

Abstract
Since its identification in 1995, TRAIL (TNF-Related Apoptosis Inducing Ligand) has sparked growing interest in oncology due to its reported ability to selectively trigger cancer cell death. Contrary to other members of the TNF superfamily, TRAIL administration in vivo is harmless. The relative absence of toxic side effects of this naturally occurring cytokine in addition to its antitumoral properties has led to its preclinical evaluation. However, despite intensive investigations, little is known with regard to the mechanisms underlying TRAIL selectivity or efficiency. Appropriate understanding of its physiological relevance, regulatory pathways and of the mechanisms controlling cancer cells escape to TRAIL-induced cell death will be required to optimally use the cytokine in clinics. The current review focuses on recent advances in the understanding of TRAIL signal transduction and discusses the current and future challenges of TRAIL-based cancer therapy development.
Keywords: Animals, Antineoplastic Agents, administration & dosage, Apoptosis, drug effects, physiology, Drug Delivery Systems, trends, Forecasting, Humans, Neoplasms, drug therapy, metabolism, pathology, Signal Transduction, drug effects, physiology, TNF-Related Apoptosis-Inducing Ligand, administration & dosage
Keywords: Apoptosis, cancer, resistance, TRAIL, TRAIL-receptor agonistic antibodies, TRAIL-Receptors
1. Introduction
During the past 20 years empirically designed chemotherapy regimens have diminished the risks of death in patients suffering from cancer. Ongoing attempts to improve chemotherapy efficacy are now entering new era with rationally designed studies based on specific understanding of the mechanisms of action, interactions and pharmacology. Recent breakthroughs promise even more successful outcomes, particularly with the advent of more selective therapeutic approaches. Among the latter, the TRAIL system should show encouraging levels of efficacy. TRAIL has now been recognized, as one of the few tumour selective agents able to eradicate cancer cells selectively by activating a signalling pathway that is used by the innate immune system, opening an entirely novel dimension to cancer research and therapy.
2. TRAIL: a natural anti-tumoral cytokine?
TRAIL, also known as Apo2L [1,2], is a type 2 membrane protein belonging to the TNF superfamily. The human TRAIL ligand interacts with 2 agonistic receptors (TRAIL-R1/DR4 and TRAIL-R2/DR5/KILLER) [3,4,5], and 3 antagonistic decoy receptors (TRAIL-R3/DcR1/TRID, TRAIL-R4/DcR2/TRUNDD and OPG) [6,7,8]. TRAIL is a remarkable member of the TNF ligand family, due to its unique ability to bind to 5 receptors. The reasons for this remain obscure. However TRAIL’s binding abilities suggest that its physiological functions as well as the molecular mechanisms involved in the regulation of its signalling pathways could be more complex than anticipated.
The primary role of TRAIL appears to be the triggering of apoptosis. Upon TRAIL binding, DR4 and/or DR5 aggregate, enabling the recruitment of the adaptor protein FADD through homotypic interactions between their respective DD (Death Domain). In turn, FADD recruits the initiator caspases-8 and -10 through interaction of their respective DED (Death Effector Domain). Initiator caspases are activated within this molecular platform, known as DISC (Death Inducing Signalling Complex) and released to the cytosol where they trigger the proteolytic cascade leading to apoptosis (Figure 1). In addition to the membrane-bound DISC, TRAIL binding to its cognate agonistic receptors was recently shown to trigger the formation of a secondary complex allowing the activation of signalling pathways such as JNK, p38 or NF-κB [9]. Interestingly, the sequential signalling pathways engaged by TRAIL and its agonistic receptors are in sharp contrast to TNFR1. TNFR1 engages primarily a membrane-bound complex that triggers cell survival in most cases, followed by a secondary complex for the trigger of apoptosis. However, activation of the apoptotic machinery by this secondary complex only occurs when the survival pathway, activated by complex I, is impaired [10]. Like TNFR1, TRAIL non-apoptotic complex II is thought to arise from complex I or TRAIL DISC. These sequentially occurring signalling platforms provide excellent molecular explanations for the pleiotropic properties of TRAIL.
Figure 1.
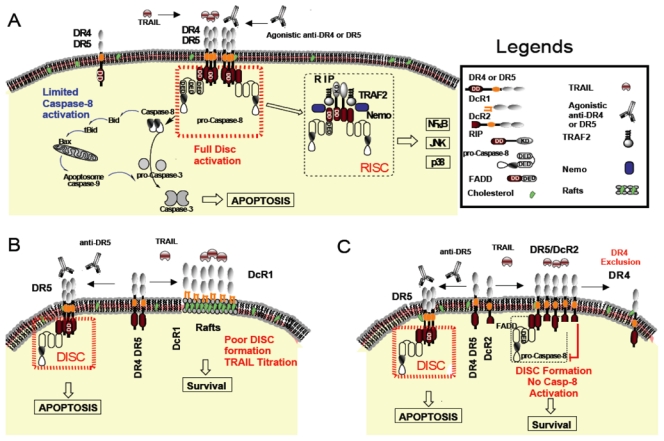
A- DR4/DR5 receptor aggregation upon TRAIL treatment allows DISC formation, and caspase-8 processing. Cleaved caspase-8 can directly activate the effector caspases, or induce the mitochondrial amplification loop. (B) In presence of DcR1, TRAIL does not induce DISC formation, because of TRAIL titration in lipid rafts, whereas an anti-DR5 agonistic antibody allows DR5 aggregation and subsequent caspase-8 activation. (C) DcR2 interacts with DR5 and the processing of caspase-8 is inhibited. The anti-DR5 antibody induces cell death in DcR2 expressing cells.
TRAIL has been shown to be expressed in diverse tissues, such as liver, lung, placenta, kidney, spleen, immune-privileged sites and immune cells, at the protein or mRNA level [11]. TRAIL is expressed by immune cells, such as human PBL (Peripheral Blood Lymphocytes) following anti-CD3 and IFNγ stimulation [12] or monocytes and neutrophils upon IFNγ treatment [13,14]. In some CD4+ T cell clones, the constitutive expression of TRAIL was shown to contribute to perforin-and granzyme B-independent cytotoxicity [15]. TRAIL was also proposed to play a role in the cytotoxic activity of dendritic cells [16,17,18,19,20], including umbilical cord blood dendritic cells [21]. Constitutive TRAIL expression has also been found in freshly isolated neutrophils [22]. In some circumstances, TRAIL is released into the circulation upon viral infections [23,24,25], and in some autoimmune [26], inflammatory [27] or malignant [23,25,28] diseases[29].
TRAIL expression pattern could be more restricted than initially expected. Accordingly, its physiological functions have mainly been shown to occur in the immune system. The first demonstrations that TRAIL played a role in immune anti-tumour response were based on studies using neutralizing anti-TRAIL antibodies and experiments obtained from TRAIL and TRAIL-receptor gene knockout mice. TRAIL deficient mice do not present developmental deficiencies, but display an increased susceptibility to experimental and spontaneous tumour formation due to impaired NK cells anti-tumour activity [30,31,32,33,34]. In line with these findings, IFNγ-induced TRAIL expression in mouse liver NK cells, was shown to be required to inhibit liver metastasis formation in vivo [30,35,36,37]. Yet, the exact function of TRAIL in modulating the anti-tumour response remains poorly defined. For instance, while on the one hand TRAIL deficient mice display reduced tumour immune surveillance, TRAIL-receptor knock out mice on the other hand exhibit enhanced innate immune response [38]. Thus the impact of recombinant TRAIL administration towards immune homeostasis still deserves careful analysis.
TRAIL appears to increase the growing list of immune cell weapons, joining Granzyme B, Fas or TNF. Like them however, TRAIL remains dispensable to elicit an efficient immune response. The redundancy in cytotoxic molecules secreted by immune cells certainly allows for a better defence against a wide panel of cancer cells but also probably accounts for our difficulty to clearly define the physiological functions of TRAIL.
3. Resistance of cancer cells
TRAIL was initially reported to kill more easily tumour than normal cells, which suggested that the cytokine could be of great interest in cancer treatment. TRAIL cancer cell selectivity was further demonstrated recently using stepwise tumorigenic cellular systems [39,40]. In both cases, while normal cells or immortalized cells remained unresponsive to TRAIL, cellular transformation-induced either by Ras or c-Myc ectopic expression restored cell sensitivity to TRAIL-induced cell death. However, numerous tumour cells also exhibit a relative resistance to TRAIL-induced apoptosis [41,42,43]. Resistance to TRAIL can be explained by the different mechanisms described below.
Upstream regulators
The first level of regulation of TRAIL-sensitivity is related to the level of expression at the cell surface of TRAIL cognate receptors. A deficit in the expression of agonistic receptors [44,45,46,47], or an unusual DR4/DR5 ratio [48,49] have been shown to account for the observed resistance in some cancer cells. Cell surface expression of these TRAIL agonistic receptors is the first requirement for the triggering of the apoptotic machinery by TRAIL. However, mutations within critical domains (such as the death domain or the ligand-binding domain) of either DR4 [50,51], or DR5 [52,53] can also hamper TRAIL-induced cell death, leading to a “decoy receptors”-like phenotype.
TRAIL antagonistic receptors DcR1 and DcR2 were initially coined decoy receptors since they lack functional death domain and cannot recruit DISC components. Originally these receptors, which confer a specific resistance to TRAIL-induced cell death, were proposed to be expressed in normal tissues and down-regulated in tumour cells [54]. Accordingly, DcR1 and DcR2 expression was found to be impaired in various tumour types, such as neuroblastomas, primary breast or lung cancers, due to promoter hypermethylation [55,56]. However, DcR are also expressed in some primary tumours such as gastrointestinal [57], prostate [58], lung [59] and acute myeloid leukemia cancer cells [60] (Table I). In addition, the correlation between DcR expression and TRAIL-resistance remains highly controversial [61,62,63,64,65]. This discrepancy could be partly explained by the fact that the expression of these receptors is not always explored at the tumour cell surface but at the mRNA or protein level in whole cell extracts [51,66,67] (Table I). Alternatively, this lack of correlation can also be related to the presence of other inhibitors of the TRAIL-apoptotic pathway, in the absence of DcR expression.
It has been suggested that DcR may compete with agonistic receptors for TRAIL binding [6,7,68]. The affinity of decoy receptors to TRAIL was initially reported to be lower than that of agonistic DR5 [69], which was not confirmed in other experimental settings [70]. If decoy receptors are not able to recruit FADD, but bind TRAIL at the membrane level with an affinity similar to that of agonistic receptors, they may act as “dominant negative” receptors. Then, the DR/DcR ratio at the cell membrane could be a determinant point as regards TRAIL-sensitivity. Ectopic over-expression of DcR1 or DcR2, in TRAIL sensitive cells, impairs TRAIL-induced apoptosis provided that DcR are expressed at the cell surface [6,7,8,71,72,73]. Accordingly, removal of DcR1 expression, by use of PI-PLC, a molecule that sheds GPI-anchored proteins from the membranes [74,75], or inhibition of TRAIL binding to DcR1 using blocking anti-DcR1 antibody (Personal communication), restores TRAIL sensitivity in DcR1-expressing cells. Similar results were obtained using an anti-DcR2 blocking antibody [48] or siRNAs to down-regulate DcR2 expression in cells that endogenously express DcR2 at their surface [58,76,77].
While both receptors share the same inhibitory function, they are structurally different. DcR1 is a GPI anchored membrane protein devoid of intracellular domain, while DcR2 is a transmembrane receptor containing a truncated death domain. We have recently shown that whereas DcR1 sequesters TRAIL within lipid rafts, thus titrating TRAIL competitively and preventing DISC formation, DcR2 allows DISC formation but impairs caspase-8 activation [72]. Interestingly, DcR1 is unable to interact with any of the agonistic receptors, while DcR2 interacts with DR5 in a TRAIL-dependent manner. It has been recently proposed that DcR2 could interact spontaneously with DR5 though the PLAD (PreLigand Associated Domain) [78]. In our hands, although weak interaction could readily be found at the steady state, a stronger interaction was obtained upon TRAIL stimulation, including in cells expressing endogenous DcR2 [72,79]. How DcR2 impairs caspase-8 activation is still unclear. Caspase-8 activation within TRAIL or Fas DISC has been shown to require close proximity, therefore it is probable that DcR2 recruitment within the DR5 DISC impinge caspase-8 activation by steric hindrance. The absence of a functional death domain within DcR2 intracellular region explains its inability to recruit initiator caspases. Yet, this trans-membrane receptor could nevertheless account for the activation of survival pathways such as NF-κB [8], or allow the recruitment of a yet to be determined inhibitory protein that would prevent caspase-8 activation within the DISC.
Downstream regulators
Besides the presence of DcR, TRAIL-induced cell death can be inhibited by a plethora of less specific intracellular mediators, at the level of the DISC or downstream. At the DISC level, a deficiency in caspase-8 and caspase-10 expression, mutations affecting their function [80], gene promoter methylation [41,81,82], or CARP-dependent mediated degradation [83] have been shown to prevent TRAIL-induced cell death. Elevated expression of FLIP (FLICE-inhibitory protein) also results in TRAIL resistance [84,85].
Downstream of the TRAIL DISC, tumour resistance can be mediated by overexpression or mutations of Bcl-2 [86] or IAP (Inhibitor of Apoptosis Proteins) family members [87,88].
While the main function of TRAIL is the induction of cell death, accumulating evidences also suggest that this ligand is able to signal non-apoptotic pathways, such as Akt, NF-κB and MAPK, involved in cell survival and proliferation [89,90,91]. Akt kinase was proposed to play a protective role against TRAIL toxicity in many tumour cells [92,93,94]. Yet some Akt inhibitors such as amiloride [95], complestatin [96], quercetin [97], sulforaphane [97], arsenic [98] or EGFR inhibitor [99] enhance TRAIL-mediated apoptosis. On the other hand however, more specific inhibitors of Akt (e.g. LY294002, Wortmannin, PI3K siRNA) were shown to be potent TRAIL inhibitors [100,101,102,103].
TRAIL is thought to induce NF-κB activation through the serine/threonine kinase RIP1 [104,105]. Likewise, the NF-kB pathway is thought to inhibit TRAIL-induced cell death, but the issue remains controversial. NF-κB activation has either been shown to inhibit [106,107] or to promote [108] TRAIL-induced apoptosis [109].
Among the MAPK superfamily, p38, ERK and JNK, would favour TRAIL-induced apoptosis [110,111], whereas ERK would display either anti-apoptotic [112,113,114,115] or pro-apoptotic functions [116]. These apparent controversies could be due to our poor understanding of the TRAIL signalling pathway, and the molecular mechanisms involved in its regulation could be more complex than anticipated.
4. Therapeutic challenges related to TRAIL
Recombinant TRAIL administration
TRAIL has attracted great expectations in the last years for oncologists. The first hints that this cytokine may hold novel antitumour properties came from the demonstration that recombinant soluble TRAIL injection in xenografted animals induced tumour regression without systemic toxicity [28,36,117,118,119,120,121,122]. TRAIL-induced tumour regression was documented in SCID mice bearing human tumours, such as colon and breast carcinomas [118,121,123], multiple myeloma [124], multiple glioma [125], pancreatic adenocarcinoma [126] or colon carcinoma [127]. Human recombinant TRAIL is now evaluated in phase I clinical trials (Genentech and Amgen). Preliminary results indicate so far that it is well tolerated in patients, but its rapid clearance from the circulation may hamper its antitumoral properties.
However, studies on xenografted animals indicated so far that the administration of recombinant TRAIL rarely induced a complete eradication of established tumours. In addition, the resistance of fresh human tumour cells to TRAIL cytotoxic effects remains poorly known [127,128]. Besides, TRAIL has been demonstrated to induce cell proliferation in some resistant cancer cells in vitro [107,129], as well as to promote tumour cell proliferation and metastasis, in vivo [130,131]. These findings could dampen the potential interest of recombinant TRAIL preparations for cancer therapy.
Other strategies have been considered to improve the efficiency and/or specificity of the TRAIL-death signalling pathway for cancer therapy. In order to avoid TRAIL-inhibitory functions of TRAIL Decoy Receptors, several recombinant TRAIL variants binding exclusively to DR5 have been developed [132]. In vitro, these variants are more efficient than TRAIL itself in inducing the apoptotic machinery in ovarian cancer cell lines. More recently, a chimeric form of TRAIL, activable by matrix metalloproteinases, proteases essentially present in human tumour sites, has been proposed as an alternative molecule with the idea of targeting specifically tumour cells [133]. It remains to be determined, however, whether these TRAIL variants will exhibit sufficient half-life in patients to retain antitumoral selectivity, whether they will be tolerated and whether these preparations will lack TRAIL proliferative properties.
TRAIL gene therapy
Several gene therapy approaches have been considered to deliver TRAIL to the tumour. TRAIL gene transfer into cancer cells can efficiently suppress established tumour growth, without major side effects [42,134,135,136,137,138], even in metastatic prone cancers cells [139]. Using a syngeneic mouse model, TRAIL retroviral-mediated gene transfer in the bone marrow was shown to induce tumour growth regression [140]. Lentiviral-mediated gene transfer can also be efficient ex-vivo, but the efficiency of such strategy could be lower in vivo [141]. The efficacy of TRAIL gene transfer could be enhanced when combined to other antitumour drugs [134,142,143]. For instance, it has been shown recently that combination of TRAIL gene therapy with actinomycin D was an efficient treatment of multiple liver metastasis in a mouse model [144].
Chemosensitization to TRAIL
In vitro, several studies showed that pretreatment of resistant cancer cells with classical chemotherapeutic drugs sensitizes them to TRAIL mediated cell death [145,146,147,148]. Chemosensitization to TRAIL-induced cell death was also observed in immunodeficient mice, resulting in the regression of xenografted colon carcinomas, osteocarcinomas or breast carcinomas [48,117,118,123,147,149].
The mechanisms underlying this chemosensitization depend on both the drug and the cell type [150]. Chemotherapeutic agents can upregulate agonistic receptor expression [48,151,152,153], facilitate the activation of the mitochondrial cell death pathway [120], or activate the caspase signalling cascade [154]. However, if chemotherapy enhances TRAIL cytotoxic effect in cancer cells, the hepatotoxicity of such combinations has to be carefully evaluated, especially as far as cisplatin is concerned [155,156,157].
Radiation sensitization to TRAIL
Ionizing radiations sensitize breast cancer, leukemic, colon carcinoma, melanoma and glioma cells to TRAIL-induced apoptosis [158,159,160,161,162]. This synergistic effect is thought to occur through a p53-dependent mediated DR5 upregulation, induction of Bax and Bak, inhibition of Bcl-2, and caspase activation [163]. Alone, endogenous TRAIL could play a role in radiation-induced cell death, since some tissues arising from the TRAIL-R knockout mice are more resistant to apoptosis following ionizing radiation, compared to wild type littermate [164].
The increasing interest in combining TRAIL with radio- or chemotherapy, which are the main treatments currently used at the clinic to cure cancer, lies on the observation that cross-resistant tumours are most of the time efficiently killed by the combination of both drugs. This strategy would therefore be useful when current treatments fail, for instance in pancreatic or lung cancers.
Agonistic TRAIL receptor antibody use
The use of agonistic antibodies targeting DR4 or DR5 could be more effective than recombinant TRAIL to treat cancers, since these compounds target selectively the agonistic receptors [165,166] and are thus able to trigger apoptosis even in cells which harbour at their cell surface the antagonistic receptors DcR1 or DcR2 [72] (Figure 1). These antibodies have been shown to induce apoptosis in human cancer cell lines and primary cells in vitro [167], as well as in a variety of human tumours grown in immunodeficient mice [168,169]. Combination of these antibodies with chemotherapy [165,168,169,170] or radiotherapy [171] enhances their anti-tumoral efficiency.
In mouse orthotopic models their administration has been shown to induce primary and metastatic tumour regression through ADCC (Antibody-Dependent Cellular Cytotoxicity) and CDC (Complement-Dependent Cytotoxicity) inducing thus a long-term protection against tumour recurrence [172,173]. Administration of murine agonistic antibodies against DR4 or DR5 induces death of tumour cell in vitro and in vivo, in xenograft models [174,175].
Some of these humanized agonistic antibodies to DR4 and DR5 are undergoing phase II (HGS-ETR1 or Mapatumumab, human genome sciences-Cambridge antibody technologies) or phase I clinical trials (HGS-ETR2, also named Lexatumumab). Compared to TRAIL, these antibodies exhibit a higher half-life in patient plasma and their administration is as safe as TRAIL [176,177]. Since DR4 and DR5 expression levels are though to be higher in cancer cells than in normal cells [65,178,179], the safe administration of these antibodies could warrant an improved tumour specificity.
Yet their efficacy remains to be demonstrated in patients. A phase II trial testing the anti-DR4 HGS-ETR1 antibody was performed in 40 patients with relapsed or refractory non-Hodgkin lymphoma and gave rise to promising results [180]. To date, 8% of the patients responded to the treatment, with one complete remission, and two partial responses, and a prolonged stabilization in 30% of patients. A phase II trial associating chemotherapy to the anti-DR4 has also been initiated and results are expected by the end of 2007.
Combination of TRAIL with other molecules
Several protein inhibitors have been shown to sensitize resistant cells to TRAIL-mediated apoptosis, such as mTOR inhibitors [181], proteasome inhibitors [182,183], HDAC inhibitors [143,184,185], Hsp90 inhibitors [186,187,188], Akt inhibitors [189], IFN [190,191], DNA methyltransferase inhibitors [41,192,193] and casein kinase 2 inhibitors [194]. Other molecules that sensitize tumour cells to TRAIL include resveratrol [195], tunicamycin [196], peroxysome proliferators-activated receptor agonists [197], betulinic acid, reactive oxygen species [198], telomerase-dependent virotherapy [199], herbal compound such as wogonin [200] or Bcl2 family inhibitory molecules and small Smac mimetics [201,202,203,204,205].
Other approaches consisting in targeting the extrinsic pathway downstream of TRAIL receptors have been proposed. Synthetic triterpenoids are antitumour compounds that have been shown to donwregulate c-FLIP and to sensitize tumour cells to TRAIL-induced cell death [206,207]. More recently, using a chemical library screen novel compounds affecting c-FLIP expression have been found [208]. These compounds, which enhance caspase-8 recruitment and activation within TRAIL DISC, could be useful combined to TRAIL in cancer therapy.
However, all these strategies, which often target a limited number of anti-apoptotic pathways in a given cell type, could be inefficient against the wide panel of TRAIL-resistant cells, especially considering the impressive variety of the molecular mechanisms allowing these cells to escape from TRAIL-induced cell death.
TRAIL and immunotherapy
Immunotherapy protocols are based on the combination of a tumour-cell apoptosis inducer with immuno-stimulators. TRAIL, as a cancer cell death inducer, may be responsible for the release of tumour antigens, up-taken by APC (Antigen Presenting Cells) and presented to CTL (Cytotoxic T Lymphocytes). Promoting APC stimulation or T-cell co-stimulation could in principle enhance the anti-tumour response, and provoke the eradication of TRAIL-resistant tumours.
Proof-of-principal for such an approach has been obtained recently, in vivo, using cells expressing TRAIL ectopically in syngeneic mice challenged for tumour regression in combination with cyclophosphamide, a compound shown to break immune tolerance [209]. Since tumour cells highly express DR5, the use of antibody against DR5 could induce a greater T cell immunity, by recruiting the Fc-receptor innate immune cells [172]. More recently, trimAb therapy combining antibodies against DR5, CD40 and CD137 has been shown to induce the rejection of established tumours in 80% of mice, and a complete cure in 60% of mice with spontaneous and distant metastases, after surgical resection of the primary orthotopic tumours [210]. This strategy allowed the rejection of tumours resistant to anti-DR5 antibody, stimulating the different steps of the immune response to give rise to an efficient tumour-specific induction of CTL, and a long-term protection against tumour recurrence.
5. Expert opinion and concluding remarks
Recombinant TRAIL or TRAIL derivatives are remarkable antitumour compounds that trigger cell death in a wide variety of cancer cells without major side effect in vivo. Yet, the ability of cancer cells to escape from TRAIL-induced cytotoxic activity is an important issue that needs to be considered in order to design novel therapeutic approaches aiming at targeting this deadly pathway. Combining TRAIL with current or recently developed anticancer drugs should overcome this resistance and provide greater therapeutic benefit than the use of these compounds alone. Given the complexity of the TRAIL system, compared to other TNF related death domain containing receptors, extensive investigations are still required to understand TRAIL regulatory pathways, since to date no simple predictive markers are available to determine the optimal combination that should be used to treat patients. Whether the use of TRAIL as an immunostimulatory cytokine in specific protocols would overcome TRAIL resistance and favour the complete eradication of tumours has also to be demonstrated. Challenging these strategies against a wider range of primary tumour cells both in syngeneic animal mouse models and in clinical trials should help to figure out whether TRAIL will hold its promises for cancer therapy.
Acknowledgments
This study was supported by grants of the Ligue Nationale contre le Cancer, the Conseil Régional de Bourgogne, the INCa (Institut National du Cancer), le Cancéropôle Grand-Est, the ANR (Agence Nationale de la Recherche), and the INSERM. We would like to thank Virginie Granci and Guillaume Jacquemin for their suggestions and helpful comments. We apologise for not citing further interesting and important papers due to space limitations.
References
- 1.PITTI RM, MARSTERS SA, RUPPERT S, et al. Induction of apoptosis by Apo-2 ligand, a new member of the tumor necrosis factor cytokine family. J Biol Chem. 1996;271(22):12687–12690. doi: 10.1074/jbc.271.22.12687. [DOI] [PubMed] [Google Scholar]
- 2.WILEY SR, SCHOOLEY K, SMOLAK PJ, et al. Identification and characterization of a new member of the TNF family that induces apoptosis. Immunity. 1995;3(6):673–682. doi: 10.1016/1074-7613(95)90057-8. [DOI] [PubMed] [Google Scholar]
- 3.PAN G, O’ROURKE K, CHINNAIYAN AM, et al. The receptor for the cytotoxic ligand TRAIL. Science. 1997;276(5309):111–113. doi: 10.1126/science.276.5309.111. [DOI] [PubMed] [Google Scholar]
- 4.CHAUDHARY PM, EBY M, JASMIN A, et al. Death receptor 5, a new member of the TNFR family, and DR4 induce FADD-dependent apoptosis and activate the NF-kappaB pathway. Immunity. 1997;7(6):821–830. doi: 10.1016/s1074-7613(00)80400-8. [DOI] [PubMed] [Google Scholar]
- 5.WALCZAK H, DEGLI-ESPOSTI MA, JOHNSON RS, et al. TRAIL-R2: a novel apoptosis-mediating receptor for TRAIL. Embo J. 1997;16(17):5386–5397. doi: 10.1093/emboj/16.17.5386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.DEGLI-ESPOSTI MA, SMOLAK PJ, WALCZAK H, et al. Cloning and characterization of TRAIL-R3, a novel member of the emerging TRAIL receptor family. J Exp Med. 1997;186(7):1165–1170. doi: 10.1084/jem.186.7.1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.PAN G, NI J, WEI YF, et al. An antagonist decoy receptor and a death domain-containing receptor for TRAIL. Science. 1997;277(5327):815–818. doi: 10.1126/science.277.5327.815. [DOI] [PubMed] [Google Scholar]
- 8.DEGLI-ESPOSTI MA, DOUGALL WC, SMOLAK PJ, et al. The novel receptor TRAIL-R4 induces NF-kappaB and protects against TRAIL-mediated apoptosis, yet retains an incomplete death domain. Immunity. 1997;7(6):813–820. doi: 10.1016/s1074-7613(00)80399-4. [DOI] [PubMed] [Google Scholar]
- 9*.VARFOLOMEEV E, MAECKER H, SHARP D, et al. Molecular determinants of kinase pathway activation by apo2 ligand/tumor necrosis factor related apoptosis-inducing ligand. J Biol Chem. 2005 doi: 10.1074/jbc.M509560200. First paper demonstrating the existence of a secondary complex regulating kinase signalling pathways by TRAIL. [DOI] [PubMed] [Google Scholar]
- 10.MICHEAU O, TSCHOPP J. Induction of TNF receptor I-mediated apoptosis via two sequential signaling complexes. Cell. 2003;114(2):181–190. doi: 10.1016/s0092-8674(03)00521-x. [DOI] [PubMed] [Google Scholar]
- 11.LIABAKK NB, ESPEVIK T. Monoclonal antibodies against TRAIL. Vitam Horm. 2004;67:65–79. doi: 10.1016/S0083-6729(04)67005-1. [DOI] [PubMed] [Google Scholar]
- 12.KAYAGAKI N, YAMAGUCHI N, NAKAYAMA M, et al. Type I interferons (IFNs) regulate tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) expression on human T cells: A novel mechanism for the antitumor effects of type I IFNs. J Exp Med. 1999;189(9):1451–1460. doi: 10.1084/jem.189.9.1451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.GRIFFITH TS, WILEY SR, KUBIN MZ, et al. Monocyte-mediated tumoricidal activity via the tumor necrosis factor-related cytokine, TRAIL. J Exp Med. 1999;189(8):1343–1354. doi: 10.1084/jem.189.8.1343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.KOGA Y, MATSUZAKI A, SUMINOE A, HATTORI H, HARA T. Neutrophil-derived TNF-related apoptosis-inducing ligand (TRAIL): a novel mechanism of antitumor effect by neutrophils. Cancer Res. 2004;64(3):1037–1043. doi: 10.1158/0008-5472.can-03-1808. [DOI] [PubMed] [Google Scholar]
- 15.KAYAGAKI N, YAMAGUCHI N, NAKAYAMA M, et al. Involvement of TNF-related apoptosis-inducing ligand in human CD4+ T cell-mediated cytotoxicity. J Immunol. 1999;162(5):2639–2647. [PubMed] [Google Scholar]
- 16.FANGER NA, MALISZEWSKI CR, SCHOOLEY K, GRIFFITH TS. Human dendritic cells mediate cellular apoptosis via tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) J Exp Med. 1999;190(8):1155–1164. doi: 10.1084/jem.190.8.1155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.LIU S, YU Y, ZHANG M, WANG W, CAO X. The involvement of TNF-alpha-related apoptosis-inducing ligand in the enhanced cytotoxicity of IFN-beta-stimulated human dendritic cells to tumor cells. J Immunol. 2001;166(9):5407–5415. doi: 10.4049/jimmunol.166.9.5407. [DOI] [PubMed] [Google Scholar]
- 18.YU Y, LIU S, WANG W, et al. Involvement of tumour necrosis factor-alpha-related apoptosis-inducing ligand in enhanced cytotoxicity of lipopolysaccharide-stimulated dendritic cells to activated T cells. Immunology. 2002;106(3):308–315. doi: 10.1046/j.1365-2567.2002.01431.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.KROLL TM, BOMMIASAMY H, BOISSY RE, et al. 4-Tertiary butyl phenol exposure sensitizes human melanocytes to dendritic cell-mediated killing: relevance to vitiligo. J Invest Dermatol. 2005;124(4):798–806. doi: 10.1111/j.0022-202X.2005.23653.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.LU G, JANJIC BM, JANJIC J, et al. Innate direct anticancer effector function of human immature dendritic cells. II. Role of TNF, lymphotoxin-alpha(1)beta(2), Fas ligand, and TNF-related apoptosis-inducing ligand. J Immunol. 2002;168(4):1831–1839. doi: 10.4049/jimmunol.168.4.1831. [DOI] [PubMed] [Google Scholar]
- 21.SHI J, IKEDA K, FUJII N, et al. Activated human umbilical cord blood dendritic cells kill tumor cells without damaging normal hematological progenitor cells. Cancer Sci. 2005;96(2):127–133. doi: 10.1111/j.1349-7006.2005.00017.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.KAMOHARA H, MATSUYAMA W, SHIMOZATO O, et al. Regulation of tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) and TRAIL receptor expression in human neutrophils. Immunology. 2004;111(2):186–194. doi: 10.1111/j.0019-2805.2003.01794.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.LIABAKK NB, SUNDAN A, TORP S, et al. Development, characterization and use of monoclonal antibodies against sTRAIL: measurement of sTRAIL by ELISA. J Immunol Methods. 2002;259(1–2):119–128. doi: 10.1016/s0022-1759(01)00501-4. [DOI] [PubMed] [Google Scholar]
- 24.CLARKE P, MEINTZER SM, GIBSON S, et al. Reovirus-induced apoptosis is mediated by TRAIL. J Virol. 2000;74(17):8135–8139. doi: 10.1128/jvi.74.17.8135-8139.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.HAN LH, SUN WS, MA CH, et al. Detection of soluble TRAIL in HBV infected patients and its clinical implications. World J Gastroenterol. 2002;8(6):1077–1080. doi: 10.3748/wjg.v8.i6.1077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.WANDINGER KP, LUNEMANN JD, WENGERT O, et al. TNF-related apoptosis inducing ligand (TRAIL) as a potential response marker for interferon-beta treatment in multiple sclerosis. Lancet. 2003;361(9374):2036–2043. doi: 10.1016/S0140-6736(03)13641-0. [DOI] [PubMed] [Google Scholar]
- 27.ROBERTSON NM, ZANGRILLI JG, STEPLEWSKI A, et al. Differential expression of TRAIL and TRAIL receptors in allergic asthmatics following segmental antigen challenge: evidence for a role of TRAIL in eosinophil survival. J Immunol. 2002;169(10):5986–5996. doi: 10.4049/jimmunol.169.10.5986. [DOI] [PubMed] [Google Scholar]
- 28.ASHKENAZI A, PAI RC, FONG S, et al. Safety and antitumor activity of recombinant soluble Apo2 ligand. J Clin Invest. 1999;104(2):155–162. doi: 10.1172/JCI6926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29**.CRETNEY E, SHANKER A, YAGITA H, SMYTH MJ, SAYERS TJ. TNF-related apoptosis-inducing ligand as a therapeutic agent in autoimmunity and cancer. Immunology and cell biology. 2006;84:1–98. doi: 10.1111/j.1440-1711.2005.01413.x. thorough review on the current understanding of the TRAIL system. [DOI] [PubMed] [Google Scholar]
- 30.CRETNEY E, TAKEDA K, YAGITA H, et al. Increased susceptibility to tumor initiation and metastasis in TNF-related apoptosis-inducing ligand-deficient mice. J Immunol. 2002;168(3):1356–1361. doi: 10.4049/jimmunol.168.3.1356. [DOI] [PubMed] [Google Scholar]
- 31.SMYTH MJ, TAKEDA K, HAYAKAWA Y, et al. Nature’s TRAIL-On a Path to Cancer Immunotherapy. Immunity. 2003;18(1):1–6. doi: 10.1016/s1074-7613(02)00502-2. [DOI] [PubMed] [Google Scholar]
- 32**.ZERAFA N, WESTWOOD JA, CRETNEY E, et al. Cutting edge: TRAIL deficiency accelerates hematological malignancies. J Immunol. 2005;175:9–5590. doi: 10.4049/jimmunol.175.9.5586. This excellent paper highlights the important role of TRAIL for the control of carcinogenesis. [DOI] [PubMed] [Google Scholar]
- 33.SEDGER LM, GLACCUM MB, SCHUH JC, et al. Characterization of the in vivo function of TNF-alpha-related apoptosis-inducing ligand, TRAIL/Apo2L, using TRAIL/Apo2L gene-deficient mice. Eur J Immunol. 2002;32(8):2246–2254. doi: 10.1002/1521-4141(200208)32:8<2246::AID-IMMU2246>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- 34.TAKEDA K, SMYTH MJ, CRETNEY E, et al. Critical role for tumor necrosis factor-related apoptosis-inducing ligand in immune surveillance against tumor development. J Exp Med. 2002;195(2):161–169. doi: 10.1084/jem.20011171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.HAYAKAWA Y, TAKEDA K, YAGITA H, et al. Critical contribution of IFN-gamma and NK cells, but not perforin-mediated cytotoxicity, to anti-metastatic effect of alpha-galactosylceramide. Eur J Immunol. 2001;31(6):1720–1727. [PubMed] [Google Scholar]
- 36.SEKI N, HAYAKAWA Y, BROOKS AD, et al. Tumor necrosis factor-related apoptosis-inducing ligand-mediated apoptosis is an important endogenous mechanism for resistance to liver metastases in murine renal cancer. Cancer Res. 2003;63(1):207–213. [PubMed] [Google Scholar]
- 37.SMYTH MJ, CRETNEY E, TAKEDA K, et al. Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) contributes to interferon gamma-dependent natural killer cell protection from tumor metastasis. J Exp Med. 2001;193(6):661–670. doi: 10.1084/jem.193.6.661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.DIEHL GE, YUE HH, HSIEH K, et al. TRAIL-R as a negative regulator of innate immune cell responses. Immunity. 2004;21(6):877–889. doi: 10.1016/j.immuni.2004.11.008. [DOI] [PubMed] [Google Scholar]
- 39.NESTEROV A, NIKRAD M, JOHNSON T, KRAFT AS. Oncogenic Ras sensitizes normal human cells to tumor necrosis factor-alpha-related apoptosis-inducing ligand-induced apoptosis. Cancer research. 2004;64(11):3922–3927. doi: 10.1158/0008-5472.CAN-03-2219. [DOI] [PubMed] [Google Scholar]
- 40.WANG Y, QUON KC, KNEE DA, NESTEROV A, KRAFT AS. RAS, MYC, and sensitivity to tumor necrosis factor-alpha-related apoptosis-inducing ligand-induced apoptosis. Cancer research. 2005;65(4):1615–1616. doi: 10.1158/0008-5472.CAN-04-2757. author reply 1616–1617. [DOI] [PubMed] [Google Scholar]
- 41.GROTZER MA, EGGERT A, ZUZAK TJ, et al. Resistance to TRAIL-induced apoptosis in primitive neuroectodermal brain tumor cells correlates with a loss of caspase-8 expression. Oncogene. 2000;19(40):4604–4610. doi: 10.1038/sj.onc.1203816. [DOI] [PubMed] [Google Scholar]
- 42.KAGAWA S, HE C, GU J, et al. Antitumor activity and bystander effects of the tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) gene. Cancer Res. 2001;61(8):3330–3338. [PubMed] [Google Scholar]
- 43.WAJANT H, PFIZENMAIER K, SCHEURICH P. TNF-related apoptosis inducing ligand (TRAIL) and its receptors in tumor surveillance and cancer therapy. Apoptosis. 2002;7(5):449–459. doi: 10.1023/a:1020039225764. [DOI] [PubMed] [Google Scholar]
- 44.JIN Z, MCDONALD ER, 3RD, DICKER DT, EL-DEIRY WS. Deficient tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) death receptor transport to the cell surface in human colon cancer cells selected for resistance to TRAIL-induced apoptosis. J Biol Chem. 2004;279(34):35829–35839. doi: 10.1074/jbc.M405538200. [DOI] [PubMed] [Google Scholar]
- 45.HORAK P, PILS D, HALLER G, et al. Contribution of epigenetic silencing of tumor necrosis factor-related apoptosis inducing ligand receptor 1 (DR4) to TRAIL resistance and ovarian cancer. Mol Cancer Res. 2005;3(6):335–343. doi: 10.1158/1541-7786.MCR-04-0136. [DOI] [PubMed] [Google Scholar]
- 46.LEE SH, SHIN MS, KIM HS, et al. Somatic mutations of TRAIL-receptor 1 and TRAIL-receptor 2 genes in non-Hodgkin’s lymphoma. Oncogene. 2001;20(3):399–403. doi: 10.1038/sj.onc.1204103. [DOI] [PubMed] [Google Scholar]
- 47.NGUYEN T, ZHANG XD, HERSEY P. Relative resistance of fresh isolates of melanoma to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis. Clin Cancer Res. 2001;7(3 Suppl):966s–973s. [PubMed] [Google Scholar]
- 48.BOURALEXIS S, FINDLAY DM, ATKINS GJ, et al. Progressive resistance of BTK-143 osteosarcoma cells to Apo2L/TRAIL-induced apoptosis is mediated by acquisition of DcR2/TRAIL-R4 expression: resensitisation with chemotherapy. Br J Cancer. 2003;89(1):206–214. doi: 10.1038/sj.bjc.6601021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.MUHLENBECK F, SCHNEIDER P, BODMER JL, et al. The tumor necrosis factor-related apoptosis-inducing ligand receptors TRAIL-R1 and TRAIL-R2 have distinct cross-linking requirements for initiation of apoptosis and are non-redundant in JNK activation. J Biol Chem. 2000;275(41):32208–32213. doi: 10.1074/jbc.M000482200. [DOI] [PubMed] [Google Scholar]
- 50.FISHER MJ, VIRMANI AK, WU L, et al. Nucleotide substitution in the ectodomain of trail receptor DR4 is associated with lung cancer and head and neck cancer. Clin Cancer Res. 2001;7(6):1688–1697. [PubMed] [Google Scholar]
- 51.KIM K, FISHER MJ, XU SQ, EL-DEIRY WS. Molecular determinants of response to TRAIL in killing of normal and cancer cells. Clin Cancer Res. 2000;6(2):335–346. [PubMed] [Google Scholar]
- 52.PARK WS, LEE JH, SHIN MS, et al. Inactivating mutations of KILLER/DR5 gene in gastric cancers. Gastroenterology. 2001;121(5):1219–1225. doi: 10.1053/gast.2001.28663. [DOI] [PubMed] [Google Scholar]
- 53.MCDONALD ER, 3RD, CHUI PC, MARTELLI PF, DICKER DT, EL-DEIRY WS. Death domain mutagenesis of KILLER/DR5 reveals residues critical for apoptotic signaling. J Biol Chem. 2001;276(18):14939–14945. doi: 10.1074/jbc.M100399200. [DOI] [PubMed] [Google Scholar]
- 54.SPIERINGS DC, DE VRIES EG, VELLENGA E, et al. Tissue distribution of the death ligand TRAIL and its receptors. J Histochem Cytochem. 2004;52(6):821–831. doi: 10.1369/jhc.3A6112.2004. [DOI] [PubMed] [Google Scholar]
- 55.VAN NOESEL MM, VAN BEZOUW S, SALOMONS GS, et al. Tumor-specific down-regulation of the tumor necrosis factor-related apoptosis-inducing ligand decoy receptors DcR1 and DcR2 is associated with dense promoter hypermethylation. Cancer Res. 2002;62(7):2157–2161. [PubMed] [Google Scholar]
- 56.SHIVAPURKAR N, TOYOOKA S, TOYOOKA KO, et al. Aberrant methylation of trail decoy receptor genes is frequent in multiple tumor types. Int J Cancer. 2004;109(5):786–792. doi: 10.1002/ijc.20041. [DOI] [PubMed] [Google Scholar]
- 57.SHEIKH MS, HUANG Y, FERNANDEZ-SALAS EA, et al. The antiapoptotic decoy receptor TRID/TRAIL-R3 is a p53-regulated DNA damage-inducible gene that is overexpressed in primary tumors of the gastrointestinal tract. Oncogene. 1999;18(28):4153–4159. doi: 10.1038/sj.onc.1202763. [DOI] [PubMed] [Google Scholar]
- 58.SANLIOGLU AD, KOKSAL IT, CIFTCIOGLU A, et al. Differential expression of TRAIL and its receptors in benign and malignant prostate tissues. J Urol. 2007;177(1):359–364. doi: 10.1016/j.juro.2006.08.087. [DOI] [PubMed] [Google Scholar]
- 59.AYDIN C, SANLIOGLU AD, KARACAY B, et al. Decoy receptor-2 small interfering RNA (siRNA) strategy employing three different siRNA constructs in combination defeats adenovirus-transferred tumor necrosis factor-related apoptosis-inducing ligand resistance in lung cancer cells. Hum Gene Ther. 2007;18(1):39–50. doi: 10.1089/hum.2006.111. [DOI] [PubMed] [Google Scholar]
- 60.RICCIONI R, PASQUINI L, MARIANI G, et al. TRAIL decoy receptors mediate resistance of acute myeloid leukemia cells to TRAIL. Haematologica. 2005;90(5):612–624. [PubMed] [Google Scholar]
- 61.GRIFFITH TS, CHIN WA, JACKSON GC, LYNCH DH, KUBIN MZ. Intracellular regulation of TRAIL-induced apoptosis in human melanoma cells. J Immunol. 1998;161(6):2833–2840. [PubMed] [Google Scholar]
- 62.HAO C, BEGUINOT F, CONDORELLI G, et al. Induction and intracellular regulation of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) mediated apotosis in human malignant glioma cells. Cancer research. 2001;61(3):1162–1170. [PubMed] [Google Scholar]
- 63.MITSIADES N, POULAKI V, MITSIADES C, TSOKOS M. Ewing’s sarcoma family tumors are sensitive to tumor necrosis factor-related apoptosis-inducing ligand and express death receptor 4 and death receptor 5. Cancer research. 2001;61(6):2704–2712. [PubMed] [Google Scholar]
- 64.ZHANG XD, FRANCO A, MYERS K, et al. Relation of TNF-related apoptosis-inducing ligand (TRAIL) receptor and FLICE-inhibitory protein expression to TRAIL-induced apoptosis of melanoma. Cancer research. 1999;59(11):2747–2753. [PubMed] [Google Scholar]
- 65.DANIELS RA, TURLEY H, KIMBERLEY FC, et al. Expression of TRAIL and TRAIL receptors in normal and malignant tissues. Cell Res. 2005;15(6):430–438. doi: 10.1038/sj.cr.7290311. [DOI] [PubMed] [Google Scholar]
- 66.LINCZ LF, YEH TX, SPENCER A. TRAIL-induced eradication of primary tumour cells from multiple myeloma patient bone marrows is not related to TRAIL receptor expression or prior chemotherapy. Leukemia. 2001;15(10):1650–1657. doi: 10.1038/sj.leu.2402251. [DOI] [PubMed] [Google Scholar]
- 67.ZHANG XD, FRANCO AV, NGUYEN T, GRAY CP, HERSEY P. Differential localization and regulation of death and decoy receptors for TNF-related apoptosis-inducing ligand (TRAIL) in human melanoma cells. J Immunol. 2000;164(8):3961–3970. doi: 10.4049/jimmunol.164.8.3961. [DOI] [PubMed] [Google Scholar]
- 68.MACFARLANE M, AHMAD M, SRINIVASULA SM, et al. Identification and molecular cloning of two novel receptors for the cytotoxic ligand TRAIL. The Journal of biological chemistry. 1997;272(41):25417–25420. doi: 10.1074/jbc.272.41.25417. [DOI] [PubMed] [Google Scholar]
- 69.TRUNEH A, SHARMA S, SILVERMAN C, et al. Temperature-sensitive differential affinity of TRAIL for its receptors. DR5 is the highest affinity receptor. The Journal of biological chemistry. 2000;275(30):23319–23325. doi: 10.1074/jbc.M910438199. [DOI] [PubMed] [Google Scholar]
- 70.LEE HW, LEE SH, LEE HW, et al. Homomeric and heteromeric interactions of the extracellular domains of death receptors and death decoy receptors. Biochem Biophys Res Commun. 2005;330(4):1205–1212. doi: 10.1016/j.bbrc.2005.03.101. [DOI] [PubMed] [Google Scholar]
- 71.SHERIDAN JP, MARSTERS SA, PITTI RM, et al. Control of TRAIL-induced apoptosis by a family of signaling and decoy receptors. Science. 1997;277(5327):818–821. doi: 10.1126/science.277.5327.818. [DOI] [PubMed] [Google Scholar]
- 72.MERINO D, LALAOUI N, MORIZOT A, et al. Differential inhibition of TRAIL-mediated DR5-DISC formation by decoy receptors 1 and 2. Mol Cell Biol. 2006;26(19):7046–7055. doi: 10.1128/MCB.00520-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.MARSTERS SA, SHERIDAN JP, PITTI RM, et al. A novel receptor for Apo2L/TRAIL contains a truncated death domain. Curr Biol. 1997;7(12):1003–1006. doi: 10.1016/s0960-9822(06)00422-2. [DOI] [PubMed] [Google Scholar]
- 74.BERNARD D, QUATANNENS B, VANDENBUNDER B, ABBADIE C. Rel/NF-kappaB transcription factors protect against tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL)-induced apoptosis by up-regulating the TRAIL decoy receptor DcR1. J Biol Chem. 2001;276(29):27322–27328. doi: 10.1074/jbc.M011183200. [DOI] [PubMed] [Google Scholar]
- 75.ZHANG XD, NGUYEN T, THOMAS WD, SANDERS JE, HERSEY P. Mechanisms of resistance of normal cells to TRAIL induced apoptosis vary between different cell types. 2000. pp. 193–199. [DOI] [PubMed] [Google Scholar]
- 76.SANLIOGLU AD, DIRICE E, AYDIN C, et al. Surface TRAIL decoy receptor-4 expression is correlated with TRAIL resistance in MCF7 breast cancer cells. BMC Cancer. 2005;5(1):54. doi: 10.1186/1471-2407-5-54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.AYDIN C, SANLIOGLU AD, KARACAY B, et al. Decoy Receptor-2 Small Interfering RNA (siRNA) Strategy Employing Three Different siRNA Constructs in Combination Defeats Adenovirus-Transferred Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand Resistance in Lung Cancer Cells. Hum Gene Ther. 2006 doi: 10.1089/hum.2006.111. [DOI] [PubMed] [Google Scholar]
- 78.CLANCY L, MRUK K, ARCHER K, et al. Preligand assembly domain-mediated ligand-independent association between TRAIL receptor 4 (TR4) and TR2 regulates TRAIL-induced apoptosis. Proc Natl Acad Sci U S A. 2005 doi: 10.1073/pnas.0507329102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.FALSCHLEHNER C, EMMERICH CH, GERLACH B, WALCZAK H. TRAIL signalling: Decisions between life and death. Int J Biochem Cell Biol. 2007 doi: 10.1016/j.biocel.2007.02.007. [DOI] [PubMed] [Google Scholar]
- 80.CHENG J, HYLANDER BL, BAER MR, CHEN X, REPASKY EA. Multiple mechanisms underlie resistance of leukemia cells to Apo2 Ligand/TRAIL. Mol Cancer Ther. 2006;5(7):1844–1853. doi: 10.1158/1535-7163.MCT-06-0050. [DOI] [PubMed] [Google Scholar]
- 81.FULDA S, KUFER MU, MEYER E, et al. Sensitization for death receptor- or drug-induced apoptosis by re-expression of caspase-8 through demethylation or gene transfer. Oncogene. 2001;20(41):5865–5877. doi: 10.1038/sj.onc.1204750. [DOI] [PubMed] [Google Scholar]
- 82.HOPKINS-DONALDSON S, ZIEGLER A, KURTZ S, et al. Silencing of death receptor and caspase-8 expression in small cell lung carcinoma cell lines and tumors by DNA methylation. Cell Death Differ. 2003;10(3):356–364. doi: 10.1038/sj.cdd.4401157. [DOI] [PubMed] [Google Scholar]
- 83.MCDONALD ER, 3RD, EL-DEIRY WS. Suppression of caspase-8- and -10-associated RING proteins results in sensitization to death ligands and inhibition of tumor cell growth. Proc Natl Acad Sci U S A. 2004;101(16):6170–6175. doi: 10.1073/pnas.0307459101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.LIU X, YUE P, SCHONTHAL AH, KHURI FR, SUN SY. Cellular FLICE-inhibitory protein down-regulation contributes to celecoxib-induced apoptosis in human lung cancer cells. Cancer Res. 2006;66(23):11115–11119. doi: 10.1158/0008-5472.CAN-06-2471. [DOI] [PubMed] [Google Scholar]
- 85.ROTH W, REED JC. FLIP protein and TRAIL-induced apoptosis. Vitam Horm. 2004;67:189–206. doi: 10.1016/S0083-6729(04)67011-7. [DOI] [PubMed] [Google Scholar]
- 86.FULDA S, MEYER E, DEBATIN KM. Inhibition of TRAIL-induced apoptosis by Bcl-2 overexpression. Oncogene. 2002;21(15):2283–2294. doi: 10.1038/sj.onc.1205258. [DOI] [PubMed] [Google Scholar]
- 87.NG CP, ZISMAN A, BONAVIDA B. Synergy is achieved by complementation with Apo2L/TRAIL and actinomycin D in Apo2L/TRAIL-mediated apoptosis of prostate cancer cells: role of XIAP in resistance. Prostate. 2002;53(4):286–299. doi: 10.1002/pros.10155. [DOI] [PubMed] [Google Scholar]
- 88.NG CP, BONAVIDA B. X-linked inhibitor of apoptosis (XIAP) blocks Apo2 ligand/tumor necrosis factor-related apoptosis-inducing ligand-mediated apoptosis of prostate cancer cells in the presence of mitochondrial activation: sensitization by overexpression of second mitochondria-derived activator of caspase/direct IAP-binding protein with low pl (Smac/DIABLO) Mol Cancer Ther. 2002;1(12):1051–1058. [PubMed] [Google Scholar]
- 89.SECCHIERO P, GONELLI A, CARNEVALE E, et al. TRAIL promotes the survival and proliferation of primary human vascular endothelial cells by activating the Akt and ERK pathways. Circulation. 2003;107(17):2250–2256. doi: 10.1161/01.CIR.0000062702.60708.C4. [DOI] [PubMed] [Google Scholar]
- 90.MILANI D, ZAULI G, RIMONDI E, et al. Tumour necrosis factor-related apoptosis-inducing ligand sequentially activates pro-survival and pro-apoptotic pathways in SK-N-MC neuronal cells. Journal of neurochemistry. 2003;86(1):126–135. doi: 10.1046/j.1471-4159.2003.01805.x. [DOI] [PubMed] [Google Scholar]
- 91.MOREL J, AUDO R, HAHNE M, COMBE B. Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) induces rheumatoid arthritis synovial fibroblast proliferation through mitogen-activated protein kinases and phosphatidylinositol 3-kinase/Akt. J Biol Chem. 2005;280(16):15709–15718. doi: 10.1074/jbc.M414469200. [DOI] [PubMed] [Google Scholar]
- 92.CHEN X, THAKKAR H, TYAN F, et al. Constitutively active Akt is an important regulator of TRAIL sensitivity in prostate cancer. Oncogene. 2001;20(42):6073–6083. doi: 10.1038/sj.onc.1204736. [DOI] [PubMed] [Google Scholar]
- 93.BORTUL R, TAZZARI PL, CAPPELLINI A, et al. Constitutively active Akt1 protects HL60 leukemia cells from TRAIL-induced apoptosis through a mechanism involving NF-kappaB activation and cFLIP(L) up-regulation. Leukemia. 2003;17(2):379–389. doi: 10.1038/sj.leu.2402793. [DOI] [PubMed] [Google Scholar]
- 94.KOBAYASHI S, WERNEBURG NW, BRONK SF, KAUFMANN SH, GORES GJ. Interleukin-6 contributes to Mcl-1 up-regulation and TRAIL resistance via an Akt-signaling pathway in cholangiocarcinoma cells. Gastroenterology. 2005;128(7):2054–2065. doi: 10.1053/j.gastro.2005.03.010. [DOI] [PubMed] [Google Scholar]
- 95.KIM KM, LEE YJ. Amiloride augments TRAIL-induced apoptotic death by inhibiting phosphorylation of kinases and phosphatases associated with the P13K-Akt pathway. Oncogene. 2005;24(3):355–366. doi: 10.1038/sj.onc.1208213. [DOI] [PubMed] [Google Scholar]
- 96.KIM EC, YUN BS, RYOO IJ, et al. Complestatin prevents apoptotic cell death: inhibition of a mitochondrial caspase pathway through AKT/PKB activation. Biochem Biophys Res Commun. 2004;313(1):193–204. doi: 10.1016/j.bbrc.2003.11.104. [DOI] [PubMed] [Google Scholar]
- 97.KIM YH, LEE YJ. TRAIL apoptosis is enhanced by quercetin through Akt dephosphorylation. J Cell Biochem. 2006;100(4):998–1009. doi: 10.1002/jcb.21098. [DOI] [PubMed] [Google Scholar]
- 98.SZEGEZDI E, CAHILL S, MEYER M, O’DWYER M, SAMALI A. TRAIL sensitisation by arsenic trioxide is caspase-8 dependent and involves modulation of death receptor components and Akt. Br J Cancer. 2006;94(3):398–406. doi: 10.1038/sj.bjc.6602954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.SHRADER M, PINO MS, LASHINGER L, et al. Gefitinib Reverses TRAIL Resistance in Human Bladder Cancer Cell Lines via Inhibition of AKT-Mediated X-Linked Inhibitor of Apoptosis Protein Expression. Cancer Res. 2007;67(4):1430–1435. doi: 10.1158/0008-5472.CAN-06-1224. [DOI] [PubMed] [Google Scholar]
- 100.MARTELLI AM, TAZZARI PL, TABELLINI G, et al. A new selective AKT pharmacological inhibitor reduces resistance to chemotherapeutic drugs, TRAIL, all-trans-retinoic acid, and ionizing radiation of human leukemia cells. Leukemia. 2003;17(9):1794–1805. doi: 10.1038/sj.leu.2403044. [DOI] [PubMed] [Google Scholar]
- 101.PUDUVALLI VK, SAMPATH D, BRUNER JM, et al. TRAIL-induced apoptosis in gliomas is enhanced by Akt-inhibition and is independent of JNK activation. Apoptosis. 2005;10(1):233–243. doi: 10.1007/s10495-005-6078-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.RYCHAHOU PG, MURILLO CA, EVERS BM. Targeted RNA interference of PI3K pathway components sensitizes colon cancer cells to TNF-related apoptosis-inducing ligand (TRAIL) Surgery. 2005;138(2):391–397. doi: 10.1016/j.surg.2005.05.012. [DOI] [PubMed] [Google Scholar]
- 103.OKA N, TANIMOTO S, TAUE R, et al. Role of phosphatidylinositol-3 kinase/Akt pathway in bladder cancer cell apoptosis induced by tumor necrosis factor-related apoptosis-inducing ligand. Cancer Sci. 2006;97(10):1093–1098. doi: 10.1111/j.1349-7006.2006.00294.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.JIN Z, EL-DEIRY WS. Distinct signaling pathways in TRAIL- versus tumor necrosis factor-induced apoptosis. Mol Cell Biol. 2006;26(21):8136–8148. doi: 10.1128/MCB.00257-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.MARTINON F, HOLLER N, RICHARD C, TSCHOPP J. Activation of a pro-apoptotic amplification loop through inhibition of NF-kappaB-dependent survival signals by caspase-mediated inactivation of RIP. FEBS letters. 2000;468(2–3):134–136. doi: 10.1016/s0014-5793(00)01212-6. [DOI] [PubMed] [Google Scholar]
- 106.KIM YS, SCHWABE RF, QIAN T, LEMASTERS JJ, BRENNER DA. TRAIL-mediated apoptosis requires NF-kappaB inhibition and the mitochondrial permeability transition in human hepatoma cells. Hepatology (Baltimore, Md) 2002;36(6):1498–1508. doi: 10.1053/jhep.2002.36942. [DOI] [PubMed] [Google Scholar]
- 107.EHRHARDT H, FULDA S, SCHMID I, et al. TRAIL induced survival and proliferation in cancer cells resistant towards TRAIL-induced apoptosis mediated by NF-kappaB. Oncogene. 2003;22(25):3842–3852. doi: 10.1038/sj.onc.1206520. [DOI] [PubMed] [Google Scholar]
- 108.SHETTY S, GLADDEN JB, HENSON ES, et al. Tumor necrosis factor-related apoptosis inducing ligand (TRAIL) up-regulates death receptor 5 (DR5) mediated by NFkappaB activation in epithelial derived cell lines. Apoptosis. 2002;7(5):413–420. doi: 10.1023/a:1020031023947. [DOI] [PubMed] [Google Scholar]
- 109.WAJANT H. TRAIL and NFkappaB signaling--a complex relationship. Vitam Horm. 2004;67:101–132. doi: 10.1016/S0083-6729(04)67007-5. [DOI] [PubMed] [Google Scholar]
- 110.JUREWICZ A, MATYSIAK M, ANDRZEJAK S, SELMAJ K. TRAIL-induced death of human adult oligodendrocytes is mediated by JNK pathway. Glia. 2006;53(2):158–166. doi: 10.1002/glia.20249. [DOI] [PubMed] [Google Scholar]
- 111.CORAZZA N, JAKOB S, SCHAER C, et al. TRAIL receptor-mediated JNK activation and Bim phosphorylation critically regulate Fas-mediated liver damage and lethality. The Journal of clinical investigation. 2006;116(9):2493–2499. doi: 10.1172/JCI27726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.TRAN SE, HOLMSTROM TH, AHONEN M, KAHARI VM, ERIKSSON JE. MAPK/ERK overrides the apoptotic signaling from Fas, TNF, and TRAIL receptors. The Journal of biological chemistry. 2001;276(19):16484–16490. doi: 10.1074/jbc.M010384200. [DOI] [PubMed] [Google Scholar]
- 113.LEE TJ, LEE JT, PARK JW, KWON TK. Acquired TRAIL resistance in human breast cancer cells are caused by the sustained cFLIP(L) and XIAP protein levels and ERK activation. Biochem Biophys Res Commun. 2006;351(4):1024–1030. doi: 10.1016/j.bbrc.2006.10.163. [DOI] [PubMed] [Google Scholar]
- 114.BANSIL S, HOLTZ CR, COOK SD, ROHOWSKY-KOCHAN C. Serum sAPO-1/Fas levels in multiple sclerosis. Acta Neurol Scand. 1997;95(4):208–210. doi: 10.1111/j.1600-0404.1997.tb00100.x. [DOI] [PubMed] [Google Scholar]
- 115.BORGES J, PANDIELLA A, ESPARIS-OGANDO A. Erk5 nuclear location is independent on dual phosphorylation, and favours resistance to TRAIL-induced apoptosis. Cell Signal. 2007 doi: 10.1016/j.cellsig.2007.01.023. [DOI] [PubMed] [Google Scholar]
- 116.FRESE S, PIRNIA F, MIESCHER D, et al. PG490-mediated sensitization of lung cancer cells to Apo2L/TRAIL-induced apoptosis requires activation of ERK2. Oncogene. 2003;22(35):5427–5435. doi: 10.1038/sj.onc.1206842. [DOI] [PubMed] [Google Scholar]
- 117.KELLEY SK, ASHKENAZI A. Targeting death receptors in cancer with Apo2L/TRAIL. Curr Opin Pharmacol. 2004;4(4):333–339. doi: 10.1016/j.coph.2004.02.006. [DOI] [PubMed] [Google Scholar]
- 118.WALCZAK H, MILLER RE, ARIAIL K, et al. Tumoricidal activity of tumor necrosis factor-related apoptosis-inducing ligand in vivo. Nat Med. 1999;5(2):157–163. doi: 10.1038/5517. [DOI] [PubMed] [Google Scholar]
- 119.YOUNES A, KADIN ME. Emerging applications of the tumor necrosis factor family of ligands and receptors in cancer therapy. J Clin Oncol. 2003;21(18):3526–3534. doi: 10.1200/JCO.2003.09.037. [DOI] [PubMed] [Google Scholar]
- 120.DEBATIN KM, KRAMMER PH. Death receptors in chemotherapy and cancer. Oncogene. 2004;23(16):2950–2966. doi: 10.1038/sj.onc.1207558. [DOI] [PubMed] [Google Scholar]
- 121.KELLEY SK, HARRIS LA, XIE D, et al. Preclinical studies to predict the disposition of Apo2L/tumor necrosis factor-related apoptosis-inducing ligand in humans: characterization of in vivo efficacy, pharmacokinetics, and safety. J Pharmacol Exp Ther. 2001;299(1):31–38. [PubMed] [Google Scholar]
- 122.LAWRENCE D, SHAHROKH Z, MARSTERS S, et al. Differential hepatocyte toxicity of recombinant Apo2L/TRAIL versions. Nat Med. 2001;7(4):383–385. doi: 10.1038/86397. [DOI] [PubMed] [Google Scholar]
- 123*.NAKA T, SUGAMURA K, HYLANDER BL, et al. Effects of tumor necrosis factor-related apoptosis-inducing ligand alone and in combination with chemotherapeutic agents on patients’ colon tumors grown in SCID mice. Cancer research. 2002;62(20):5800–5806. Interesting example of tumour regression in vivo using TRAIL combined to chemotherapy. [PubMed] [Google Scholar]
- 124.MITSIADES CS, TREON SP, MITSIADES N, et al. TRAIL/Apo2L ligand selectively induces apoptosis and overcomes drug resistance in multiple myeloma: therapeutic applications. Blood. 2001;98(3):795–804. doi: 10.1182/blood.v98.3.795. [DOI] [PubMed] [Google Scholar]
- 125.POLLACK IF, ERFF M, ASHKENAZI A. Direct stimulation of apoptotic signaling by soluble Apo2l/tumor necrosis factor-related apoptosis-inducing ligand leads to selective killing of glioma cells. Clin Cancer Res. 2001;7(5):1362–1369. [PubMed] [Google Scholar]
- 126.HYLANDER BL, PITONIAK R, PENETRANTE RB, et al. The anti-tumor effect of Apo2L/TRAIL on patient pancreatic adenocarcinomas grown as xenografts in SCID mice. J Transl Med. 2005;3(1):22. doi: 10.1186/1479-5876-3-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.OIKONOMOU E, KOTHONIDIS K, ZOGRAFOS G, et al. Newly established tumourigenic primary human colon cancer cell lines are sensitive to TRAIL-induced apoptosis in vitro and in vivo. Br J Cancer. 2007 doi: 10.1038/sj.bjc.6603835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.KOSCHNY R, HOLLAND H, SYKORA J, et al. Bortezomib Sensitizes Primary Human Astrocytoma Cells of WHO Grades I to IV for Tumor Necrosis Factor-Related Apoptosis- Inducing Ligand-Induced Apoptosis. Clin Cancer Res. 2007;13(11):3403–3412. doi: 10.1158/1078-0432.CCR-07-0251. [DOI] [PubMed] [Google Scholar]
- 129.BAADER E, TOLOCZKO A, FUCHS U, et al. Tumor necrosis factor-related apoptosis-inducing ligand-mediated proliferation of tumor cells with receptor-proximal apoptosis defects. Cancer research. 2005;65(17):7888–7895. doi: 10.1158/0008-5472.CAN-04-4278. [DOI] [PubMed] [Google Scholar]
- 130**.TRAUZOLD A, SIEGMUND D, SCHNIEWIND B, et al. TRAIL promotes metastasis of human pancreatic ductal adenocarcinoma. Oncogene. 2006;25(56):7434–7439. doi: 10.1038/sj.onc.1209719. Recent paper suggesting limitations for the use of TRAIL in cancer therapy. [DOI] [PubMed] [Google Scholar]
- 131.THAI LE M, LABRINIDIS A, HAY S, et al. Apo2l/Tumor necrosis factor-related apoptosis-inducing ligand prevents breast cancer-induced bone destruction in a mouse model. Cancer Res. 2006;66(10):5363–5370. doi: 10.1158/0008-5472.CAN-05-4386. [DOI] [PubMed] [Google Scholar]
- 132.VAN DER SLOOT AM, TUR V, SZEGEZDI E, et al. Designed tumor necrosis factor-related apoptosis-inducing ligand variants initiating apoptosis exclusively via the DR5 receptor. Proc Natl Acad Sci U S A. 2006;103(23):8634–8639. doi: 10.1073/pnas.0510187103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.SHIN JN, PARK SY, CHA JH, et al. Generation of a novel proform of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) protein that can be reactivated by matrix metalloproteinases. Exp Cell Res. 2006;312(19):3892–3898. doi: 10.1016/j.yexcr.2006.08.015. [DOI] [PubMed] [Google Scholar]
- 134.WENGER T, MATTERN J, HAAS TL, et al. Apoptosis mediated by lentiviral TRAIL transfer involves transduction-dependent and -independent effects. Cancer Gene Ther. 2006 doi: 10.1038/sj.cgt.7701016. [DOI] [PubMed] [Google Scholar]
- 135.KIM KU, SEO SY, HEO KY, et al. Antitumor activity of TRAIL recombinant adenovirus in human malignant glioma cells. J Korean Med Sci. 2005;20(6):1046–1052. doi: 10.3346/jkms.2005.20.6.1046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.YOO J, CHOI S, HWANG KS, et al. Adeno-associated virus-mediated gene transfer of a secreted form of TRAIL inhibits tumor growth and occurrence in an experimental tumor model. J Gene Med. 2006;8(2):163–174. doi: 10.1002/jgm.832. [DOI] [PubMed] [Google Scholar]
- 137.MOHR A, HENDERSON G, DUDUS L, et al. AAV-encoded expression of TRAIL in experimental human colorectal cancer leads to tumor regression. Gene Ther. 2004;11(6):534–543. doi: 10.1038/sj.gt.3302154. [DOI] [PubMed] [Google Scholar]
- 138.SHAH K, TUNG CH, BREAKEFIELD XO, WEISSLEDER R. In vivo imaging of S-TRAIL- mediated tumor regression and apoptosis. Mol Ther. 2005;11(6):926–931. doi: 10.1016/j.ymthe.2005.01.017. [DOI] [PubMed] [Google Scholar]
- 139.SOVA P, REN XW, NI S, et al. A tumor-targeted and conditionally replicating oncolytic adenovirus vector expressing TRAIL for treatment of liver metastases. Mol Ther. 2004;9(4):496–509. doi: 10.1016/j.ymthe.2003.12.008. [DOI] [PubMed] [Google Scholar]
- 140.SONG K, BENHAGA N, ANDERSON RL, KHOSRAVI-FAR R. Transduction of tumor necrosis factor-related apoptosis-inducing ligand into hematopoietic cells leads to inhibition of syngeneic tumor growth in vivo. Cancer Res. 2006;66(12):6304–6311. doi: 10.1158/0008-5472.CAN-05-3501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.WENGER T, MATTERN J, HAAS TL, et al. Apoptosis mediated by lentiviral TRAIL transfer involves transduction-dependent and -independent effects. Cancer Gene Ther. 2007;14(3):316–326. doi: 10.1038/sj.cgt.7701016. [DOI] [PubMed] [Google Scholar]
- 142.MATSUBARA H, MIZUTANI Y, HONGO F, et al. Gene therapy with TRAIL against renal cell carcinoma. Mol Cancer Ther. 2006;5(9):2165–2171. doi: 10.1158/1535-7163.MCT-05-0522. [DOI] [PubMed] [Google Scholar]
- 143.KASMAN L, LU P, VOELKEL-JOHNSON C. The histone deacetylase inhibitors depsipeptide and MS-275, enhance TRAIL gene therapy of LNCaP prostate cancer cells without adverse effects in normal prostate epithelial cells. Cancer Gene Ther. 2006 doi: 10.1038/sj.cgt.7701017. [DOI] [PubMed] [Google Scholar]
- 144.ISHII M, IWAI M, HARADA Y, et al. Soluble TRAIL gene and actinomycin D synergistically suppressed multiple metastasis of TRAIL-resistant colon cancer in the liver. Cancer Lett. 2007;245(1–2):134–143. doi: 10.1016/j.canlet.2005.12.040. [DOI] [PubMed] [Google Scholar]
- 145.KEANE MM, ETTENBERG SA, NAU MM, RUSSELL EK, LIPKOWITZ S. Chemotherapy augments TRAIL-induced apoptosis in breast cell lines. Cancer research. 1999;59(3):734–741. [PubMed] [Google Scholar]
- 146.LACOUR S, MICHEAU O, HAMMANN A, et al. Chemotherapy enhances TNF-related apoptosis-inducing Ligand DISC assembly in HT29 human colon cancer cells. Oncogene. 2003;22(12):1807–1816. doi: 10.1038/sj.onc.1206127. [DOI] [PubMed] [Google Scholar]
- 147.SINGH TR, SHANKAR S, CHEN X, ASIM M, SRIVASTAVA RK. Synergistic interactions of chemotherapeutic drugs and tumor necrosis factor-related apoptosis-inducing ligand/Apo-2 ligand on apoptosis and on regression of breast carcinoma in vivo. Cancer research. 2003;63(17):5390–5400. [PubMed] [Google Scholar]
- 148.BUCHSBAUM DJ, ZHOU T, LOBUGLIO AF. TRAIL receptor-targeted therapy. Future Oncol. 2006;2(4):493–508. doi: 10.2217/14796694.2.4.493. [DOI] [PubMed] [Google Scholar]
- 149.GLINIAK B, LE T. Tumor necrosis factor-related apoptosis-inducing ligand’s antitumor activity in vivo is enhanced by the chemotherapeutic agent CPT-11. Cancer research. 1999;59(24):6153–6158. [PubMed] [Google Scholar]
- 150.MICHEAU O. Cellular FLICE-inhibitory protein: an attractive therapeutic target? Expert Opin Ther Targets. 2003;7(4):559–573. doi: 10.1517/14728222.7.4.559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.ARTS HJ, DE JONG S, HOLLEMA H, et al. Chemotherapy induces death receptor 5 in epithelial ovarian carcinoma. Gynecol Oncol. 2004;92(3):794–800. doi: 10.1016/j.ygyno.2003.11.054. [DOI] [PubMed] [Google Scholar]
- 152.RYU HS, CHANG KH, CHANG SJ, et al. Expression of TRAIL (TNF-related apoptosis-inducing ligand) receptors in cervical cancer. Int J Gynecol Cancer. 2000;10(5):417–424. doi: 10.1046/j.1525-1438.2000.010005417.x. [DOI] [PubMed] [Google Scholar]
- 153.REESINK-PETERS N, HOUGARDY BM, VAN DEN HEUVEL FA, et al. Death receptors and ligands in cervical carcinogenesis: an immunohistochemical study. Gynecol Oncol. 2005;96(3):705–713. doi: 10.1016/j.ygyno.2004.10.046. [DOI] [PubMed] [Google Scholar]
- 154.GANTEN TM, HAAS TL, SYKORA J, et al. Enhanced caspase-8 recruitment to and activation at the DISC is critical for sensitisation of human hepatocellular carcinoma cells to TRAIL-induced apoptosis by chemotherapeutic drugs. Cell Death Differ. 2004;11 (Suppl 1):S86–96. doi: 10.1038/sj.cdd.4401437. [DOI] [PubMed] [Google Scholar]
- 155.GANTEN TM, KOSCHNY R, SYKORA J, et al. Preclinical differentiation between apparently safe and potentially hepatotoxic applications of TRAIL either alone or in combination with chemotherapeutic drugs. Clin Cancer Res. 2006;12(8):2640–2646. doi: 10.1158/1078-0432.CCR-05-2635. [DOI] [PubMed] [Google Scholar]
- 156.MEURETTE O, FONTAINE A, REBILLARD A, et al. Cytotoxicity of TRAIL/anticancer drug combinations in human normal cells. Annals of the New York Academy of Sciences. 2006;1090:209–216. doi: 10.1196/annals.1378.023. [DOI] [PubMed] [Google Scholar]
- 157.KOSCHNY R, WALCZAK H, GANTEN TM. The promise of TRAIL-potential and risks of a novel anticancer therapy. J Mol Med. 2007 doi: 10.1007/s00109-007-0194-1. [DOI] [PubMed] [Google Scholar]
- 158.CHINNAIYAN AM, PRASAD U, SHANKAR S, et al. Combined effect of tumor necrosis factor-related apoptosis-inducing ligand and ionizing radiation in breast cancer therapy. Proc Natl Acad Sci U S A. 2000;97(4):1754–1759. doi: 10.1073/pnas.030545097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.CIUSANI E, CROCI D, GELATI M, et al. In vitro effects of topotecan and ionizing radiation on TRAIL/Apo2L-mediated apoptosis in malignant glioma. J Neurooncol. 2005;71(1):19–25. doi: 10.1007/s11060-004-9180-4. [DOI] [PubMed] [Google Scholar]
- 160.GONG B, ALMASAN A. Apo2 ligand/TNF-related apoptosis-inducing ligand and death receptor 5 mediate the apoptotic signaling induced by ionizing radiation in leukemic cells. Cancer Res. 2000;60(20):5754–5760. [PubMed] [Google Scholar]
- 161.MARINI P, SCHMID A, JENDROSSEK V, et al. Irradiation specifically sensitises solid tumour cell lines to TRAIL mediated apoptosis. BMC Cancer. 2005;5:5. doi: 10.1186/1471-2407-5-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.IVANOV VN, ZHOU H, HEI TK. Sequential Treatment by Ionizing Radiation and Sodium Arsenite Dramatically Accelerates TRAIL-Mediated Apoptosis of Human Melanoma Cells. Cancer research. 2007;67(11):5397–5407. doi: 10.1158/0008-5472.CAN-07-0551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.SHANKAR S, SINGH TR, SRIVASTAVA RK. Ionizing radiation enhances the therapeutic potential of TRAIL in prostate cancer in vitro and in vivo: Intracellular mechanisms. Prostate. 2004;61(1):35–49. doi: 10.1002/pros.20069. [DOI] [PubMed] [Google Scholar]
- 164.FINNBERG N, GRUBER JJ, FEI P, et al. DR5 knockout mice are compromised in radiation-induced apoptosis. Mol Cell Biol. 2005;25(5):2000–2013. doi: 10.1128/MCB.25.5.2000-2013.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.GONG J, YANG D, KOHANIM S, et al. Novel in vivo imaging shows up-regulation of death receptors by paclitaxel and correlates with enhanced antitumor effects of receptor agonist antibodies. Mol Cancer Ther. 2006;5(12):2991–3000. doi: 10.1158/1535-7163.MCT-06-0188. [DOI] [PubMed] [Google Scholar]
- 166.ZENG Y, WU XX, FISCELLA M, et al. Monoclonal antibody to tumor necrosis factor-related apoptosis-inducing ligand receptor 2 (TRAIL-R2) induces apoptosis in primary renal cell carcinoma cells in vitro and inhibits tumor growth in vivo. Int J Oncol. 2006;28(2):421–430. doi: 10.3892/ijo.28.2.421. [DOI] [PubMed] [Google Scholar]
- 167.GEORGAKIS GV, LI Y, HUMPHREYS R, et al. Activity of selective fully human agonistic antibodies to the TRAIL death receptors TRAIL-R1 and TRAIL-R2 in primary and cultured lymphoma cells: induction of apoptosis and enhancement of doxorubicin- and bortezomib-induced cell death. Br J Haematol. 2005;130(4):501–510. doi: 10.1111/j.1365-2141.2005.05656.x. [DOI] [PubMed] [Google Scholar]
- 168.MOTOKI K, MORI E, MATSUMOTO A, et al. Enhanced apoptosis and tumor regression induced by a direct agonist antibody to tumor necrosis factor-related apoptosis-inducing ligand receptor 2. Clin Cancer Res. 2005;11(8):3126–3135. doi: 10.1158/1078-0432.CCR-04-1867. [DOI] [PubMed] [Google Scholar]
- 169.PUKAC L, KANAKARAJ P, HUMPHREYS R, et al. HGS-ETR1, a fully human TRAIL-receptor 1 monoclonal antibody, induces cell death in multiple tumour types in vitro and in vivo. Br J Cancer. 2005;92(8):1430–1441. doi: 10.1038/sj.bjc.6602487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.VOELKEL-JOHNSON C. An antibody against DR4 (TRAIL-R1) in combination with doxorubicin selectively kills malignant but not normal prostate cells. Cancer Biol Ther. 2003;2(3):283–290. doi: 10.4161/cbt.2.3.398. [DOI] [PubMed] [Google Scholar]
- 171.MARINI P, DENZINGER S, SCHILLER D, et al. Combined treatment of colorectal tumours with agonistic TRAIL receptor antibodies HGS-ETR1 and HGS-ETR2 and radiotherapy: enhanced effects in vitro and dose-dependent growth delay in vivo. Oncogene. 2006;25(37):5145–5154. doi: 10.1038/sj.onc.1209516. [DOI] [PubMed] [Google Scholar]
- 172**.TAKEDA K, YAMAGUCHI N, AKIBA H, et al. Induction of Tumor-specific T Cell Immunity by Anti-DR5 Antibody Therapy. J Exp Med. 2004;199(4):437–448. doi: 10.1084/jem.20031457. First paper to demonstrate that agonistic anti-DR5 antibodies induce Tumor-specific T Cell Immunity and tumor regression in a syngeneic mouse model. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.ZHANG L, ZHANG X, BARRISFORD GW, OLUMI AF. Lexatumumab (TRAIL-receptor 2 mAb) induces expression of DR5 and promotes apoptosis in primary and metastatic renal cell carcinoma in a mouse orthotopic model. Cancer Lett. 2006 doi: 10.1016/j.canlet.2006.11.013. [DOI] [PubMed] [Google Scholar]
- 174.CHUNTHARAPAI A, DODGE K, GRIMMER K, et al. Isotype-dependent inhibition of tumor growth in vivo by monoclonal antibodies to death receptor 4. J Immunol. 2001;166(8):4891–4898. doi: 10.4049/jimmunol.166.8.4891. [DOI] [PubMed] [Google Scholar]
- 175.ICHIKAWA K, LIU W, ZHAO L, et al. Tumoricidal activity of a novel anti-human DR5 monoclonal antibody without hepatocyte cytotoxicity. Nat Med. 2001;7(8):954–960. doi: 10.1038/91000. [DOI] [PubMed] [Google Scholar]
- 176.HOTTE SJ, HIRTE HW, CHEN EX, et al. HGS-ETR1, a Fully Human Monoclonal Antibody to the Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand Death Receptor 1 (TRAIL-R1) in Patients with Advanced Solid Cancer: Results of a Phase 1 Trial. Journal of Clinical Oncology, 2005 ASCO Annual Meeting Proceedings. 2005;23(16S) [Google Scholar]
- 177.PACEY S, PLUMMER RE, ATTARD G, et al. Phase I and pharmacokinetic study of HGS-ETR2, a human monoclonal antibody to TRAIL R2, in patients with advanced solid malignancies. Journal of Clinical Oncology, ASCO Annual Meeting Proceedings. 2005;23(16S):3055. [Google Scholar]
- 178.KOORNSTRA JJ, KLEIBEUKER JH, VAN GEELEN CM, et al. Expression of TRAIL (TNF-related apoptosis-inducing ligand) and its receptors in normal colonic mucosa, adenomas, and carcinomas. J Pathol. 2003;200(3):327–335. doi: 10.1002/path.1364. [DOI] [PubMed] [Google Scholar]
- 179.OZAWA F, FRIESS H, KLEEFF J, et al. Effects and expression of TRAIL and its apoptosis-promoting receptors in human pancreatic cancer. Cancer Lett. 2001;163(1):71–81. doi: 10.1016/s0304-3835(00)00660-1. [DOI] [PubMed] [Google Scholar]
- 180.YOUNES A, VOSE J, ZELENETZ AD, et al. Results of a Phase 2 trial of HGS-ETR1 (agonistic human monoclonal antibody to TRAIL receptor 1) in subjects with relapsed/refractory non-Hodgkin’s lymphoma (NHL) (ETR1-HM01). 47th Annual Meeting of the American Society of Hematology; 2005. [Google Scholar]
- 181.PANNER A, PARSA AT, PIEPER RO. Use of APO2L/TRAIL with mTOR inhibitors in the treatment of glioblastoma multiforme. Expert Rev Anticancer Ther. 2006;6(9):1313–1322. doi: 10.1586/14737140.6.9.1313. [DOI] [PubMed] [Google Scholar]
- 182.INOUE T, SHIRAKI K, FUKE H, et al. Proteasome inhibition sensitizes hepatocellular carcinoma cells to TRAIL by suppressing caspase inhibitors and AKT pathway. Anticancer Drugs. 2006;17(3):261–268. doi: 10.1097/00001813-200603000-00004. [DOI] [PubMed] [Google Scholar]
- 183.SAYERS TJ, MURPHY WJ. Combining proteasome inhibition with TNF-related apoptosis-inducing ligand (Apo2L/TRAIL) for cancer therapy. Cancer Immunol Immunother. 2006;55(1):76–84. doi: 10.1007/s00262-005-0676-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.KAUFMANN SH, STEENSMA DP. On the TRAIL of a new therapy for leukemia. Leukemia. 2005;19(12):2195–2202. doi: 10.1038/sj.leu.2403946. [DOI] [PubMed] [Google Scholar]
- 185.ZHANG XD, GILLESPIE SK, BORROW JM, HERSEY P. The histone deacetylase inhibitor suberic bishydroxamate: a potential sensitizer of melanoma to TNF-related apoptosis-inducing ligand (TRAIL) induced apoptosis. Biochem Pharmacol. 2003;66(8):1537–1545. doi: 10.1016/s0006-2952(03)00509-4. [DOI] [PubMed] [Google Scholar]
- 186.VASILEVSKAYA IA, O’DWYER PJ. 17-Allylamino-17-demethoxygeldanamycin overcomes TRAIL resistance in colon cancer cell lines. Biochem Pharmacol. 2005;70(4):580–589. doi: 10.1016/j.bcp.2005.05.018. [DOI] [PubMed] [Google Scholar]
- 187.MA Y, LAKSHMIKANTHAN V, LEWIS RW, KUMAR MV. Sensitization of TRAIL-resistant cells by inhibition of heat shock protein 90 with low-dose geldanamycin. Mol Cancer Ther. 2006;5(1):170–178. doi: 10.1158/1535-7163.MCT-05-0129. [DOI] [PubMed] [Google Scholar]
- 188.WANG X, JU W, RENOUARD J, et al. 17-allylamino-17-demethoxygeldanamycin synergistically potentiates tumor necrosis factor-induced lung cancer cell death by blocking the nuclear factor-kappaB pathway. Cancer Res. 2006;66(2):1089–1095. doi: 10.1158/0008-5472.CAN-05-2698. [DOI] [PubMed] [Google Scholar]
- 189.ALLADINA SJ, SONG JH, DAVIDGE ST, HAO C, EASTON AS. TRAIL-induced apoptosis in human vascular endothelium is regulated by phosphatidylinositol 3- kinase/Akt through the short form of cellular FLIP and Bcl-2. J Vasc Res. 2005;42(4):337–347. doi: 10.1159/000086599. [DOI] [PubMed] [Google Scholar]
- 190.MERCHANT MS, YANG X, MELCHIONDA F, et al. Interferon gamma enhances the effectiveness of tumor necrosis factor-related apoptosis-inducing ligand receptor agonists in a xenograft model of Ewing’s sarcoma. Cancer Res. 2004;64(22):8349–8356. doi: 10.1158/0008-5472.CAN-04-1705. [DOI] [PubMed] [Google Scholar]
- 191.PAPAGEORGIOU A, LASHINGER L, MILLIKAN R, et al. Role of tumor necrosis factor-related apoptosis-inducing ligand in interferon-induced apoptosis in human bladder cancer cells. Cancer Res. 2004;64(24):8973–8979. doi: 10.1158/0008-5472.CAN-04-1909. [DOI] [PubMed] [Google Scholar]
- 192.FULDA S, DEBATIN KM. 5-Aza-2′-deoxycytidine and IFN-gamma cooperate to sensitize for TRAIL-induced apoptosis by upregulating caspase-8. Oncogene. 2006;25(37):5125–5133. doi: 10.1038/sj.onc.1209518. [DOI] [PubMed] [Google Scholar]
- 193.ERAMO A, PALLINI R, LOTTI F, et al. Inhibition of DNA methylation sensitizes glioblastoma for tumor necrosis factor-related apoptosis-inducing ligand-mediated destruction. Cancer Res. 2005;65(24):11469–11477. doi: 10.1158/0008-5472.CAN-05-1724. [DOI] [PubMed] [Google Scholar]
- 194.RAVI R, BEDI A. Sensitization of tumor cells to Apo2 ligand/TRAIL-induced apoptosis by inhibition of casein kinase II. Cancer Res. 2002;62(15):4180–4185. [PubMed] [Google Scholar]
- 195.FULDA S, DEBATIN KM. Sensitization for anticancer drug-induced apoptosis by the chemopreventive agent resveratrol. Oncogene. 2004;23(40):6702–6711. doi: 10.1038/sj.onc.1207630. [DOI] [PubMed] [Google Scholar]
- 196.SHIRAISHI T, YOSHIDA T, NAKATA S, et al. Tunicamycin enhances tumor necrosis factor-related apoptosis-inducing ligand-induced apoptosis in human prostate cancer cells. Cancer Res. 2005;65(14):6364–6370. doi: 10.1158/0008-5472.CAN-05-0312. [DOI] [PubMed] [Google Scholar]
- 197.LU M, KWAN T, YU C, et al. Peroxisome proliferator-activated receptor gamma agonists promote TRAIL-induced apoptosis by reducing survivin levels via cyclin D3 repression and cell cycle arrest. J Biol Chem. 2005;280(8):6742–6751. doi: 10.1074/jbc.M411519200. [DOI] [PubMed] [Google Scholar]
- 198.IZERADJENE K, DOUGLAS L, TILLMAN DM, DELANEY AB, HOUGHTON JA. Reactive oxygen species regulate caspase activation in tumor necrosis factor-related apoptosis-inducing ligand-resistant human colon carcinoma cell lines. Cancer Res. 2005;65(16):7436–7445. doi: 10.1158/0008-5472.CAN-04-2628. [DOI] [PubMed] [Google Scholar]
- 199.WIRTH T, KUHNEL F, FLEISCHMANN-MUNDT B, et al. Telomerase-dependent virotherapy overcomes resistance of hepatocellular carcinomas against chemotherapy and tumor necrosis factor-related apoptosis-inducing ligand by elimination of Mcl-1. Cancer Res. 2005;65(16):7393–7402. doi: 10.1158/0008-5472.CAN-04-3664. [DOI] [PubMed] [Google Scholar]
- 200.FAS SC, BAUMANN S, ZHU JY, et al. Wogonin sensitizes resistant malignant cells to TNFalpha- and TRAIL-induced apoptosis. Blood. 2006;108(12):3700–3706. doi: 10.1182/blood-2006-03-011973. [DOI] [PubMed] [Google Scholar]
- 201.OLTERSDORF T, ELMORE SW, SHOEMAKER AR, et al. An inhibitor of Bcl-2 family proteins induces regression of solid tumours. Nature. 2005;435(7042):677–681. doi: 10.1038/nature03579. [DOI] [PubMed] [Google Scholar]
- 202.YEOW WS, BARAS A, CHUA A, et al. Gossypol, a phytochemical with BH3-mimetic property, sensitizes cultured thoracic cancer cells to Apo2 ligand/tumor necrosis factor-related apoptosis-inducing ligand. J Thorac Cardiovasc Surg. 2006;132(6):1356–1362. doi: 10.1016/j.jtcvs.2006.07.025. [DOI] [PubMed] [Google Scholar]
- 203.FULDA S, WICK W, WELLER M, DEBATIN KM. Smac agonists sensitize for Apo2L/TRAIL- or anticancer drug-induced apoptosis and induce regression of malignant glioma in vivo. Nat Med. 2002;8(8):808–815. doi: 10.1038/nm735. [DOI] [PubMed] [Google Scholar]
- 204.LI L, THOMAS RM, SUZUKI H, et al. A small molecule Smac mimic potentiates TRAIL- and TNFalpha-mediated cell death. Science. 2004;305(5689):1471–1474. doi: 10.1126/science.1098231. [DOI] [PubMed] [Google Scholar]
- 205.PETRUCCI E, PASQUINI L, PETRONELLI A, et al. A small molecule Smac mimic potentiates TRAIL-mediated cell death of ovarian cancer cells. Gynecol Oncol. 2007 doi: 10.1016/j.ygyno.2007.01.011. [DOI] [PubMed] [Google Scholar]
- 206.SUH WS, KIM YS, SCHIMMER AD, et al. Synthetic triterpenoids activate a pathway for apoptosis in AML cells involving downregulation of FLIP and sensitization to TRAIL. Leukemia. 2003;17(11):2122–2129. doi: 10.1038/sj.leu.2403112. [DOI] [PubMed] [Google Scholar]
- 207.HYER ML, CROXTON R, KRAJEWSKA M, et al. Synthetic triterpenoids cooperate with tumor necrosis factor-related apoptosis-inducing ligand to induce apoptosis of breast cancer cells. Cancer Res. 2005;65(11):4799–4808. doi: 10.1158/0008-5472.CAN-04-3319. [DOI] [PubMed] [Google Scholar]
- 208.SCHIMMER AD, THOMAS MP, HURREN R, et al. Identification of small molecules that sensitize resistant tumor cells to tumor necrosis factor-family death receptors. Cancer research. 2006;66(4):2367–2375. doi: 10.1158/0008-5472.CAN-05-1061. [DOI] [PubMed] [Google Scholar]
- 209.DI PAOLO NC, TUVE S, NI S, et al. Effect of adenovirus-mediated heat shock protein expression and oncolysis in combination with low-dose cyclophosphamide treatment on antitumor immune responses. Cancer Res. 2006;66(2):960–969. doi: 10.1158/0008-5472.CAN-05-2388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210**.UNO T, TAKEDA K, KOJIMA Y, et al. Eradication of established tumors in mice by a combination antibody-based therapy. Nat Med. 2006;12(6):693–698. doi: 10.1038/nm1405. Good example of the use of the TRAIL system in immunotherapy. [DOI] [PubMed] [Google Scholar]