clearly symmetrical manner: It responded more to the comparisons on probe trials that reversed the “positive” (responding-reinforced) baseline relations than on probe trials that reversed the “negative” (not-responding-reinforced) baseline relations, a result which disconfirms the prediction derived from Urcuioli’s (2008) theory.
One way to reconcile the latter result with the theory is to argue that (for some pigeons, at least) withholding responding for a somewhat lengthy period of time in order to obtain reinforcement has aversive properties akin to non-reinforcement (see, for example, Singer, Berry, & Zentall, 2007; Urcuioli & DeMarse, 1994). If so, this commonality juxtaposed with a more preferred/positive situation associated with reinforced responding could generate the requisite stimulus classes for class merger and for symmetry.
Alternatively, another basis of class formation could be responding versus not-responding to the comparisons. Under this scenario, classes contain the elements of those sample-comparison combinations for which pigeons peck at the comparisons at relatively high rates. Not-pecking the comparisons for the other combinations helps to segregate the matching stimuli into separate classes. Note that in the standard successive matching task (e.g., Frank & Wasserman, 2005; Urcuioli, 2008, Experiments 3 and 4), pigeons eventually do not (or rarely) peck the comparisons on the non-reinforced baseline trials. Perhaps this not-responding contrasted with vigorous comparison responding on the other, reinforced trials is a basis of class formation.
The next experiment explored this response-based-class idea by training pigeons in a left versus right choice task in which reinforcement could be obtained on every baseline trial.
Experiment 2
In Experiment 2, pigeons were trained on a pair-comparison task (Pontecorvo, 1985; Shimp & Moffitt, 1977; Urcuioli & DeMarse, 1997) in which samples and comparisons were presented in the same fashion as in successive matching but with a left- versus right-key choice following each sample-comparison combination. After some combinations, the left-key choice was reinforced; after others, the right-key choice was reinforced. As pigeons learn these contingencies, the frequency of non-reinforcement drops substantially. Indeed, if pigeons always make the correct choice, it drops to zero (i.e., reinforcement on 100% of the trials). Thus, if the continual and equally frequent juxtaposition of non-reinforcement with reinforcement throughout training is necessary for stimulus-class formation, pair-comparison training should preclude class formation and, consequently, the possibility of observing symmetry.
At first glance, however, it would appear that such training might generate left- and right-choice stimulus classes and, with them, the prerequisites for symmetry. In other words, if classes contain the elements of the sample-comparison combinations for which a particular choice had been reinforced, merger of these classes with others sharing common elements could generate symmetry-consistent choice responding in a manner like that depicted in Figure 1. However, for reasons to be fully explained later (cf. Dube & McIlvane, 1996), the net result of this hypothesized process would be a single large class containing all of the matching stimuli, and this would equally predict symmetry-consistent and symmetry-inconsistent responding. Thus, it is of some interest to see how pigeons actually respond in testing following pair-comparison training.
Method
Subjects and Apparatus
Eight White Carneau pigeons (retired breeders) from the Palmetto Pigeon Plant (Sumter, SC) served in this experiment. All had previously participated in unrelated experiments, some of which involved choice contingencies but none of which involved within-trial presentations of multiple center-key stimuli. They were maintained at 80% of the free-feeding body weights throughout the experiment and were housed in the same fashion as the pigeons used in Experiment 1. Two experimental chambers similar to those of Experiment 1 were used with side-key projectors equipped to project a homogeneous white field onto the left and right keys.
Procedure
Preliminary training
Preliminary training was very similar to that of Experiment 1, although food-magazine training and key-peck shaping were unnecessary for these experienced pigeons. Preliminary training also included reinforced pecking to the left and right side keys whenever either was lit by a white stimulus.
Matching acquisition
Each 96-trial acquisition session involved an equal number of trials (32) with the symbolic (AB), hue-identity (AA), and form-identity (BB) sample-comparison sequences shown in Table 2. The sample and comparison stimulus for each trial appeared on the center-response key. A FI 5-s requirement was in effect for responding to both samples and comparisons timed from the first sample or comparison peck. The last sample-stimulus peck turned off the sample and initiated a blank 500-ms interval after which a single comparison stimulus appeared on the same key. The last comparison-stimulus peck turned off the comparison and immediately illuminated both side keys with a white stimulus. A single peck to either side key turned off both keys and produced either food reinforcement or an equivalent timeout period with the house light off. A left side-key choice was reinforced on all hue- and form-identity matching trials (e.g., red – red (R – R) and triangle – triangle (T – T)) and a right side-key choice was reinforced on all non-matching trials (e.g., red – green (R – G) and triangle – horizontal (T – H)). For symbolic matching, a left choice was reinforced on R – T and G – H trials and a right choice was reinforced on R – H and G – T trials for half of the pigeons (cf. Table 2); the opposite set of contingencies were in effect for the remaining pigeons. All other details of these sessions (viz., trial randomization, house light conditions, ITI length, reinforcement durations, etc.) were identical to those in Experiment 1.
Table 2.
Pair-comparison Training Contingencies in Experiment 2
| R → T – Left+ | Symbolic (AB) |
| R → H – Right+ | |
| G → T – Right+ | |
| G → H – Left+ | |
| R → R – Left+ | Hue identity (AA) |
| R → G – Right+ | |
| G → R – Right+ | |
| G → G – Left+ | |
| T → T – Left+ | Form identity (BB) |
| T → H – Right+ | |
| H → T – Right+ | |
| H → H – Left+ |
Note. R = red, G = green, T = triangle, H = horizontal, FI = fixed interval schedule, Left+ = left side-key choice reinforced, Right+ = right side-key choice reinforced. A and B denote hue and form stimulus sets, respectively. Samples are shown to the left of the arrows; comparisons are shown to the right of the arrows. Incorrect (non-reinforced) choices and counterbalancing of the symbolic left versus right contingencies are omitted for clarity.
The acquisition criterion was 5 of 6 consecutive sessions in which overall accuracy was 85% correct or better and a minimum of 26/32 correct trials (81.25%) on each of the three matching tasks. Once this criterion was met, pigeons received a minimum of 10 overtraining sessions, the last five of which were required to be at or above the acquisition criteria. Because this pair-comparison task proved to be very difficult to learn, a correction procedure was instituted for 7 of the 8 pigeons beginning on Session 36 (six pigeons) or Session 66 (one pigeon). During these sessions, any incorrect choice caused the trial to be repeated after the usual timeout period and ITI, and repeats continued until a correct choice was made. (In later sessions, the number of repeats was limited to 3.) The correction procedure remained in effect until the acquisition criteria were met, at which point it was discontinued and pigeons run to criteria without it. Even with the correction procedure, three pigeons were unable to reach criterion levels of performance after 117 – 148 additional sessions, so their experimental participation was discontinued. Choice accuracy of another pigeon that met criterion with the correction procedure dropped without it, so eventually this pigeon was kept on the correction procedure throughout the remainder of the acquisition phase (including overtraining).
Symmetry testing
Twelve symmetry tests, run in blocks of 2 sessions with each block separated by 5 of 6 consecutive baseline sessions at criterion levels of accuracy, began following overtraining. These 104-trial test sessions consisted of 96 trials divided equally among the symbolic, hue-identity, and form-identity baseline tasks and 8 symmetry probe trials. Symmetry probes reversed the symbolic sample-comparison baseline sequences: A triangle or horizontal-lines sample was followed by a red or green comparison and, then, a left versus right choice. Each of the 4 possible probe sequences occurred twice in each test session with probe-trial choices non-differentially reinforced with a probability of 0.5. When a probe-trial choice was not followed by food, the house light remained on after the side keys went off for the equivalent of the reinforcement duration. The reason these trials were not run in extinction was because it was felt that a contrast between novel, non-reinforced sample-comparison sequences and the routinely reinforced baseline sequences might quickly yield a side-key preference or entirely random responding that could obscure the detection of symmetry.
Symmetry test sessions were run without a correction procedure, including for the one pigeon that completed acquisition and overtraining with it in effect. This pigeon’s baseline recovery sessions between successive tests, however, were run with correction.
Results
Acquisition of the three pair-comparison tasks was slow and, with the exception of one pigeon (DPC6), required the use of a correction procedure. Even with the correction procedure, three of the eight pigeons were dropped from the experiment after 184, 153, and 153 sessions of training, respectively, when their performances failed to show any indication of steady improvement toward the acquisition criteria. The average accuracy for these pigeons over their last 10 training sessions was 66.7% correct for symbolic matching (range: 66.2 – 67.2% correct), 75.0% for hue-identity matching (range: 63.4 – 81.2% correct), and 62.1% correct for form-identity matching (range: 54.7 – 66.7% correct).
Four of the 5 remaining pigeons eventually met the acquisition criteria with the correction procedure in effect; the fifth did so without it. Figure 3 plots the number of training sessions required, on average, for these pigeons to match at 80% correct or better accuracy on each task. A randomized-blocks ANOVA showed a significant between-task difference, F(2, 8) = 4.42. Post-hoc contrasts indicated that pigeons required comparable amounts of training to reach criterion on symbolic matching and hue-identity matching (72.0 and 72.6 sessions, respectively; F(2, 8) = 0.00), but fewer sessions of training on these tasks than on form-identity matching (94.4 sessions), F(2, 8) = 4.42. Despite the acquisition differences, there were no between-task differences in baseline accuracies averaged over the 5 sessions preceding the first block of symmetry test sessions: 90.6%, 91.0% and 89.5% correct for symbolic, hue-identity, and form-identity matching, respectively, F(2, 8) = 0.22. The same was true for the five baseline sessions preceding each subsequent block of test sessions, all Fs(2, 8) < 1.05.
Figure 3.
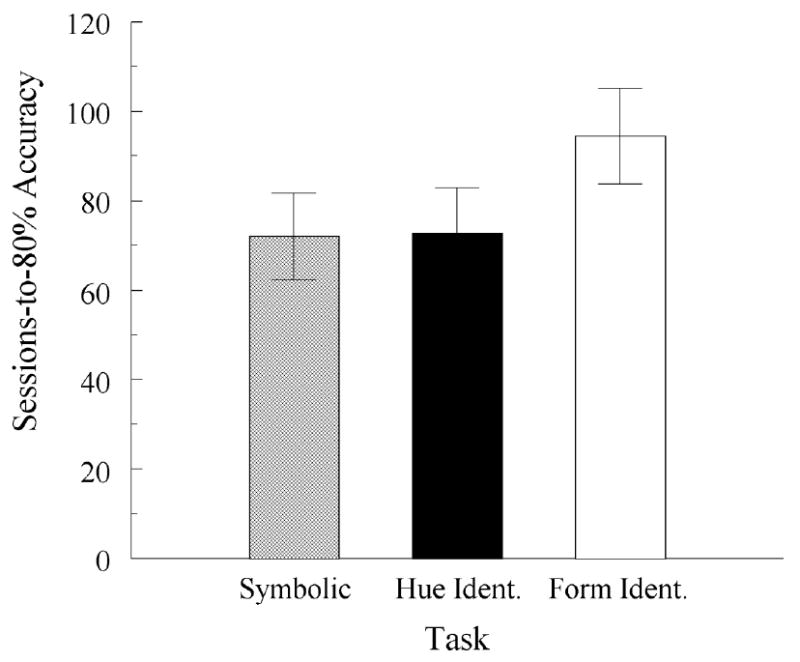
Average number of training sessions to reach 80% correct or better matching accuracy (± 1 SEM) on each component pair-comparison task in Experiment 2 for the five pigeons that met the acquisition criteria.
The symmetry tests yielded considerable session-to-session variability in probe-trial performances. For example, the percentages of symmetry-consistent choices for four of the five pigeons (DPC2, DPC3, DPC4, and DPC6) were periodically below 50%. Figure 4 shows the percentage of these choices averaged over all 12 test sessions for each of the five pigeons. The percentages were all above the chance level (dotted line) albeit not substantially so, ranging from 55.7% to 63.5% correct. Nevertheless, two-tailed t-tests indicated that accuracies were significantly above chance (noted by the stars) for all pigeons except one, ts (11) = 4.06, 2.28, 2.86, 2.73, and 1.37, respectively.
Figure 4.
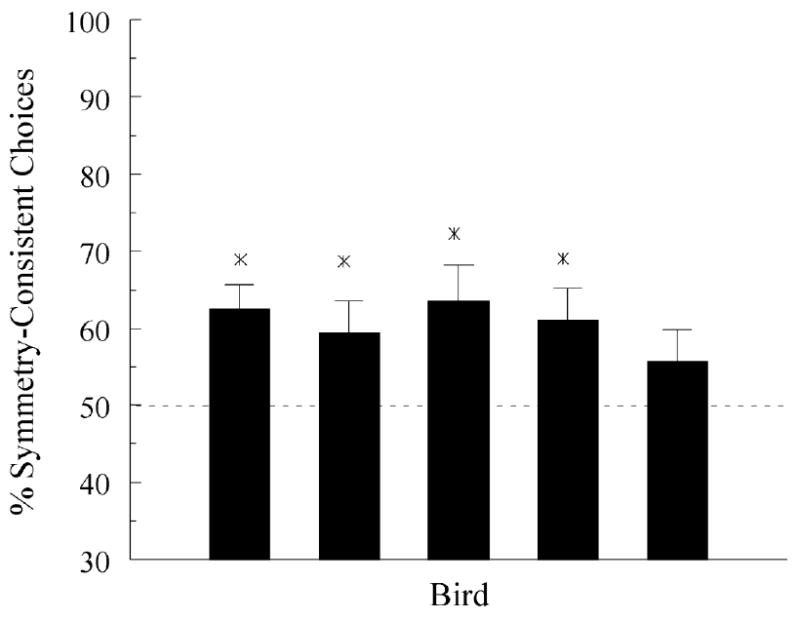
The percentage of symmetry-consistent choices for each pigeon meeting the acquisition criteria in Experiment 2 averaged over all test sessions. Stars indicate performances significantly above chance (indicated by the dashed line).
Discussion
Two findings from this experiment stand out. One was the difficulty of pigeons to acquire the concurrent pair-comparison tasks. For example, 3 of the 8 pigeons were unable to reach accuracy levels consistently above 80% correct despite over 5 months of training, most of which involved a correction procedure in a vain attempt to improve performances. Of the remaining pigeons, one was able to learn the task to high levels of accuracy without a correction procedure, but all others required it and, of the latter, one pigeon could not subsequently maintain high accuracy levels when the correction procedure was removed. The choice phase following center-key sequences appears to be responsible for these difficulties. After all, when trained with exactly the same set of 12 sample-comparison combinations in standard successive matching (i.e., with half of the trials ending in reinforcement and the other half not), pigeons are able to learn all component tasks to criterion with 1/2 the amount of training or less (Urcuioli, 2008, Experiment 3).
The second finding of note was symmetry-consistent test performances that were significantly above chance for 4 of the 5 pigeons meeting the acquisition criteria. Although there was considerable session-to-session variability in these choices, that variability may have been fueled by the fact that any probe-trial choice, symmetry-consistent or symmetry-inconsistent, was reinforced with a probability of 0.5. If pigeons quickly detected these contingencies early in testing, this variability is not surprising. From this vantage point, the average test results are noteworthy precisely because they occurred despite probe-trial contingencies that should work against symmetry. That said, reconciling the findings from this experiment with an explanation based on stimulus classes is challenging for reasons discussed in the final section.
General Discussion
The two experiments reported here were specifically designed to evaluate one of the assumptions of Urcuioli’s (2008) theory of stimulus class formation by pigeons. In particular, they tested whether continual juxtaposition of non-reinforced with reinforced sample-comparison relations throughout successive matching training is necessary for class formation and the emergent effects it generates (Sweeney & Urcuioli, in press; Urcuioli, 2008). The results of Experiment 1 provided some evidence that one of those emergent effects – symmetry – does not require equal frequencies of non-reinforced and reinforced sample-comparison relations throughout training. By using DRO contingencies, all of the sample-comparison relations for one of the three concurrently trained baseline tasks were reinforced. Nevertheless, one of four pigeons showed clear evidence of symmetry: In testing, it responded more to the comparisons on probe trials that were the reverse of the baseline symbolic relations in which comparison responding (as opposed to not-responding) was reinforced.
Although this finding contradicts the theoretical assumption mentioned above, a relatively minor modification or expansion of that assumption could encompass this result. Specifically, stimulus classes might arise from training if there is continual contrast between the aversive aspects of non-reinforcement or the requirement to withhold responding to obtain reinforcement (Singer et al., 2007) and the positive aspects of reinforced responding. Alternatively, the contrast of not-responding to the comparisons on non-reinforced trials (or on trials in which not-responding is explicitly reinforced) versus responding to the comparisons on the remaining trials might provide another mechanism for stimulus class formation. Still, these adjusted or alternative formulations do not appear to be broadly applicable given that most (3 of the 4) pigeons in Experiment 1 showed no signs of symmetry. Moreover, those 4 pigeons themselves were a select group out of the 8 that began the experiment. The rest were unable to acquire the baseline tasks to high levels of discriminative performance, suggesting that class formation might be hindered when there aren’t equal numbers of non-reinforced and reinforced baseline relations during successive matching training.
Experiment 2 used a different strategy to test the aforementioned assumption while, at the same time, creating conditions for empirically evaluating the possibility that symmetry might emerge from explicit differential response contingencies for all baseline tasks. Pigeons were presented with exactly the same sample-comparison combinations as the pigeons in Experiment 1 but were required to make a left or right choice response after each combination in order to obtain reinforcement. In principle, then, food reinforcement could occur on all pair-comparison trials if pigeons always made a correct spatial choice. But even without “perfect” performances, highly accurate performances would greatly reduce the relative frequency of non-reinforcement vis-à-vis the standard successive matching task, thus removing one of the hypothesized “supports” for stimulus-class formation. Nevertheless, of the 5 pigeons that eventually learned the pair-comparison contingencies, 4 exhibited significant symmetry-consistent choice responding in their averaged probe-trial performances, again contradicting a prediction from Urcuioli (2008). But as mentioned in the introduction to Experiment 2, these results also cannot be derived from a stimulus-class analysis in which the samples and comparisons are segregated into “left-response” versus “right-response” classes.
To understand why this is so, Table 3 shows hypothesized left- and right-response classes presumably resulting from the three pair-comparison baseline tasks of Experiment 2 (cf. Table 2). Each class consists of a sample and a comparison stimulus, denoted by the “1” and “2”, respectively, after the nominal-stimulus abbreviations. Thus, [R1, T2] refers to a [red sample, triangle comparison] class – in this case, a left-response class because a left spatial choice was reinforced after this sample-comparison sequence. Similarly, [R1, H2] refers to a [red sample, horizontal comparison] right-response class because a right spatial choice was reinforced after this particular sequence. The remaining bracketed sample-comparison classes in each column complete the list arising from the 6 baseline combinations for which a left choice response was reinforced (2 per component task) and 6 baseline combinations for which a right choice response was reinforced (2 per component task).
Table 3.
Hypothesized Stimulus Classes Resulting from the Pair-Comparison Training Tasks in Experiment 2.
| Left-Response Classes | Right-Response Classes |
| [R1, T2] | [R1, H2] |
| [G1, H2] | [G1, T2] |
| [R1, R2] | [G1, G2] |
| [G1, G2] | [G1, R2] |
| [T1, T2] | [T1, H2] |
| [H1, H2] | [H1, T2] |
| Merged Left-Response Class | |
| [R1, R2, G1, G2, T1, T2, H1, H2] | |
Note. R = red, G = green, T = triangle, H = horizontal, 1 = first ordinal position in sample-comparison sequence, 2 = second ordinal position in a sample-comparison sequence. Brackets encompass stimuli presumed to be members of a stimulus class. Italics designate individual stimuli that are common to more than one left-response class.
Some stimuli have been italicized in the Left-Response Classes column to highlight the fact that they appear in more than one left-response class – e.g., R1 is both a member of the [R1, T2] class and the [R1, R2] class. If common members cause their respective classes to merge (Sidman, Kirk, & Willson-Morris, 1985; Sidman & Tailby, 1982; see also Dube, McIlvane, Maguire, Mackay, & Stoddard, 1989), the net result would be one large 8-member class (“Merged Left-Response Class”) shown at the bottom of Table 3. Note that this merged class contains all of the matching stimuli from the 3 baseline tasks. What this means if that even though the red sample – triangle comparison (R1-T2) sequence occasioned a left response in training, the symmetrical sequence (T1-R2) and the non-symmetrical horizontal sample – red comparison (H1-R2) sequence are predicted to occasion a left response in testing – viz., a level of symmetry-consistent responding of 50%.
The common elements shared by the 6 individual right-response classes should also, via class merger, yield a corresponding 8-member right-response class (not shown). But the members of that class would be exactly the same as the members of the 8-member left-response class! Consequently, every form-hue probe-trial sequence is predicted to occasion both left and right choice responses – i.e., a level of symmetry-consistent responding of 50%. Obviously, there are no theoretical grounds here to predict symmetry following acquisition of baseline pair-comparison tasks, assuming ordinal information is coded in the functional matching stimuli and common elements cause class merger.
Despite considerable data indicating that ordinal/temporal information is naturally coded as part of the functional stimuli controlling the behavior of non-human animals (e.g., Balsam & Gallistel, 2008; Miller & Barnet, 1993; Savastano & Miller, 1998; see also Sigurdardottir, Green, & Saunders, 1990), could the symmetry test results from Experiment 2 be accommodated if the ordinal coding assumption were relinquished? In other words, could a viable stimulus-class explanation of the data be constructed by assuming that the left- and right-response-class elements are simply the nominal matching stimuli (i.e., R, G, T, and H)? The answer is “no”.
If the functional matching stimuli are the nominal ones, then the sample-comparison relations comprising the symbolic pair-comparison task would, for example, yield a [R, H] right-response class and a [G, T] right-response class (see Table 4) given that a right choice was reinforced after the red sample – horizontal and green sample – triangle comparison sequences (cf. Table 2). Similarly, the hue-identity relations would yield a [R, G] and a [G, R] right-response class (or, more simply, a single [R, G] class because stimulus order does not matter by assumption). Likewise, the form-identity relations would yield a [T, H] right-response class. Again, italicized stimuli indicate membership in more than one class. If common elements cause their respective classes to merge, the result would be a single 4-member class containing all of the nominal matching stimuli. This analysis, then, predicts a right choice in testing for both symmetrical (e.g., H-R) and non-symmetrical (e.g., H-G) probe-trial sequences. Stated otherwise, the predicted percentages of symmetry-consistent choice responding would also be 50% by this analysis.
Table 4.
Hypothesized Right-Response Stimulus Classes (Assuming No Ordinal Coding) Resulting from the Pair-Comparison Training Tasks in Experiment 2.
| Right-Response Classes | |
| [R, H] | |
| [G, T] | Merged Right-Response Class |
| [R, G] | [R, G, T, H] |
| [T, H] |
Note. R = red, G = green, T = triangle, H = horizontal. Brackets encompass stimuli presumed to be members of a stimulus class. Italics designate individual stimuli that are common to more than one right-response class.
Apparently, any stimulus-class analysis that includes an assumption about class merger via common elements will not accommodate the findings of Experiment 2. Relinquishing that assumption may solve the problem but would create new problems because without it, the symmetry and “anti-symmetry” effects reported by Urcuioli (2008) cannot be explained. Besides, it would leave the nagging question of why the pair-comparison task should ostensibly operate by a different set of stimulus class “rules” than the same procedure minus the choice phase (viz., successive matching).
Of course, the conundrum posed by the results of Experiment 2 presupposes that the significant deviations of symmetry-consistent responding from chance represent a true symmetry effect. Perhaps a fruitful next step, then, ought to be an empirical rather than a theoretical evaluation of these results. For instance, given the concern that non-differentially reinforcing pigeons’ probe-trial choices in Experiment 2 may have lowered the percentages of symmetry-consistent choices, a between-group comparison in testing might be a better approach. Specifically, pigeons could be trained on the same pair-comparison contingencies in Experiment 2 and those reaching criterion tested in one of two differentially reinforced probe-trial conditions. For one group, only probe-trial choices consistent with symmetry would be reinforced whereas for another group, only probe-trial choices inconsistent with symmetry would be reinforced. If the test results from Experiment 2 truly reflect an emergent relation of symmetry, the prediction is that the former (“consistent”) group will exhibit higher probe-trial accuracies during the course of repeated testing than the latter (“inconsistent”) group (e.g., Urcuioli, Zentall, Jackson-Smith, & Steirn, 1989, Experiment 2). If confirmed, the onus would then revert back to developing a viable theoretical analysis to explain the effect. If not, then there may not be an emergent effect to explain.
Acknowledgments
This research was supported in part by NIMH Grant 66915 and its preparation by NICHD Grant HD061322. The author thanks Timothy Burnight and Maggie Sweeney for their assistance in conducting this research.
Footnotes
The data for these analyses were the response rates on each individual matching and non-matching probe trial (4 each per test session) over all (8) test sessions.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Balsam P, Gallistel CR. Temporal maps and informativeness in associative learning. Trends in Neuroscience. 2008;32:73–78. doi: 10.1016/j.tins.2008.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cowley BJ, Green G, Braunling-McMorrow D. Using stimulus equivalence procedures to teach name-face matching to adults with brain injuries. Journal of Applied Behavior Analysis. 1992;25:461–475. doi: 10.1901/jaba.1992.25-461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D’Amato MR, Salmon DP, Loukas E, Tomie A. Symmetry and transitivity in the conditional relations in monkeys (Cebus apella) and pigeons (Columba livia) Journal of the Experimental Analysis of Behavior. 1985;44:35–47. doi: 10.1901/jeab.1985.44-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dube WV, McIlvane WJ. Some implications of a stimulus control topography analysis for emergent behavior and stimulus classes. In: Zentall TR, Smeets PM, editors. Stimulus class formation in humans and animals. New York: Elsevier; 1996. pp. 197–218. [Google Scholar]
- Dube WV, McIlvane WJ, Maguire RW, Mackay HA, Stoddard LT. Stimulus class formation and stimulus-reinforcer relations. Journal of the Experimental Analysis of Behavior. 1989;51:65–76. doi: 10.1901/jeab.1989.51-65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dugdale N, Lowe CF. Testing for symmetry in the conditional discriminations of language-trained chimpanzees. Journal of the Experimental Analysis of Behavior. 2000;73:5–22. doi: 10.1901/jeab.2000.73-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fields L, Adams BJ, Verhave T, Newman S. The effects of nodality on the formation of equivalence classes. Journal of the Experimental Analysis of Behavior. 1990;53:345–358. doi: 10.1901/jeab.1990.53-345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank AJ, Wasserman EA. Associative symmetry in the pigeon after successive matching-to-sample training. Journal of the Experimental Analysis of Behavior. 2005;84:147–165. doi: 10.1901/jeab.2005.115-04. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lionello KM, Urcuioli PJ. Control by sample location in pigeons’ matching to sample. Journal of the Experimental Analysis of Behavior. 1998;70:235–251. doi: 10.1901/jeab.1998.70-235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lionello-DeNolf KM, Urcuioli PJ. Transfer of pigeons’ matching to sample to novel sample locations. Journal of the Experimental Analysis of Behavior. 2000;73:141–161. doi: 10.1901/jeab.2000.73-141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lionello-DeNolf KM, Urcuioli PJ. Stimulus control topographies and tests of symmetry in pigeons. Journal of the Experimental Analysis of Behavior. 2002;78:467–495. doi: 10.1901/jeab.2002.78-467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller RR, Barnet RC. The role of time in elementary associations. Current Directions in Psychological Science. 1993;2:106–111. [Google Scholar]
- Nelson KR, Wasserman EA. Temporal factors influencing the pigeon’s successive matching-to-sample performance: Sample duration, intertrial interval, and retention interval. Journal of the Experimental Analysis of Behavior. 1978;30:153–162. doi: 10.1901/jeab.1978.30-153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pontecorvo MJ. Memory for a stimulus versus anticipation of a response: Contrasting effects of proactive interference in delayed comparison tasks. Animal Learning & Behavior. 1985;13:355–364. [Google Scholar]
- Richards RW. The question of bidirectional associations in pigeons’ learning of conditional discrimination tasks. Bulletin of the Psychonomic Society. 1988;26:577–579. [Google Scholar]
- Rodger RS. The number of non-zero, post hoc contrasts from ANOVA and error rate. I. British Journal of Mathematical and Statistical Psychology. 1975;28:71–78. [Google Scholar]
- Saunders RR, Drake KM, Spradlin JE. Equivalence class establishment, expansion, and modification in preschool children. Journal of the Experimental Analysis of Behavior. 1999;71:195–214. doi: 10.1901/jeab.1999.71-195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savastano HI, Miller RR. Time as content in Pavlovian conditioning. Behavioural Processes. 1998;44:147–162. doi: 10.1016/s0376-6357(98)00046-1. [DOI] [PubMed] [Google Scholar]
- Shimp CP, Moffitt M. Short-term memory in the pigeon: Delayed-pair-comparison procedures and some results. Journal of the Experimental Analysis of Behavior. 1977;28:13–25. doi: 10.1901/jeab.1977.28-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidman M, Kirk B, Willson-Morris M. Six-member stimulus classes generated by conditional-discrimination procedures. Journal of the Experimental Analysis of Behavior. 1985;43:21–42. doi: 10.1901/jeab.1985.43-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidman M, Rauzin R, Lazar R, Cunningham S, Tailby W, Carrigan P. A search for symmetry in the conditional discrimination of rhesus monkeys, baboons, and children. Journal of the Experimental Analysis of Behavior. 1982;37:23–44. doi: 10.1901/jeab.1982.37-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidman M, Tailby W. Conditional discrimination vs. matching-to-sample: An expansion of the testing paradigm. Journal of the Experimental Analysis of Behavior. 1982;37:5–22. doi: 10.1901/jeab.1982.37-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sigurdardottir ZG, Green G, Saunders RR. Equivalence classes generated by sequence training. Journal of the Experimental Analysis of Behavior. 1990;53:47–63. doi: 10.1901/jeab.1990.53-47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singer RA, Berry LM, Zentall TR. Preference for a stimulus that follows a relatively aversive event: Contrast or delay reduction? Journal of the Experimental Analysis of Behavior. 2007;87:275–285. doi: 10.1901/jeab.2007.39-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sweeney MM, Urcuioli PJ. Reflexivity in pigeons. Journal of the Experimental Analysis of Behavior. doi: 10.1901/jeab.2010.94-267. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomonaga M, Matsuzawa T, Fujita K, Yamamoto J. Emergence of symmetry in a visual discrimination by chimpanzees (Pan troglodytes) Psychological Reports. 1991;68:51–60. doi: 10.2466/pr0.1991.68.1.51. [DOI] [PubMed] [Google Scholar]
- Urcuioli PJ. Sample and comparison location as factors in matching acquisition, transfer, and acquired equivalence. Learning & Behavior. 2007;35:252–261. doi: 10.3758/bf03206431. [DOI] [PubMed] [Google Scholar]
- Urcuioli PJ. Associative symmetry, “anti-symmetry”, and a theory of pigeons’ equivalence-class formation. Journal of the Experimental Analysis of Behavior. 2008;90:257–282. doi: 10.1901/jeab.2008.90-257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urcuioli PJ, DeMarse T. On the relationship between differential outcomes and differential sample responding in matching-to-sample. Journal of Experimental Psychology: Animal Behavior Processes. 1994;20:249–263. doi: 10.1037//0097-7403.20.3.249. [DOI] [PubMed] [Google Scholar]
- Urcuioli PJ, DeMarse T. Memory processes in delayed spatial discriminations: Response intentions or response mediation? Journal of the Experimental Analysis of Behavior. 1997;67:323–336. doi: 10.1901/jeab.1997.67-323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urcuioli PJ, Zentall TR, Jackson-Smith P, Steirn JN. Evidence for common coding in many-to-one matching: Retention, intertrial interference, and transfer. Journal of Experimental Psychology: Animal Behavior Processes. 1989;15:264–273. [Google Scholar]
- Wasserman EA. Successive matching-to-sample in the pigeon: Variation on a theme by Konorski. Behavior Research Methods & Instrumentation. 1976;8:278–282. [Google Scholar]