

Abstract
A new type II restriction endonuclease designated OliI has been partially purified from the halophilic bacterium Oceanospirillum linum 4-5D. OliI recognizes the interrupted hexanucleotide palindrome 5′-CACNN↓NNGTG-3′ and cleaves it in the center generating blunt-ended DNA fragments.
INTRODUCTION
Among the 229 different type II restriction endonucleases (REBASE v.011) (1), there is a significant group of enzymes that cleave palindromic sequences interrupted by some ambiguous nucleotides. We describe here the isolation and characterization of a new representative of this group of enzymes, the restriction endonuclease OliI from Oceanospirillum linum 4-5D, which recognizes the interrupted hexanucleotide 5′-CACNN↓NNGTG-3′ and cleaves it in the centre of the recognition sequence yielding blunt-ended DNA fragments.
MATERIALS AND METHODS
Biological materials
Oceanospirillum linum strain 4-5D was from MBI Fermentas culture collection. The cells were grown aerobically at 37°C in a medium containing 10 g/l peptone, 5 g/l yeast extract (both from Oxoid), 30 g/l NaCl and 10 g/l MgSO4·7H2O, pH 7.3, up to late-logarithmic phase, collected by centrifugation and stored at –20°C until use. Phage λ, plasmid DNAs, Cycle Reader DNA Sequencing kit and all restriction endonucleases used were products of MBI Fermentas. SssI methylase was purchased from New England Biolabs, Ad2 DNA from Sigma and [α-33P]dATP from Amersham.
Endonuclease assay
The restriction endonuclease activity was assayed in 50 µl of MBI Fermentas R+ buffer (10 mM Tris–HCl pH 8.5, 100 mM KCl, 10 mM MgCl2 and 0.1 mg/ml BSA). Column samples (1 µl) were added to the reaction mixture, incubated for 1 min at 37°C and an aliquot from each solution was then electrophoresed in a 0.7% agarose gel.
Purification of restriction endonuclease
All steps were carried out at 4°C. Frozen cells (50 g wet weight) of a log-phase culture were thawed in 200 ml buffer A (10 mM potassium phosphate pH 7.0, 1 mM EDTA, 7 mM 2-mercaptoethanol), containing 0.1 M NaCl. After sonication, insoluble material was removed from the crude extract by centrifugation at 30 000 g for 1 h.
The crude extract was applied to a heparin–Sepharose column (1.5 × 26 cm) pre-equilibrated with buffer A, 0.1 M NaCl. The column was washed with the same buffer and eluted with a 500 ml linear gradient from 0.1 to 1.0 M NaCl. Fractions of 10 ml were collected and assayed for endonuclease activity. The restriction endonuclease OliI eluted at ~0.75–0.83 M NaCl. Active fractions were pooled and dialysed against buffer A, 0.1 M NaCl.
The enzyme pool from heparin–Sepharose was applied to an AH–Sepharose column (1.5 × 24 cm) equilibrated with the dialysis buffer. The column was washed with the same buffer and eluted with a 500 ml linear gradient from 0.1 to 1.0 M NaCl in buffer A. Fractions of 10 ml were collected. OliI activity eluted from the column at ~0.38–0.46 M NaCl. These fractions were pooled and dialysed against buffer A, 0.1 M NaCl.
Active fractions from AH–Sepharose after dialysis were applied to a phosphocellulose P11 column (1.5 × 20 cm), equilibrated with buffer A, 0.1 M NaCl. The column was washed with the same buffer and subsequently developed with a 400 ml linear gradient from 0.1 to 1.0 M NaCl in buffer A. Fractions of 8 ml were collected. The peak of the restriction endonuclease activity eluted at 0.64–0.73 M NaCl. Active fractions were pooled and dialysed against 10 mM Tris–HCl pH 7.4, 100 mM KCl, 1 mM EDTA, 1 mM DTT and 50% glycerol. The final preparation was stored at –20°C. The yield of the enzyme was ~30 000 U/g wet weight of cells (1 U of enzyme is the amount required to hydrolyze 1 µg of λ DNA in 60 min in a total reaction volume of 50 µl).
Determination of the recognition sequence and cleavage site
The recognition sequence of OliI was inferred by restriction mapping of the recognition sites on the DNAs of phages λ, T7, φX174 and M13mp18, Adenovirus-2 and plasmids pBR322 and pUC18. Then the fragments predicted by cleavage of the inferred recognition sites were compared with the observed restriction fragments from OliI cleavage of the DNAs.
λ DNA was used as a template to characterize the cleavage site of OliI. A 20mer oligodeoxyribonucleotide complementary to λ between positions 1640 and 1660 was used in direct sequencing through the OliI site located at position 1712. Four dideoxy sequencing reactions (lanes G, A, T and C) using [α-33P]dATP and the Cycle Reader DNA Sequencing kit (MBI Fermentas) were carried out. The same primer and template were used in an extension reaction, which also included T7 DNA polymerase, dNTP and [α-33P]dATP. The extension reaction was heat inactivated, and then radiolabeled DNA was digested by OliI and the reaction mixture was divided into two. One sample was treated with T4 DNA polymerase. Both samples were diluted with sequencing dye solution and loaded onto a standard sequencing gel together with the dideoxy sequencing reactions.
RESULTS
The recognition sequence and cleavage site of OliI
To determine the substrate specificity of the enzyme, DNA substrates (λ phage, T7 phage, Adenovirus-2, plasmid pBR322, pUC18, φX174 and M13mp18 DNAs) were incubated at 37°C in 50 µl of MBI Fermentas R+ buffer (10 mM Tris–HCl pH 8.5, 100 mM KCl, 10 mM MgCl2 and 0.1 mg/ml BSA) containing 1 µg DNA (Fig. 1). OliI did not cleave pBR322 and pUC18 DNAs. The single OliI cleavage site on φX174 DNA was mapped to approximate position 2900 using Eam1105I, AatII, BcnI and MunI. Restriction endonucleases BglII, Bsp1407I, Cfr10I, NsbI and SmiI were used to find the OliI cleavage position in M13mp18 DNA (approximate position 6580). Seven of 10 OliI cleavage sites on Adenovirus-2 DNA (approximate positions 580, 10 100, 11 600, 14 350, 20 350, 28 700 and 29 150) were mapped by double digestion with BamHI, BoxI, Bsp119I, Bsu15I, CpoI, MssI, SmiI and XmaJI (data not shown). After digestion of T7 DNA with OliI in combination with Alw44I, MluI, Eco72I, PsyI, BglII, Eco24I, Eco147I, NheI and NcoI, five OliI cleavage sites at approximate positions 14 650, 19 200, 23 600, 23 750 and 43 300 were localized. A computer-aided search of homologous nucleotide sequences at all the mapped OliI sites revealed only one common sequence, 5′-CACNNNNGTG-3′, for all of them except the cleavage site on M13mp18 DNA. In contrast to our observation (Fig. 1, lane 6) that OliI cleaves M13mp18 DNA at approximate position 6580, the sequence 5′-CACNNNNGTG-3′ was not found in M13mp18 DNA. The computer-generated cleavage pattern of various DNAs suggested this to be the correct recognition sequence for all DNAs (λ, Adenovirus-2, T7, φX174, pUC18 and pBR322) except for M13mp18. Examination of the published sequence surrounding the supposed non-existing cleavage site in M13mp18 DNA (GenBank/EMBL accession no. M77815) showed that there was a sequence 5′-CACNNNNGTA-3′ at positions 6572–6581, which differs from the OliI target sequence by 1 nt. This region was resequenced and a ‘G’ band was found instead of the ‘A’ at position 6581 (data not shown). These results confirm the interrupted hexanucleotide 5′-CACNNNNGTG-3′ as a recognition sequence of OliI.
Figure 1.
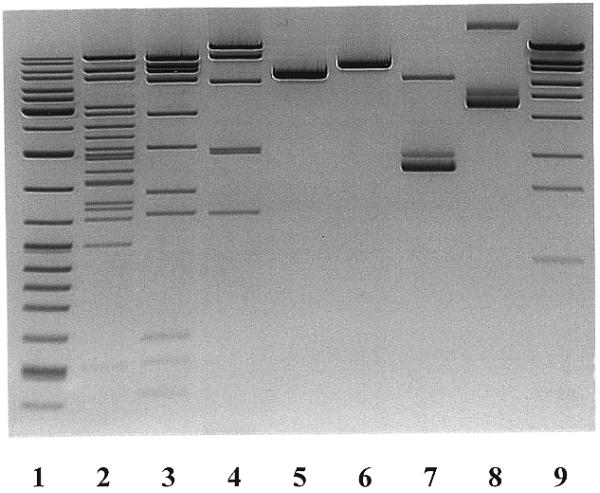
Cleavage of various DNAs with OliI. Lane 1, GeneRulerTM DNA Ladder Mix as size markers; lane 2, λ DNA + OliI; lane 3, Adenovirus-2 DNA + OliI; lane 4, T7 DNA + OliI; lane 5, φX174 DNA + OliI; lane 6, M13mp19 DNA + OliI; lane 7, pUC19 DNA + OliI; lane 8, pBR322 DNA + OliI; lane 9, λ DNA–Eco130I digest as size markers.
The cleavage site of OliI was determined by comparison of dideoxy sequencing ladders with fragments generated by OliI cleavage and T4 DNA polymerase action on the digestion product. The results of the determination of the OliI cleavage site are shown in Figure 2. The fragment generated by OliI digestion co-migrates with the T-band of the sequence ladder (Fig. 2, lane 1) through the OliI recognition site. From these results it can be inferred that OliI cleaves DNA between the fifth and sixth nucleotides, starting from the 5′-end of the OliI recognition sequence as indicated by 5′-CACCT↓GGGTG-3′. Figure 2 (lane 2) shows the result when the fragment produced by OliI digestion was further treated with T4 DNA polymerase. The single band obtained by the exonuclease activity of the T4 DNA polymerase also co-migrates with the T-band of the sequence ladder CACCTGGGTG and indicates the cleavage point on the complementary DNA strand. These results would therefore indicate that DNA cleavage by OliI generates blunt ends and the cleavage specificity of OliI is 5′-CACNN↓NNGTG-3′.
Figure 2.
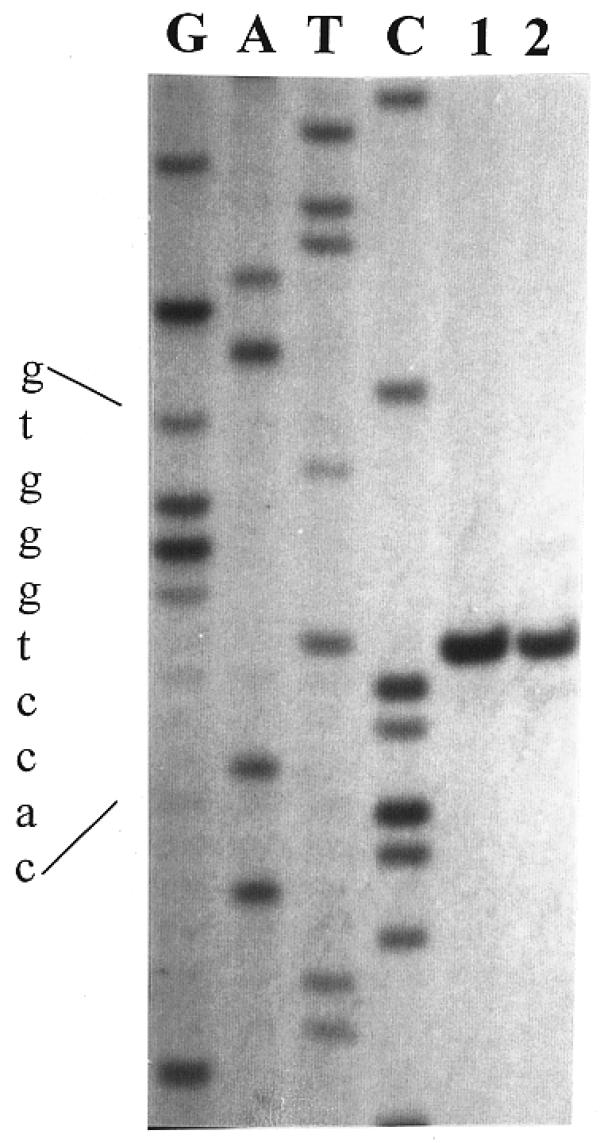
Determination of the OliI cleavage site. Lanes G, A, T and C, the sequence ladders through the OliI site; lane 1, the product of the primed synthesis reaction cleaved with OliI; lane 2, the T4 DNA polymerase action on the OliI digest.
Enzymatic properties
As would be expected for a restriction enzyme isolated from a halophilic microorganism, the enzyme shows the highest activity in a high salt concentration buffer (100–300 mM KCl) and the optimum pH is ∼8.5.
Since the OliI recognition sequence may overlap with CG sequences due to the unspecified internal nucleotides, the enzyme sensitivity to such methylation was investigated. Eight of 20 OliI sites on λ DNA overlap CG at one part of the recognition sequence (5′-CACGNNNGTG-3′) and one at both parts (5′-CACGNNCGTG-3′). After methylation of λ DNA with SssI methylase, 5 U of OliI was sufficient to cleave 1 µg of DNA at all sites except the one that overlaps CG at both parts of recognition sequence. Even with 30 U of enzyme only partial cleavage of this site was achieved.
DISCUSSION
At present, the known discontinuous palindromic target sequences for restriction endonucleases contain from one to nine unspecified nucleotides between the flanking nucleotides recognized by the enzyme. Only three among them, BsaBI, PshAI and XmnI (Z.Chen and H.Kong, unpublished observations; 2,3), recognize hexanucleotide sequences interrupted by four unspecified nucleotides. All of them cleave DNA generating blunt ends. OliI belongs to this group of specific endonucleases. 5′-CACNN↓NNGTG-3′ is a new addition to the list of nucleotide sequences known to be recognized by restriction enzymes. OliI should cleave some MslI sites (5′-CAYNN↓NNRTG-3′) if Y is C and R is G. The enzyme represents a new tool for structural analysis and molecular cloning of DNA.
Acknowledgments
ACKNOWLEDGEMENTS
The authors would like to thank Audra Rukenaite for resequencing of the M13mp18 DNA region containing the OliI site and Egle Cesnaviciene for help in preparation of the manuscript.
References
- 1.Roberts R.J. and Macelis,D. (2000) REBASE – restriction enzymes and methylases. Nucleic Acids Res., 28, 306–307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Miyahara M., Nakajima,K., Shimada,T. and Mise,K. (1990) Restriction endonuclease PshAI from Plesiomonas shigelloides with the novel recognition site 5′ GACNN/NNGTC. Gene, 87, 119–122. [DOI] [PubMed] [Google Scholar]
- 3.Lin B.-C., Chien,M.-C. and Lou,S.-Y. (1980) A sequence-specific endonuclease (XmnI) from Xanthomonas manihotis. Nucleic Acids Res., 8, 6189–6198. [DOI] [PMC free article] [PubMed] [Google Scholar]