

Abstract
Background: The specific resting metabolic rates (Ki; in kcal · kg−1 · d−1) of major organs and tissues in adults were suggested by Elia (in Energy metabolism: tissue determinants and cellular corollaries. New York, NY: Raven Press, 1992) to be as follows: 200 for liver, 240 for brain, 440 for heart and kidneys, 13 for skeletal muscle, 4.5 for adipose tissue, and 12 for residual organs and tissues. However, Elia's Ki values have never been fully evaluated.
Objectives: The objectives of the present study were to evaluate the applicability of Elia's Ki values across adulthood and to explore the potential influence of age on the Ki values.
Design: A new approach was developed to evaluate the Ki values of major organs and tissues on the basis of a mechanistic model: REE = Σ(Ki × Ti), where REE is whole-body resting energy expenditure measured by indirect calorimetry, and Ti is the mass of individual organs and tissues measured by magnetic resonance imaging. With measured REE and Ti, marginal 95% CIs for Ki values were calculated by stepwise univariate regression analysis. An existing database of nonobese, healthy adults [n = 131; body mass index (in kg/m2) <30] was divided into 3 age groups: 21–30 y (young, n = 43), 31–50 y (middle-age, n = 51), and >50 y (n = 37).
Results: Elia's Ki values were within the range of 95% CIs in the young and middle-age groups. However, Elia's Ki values were outside the right boundaries of 95% CIs in the >50-y group, which indicated that Elia's study overestimated Ki values by 3% in this group. Age-adjusted Ki values for adults aged >50 y were 194 for liver, 233 for brain, 426 for heart and kidneys, 12.6 for skeletal muscle, 4.4 for adipose tissue, and 11.6 for residuals.
Conclusion: The general applicability of Elia's Ki values was validated across adulthood, although age adjustment is appropriate for specific applications.
INTRODUCTION
One of the primary aims of human energy metabolism research is to explore the specific metabolic rate (Ki value) for individual organs and tissues under resting conditions. Estimating the Ki values forms the basis for exploring the relation between resting energy expenditure (REE) and body composition and for understanding the daily energy requirements in humans (1–3). Different approaches, including in vitro and in vivo methods, have been used to estimate the Ki values of individual organs and tissues. When isolated organ and tissue slices are used, the in vitro method usually underestimates Ki values (4), whereas the in vivo method remains technically demanding (5–7).
On the basis of reported experimental results in humans and other mammals, Elia (1) presented a review on the Ki values (in kcal · kg−1 · d−1) for 7 organs and tissues in adults: liver (200), brain (240), heart (440), kidneys (440), skeletal muscle (13), adipose tissue (4.5), and residual mass (12). The residual mass includes other organs and tissues, such as skin, intestines, bones, and lungs. According to Elia, the heart and kidneys have the highest Ki values, twice those for liver and brain. In contrast, the Ki value of skeletal muscle is only 1/35 that of the heart and kidneys, and adipose tissue has the lowest Ki value of the 7 organs and tissues.
During the past decade, another approach was applied that measures masses of major organs and tissues by magnetic resonance imaging and then calculates whole-body REE based on Elia's Ki values (8, 9). Published studies showed that the predicted REE well matched the measured REE in young adults, which supports the applicability of Elia's Ki values. However, the predicted REE was significantly higher than the measured REE in elderly adults, which indicated that the Ki value may decline from a young age to an elderly age (3, 9, 10). The objectives of the present study were to critically evaluate the applicability of Elia's Ki values across adulthood and to explore the potential influence of age on Ki values.
METHODS
Development of approach
A new approach was developed by combining a mechanistic REE model with stepwise univariate regression analysis to evaluate the applicability of Elia's Ki values.
Mechanistic model of REE
This model is based on the concept that whole-body REE reflects the sum of resting energy consumption in all organs and tissues in the human body. A mechanistic model was developed that represents REE as the sum of the products of individual organ and tissue masses and their corresponding specific resting metabolic rates (11):
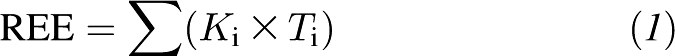 |
where Ti is the individual organ/tissue mass, i is the organ/tissue number (i = 1, 2, …, n), and Ki is the specific resting metabolic rate of the individual organs and tissues.
Because 4 organs (ie, liver, brain, heart, and kidneys) have particularly high basal specific metabolic rate and 2 tissues (ie, skeletal muscle and adipose tissue) have particularly large masses, the following body-composition model at the organ-tissue level was applied in the present study:
where BM is body mass and SM and AT are skeletal muscle and adipose tissue, respectively. Residual mass includes other tissues and organs such as skeleton, blood, skin, gastrointestinal tract, lung, and spleen. Residual mass in our study was calculated as BM minus the sum of liver, brain, heart, kidneys, skeletal muscle, and adipose tissue.
On the basis of the Ki values suggested by Elia, a working REE model was expressed as follows:
Stepwise univariate regression analysis
Our approach to evaluating Elia's Ki values was to examine each organ and tissue separately. We constructed separate marginal 95% CIs for each of the 7 Ki values via univariate linear regression analysis (12).
First, we evaluated the specific metabolic rate of liver (Kliver) with the statistical hypothesis Kliver = 200, suggested by Elia (1). According to Equation 3, we fitted a linear regression model:
In other words, we considered the following univariate linear regression:
where
An estimate with SE (95% CIs) for Kliver was obtained by using the least-squares method. We then compared the 95% CIs with the hypothesized Kliver value suggested by Elia. Testing statistical hypothesis Kliver = 200 at a significance level 0.05 was tantamount to checking whether Kliver = 200 falls inside the 95% CIs.
Second, we evaluated the specific metabolic rate of brain (Kbrain) and fitted a linear regression model according to Equation 3:
An estimate with SE (95% CIs) for Kbrain was obtained by using the least-squares method. We then compared it with the hypothesized value Kbrain = 240 suggested by Elia (1). The same process was repeated for each of the remaining coefficients, ie, Kheart, Kkidneys, KSM, KAT, and Kresidual.
Age-stratified mechanistic model of REE
The working REE model (ie, Equation 3) was based on an assumption that the Ki value of each organ and tissue is stable across adulthood. However, previous observations showed that the Ki value may decline from a young age to an elderly age (3). In the present study, the adult subject pool was divided into 3 age groups to assess the potential influence of age on the Ki values of major organs and tissues. An age-stratified mechanistic model of resting energy expenditure was applied:
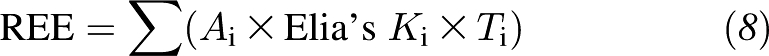 |
where Ai is the age-adjusted coefficient for Elia's Ki value for each age group. In the present study, an assumption was made that the age-adjusted Ai are the same across all organs and tissues for each age group, or Ai = A. A simplified age-stratified REE model was thus applied:
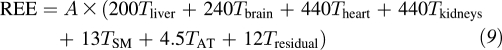 |
The A values of major organs and tissues were predicted in different age groups, respectively. The age-adjusted Ki values can then be calculated in each age group as
Subjects
Existing REE and organ and tissue data were collected at the Institute of Human Nutrition and Food Science, Christian-Albrechts University, Kiel, Germany. The Institutional Review Board at Christian-Albrechts University approved the original studies, and the subject signed an informed consent form. The subjects have been recruited since 2000. Healthy nonobese adult subjects (n = 131) were divided into 3 age groups: 21–30 y (young), 31–50 y (middle-age), and >50 y. Obese subjects [body mass index (BMI; in kg/m2) ≥ 30] were excluded from this study. The subjects in the present study had participated in other studies (9, 13).
Measurements
The existing database contains anthropometric measures; measured REE; measured organ and tissue masses including liver, brain, heart, kidneys, skeletal muscle, and adipose tissue; and percentage of body mass as fat.
Anthropometric measures
Anthropometric measurements were made by experienced observers using standardized procedures as reported by Lohman et al (14). Body mass was measured to the nearest 0.1 kg in fasting subjects wearing minimal clothing. Height was measured with a stadiometer to the nearest 0.1 cm.
REE
Indirect calorimetry was applied to estimate REE with participants in a postabsorptive state. No food or calorie-containing beverages were consumed after 1900 until the REE and all body-composition tests were completed. The subjects did not drink water before measurement. All subjects had been investigated in a metabolic ward at constant room temperature and humidity. REE was measured between 0700 and 0900 with subjects resting comfortably on a bed with a plastic transparent ventilated hood placed over their heads for 30 min. Continuous gas exchange measurements were taken to analyze the rates of O2 consumption and CO2 production by using a SensorMedics calorimeter (Vmax Spectra 29n; SensorMedics, Bilthoven, Netherlands). All gas exchange data were collected in the early morning after waking up, in a resting state 8 h after physical activity, at an environmental temperature of ≈ 25°C, and in a postabsorptive state 10–12 h after feeding (2).
Organ and tissue mass
Organ and tissue masses were obtained by summing pixels from images obtained with a 1.5-T Magnetom Vision scanner (Siemens, Erlangen, Germany). The magnetic resonance imaging (MRI) protocol details have been previously described (13).
In brief, for the brain, a T1-weighted fast low-angle shot (FLASH) breath-hold sequence was performed with repetition time of 174.9 ms, echo time of 4.1 ms, and flip angle of 80°. For the heart, ultrashort scans were made by electrocardiogram-triggered, T2-weighted half-single-shot turbo spin-echo (HASTE) sequences (breath-hold, repetition time 800 ms, echo time 43.0 ms, and acquisition time 20 ms). Liver and kidney images were produced by using an axial T1-weighted spin echo sequence. Total body SM and AT volumes were evaluated and involved the acquisition of ≈40 axial images across the whole body.
All MRI images were manually segmented with a software (TomoVision 4.3 Software; Slice-O-Matic, Montreal, Canada). Each organ and tissue was analyzed by the same observer who was blinded to the time point and subject identity. Visible AT areas within organ/tissue cross-sectional area were removed, including the small amount of visible AT within skeletal muscle bundles. The intraobserver CVs based on comparison of repeated segmentations were 0.07% for liver, 1.8% for brain, 1.7% for heart, and 1.0% for kidneys. The technical errors for measurement of the same scan on 2 separate days by the same observer of skeletal muscle and adipose tissue volumes were 0.7 ± 0.1% and 1.1 ± 1.2% (mean ± SD), respectively.
The mass of each organ and tissue was determined from the sum of all cross-sectional areas multiplied by the slice thickness, the gaps between slices, and the density of each organ and tissue (15):
where S is the cross-sectional area of each organ/tissue image, i is the image number, t is the thickness of each image, g is the gap (distance) between consecutive images, and d is the density of organ/tissue. The t, g, and d values of major organs and tissues are summarized in Table 1.
TABLE 1.
Thickness and gap of magnetic resonance imaging (MRI) for major organs and tissues1
| Liver | Brain | Heart | Kidneys | SM | AT | |
| t (cm) | 0.8 | 0.6 | 0.7 | 0.8 | 1.0 | 1.0 |
| g (cm) | 0.24 | 0.12 | 0.21 | 0.24 | 4.0 | 4.0 |
| d (g/cm3) | 1.06 | 1.036 | 1.06 | 1.05 | 1.04 | 0.92 |
d, density of organ and tissue; g, gap (distance) between consecutive MRI scans; t, thickness of each MRI scan; AT, adipose tissue; SM, skeletal muscle.
Dual-energy X-ray absorptiometry
Total body fat and bone mineral content were measured with a dual-energy X-ray absorptiometry scanner (software version V8.26a:3, model QDR 4500A; Hologic, Waltham, MA). The subjects lay supine with arms and legs at their sides during the 10-min scan. The between-measurement technical error for fat in the same person is 1.2% (16). The percentage of body mass as fat was then calculated. In some subjects, skeletal muscle and adipose tissue masses were calculated from dual-energy X-ray absorptiometry estimation, as previously described (13). Skeletal muscle mass was predicted from appendicular lean soft tissue (17), and adipose tissue mass was predicted from fat mass, assuming a fat content of 80% (18).
Statistical analysis
Descriptive statistics from the data were expressed as the group mean ± SD. Statistical analyses were performed by using SPSS for Windows 13.0 (SPSS Inc, Chicago, IL). Differences in body composition and REE between the 3 age groups were analyzed by analysis of variance (ANOVA) and pairwise comparisons with Bonferroni adjustment. Statistical significance was set at P < 0.05. Elia's Ki values for the 7 organs and tissues were applied to predict REE and examine the association between measured REE (REEm) and predicted REE (REEp) with the use of simple linear regression analysis. We then explored for bias in the REEm and REEp relation by using the method reported by Bland and Altman (19). The marginal 95% CIs for the 7 Ki values were predicted via the simple univariate linear regression method (12). The database was analyzed by the programming in R version 2.10.1, a software program for statistical computing and graphics, initially written by Robert Gentleman and Ross Ihaka, Statistics Department, University of Auckland.
RESULTS
Physical characteristics
In this secondary analysis study, 131 (67 men, 64 women) nonobese healthy adults were included and divided into 3 age groups: 43 young adults aged 21–30 y, 51 middle-age adults aged 31–50 y, and 37 adults aged 51–73 y. The baseline characteristics and body composition of all subjects and the 3 age groups are presented in Table 2. No differences in height, fat mass, and bone mineral content were observed between the 3 age groups (both ANOVA test and pairwise comparisons with Bonferroni adjustment, all P > 0.1). However, body mass and BMI were significantly different between the young, middle-age, and >50-y groups (ANOVA, P = 0.034 for body mass, P = 0.001 for BMI). Pairwise comparisons with Bonferroni adjustment showed that the differences in body mass and BMI were significant only between the young and >50-y groups (P = 0.04 for body mass, P < 0.001 for BMI). Fat-free mass in the >50-y group (58.4 ± 11.0 kg) was greater than that in the young group (53.3 ± 10.0 kg), by 5.1 kg (Table 3). The major reason was the imbalance in subject number between men and women (26 men and 11 women in the >50-y group compared with 16 men and 27 women in the young group). However, this difference was not statistically significant (pairwise comparisons with Bonferroni adjustment, P > 0.1).
TABLE 2.
Subject characteristics and body composition by age group1
| All subjects | 21–30 y (young) | 31–50 y (middle-age) | 51–73 y | P2 | |
| Subjects (n) | 131 | 43 | 51 | 37 | — |
| Men | 67 | 16 | 25 | 26 | |
| Women | 64 | 27 | 26 | 11 | |
| Age (y) | 41.8 ± 14.83 | 25.9 ± 2.0 | 40.5 ± 5.1 | 62.0 ± 5.3 | — |
| Body mass (kg) | 73.7 ± 12.4 | 70.0 ± 11.3 | 74.8 ± 12.5 | 76.7 ± 12.7 | 0.0344 |
| Height (m) | 1.74 ± 0.08 | 1.74 ± 0.06 | 1.75 ± 0.09 | 1.73 ± 0.08 | 0.5945 |
| BMI (kg/m2) | 24.2 ± 2.9 | 23.0 ± 2.7 | 24.4 ± 2.8 | 25.4 ± 2.7 | 0.0016 |
| Fat (kg) | 17.9 ± 6.7 | 16.6 ± 6.7 | 18.6 ± 6.6 | 18.3 ± 6.7 | 0.3245 |
| Fat (%) | 25.0 ± 8.3 | 24.3 ± 8.7 | 25.8 ± 8.2 | 24.6 ± 8.1 | 0.6615 |
| FFM (kg) | 55.9 ± 11.4 | 53.3 ± 10.0 | 56.2 ± 12.3 | 58.4 ± 11.0 | 0.1225 |
| BMC (kg) | 2.58 ± 0.46 | 2.53 ± 0.40 | 2.61 ± 0.46 | 2.62 ± 0.54 | 0.6035 |
BMC, bone mineral content; FFM, fat-free mass.
Tests of group mean differences by ANOVA and pairwise analyses with Bonferroni adjustment.
Mean ± SD (all such values).
Young compared with middle-age (P > 0.1), young compared with >50 y (P = 0.04), and middle-age compared with >50 y (P > 0.1).
Pairwise comparisons with Bonferroni adjustment, P > 0.1.
Young compared with middle-age (P = 0.07), young compared with >50 y (P < 0.001), and middle-age compared with >50 y (P > 0.1).
TABLE 3.
Major organ and tissue masses and whole-body resting energy expenditure (REE)1
| All subjects | 21–30 y (young) | 31–50 y (middle-age) | 51–73 y | P2 | |
| Liver (kg) | 1.39 ± 0.253 | 1.35 ± 0.23 | 1.41 ± 0.25 | 1.41 ± 0.28 | 0.5134 |
| Brain (kg) | 1.33 ± 0.11 | 1.33 ± 0.11 | 1.34 ± 0.10 | 1.32 ± 0.12 | 0.7664 |
| Heart (kg) | 0.31 ± 0.08 | 0.31 ± 0.09 | 0.30 ± 0.08 | 0.33 ± 0.07 | 0.3274 |
| Kidneys (kg) | 0.29 ± 0.06 | 0.28 ± 0.06 | 0.28 ± 0.05 | 0.31 ± 0.06 | 0.0425 |
| SM (kg) | 26.3 ± 6.3 | 25.0 ± 5.9 | 26.7 ± 6.6 | 26.9 ± 6.4 | 0.5324 |
| AT (kg) | 19.4 ± 6.4 | 18.4 ± 6.4 | 19.9 ± 6.3 | 19.9 ± 6.5 | 0.4434 |
| Residual mass (kg) | 24.7 ± 5.2 | 22.8 ± 3.9 | 24.9 ± 6.0 | 26.5 ± 4.6 | 0.0046 |
| REEm (kcal/d) | 1575 ± 241 | 1547 ± 241 | 1590 ± 248 | 1586 ± 234 | 0.6494 |
| REEp (kcal/d) | 1588 ± 234 | 1535 ± 220 | 1596 ± 239 | 1636 ± 236 | 0.1474 |
| REEm − REEp (kcal/d)7 | −13 ± 80 (0.068) | 11 ± 80 (0.36) | −6 ± 79 (0.59) | −50 ± 67 (<0.001) | — |
AT, adipose tissue; REEm, REE measured by indirect calorimetry; REEp, REE predicted by the Ki values suggested by Elia (1); SM, skeletal muscle.
Tests of group mean differences for organ and tissue masses and REE by ANOVA and pairwise analyses with Bonferroni adjustment.
Mean ± SD (all such values).
Pairwise comparisons with Bonferroni adjustment, P > 0.1.
Young compared with middle-age (P > 0.1), young compared with >50 y (P = 0.06), and middle-age compared with >50 y (P > 0.1).
Young compared with middle-age (P > 0.1), young compared with >50 y (P < 0.01), and middle-age compared with >50 y (P > 0.1).
Values in parentheses are the P values for the difference between REEm and REEp within relevant groups by paired t test.
Organ and tissue masses and REE
The masses of 4 high-metabolic-rate organs (ie, liver, brain, heart, and kidneys) and 3 low-metabolic-rate tissues (ie, skeletal muscle, adipose tissue, and residual mass) for all subjects and 3 age groups are presented in Table 3. No significant differences in liver, brain, heart, skeletal muscle, and adipose tissue mass were observed between the 3 groups (both ANOVA and pairwise comparisons with Bonferroni adjustment, all P > 0.1). For kidneys, a significant difference was observed between the 3 groups (ANOVA, P = 0.042). Pairwise comparisons with Bonferroni adjustment showed that the differences in kidneys were not significant, although it was nearly significant between the young and >50-y groups (P = 0.06). For residual mass, a significant difference was observed between the 3 groups (ANOVA, P = 0.004). Pairwise comparisons with Bonferroni adjustment showed that the difference in residual mass was significant only between the young and >50-y groups (P < 0.01).
The REEm for all subjects and the 3 age groups are presented in Table 3. There was no significant difference in REEm between the 3 groups (both ANOVA and pairwise comparisons with Bonferroni adjustment, all P > 0.1).
Evaluation of Eliarsquos Ki values in all subjects
According to Equation 3, REEp was calculated for all subjects. The REEm and REEp (mean ± SD) were 1575 ± 241 and 1588 ± 234 kcal/d, respectively (Table 3). No significant difference was observed between REEm and REEp for all subjects (paired Student's t test, P = 0.068), with a mean difference (ie, REEm − REEp) of −13 ± 80 kcal/d. A Bland-Altman plot showed that no significant trend (r = 0.09, P > 0.20) was observed between the REEm and REEp difference compared with the average of REEm and REEp (Figure 1).
FIGURE 1.

The difference between measured and predicted resting energy expenditure (ie, REEm – REEp) compared with the mean of REEm and REEp for all subjects. (REEm – REEp) = 0.0306 × mean – 61.2; r = 0.09, P > 0.20; n = 131. REEp was calculated by using the Ki values suggested by Elia (1), according to Equation 3. The regression line, zero difference line, and the lines representing 2 SDs for the differences (146, −172 kcal/d; indicated by the upper and lower lines) are shown.
The stepwise univariate analysis method was used to calculate the 95% CIs for each Ki value of the organs and tissues for all subjects (Table 4). As shown in Figure 2, Elia's Ki values were within but close to the right boundary of 95% CIs for the 7 organs and tissues.
TABLE 4.
95% CIs of specific metabolic rates of major organs and tissues1
| Organ/tissue and Elia's Ki value | All subjects | 21–30 y (young) | 31–50 y (middle-age) | 51–73 y |
| Liver (200 kcal · kgminus1 · dminus1) | ||||
| 95% CIs for corresponding Ki values | (182, 201) | (191, 227) | (181, 212) | (150, 181) |
| P value from t test of H02 | 0.087 | 0.31 | 0.67 | <0.001 |
| Marginal 2 value3 | 0.92 (0.92) | 0.93 (0.93) | 0.93 (0.93) | 0.93 (0.89) |
| Brain (240 kcal · kgminus1 · dminus1) | ||||
| 95% CIs for corresponding Ki values | (220, 241) | (230, 267) | (219, 252) | (186, 219) |
| P value from t test of H02 | 0.073 | 0.34 | 0.57 | <0.001 |
| Marginal 2 value3 | 0.94 (0.94) | 0.95 (0.95) | 0.94 (0.94) | 0.94 (0.91) |
| Heart (440 kcal · kgminus1 · dminus1) | ||||
| 95% CIs for corresponding Ki values | (348, 434) | (402, 554) | (338, 479) | (213, 338) |
| P value from t test of H02 | 0.024 | 0.32 | 0.37 | <0.001 |
| Marginal 2 value3 | 0.72 (0.71) | 0.79 (0.79) | 0.73 (0.72) | 0.68 (0.44) |
| Kidneys (440 kcal · kgminus1 · dminus1) | ||||
| 95% CIs for corresponding Ki values | (336, 430) | (396, 568) | (328, 481) | (200, 332) |
| P value from t test of H02 | 0.016 | 0.34 | 0.35 | <0.001 |
| Marginal 2 value3 | 0.68 (0.66) | 0.75 (0.74) | 0.69 (0.69) | 0.65 (0.37) |
| SM (13 kcal · kgminus1 · dminus1) | ||||
| 95% CIs for corresponding Ki values | (12.0, 13.0) | (12.5, 14.4) | (12.0, 13.6) | (10.3, 11.9) |
| P value from t test of H02 | 0.051 | 0.35 | 0.59 | <0.001 |
| Marginal 2 value3 | 0.95 (0.95) | 0.95 (0.95) | 0.95 (0.95) | 0.96 (0.93) |
| AT (4.5 kcal · kgminus1 · dminus1) | ||||
| 95% CIs for corresponding Ki values | (3.21, 4.57) | (3.85, 6.37) | (3.14, 5.26) | (1.16, 3.34) |
| P value from t test of H02 | 0.077 | 0.34 | 0.58 | <0.001 |
| Marginal 2 value3 | 0.50 (0.49) | 0.61 (0.60) | 0.56 (0.55) | 0.32 (0.00) |
| Res (12 kcal · kgminus1 · dminus1) | ||||
| 95% CIs for corresponding Ki values | (10.9, 12.0) | (11.5, 13.6) | (10.9, 12.6) | (9.3, 11.0) |
| P value from t test of H02 | 0.051 | 0.32 | 0.59 | <0.001 |
| Marginal 2 value3 | 0.93 (0.93) | 0.93 (0.93) | 0.94 (0.94) | 0.94 (0.91) |
AT, adipose tissue; Res, residual mass; SM, skeletal muscle.
H0: Ki = Elia's Ki value.
The proportion of marginal variability reduction; for liver, reduction was due to fitting least squares with Equation 5. The value within parentheses is the corresponding reduction by taking Kliver = 200 (Elia's value). This is likewise defined for other organs and tissues.
FIGURE 2.

The 95% marginal CIs for Ki values of major organs and tissues, fitted by stepwise univariate analysis and shown on a logarithmic scale, for all adult subjects. The Xs represent the Ki values suggested by Elia (1). AT, adipose tissue; Res, residual mass; SM, skeletal muscle.
Evaluation of Eliarsquos Ki values in 3 age groups
A plot between REEm – REEp and age for all subjects showed that the REEm and REEp difference was significantly associated with age (r = −0.27, P < 0.01; Figure 3). We thus further evaluated Elia's Ki values in different age groups.
FIGURE 3.

The difference between measured and predicted resting energy expenditure (ie, REEm – REEp) compared with age for all subjects. (REEm – REEp) = 47.7 − 1.45 × age; r = −0.27, P < 0.01; n = 131. REEp was calculated by using the Ki values suggested by Elia (1), according to Equation 3. The zero difference line is shown.
According to Equation 3, REEp, as well as the differences between REEm and REEp, were calculated for the 3 age groups (Table 3). No significant differences were observed between REEm and REEp within the young (11 ± 80 kcal/d; P = 0.36) and middle-age (−6 ± 79 kcal/d; P = 0.59) groups. However, the REEm – REEp values were significant in the >50-y group (−50 ± 67 kcal/d, P < 0.001).
The stepwise univariate analysis method was used to calculate the 95% CIs of Ki values for the 3 age groups (Table 4). For the young and middle-age adults, Elia's Ki values were located within the 95% CIs for the 7 organs and tissues. For the >50-y group, however, Elia's Ki values were outside the right boundary of 95% CIs for the 7 organs and tissues (Figure 4). This observation indicated that Elia's study overestimated Ki values in the adults aged >50 y.
FIGURE 4.

The 95% marginal CIs for Ki values of major organs and tissues, fitted by stepwise univariate analysis and shown on a logarithmic scale, for the young (upper line), middle-age (middle line), and >50-y (lower line) groups. The Xs represent the Ki values suggested by Elia (1). AT, adipose tissue; Res, residual mass; SM, skeletal muscle.
Modification of Eliarsquos Ki values in the gt50-y group
According to Equation 7, age-adjusted coefficients were derived: A = 1.008 (P = 0.30) for the young adults, A = 0.996 (P = 0.54) for the middle-age adults, and A = 0.969 (P < 0.001) for the adults aged >50 y. According to Equation 8, the age-adjusted Ki values (in kcal · kg−1 · d−1) and their 95% CIs were calculated for the >50-y group (Table 5): 194 for liver, 233 for brain, 426 for heart and kidneys, 12.6 for skeletal muscle, 4.4 for adipose tissue, and 11.6 for residual mass.
TABLE 5.
Age-adjusted Ki values (and their 95% CIs) of specific metabolic rates of major organs and tissues1
| Organ/tissue | Elia's Ki value | 21–30 y (young) | 31–50 y (middle-age) | 51–73 y |
| kcal · kg−1 · d−1 | kcal · kg−1 · d−1 | kcal · kg−1 · d−1 | kcal · kg−1 · d−1 | |
| Liver | 200 | 202 (199, 205) | 199 (197, 202) | 194 (191, 197) |
| Brain | 240 | 242 (238, 245) | 239 (236, 242) | 233 (229, 236) |
| Heart | 440 | 443 (437, 450) | 438 (432, 444) | 426 (420, 433) |
| Kidneys | 440 | 443 (437, 450) | 438 (432, 444) | 426 (420, 433) |
| SM | 13 | 13.1 (12.9, 13.3) | 12.9 (12.8, 13.1) | 12.6 (12.4, 12.8) |
| AT | 4.5 | 4.54 (4.47, 4.60) | 4.48 (4.42, 4.54) | 4.36 (4.29, 4.43) |
| Res | 12 | 12.1 (11.9, 12.3) | 12.0 (11.8, 12.1) | 11.6 (11.4, 11.8) |
AT, adipose tissue; Res, residual mass; SM, skeletal muscle.
DISCUSSION
Exploring the specific resting metabolic rates of individual organs and tissues is one of the primary aims for energy metabolism research. Several approaches have been applied to estimate the Ki values in humans and several species of mammals. The innovation of the present study was to develop and apply a new approach to critically evaluate the applicability of Elia's Ki values across adulthood.
In vitro and in vivo approaches of estimating Ki values in humans
Since the 1920s, major efforts have been applied to the estimation of Ki values in isolated organ and tissue slices in several mammalian species by varying the substrates and estimation conditions (20, 21). However, a critical question raised by the in vitro technique is whether the estimated Ki values from isolated organ and tissue slices are comparable with the in vivo specific metabolic rates of the same organ and tissue in the intact living animal. On the whole, in vitro estimated Ki values are underestimated (4).
In vivo studies were then developed so that the Ki value of an organ or tissue could be estimated by measuring the arteriovenous difference in oxygen concentrations across the organ, combined with the assessment of blood flow perfusing the organ (6, 7). More recently, the Ki values of the individual organs were assessed by positron emission tomography with 13C, 15O, 18F, or 31P markers, such as 2-deoxy-2[18F]fluoro-d-glucose (5, 22). However, assessment of Ki by positron emission tomography has limited value depending on the tracer used, because the most frequently used glucose tracer is not specific for energy metabolism. Moreover, the in vivo techniques are expensive and remain technically demanding for humans.
The REEm minus REEp approach of evaluating Eliarsquos Ki values in humans
Another approach has been applied to evaluate the applicability of Elia's Ki values that compares REEp with REEm (8). The predicted REE is calculated by Equation 3 with the Ki values suggested by Elia. If the REEm – REEp difference is close to zero, we may consider that Elia's Ki values should be applicable as a whole.
This approach is dependent on simultaneous measurements of REE and organs and tissues from the same subjects. Currently, only 2 laboratories have measured REE and organs and tissues using magnetic resonance imaging: Christian-Albrechts University, Kiel, Germany and St Luke's–Roosevelt Hospital, Columbia University, New York, NY (8, 23, 24). The REEm – REEp differences reported in the previous and present studies are summarized in Table 6.
TABLE 6.
Differences between measured and predicted resting energy expenditure (REE) for the different age groups1
| Group | Age (range) | BMI | n | REEm − REEp (P value) | Laboratory | Reference |
| y | kg/m2 | |||||
| Children | 9.3 ± 1.72 (6–12) | 16.3 ± 3.6 | 15 (8 M, 7 F) | 299 ± 121 (<0.001) | NY | (25) |
| Adolescence | 14.7 ± 0.6 | 20.5 ± 3.5 | 20 M | 118 ± 165 (<0.01) | NY | (26) |
| Young | 22.9 ± 2.5 (18–50) | 22.9 ± 2.5 | 13 (8 M, 5 F) | 19.9 ± 126 (>0.05) | NY | (8) |
| Young | 24.8 ± 2.4 | 21.8 ± 2.2 | 13 F | −10 ± 72 | Kiel | (9) |
| Young | 25.9 ± 2.0 (21–30) | 23.0 ± 2.7 | 43 (16 M, 27 F) | 11 ± 80 (>0.05) | Kiel | Current study |
| Young | 26.2 ± 2.1 | 22.5 ± 1.8 | 13 M | 24 ± 120 | Kiel | (9) |
| Middle-age | 40.5 ± 5.1 (31–50) | 24.4 ± 2.8 | 51 (25 M, 26 F) | −6 ± 79 (>0.05) | Kiel | Current study |
| >50 y | 62.0 ± 5.3 (51–73) | 25.4 ± 2.7 | 37 (26 M, 11 F) | −50 ± 67 (<0.001) | Kiel | Current study |
| Elderly | 64.9 ± 2.7 | 25.0 ± 3.0 | 7 M | −79 ± 96 | Kiel | (9) |
| Elderly | 76.5 ± 5.5 (>70) | 24.5 ± 3.9 | 6 M | −144 ± 64 (<0.01) | NY | (10) |
| Elderly | 80.3 ± 7.5 (>70) | 23.1 ± 3.6 | 7 F | −146 ± 78 (<0.001) | NY | (10) |
Kiel, Institute of Human Nutrition and Food Science, Christian-Albrechts University, Kiel, Germany; NY, Obesity Research Center, St Luke's–Roosevelt Hospital, Columbia University College of Physicians and Surgeons, New York, NY; REEm, REE measured by indirect calorimetry; REEp, REE predicted by the Ki values suggested by Elia (1).
Mean ± SD (all such values).
The St Luke's research group reported that Equation 3 underpredicted REE in children and adolescents, which indicated that Ki values are higher during growth and development than in adulthood (25, 26).
In contrast, the present study found that the REEm – REEp was < 0 in the >50-y group and was negatively associated with age (r = −0.27, P < 0.01; Figure 3). The St Luke's research group also showed that Equation 3 overpredicts REE in the elderly adults, although well-predicts REE in young and middle-age adults (3, 10). Caution should be taken that the age ranges in the previous studies of elderly adults were at the upper limit or even above the range of the subjects in the present study. This may explain the even greater REEm – REEp difference observed in the previous studies (9, 10).
New approach of evaluating Eliarsquos Ki values in adult humans
To critically evaluate the applicability of Elia's Ki values across adulthood, a new approach was developed that combines the mechanistic REE model with the stepwise univariate analysis method. Our results showed that Elia's Ki values were close to the right boundary of 95% CIs for the 7 organs and tissues in all subjects (Figure 2). This observation validated the applicability of Elia's Ki values in whole adulthood. Also, Elia's Ki values were located within the 95% CIs for the 7 organs and tissues in the young and middle-age adults (Figure 4). The age-adjusted coefficients were close to 1 and nonsignificant: A = 1.008 (P = 0.30) for the young group and A = 0.996 (P = 0.54) for the middle-age group. This observation supports the applicability of Elia's Ki values in young and middle-age adults.
For the >50-y group, Elia's Ki values were outside the right boundary of 95% CIs for the 7 organs and tissues (Figure 4). A significantly age-adjusted coefficient, A = 0.969 (P < 0.001), should be applied for this group of adults. On the basis of the predicted age-adjusted coefficient, the age-modified Ki values were suggested (Table 5).
Our results showed that the Ki values were significantly lower by 3% in the adults aged >50 y than in the young and middle-age adults. There are 3 possible explanations for the observations. First, the cellular fraction of organs and tissues is lower in elderly adults than in young and middle-age adults. In support of this explanation, well-established changes in the whole body and liver showed an expansion of the extracellular compartments and a relative loss of cellularity (3, 27, 28). The effect of organ fat, which may differ between age groups, on the Ki values should also be considered. Second, the specific metabolic rate of individual cell category may be lower in the elderly than in young adults. In other words, elderly adults may have a lower resting energy per unit cell mass than do young adults. Third, the lower Ki values in the elderly adults may have been caused by the combination of both the lower cellularity of organs and tissues and the lower specific metabolic rate of individual cell category. Further study is needed to find the actual reason of decline in Ki values across adulthood.
The association between REE and organ and tissue masses (ie, Equation 1) appears to be a natural candidate for the use of multiple linear regression (MLR), with 7 explanatory variables. Implementation of MLR method does not require any prior knowledge on the Ki values of major organs and tissues. However, because of the high collinearity among some organs and tissues, the MLR approach produced unstable results in the database used in the current study. Specifically, the SEs for the resulting estimators of regression coefficients were exceptionally large; therefore, the resulting CIs provided little information to the true values. For example, the 95% CIs for Kheart were as wide as (−185, 643) from fitting an MLR model.
Our new approach, which combines a mechanistic REE model with stepwise univariate regression analysis, can avoid the limitations of the MLR method. The existence of Elia's REE model (ie, Equation 3) allows us to evaluate each Ki value separately by holding the remaining Ki values at Elia's proposed values. The new approach thus provides an effective way to evaluate Elia's Ki values. In particular, we were able to construct tight CIs that can be used to assess Elia's Ki values and to predict the direction of any deviation from their coefficients. By introducing a common scaling factor, we were also able to quantify the adjustment needed for different age groups to better predict their respective REE.
In conclusion, the current study validated the applicability of Elia's Ki values in whole adulthood. Although correctly estimated in young and middle-age adults, Elia's study overestimated Ki values in the >50-y group; therefore, age-modified Ki values should be applied in the elderly. This study can thus help in understanding the inherent relation between REE and body composition at the organ-tissue level.
Acknowledgments
We are grateful to those subjects who were included in this study.
The authors’ responsibilities were as follows—ZW: responsible for all aspects of the study, including study design, model development, and manuscript writing; BS and WL: responsible for the magnetic resonance imaging organ and tissue segmentation procedures; ZY and JZ: responsible for model development, statistical analysis, and manuscript writing; AB-W and MJM: provided existing REE and organ and tissues database and responsible for manuscript writing; and SBH: provided consultation and assisted with manuscript writing. None of the authors had a conflict of interest concerning any company or organization sponsoring this study.
REFERENCES
- 1.Elia M. Organ and tissue contribution to metabolic rate. Kinney JM, Tucker HN, Energy metabolism: tissue determinants and cellular corollaries. New York, NY: Raven Press, 1992:61–80 [Google Scholar]
- 2.FAO/WHO/UNU Human energy requirements. Rome, Italy: Joint FAO/WHO/UNU Expert Consultation, 2004:35–50 [Google Scholar]
- 3.Wang Z, Heshka S, Heymsfield SB, Shen W, Gallagher D. A cellular level approach to predicting resting energy expenditure across the adult years. Am J Clin Nutr 2005;81:799–806 [DOI] [PubMed] [Google Scholar]
- 4.Schmidz-Nielsen K. Scaling: why is animal size so important? Cambridge, United Kingdom: Cambridge University Press, 1984 [Google Scholar]
- 5.Chugani HT, Phelps ME, Mazziotta JC. Positron emission tomography study of human brain functional development. Ann Neurol 1987;22:487–97 [DOI] [PubMed] [Google Scholar]
- 6.Linde B, Hjemdahl P, Freyschuss U, Juhlin-Dunndelt A. Adipose tissue and skeletal muscle blood flow during mental stress. Am J Physiol 1989;256:E12–8 [DOI] [PubMed] [Google Scholar]
- 7.McCully KK, Posner JD. The application of blood flow measurements to the study of aging muscle. J Gerontol 1995;50:130–6 [DOI] [PubMed] [Google Scholar]
- 8.Gallagher D, Belmonte D, Deurenberg P, et al. Organ-tissue mass measurement allows modeling of REE and metabolically active tissue mass. Am J Physiol 1998;275:E249–58 [DOI] [PubMed] [Google Scholar]
- 9.Bosy-Westphal A, Eichhorn C, Kutzner D, Illner K, Heller M, Müller MJ. The age-related decline in resting energy expenditure in humans is due to the loss of fat-free mass and to alterations in its metabolically active components. J Nutr 2003;133:2356–62 [DOI] [PubMed] [Google Scholar]
- 10.Gallagher D, Allen A, Wang ZM, Heymsfield SB, Krasnow N. Smaller organ tissue mass in the elderly fails to explain lower resting metabolic rate. Ann N Y Acad Sci 2000;904:449–55 [DOI] [PubMed] [Google Scholar]
- 11.Wang Z, Heshka S, Zhang K, Boozer C, Heymsfield SB. Resting energy expenditure: systematic organization and critique of prediction methods. Obes Res 2001;9:331–6 [DOI] [PubMed] [Google Scholar]
- 12.Weisberg S. Applied linear regression. 3rd ed Hoboken, NJ: John Wileys & Sons, 2005 [Google Scholar]
- 13.Bosy-Westphal A, Reinecke U, Schlorke T, et al. Effect of organ and tissue masses on resting energy expenditure in underweight, normal weight and obese adults. Int J Obes Relat Metab Disord 2004;28:72–9 [DOI] [PubMed] [Google Scholar]
- 14.Lohman TC, Roche AF, Martorell R, Anthropometric standardization reference manual. Champaign, IL: Human Kinetics, 1988 [Google Scholar]
- 15.Ross R, Janssen I. Computed tomography and magnetic resonance imaging. Heymsfield SB, Lohman TG, Wang ZM, Going SB, Human body composition. 2nd ed Champaign, IL: Human Kinetics, 2005:89–108 [Google Scholar]
- 16.Lohman TG, Chen Z. Dual energy X-ray absorptiometry. Heymsfield SB, Lohman TG, Wang ZM, Going SB, Human body composition. 2nd ed Champaign, IL: Human Kinetics, 2005:63–78 [Google Scholar]
- 17.Kim J, Wang ZM, Heymsfield ZM, Baumgartner RN, Gallagher D. Total-body skeletal muscle mass: estimation by new dual-energy X-ray absorptiometry method. Am J Clin Nutr 2002;76:378–83 [DOI] [PubMed] [Google Scholar]
- 18.Snyder WS, Cook MJ, Nasset ES, Karhausen LR, Howells GP, Tipton IH. Report of the Task Group on Reference Man. Oxford, United Kingdom: Pergamon Press, 1975 [Google Scholar]
- 19.Bland JM, Altman DG. Statistical methods for assessing agreement between two methods of clinical measurement. Lancet 1986;1:307–10 [PubMed] [Google Scholar]
- 20.Terroine EF, Roche J. La respiration des tissue. I. Production calorique des Homeothermas et intensite de la respiration in vitro des homologues. [Tissue respiration. I. Caloric expenditure of Homeotherms and intensity of the in vitro respiration of homologues.] Arch Int Physiol 1925;24:356–99 [Google Scholar]
- 21.Couture P, Hulbert AJ. Relationship between body mass, tissue metabolic rate, and sodium pump activity in mammalian liver and kidney. Am J Physiol 1995;268:R641–50 [DOI] [PubMed] [Google Scholar]
- 22.Petersen KF, Beftoy D, Dufour S, et al. Mitochondrial dysfunction in the elderly: possible role in insulin resistance. Science 2003;300:1140–2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Illner K, Brinkmann G, Heller M, Bosy-Westphal A, Müller MJ. Metabolically active components of fat free mass and resting energy expenditure in non obese adults. Am J Physiol 2000;278:E308–15 [DOI] [PubMed] [Google Scholar]
- 24.Müller MJ, Bosy-Westphal A, Kutzner D, Heller M. Metabolically active components of fat-free mass and resting energy expenditure in humans: recent lessons from imaging technologies. Obesity (Silver Spring) 2002;3:113–22 [DOI] [PubMed] [Google Scholar]
- 25.Hsu A, Heshka S, Janumala I, et al. Larger mass of high-metabolic-rate organs does not explain higher resting energy expenditure in children. Am J Clin Nutr 2003;77:1506–11 [DOI] [PubMed] [Google Scholar]
- 26.Wang Z, Heymsfield SB, Ying Z, Pierson RN, Jr, Gallagher D, Gidwani S. A cellular level approach to predicting resting energy expenditure: Evaluation of applicability in adolescents. Am J Hum Biol 2010;22:476–83 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wakabayashi H, Nishiyama Y, Ushivama T, Maeba T, Maeta H. Evaluation of the effect of age on functioning hepatocyte mass and liver blood flow using liver scintigraphy in preoperative estimations for surgical patients: comparison with CT volumetry. J Surg Res 2002;106:246–53 [DOI] [PubMed] [Google Scholar]
- 28.Wang Z, Heshka S, Wang J, et al. Metabolically active portion of fat-free mass: a cellular body composition level modeling analysis. Am J Physiol Endocrinol Metab 2007;292:E49–53 [DOI] [PMC free article] [PubMed] [Google Scholar]