

Abstract
To maximize fitness, animals must respond to a variety of processes that operate at different rates or timescales. Appropriate decisions could therefore involve complex interactions among these processes. For example, eiders wintering in the arctic sea ice must consider locomotion and physiology of diving for benthic invertebrates, digestive processing rate and a nonlinear decrease in profitability of diving as currents increase over the tidal cycle. Using a multi-scale dynamic modelling approach and continuous field observations of individuals, we demonstrate that the strategy that maximizes long-term energy gain involves resting during the most profitable foraging period (slack currents). These counterintuitive foraging patterns are an adaptive trade-off between multiple overlapping rate processes and cannot be explained by classical rate-maximizing optimization theory, which only considers a single timescale and predicts a constant rate of foraging. By reducing foraging and instead digesting during slack currents, eiders structure their activity in order to maximize long-term energetic gain over an entire tide cycle. This study reveals how counterintuitive patterns and a complex functional response can result from a simple trade-off among several overlapping rate processes, emphasizing the necessity of a multi-scale approach for understanding adaptive routines in the wild and evaluating mechanisms in ecological time series.
Keywords: foraging, diving, digestion, scale, time series, arctic ecology
1. Introduction
A variety of processes occurring at different rates or timescales can influence the dynamics of animal behaviour. Classic rate-maximizing approaches to the economics of behaviour (e.g. Charnov 1976) only consider a single timescale in isolation, assuming optimization currencies scale to longer term fitness, and predicting a constant rate of activity. However, complex and seemingly counterintuitive dynamics could result from the interactions among behavioural, physiological and abiotic rate processes with different timescales, necessitating a multi-scale approach (McNamara et al. 1987; Wiens 1989; Levin 1992; Schneider 1994, 2001). For example, research on locomotion and exercise physiology indicates that a short-term reduction in the rate of locomotion can actually increase the amount of work performed in the longer term, because endurance increases (Kramer & McLaughlin 2001; Wienstein 2001; see also Ydenberg & Hurd 1998; Heath et al. 2008). Similarly, Heath et al. (2007) demonstrated that diving models based on rate maximization (Kramer 1988; Houston & Carbone 1992) were inadequate to explain diving patterns of foraging eider ducks, probably owing to longer term processes such as digestion. Behavioural routines can be influenced by a variety of physiological and ecological processes with different timescales, resulting in dynamics that may not be easily explained without formal analysis and precluding their study in isolation (McNamara et al. 1987; McNamara & Houston 2008).
Here, we begin with the notion that to maximize energetic returns, animals must simultaneously consider processes happening in both the short and the long term. Dynamic state variable (DSV) models provide a framework in which multiple states and therefore rate processes can be simultaneously incorporated (Houston et al. 1988). In this manner, predictions can be made about the behavioural routines that best compromise between competing physiological and/or environmental processes at multiple timescales. Using a case study of eiders wintering in arctic sea ice, we demonstrate how this framework can help explain diving activity patterns that initially appeared to be counterintuitive. This analysis provides insight into the dynamics of trade-offs among multiple rate processes. Our results indicate that complex and seemingly counterintuitive dynamics can arise from these interactions, and that understanding the adaptive significance of these patterns requires simultaneously considering mechanisms that operate at different rates or timescales in a dynamic framework.
2. Background and study system
Groups of common eiders, Somateria mollissima sedentaria, over-winter in persistent open-water habitats called polynyas that are maintained by strong tidal currents in the sea ice around the Belcher Islands, Hudson Bay, Nunavut. Eiders remained in our study area at Ulutsatuq polynya (56.305° N–78.864° W) throughout the winter (January–March), providing a system essentially closed to local movements (see Heath 2007; Gilchrist & Robertson 2000 for additional details). Their foraging decisions are therefore primarily temporal (when to dive) rather than spatial (where to forage), removing the effects of spatial variation and movements that could influence their behavioural routines. Eiders wintering in the Ulutsatuq polynya primarily foraged on benthic prey (primarily blue mussels, Mytilus edulis) obtained by repeated dives to the bottom (depth 11 m).
Current speed in the polynya fluctuated tidally from near zero at slack tide to a maximum of about 1.4 m s−1 in full flood or ebb. In strong currents eiders rest on the ice edge, almost never diving in currents faster than 1.2 m s−1 (Heath & Gilchrist 2010). This limits time available for foraging to about 250 min of a full 372 min tidal cycle from peak flood to peak ebb. Heath et al. (2007) measured the diving behaviour of eiders wintering at Ulutsatuq polynya using underwater video recordings, and found that the time required to descend to the bottom varied directly with current speed, but the total dive duration did not change. As current speed slowed and travel time decreased, foraging time at depth increased (e.g. by approx. 10 s as currents changed from 1 to 0 m s−1). This indicated that foraging is most energetically profitable in the slack currents during the middle of the foraging interval (Heath et al. 2007; Heath & Gilchrist 2010). Paradoxically, in some cases the diving activity of eiders was greatest at the start and end of the interval during strong currents and when foraging was least profitable. These findings do not support the predictions of classic rate-maximizing dive cycle models (Kramer 1988; Houston & Carbone 1992).
Heath et al. (2007) also found that surface pause durations between successive dives were much longer than anticipated based on physiological recovery alone. They suggested that the rate of digestive processing could constrain the rate of diving. The benthic invertebrates consumed by eiders are swallowed whole along with the shell and processed by grinding in the gizzard, making it plausible that the rate of intake of these bulky prey exceeds the rate at which they can be processed (Guillemette 1994; Kersten & Visser 1996). In this paper we investigate whether and how processes of tidal current change, diving energetics and digestive processing interact to influence patterns of foraging behaviour (Wiener 1992; Guillemette 1994, 1998; Heath et al. 2007). We present this as a quantitative case study of how the interaction between processes with different rates or timescales can have a complex result on the ecological dynamics of an organism.
3. Material and methods
(a). Empirical observations of behavioural patterns
We compiled continuous time series of eider diving behaviour using radio telemetry. Radioed birds remained in the polynya and within range of the receiver for the duration of the study. We affixed small radio telemeters (RI-2B Transmitter, mass 9.0 g, frequency 164.027–164.575 Hz, Holohil Systems Ltd, Carp, Ontario, Canada) to the plumage on the backs of eight individual eiders with super-glue. (Procedures followed guidelines of the Sanikiluaq Hunters and Trappers Association and Environment Canada.) Transmitters remained on the birds for approximately one week. Data collection computers (DCC; ATS Model R4000 Scientific Receiver with model DCC II, Advanced Telemetry Systems, Isanti, MN, USA), measured the number of pulses transmitted by each radio during 2 s, at 12 s intervals. Two or three pulses were received during the 2 s, but no pulses were received during dives. Dives last on average 1 min (Heath et al. 2007; minimum observed dive length was 49 s), and could thus be identified as two or more successive silent intervals. These data were organized into a binomial (dive/rest) time series for each individual, with a grain (resolution) of 1 min. Each observed dive was assigned to the 1 min interval corresponding most closely with the mid-point of the dive. Time series from eight individuals were obtained for the period investigated in this study, a single tidal cycle that occurred during daylight hours on 6 March 2003, which had a strong current profile owing to the influence of spring tides. An Aquadopp current logger deployed at the polynya measured tidal current velocity at 5 min intervals throughout the study (Heath et al. 2006).
(b). Predictions
The strong tidal nature of polynyas at the Belcher Islands creates a temporal foraging environment with periodic access to prey. The costs of foraging vary regularly during these periods, being highest at the start and finish of each tidal cycle when current speed is fastest, and lowest in the middle when current is slack. During peak currents, diving is dangerous and unprofitable, and eiders leave the water and rest on the ice edge (Heath & Gilchrist 2010). We evaluate three potential hypotheses to explain the foraging patterns of diving eiders:
— Eiders are not constrained by either digestion or tidal currents and simply forage in proportion to time available. Given the results of Heath & Gilchrist (2010), we assume that time was only available for foraging when current speed was less than 1.2 m s−1.
— Eiders allocate foraging effort only in relation to the profitability of diving. Based on empirical data from underwater observations and published values of energy consumption rates, Heath & Gilchrist (2010) present an energetic model that estimates the net energy gain per dive cycle as a function of tidal current speed. This hypothesis predicts that foraging effort will be positively correlated with the profitability of diving, which we determined by evaluating the regression equations of Heath & Gilchrist (2010) by the tidal current profile.
— Eiders consider a trade-off between the profitability of diving, a digestive processing constraint and the cyclical nature of the tidal current profile in order to maximize net energy gain over the tidal cycle. Predictions of this hypothesis were evaluated by developing a DSV model (Clarke & Mangel 2000) to predict the diving pattern that maximizes total net energy gain over an entire tidal period.
We tested predictions of each of these hypotheses against the continuous activity records of wild eiders obtained at the polynya using radio telemetry.
(c). Dynamic model
We assumed that fitness is a direct function of the total net energy gained, which is reasonable given that we are considering a relatively short period (a single tide cycle), far away from the breeding season in the middle of the arctic winter, where strong winds (greater than 60 km h−1) and low temperatures (less than −40°C) commonly occur.
We considered two state variables in the analysis, stomach contents z and energy stores x. Stomach contents z could range from 0 to 30 kJ (see §3d), discretized for model implementation into 100 categories. Energy reserves x ranged between 0 and 10 000 kJ, which was well above the range that could be acquired or depleted in a single tidal cycle under even the most intensive foraging regime. This range was discretized into 1000 categories.
The model considered an entire tidal cycle (372 min) in 1 min intervals. For implementation in the model, we used 13 current speed categories, corresponding to currents ranging between 0.1 and 1.3 m s−1 in 0.1 m s−1 bins. In each time interval, eiders could decide to rest on the surface (i = 1); to dive and forage (i = 2); or rest by hauling out onto the ice edge (i = 3). A decision to dive (i = 2) necessitated a surface pause (i = 1) in the following time step. Each behaviour had an associated rate of energy expenditure ei and gain gi, some of which were a function of current speed (table 1). While in the model it was important to consider the energetics of resting on the ice versus water, we were only interested in analysing diving patterns. We therefore considered binomial time series of diving or resting (on water or ice) in further analyses.
Table 1.
Energetic costs e and benefits g (Joules) of the decision to rest or dive (i = 1 or 2, respectively), as a function of current speed c (m s−1) and rates of expenditure (surfacerate, descentrate, bottomrate, ascentrate in Watts for resting on the surface, descent, foraging at depth and ascending, respectively) and gain (intakerate in Watts per time at depth). Equations are based on empirical measurements of diving activities in the wild (Heath et al. 2006, 2007) and on baseline rates of expenditure presented by Heath & Gilchrist (2010). Surfacerate = 27.5 W, descentrate =44.0 W, bottomrate = 30.25 W, ascentrate = 27.5 W and intakerate = 174.0 W.
| equation | |
|---|---|
| resting expenditure | e1(c) = surfacerate·60· (1.016 + 0.7977e−e·10.24(0.9631c)+1) |
| diving expenditure | e2(c) = descent + bottom + ascent |
| descent = descentrate(8.75 + 3.6c + 5.35(c − 0.349)2) | |
| bottom = bottomrate(42.094 − 10.49c2) | |
| ascent = ascentrate(7.37 + 1.91c + 8.33(c − 0.349)2) | |
| intake | g1(c) = 0 |
| g2(c) = intakerate(42.094 − 10.49c2) |
We incorporated a mortality factor for diving, as we had observed birds being swept under the ice in very strong currents (Heath 2007). The probability of mortality m was set to zero for currents less than or equal to 0.8 m s−1. For currents greater than 0.8 m s−1, the risk m(c) was set to equal 0.005, 0.0075, 0.01, 0.05 and 0.1 for current speeds of 0.9, 1.0, 1.1, 1.2 and 1.3 m s−1, respectively. Implementation of this factor did not change the major results of the model, but gave a better fit to the current speed at which eiders stopped foraging and rested on the ice, which was desirable (Heath & Gilchrist 2010).
The digestive rate d was equal to 3 kJ per time step (see §3d). Like other physical exercise, diving entails a redirection of blood flow from internal organs to the limbs (Bevan & Butler 1992), and we therefore assumed that digestive processing did not proceed during a dive and the subsequent pause. Therefore, digestive rate was set to zero during a dive and the subsequent surface pause. If the stomach was full (i.e. z = zmax), additional energy could not be gained by diving.
The dynamic programming equation for the model was defined as
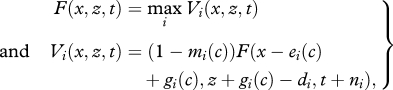 |
3.1 |
where n1 and n3 = 1 (fitness is evaluated at t + 1 for i = 1 (rest) or 3 (rest on ice)), and n2 = 2 (fitness is evaluated at t + 2 for i = 2 (dive)). The terminal fitness condition is a function of total energy at the end of a tide cycle (T = 372 1 min time steps) and is therefore the sum of energy stores x and extractable energy contained in the oesophagus and gizzard (hereafter called ‘stomach’) contents z
| 3.2 |
Using the dynamic programming equation (3.1), the decision that maximizes total net energy gain at time T can be determined for each combination of states z and x. The full decision matrix (i.e. the optimal decision at all possible states in the preceding time steps) is calculated by backwards iteration. The dynamic programming routine was implemented in Matlab v. 6.5 (Mathworks, Inc., Natick, MA, USA). Parameter estimation is described below and equations for energetic costs and benefits are presented in table 1.
The decision matrix was used in a forward iteration to compute the foraging routine that provided maximum net energy gain over the tide cycle (see Clarke & Mangel 2000; Houston & McNamara 1999 for details). We considered the predictions of the model for individuals beginning the period with a medium value of energy stores (x = 5000). The range of x approximated the range in full body stores of wintering eiders, exceeding greatly that which could be lost or gained over a single tidal period (i.e. it therefore never reached its upper or lower limit in the model). To evaluate net energy gain over the tide cycle, the model kept track of total energy stores (state variable x), but we are interested here in the influence of digestive processing on behavioural patterns, rather than the seasonal dynamics of body reserves acquisition and sequestering.
(d). Parameter estimation
We used regression equations based on empirical observations of eiders diving at different current speeds at our field site (Heath et al. 2006, 2007), combined with energetic rate estimates from the literature (Heath & Gilchrist 2010) to estimate both energy expenditure and gain as a function of current speed. Parameter values are summarized in table 1. We used a value of 30 kJ of digestible energy held in the stomach (gizzard plus oesophagus), estimated as follows. Guillemette (1994) found that the gizzard of an eider could contain about 20 g of mussels, or about 12.5 kJ energetic equivalent (accounting for digestive processing costs; Heath & Gilchrist 2010). The eider subspecies at the Belcher Islands is larger than that studied by Guillemette, and we estimated a slightly larger gizzard size could contain up to about 15 kJ of digestible energy from mussels. We doubled this value to account for mussels held in the oesophagus prior to grinding in the gizzard. Energetic costs of grinding mussels in the crop and additional digestive processing were included in our energetic intake rates (Heath & Gilchrist 2010).
Guillemette (1994) also estimated that mussels could be processed at a rate of 2.4–6.1 g min−1. Converted to digestible energy as above (Heath & Gilchrist 2010), this means about 3 kJ min−1 is processed in the gizzard. We used this as a baseline value in the model.
Sensitivity analysis was conducted by varying both digestive rate d and digestive capacity zmax (i.e. maximum stomach contents) by ±50 per cent, both independently and together. Including baseline values, there were nine scenarios considered: both rate and capacity at baseline (1); rate or capacity at baseline, the other at ±50 per cent (2–5); both rate and capacity at either ±50 per cent (6 and 7); and rate at +50 per cent, capacity at −50 per cent and vice versa (8 and 9). While this is an extensive range of sensitivity analysis, our purpose was to demonstrate that results were substantially robust. Consideration of a stochastic intake rate also had no influence on results of the model and was therefore not investigated further here.
(e). Analysis
The entire tide cycle was categorized into time blocks falling into the 13 current speed categories corresponding to currents (0.1–1.3 m s−1). For each current speed category, we calculated the proportion of total time available, the proportion of dives observed for each of the eight individual eiders (empirical data), the profitability (net energy gain rate of a dive cycle) and the proportion of dives predicted (model results). Means and standard deviations were computed across individuals for the empirical data. Error estimates from model baseline values are represented as the standard deviation across all nine sensitivity analysis scenarios.
The empirical results (proportion of dives made by wild eiders in each current speed category) were compared with the distribution expected if diving behaviour (i) matched the time available, (ii) matched the relative profitability of each current speed category, and (iii) matched the predictions of the DSV model. The distributions were compared and deviances (sums of squares) calculated to determine the comparison with the highest goodness of fit (lowest sums of squared differences between model and data). We also report the correlation between the observed and predicted proportion of dives across the 13 current speed categories.
To facilitate graphical presentation of the results, we converted the binomial (dive/rest) time series into a continuous plot of foraging effort by applying a moving average window to the time series (window size = 10% of time series length) and then fitted the most appropriate polynomial curve using the least-squares ‘polyfit’ function in Matlab v. 6.5 (Mathworks, Inc.).
4. Results
The simplest null hypothesis was that eiders did not consider tidal currents or digestion during diving, allocating their foraging effort only in relation to time available (figure 1a). Figure 2a shows the distribution of time available across current speeds in relation to the observed patterns of diving. Across the 13 current speed categories, there was no correlation between the proportion of time available and field observations (proportion of dives; r = −0.126, n = 13, p = 0.681; sums of squares = 309.11). Relative to time available, eiders undermatched fast current speeds and overmatched medium current speeds.
Figure 1.

Predicted allocation of foraging effort (dashed lines) by diving eiders over a strong spring tide current profile (solid lines) in relation to (a) time available, (b) the profitability of diving and (c) the dynamic model that considered the interaction between profitability, digestion and the tide cycle. (d) An example time series of observed foraging effort by a diving eider. Solid circles indicate an individual dive. Dashed lines in (c) and (d) are a graphical representation of foraging effort obtained using a moving average and curve fitting of the binomial (dive/rest) data (§3). As predicted by the dynamic model eiders, concentrated foraging effort at the beginning and end of the foraging period, and reduced effort in slack currents even though this was the most profitable time to dive. This allowed them to finish the foraging period with a full stomach and maximized net energy gain over the entire tide cycle.
Figure 2.

(a) Predictions of the dynamic model and (b) radio telemetry observations of eight individuals in terms of time allocation spent diving versus time available to forage in current velocities ranging from 0.1 to 1.3 m s−1. In both the model and observations, eiders avoided fast currents when foraging was unprofitable. Surprisingly, foraging effort was primarily concentrated during medium currents, and relatively little foraging effort was exhibited during slack currents when it was most profitable to dive. Error bars are standard deviations across sensitivity analysis or individuals, for the model and empirical data, respectively. Black bars, per cent of total dives; grey bars, per cent time available.
Another simple and intuitive hypothesis was that eiders matched foraging effort with the energetic profitability of diving (figure 1b). This model predicts that maximum foraging effort should occur during slack currents, when it takes less time to travel to the bottom and there is more time available to feed on a given dive (see also Heath et al. 2007). Relative to predictions of the profitability model, eiders undermatched slow current speeds and overmatched faster currents, providing weak support for this hypothesis (r = 0.588, n = 13, p = 0.035; sum of squares = 175.91).
The more complex DSV model, which considered the influence of profitability, digestion and the cyclical tide cycle, provided the best fit with the empirical data (r = 0.847, n = 13, p = 0.0003; sums of squares = 111.75). Both the model and empirical observations indicate the avoidance of diving in fast currents, the concentration of diving in currents between about 0.4 and 0.9 m s−1 and lower foraging effort in slack currents (figure 1c,d).
The optimal decisions predicted by the dynamic model (rest in white region; dive in grey region) are presented as a function of stomach content (state variable z) in figure 3a. While the decision matrix is complex and dynamic in some regions, several general features are apparent. First, individuals should dive, regardless of stomach contents, at the beginning and end of the tide cycle, when currents are strongest. Second, individuals dive throughout the tidal cycle across a range of stomach contents. Finally, individuals should always dive if the stomach contents drop below a threshold (about 15 per cent of capacity). Figure 3b illustrates a sample sequence of stomach contents, derived from forward iteration, indicating how it fills and empties throughout the tide cycle. Note that as the end of the foraging period approaches, the model predicts the stomach is maintained fuller, until eiders end the foraging period with a full stomach.
Figure 3.

(a) The decision state space generated by the dynamic model, indicating the optimal decision (white regions indicate resting, while grey regions indicate a decision to dive) as a function of the state of the stomach contents (proportion of capacity) and time across the tidal cycle. (b) Changes in the proportion of the stomach that is full, over the tidal cycle, as predicted by forward iteration through decision state space.
5. Discussion
Eider ducks wintering at polynyas in Hudson Bay reduced their foraging effort during the most profitable time to feed, despite limited time available for foraging during fast tidal current profiles (spring tides). To understand this counterintuitive behaviour, this study considered the interactions among three processes with different timescales that influence diving and foraging. A dynamic model indicated the observed patterns of diving could arise from simple interactions between changing profitability during dive cycles (short term), digestive processing of benthic invertebrate prey (medium term) and the profile of current speeds over a tidal cycle (longer term; figure 1). As a result of these interactions, eiders can obtain the greatest net energy gains by diving intensely at the beginning and end of a tide cycle (when current speed is higher and diving relatively unprofitable), and reducing foraging during slack currents even though this is the most profitable time to dive. Resting and digesting in slack currents allows adequate time to fill up the stomach again before resting on the ice edge during peak tidal currents, when diving is dangerous and rarely profitable.
While digestive constraints have been proposed to limit intake rate (e.g. Zwarts et al. 1996; Guillemette 1998), and to influence the distribution of foraging effort under predation risk (Bednekoff & Houston 1994), this study is unique in explicitly and quantitatively evaluating how digestive bottlenecks can interact with other rate processes, resulting in a complex functional response. By considering multiple physiological and environmental rate processes in a dynamic framework, counterintuitive patterns can be understood as adaptive. The complex foraging routines observed in this study directly emerge from our dynamic model (figure 1). No spatial or temporal differences in ecological factors such as prey availability or predation risk were responsible for this patterning; it emerged solely from a consideration of digestive processing against a backdrop of natural tidal variation and associated changes in the energetics of diving and foraging. Specifically, by resting and digesting in slack currents at the middle of a tide cycle, eiders can put in most effort when their stomach is empty at the beginning of the tide cycle, and can end the tidal cycle and get out of the water during dangerously fast and unprofitable current velocities with a full stomach. These results could not be explained by the simpler hypotheses of matching time available, matching profitability or by the classic dive cycle models.
Existing diving models are based on a static approach that only considers a single rate process in isolation. This approach could not explain the observed patterns of foraging behaviour, instead incorrectly predicting that eiders should concentrate their foraging effort in slack currents (Heath et al. 2007). Static approaches employ classic economic theory (e.g. Charnov 1976), and base their predictions on maximizing a currency such as the rate or efficiency of energy intake. However, these currencies do not necessarily represent longer term fitness (Ydenberg 1998), which is often assumed, but rarely tested. The appropriate currency can depend on whether time or energy constraints are limiting (Ydenberg & Hurd 1998) and as demonstrated here, both time (tide cycle) and energy (digestive bottleneck) constraints can act together in a dynamic manner.
Maximizing rate or efficiency over the long term in the wild can be a complex process. For example, animals cannot run indefinitely at a constant maximum dash speed, and similarly may optimize energy intake at different timescales (Ydenberg 1998; Fortin et al. 2002; Heath et al. 2007). In some situations, long-term rewards may be best achieved by a short-term reduction in effort so that the best strategies involve intermittent bouts of inactivity (Williams et al. 2000; Kramer & McLaughlin 2001; Wienstein 2001; Heath et al. 2008), or more complex temporal structuring of activity patterns (Cole 1994). Investigating behavioural patterns therefore requires simultaneous consideration of the trade-offs that can occur between multiple overlapping physiological and abiotic rate processes in a dynamic multi-scale framework.
Dynamic approaches similar to the one used here have provided significant insights into the routines of individuals, such as those of small birds in winter responding to predation risk (Houston & McNamara 1999; Clarke & Mangel 2000). In several cases, these models have made unexpected predictions, such as how temporal variation in predation risk can lead to counterintuitive food-caching patterns (Pravosudov & Lucas 2001), or that bird song can peak when food availability is highest (McNamara et al. 1987). This study is the first to demonstrate that these counterintuitive effects are not just special theoretical cases, but rather can help explain complex functional responses and ecology of organisms in the wild.
The behavioural ecology approach and dynamic modelling to date have primarily considered trade-offs between ecological factors such as predation risk, habitat, food availability and prey or host selection. The present study focused exclusively on temporal activity patterns, and demonstrates the importance of considering trade-offs between short-term and long-term processes in an otherwise constant ecological context. Behaviour can be understood as the means by which an organism negotiates a coupling between its internal physiological processes and processes in its environment. Particularly when these processes are nonlinear (such as many components of diving activities), it should not be surprising that an understanding of behavioural patterns can vary as a function of the scale of the investigation (Schneider 2001). Dynamic modelling provides an approach in which the grain and extent of observation can be changed and multiple processes occurring at different frequencies can be considered simultaneously. This provides a promising approach for understanding how a variety of complex patterns can emerge from simple processes. For eiders wintering in the arctic, this demonstrated that counterintuitive foraging patterns and a complex functional response could be understood as an adaptive response to a trade-off between short-term energetics in dive cycles, longer term digestive constraints and the cyclical nature of tidal currents. The lessons learned from this research indicate that a multi-scale dynamic approach can help explore the mechanisms underlying behaviour, such as the coupling and coevolution of physiological and abiotic rate processes, the role of behaviour in negotiating these interactions, and provide adaptive explanations for complex and seemingly counterintuitive strategies adopted by individuals in the wild.
Acknowledgements
This project was a collaborative effort between the Canadian Wildlife Service and Science and Technology Branch of Environment Canada, the Center for Wildlife Ecology at Simon Fraser University, the Nunavut Wildlife Management Board and the Community of Sanikiluaq. Particular thanks are due to Lucassie Arragutainaq of the Sanikiluaq Hunters and Trappers Association, and Dwayne Searle of the Mitiq Cooperative Ltd. Fieldwork relied on the knowledge and experience of Inuit hunters, particularly Simeonie Kavik, Elijah Oquaituk and Lucassie Ippaq. We also thank Karel Allard, Joel Bety, Rachael Bryant, Scott Gilliland and Paul Smith who assisted with fieldwork. Research funding was provided by the Canadian Wildlife Service, the Nunavut Research Trust, the Sea Duck Joint Venture, World Wildlife Fund Canada, Simon Fraser University and the Northern Scientific Training Program. Scholarship funding to J.P.H. was provided by the Natural Sciences and Engineering Research Council of Canada, the Center for Wildlife Ecology at Simon Fraser University and the Glen Geen scholarship in marine biology.
References
- Bednekoff P. A., Houston A. I.1994Avian daily foraging patterns: effects of digestive constraints and variability. Evol. Ecol. 8, 36–52 (doi:10.1007/BF01237664) [Google Scholar]
- Bevan R. M., Butler P. J.1992Cardiac output and blood flow distribution during swimming and voluntary diving of the tufted duck (Aythya fuligula). J. Exp. Biol. 168, 199–217 [DOI] [PubMed] [Google Scholar]
- Charnov E. L.1976Optimal foraging: the marginal value theorem. Theor. Popul. Biol. 9, 129–136 (doi:10.1016/0040-5809(76)90040-X) [DOI] [PubMed] [Google Scholar]
- Clarke C. W., Mangel M.2000Dynamic state variable models in ecology: methods and applications. New York, NY: Oxford University Press [Google Scholar]
- Cole B. J.1994Chaos and behavior: the perspective of non-linear dynamics. In Behavioral mechanisms in evolutionary ecology (ed. Real L.), pp. 423–443 Chicago, IL: University of Chicago Press [Google Scholar]
- Fortin D., Fryxell J. M., Pilote R.2002The temporal scale of foraging decisions in bison. Ecology 83, 970–982 (doi:10.1890/0012-9658(2002)083[0970:TTSOFD]2.0.CO;2) [Google Scholar]
- Gilchrist H. G., Robertson G. J.2000Observations of marine birds and mammals wintering at polynyas and ice edges in the Belcher Islands, Nunavut, Canada. Arctic 53, 61–68 [Google Scholar]
- Guillemette M.1994Digestive-rate constraint in wintering common eiders (Somateria mollissima): implications for flying capabilities. Auk 111, 900–909 [Google Scholar]
- Guillemette M.1998The effect of time and digestion constraints in common eiders while feeding and diving over blue mussel beds. Funct. Ecol. 12, 123–131 (doi:10.1046/j.1365-2435.1998.00164.x) [Google Scholar]
- Heath J. P.2007Diving and foraging by common eiders wintering in the Canadian arctic: managing energy at multiple timescales. PhD thesis, Simon Fraser University [Google Scholar]
- Heath J. P., Gilchrist H. G.2010When foraging becomes unprofitable: energetics of diving in tidal currents by common eiders wintering in the arctic. Mar. Ecol. Prog. Ser. 403, 279–290 (doi:10.3354/meps08482) [Google Scholar]
- Heath J. P., Gilchrist H. G., Ydenberg R. C.2006Regulation of stroke patterns and swim speed across a range of current velocities: diving by common eiders wintering in polynyas in the Canadian arctic. J. Exp. Biol. 209, 3974–3983 (doi:10.1242/jeb.02482) [DOI] [PubMed] [Google Scholar]
- Heath J. P., Gilchrist H. G., Ydenberg R. C.2007Can dive cycle models predict patterns of foraging behaviour? Diving by common eiders in an Arctic polynya. Anim. Behav. 73, 877–884 (doi:10.1016/j.anbehav.2006.10.015) [Google Scholar]
- Heath J. P., Montevecchi W. A., Robertson G. J.2008Allocating foraging effort across multiple timescales: behavioural responses to environmental conditions by harlequin ducks wintering at Cape St Mary's, Newfoundland. Waterbirds 31, 71–80 [Google Scholar]
- Houston A. I., Carbone C.1992The optimal allocation of time during the dive cycle. Behav. Ecol. 3, 255–265 (doi:10.1093/beheco/3.3.255) [Google Scholar]
- Houston A. I., McNamara J. M.1999Models of adaptive behaviour: an approach based on state. Cambridge, UK: Cambridge University Press [Google Scholar]
- Houston A. I., Clarke C., McNamara J., Mangel M.1988Dynamic models in behavioural and evolutionary ecology. Nature 332, 29–34 (doi:10.1038/332029a0) [Google Scholar]
- Kersten M., Visser W.1996The rate of food processing in the oystercatcher: food intake and energy expenditure constrained by a digestive bottleneck. Funct. Ecol. 10, 440–448 (doi:10.2307/2389936) [Google Scholar]
- Kramer D. L.1988The behavioural ecology of air breathing by aquatic animals. Can. J. Zool. 66, 89–94 (doi:10.1139/z88-012) [Google Scholar]
- Kramer D. L., Mclaughlin R. L.2001The behavioural ecology of intermittent locomotion. Am. Zool. 41, 137–153 (doi:10.1668/0003-1569(2001)041[0137:TBEOIL]2.0.CO;2) [Google Scholar]
- Levin S. A.1992The problem of pattern and scale in ecology. Ecology 73, 1943–1967 (doi:10.2307/1941447) [Google Scholar]
- McNamara J. M., Houston A. I.2008Optimal annual routines: behaviour in the context of physiology and ecology. Phil. Trans. R. Soc. B 363, 301–319 (doi:10.1098/rstb.2007.2141) [DOI] [PMC free article] [PubMed] [Google Scholar]
- McNamara J. M., Mace R. H., Houston A. I.1987Optimal daily routines of singing and foraging in a bird singing to attract a mate. Behav. Ecol. Sociobiol. 20, 399–405 (doi:10.1007/BF00302982) [Google Scholar]
- Pravosudov V. V., Lucas J. R.2001Daily patterns of energy storage in food-caching birds under variable daily predation risk: a dynamic state variable model. Behav. Ecol. Sociobiol. 50, 239–250 (doi:10.1007/s002650100361) [Google Scholar]
- Schneider D. C.1994Quantitative ecology: spatial and temporal scaling. San Diego, CA: Academic Press [Google Scholar]
- Schneider D. C.2001The rise of the concept of scale in ecology. Bioscience 51, 545–553 (doi:10.1641/0006-3568(2001)051[0545:TROTCO]2.0.CO;2) [Google Scholar]
- Wiener J.1992Physiological limits to sustainable energy budgets in birds and mammals: ecological implications. Trends Ecol. Evol. 7, 384–388 [DOI] [PubMed] [Google Scholar]
- Wiens J. A.1989Spatial scaling in ecology. Funct. Ecol. 3, 385–397 (doi:10.2307/2389612) [Google Scholar]
- Wienstein R. B.2001Terrestrial intermittent locomotion. Am. Zool. 41, 219–228 [Google Scholar]
- Williams T. M., Davis R. W., Fuiman L. A., Francis J., LeBoeuf B. J., Horning M., Calambokids J., Croll D. A.2000Sink or swim: strategies for cost-efficient diving by marine mammals. Science 288, 133–136 (doi:10.1126/science.288.5463.133) [DOI] [PubMed] [Google Scholar]
- Ydenberg R. C.1998Behavioral decisions about foraging and predator avoidance. In Cognitive ecology: the evolutionary ecology of information processing and decision making (ed. Dukas R.), pp. 343–378 Chicago, IL: University of Chicago Press [Google Scholar]
- Ydenberg R. C., Hurd P.1998Simple models of feeding with time and energy constraints. Behav. Ecol. 9, 49–53 (doi:10.1093/beheco/9.1.49) [Google Scholar]
- Zwarts L., Ens B. J., Goss-Custard J. D., Hulscher J. B., Kersten M.1996Why oystercatchers Haematopus ostralegus cannot meet their daily energy requirements in a single low water period. Ardea 84A, 269–290 [Google Scholar]