

Abstract
Adaptation to an environmental stressor is usually studied in isolation, yet these stressors are often encountered in combination in the field, an example being cold and hypoxia at altitude. There has been a paucity of research in this area, although work with rodents indicates that habituation to repeated short cold exposures has a cross-adaptive effect during hypoxia. The present study tested the hypothesis that cross-adaptation is also possible with humans. Thirty-two male volunteers were exposed to 10 min bouts of normoxic and hypoxic (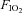 0.12) rest and exercise (100 W on a recumbent cycle ergometer). These were repeated after a 96 h interval, during which participants completed six, 5 min immersions in either cold (12°C, CW) or thermoneutral water (35°C, TW). Venous blood samples were taken immediately after each bout, for determination of catecholamine concentrations. A three-lead ECG was recorded throughout and the final 5 min of each bout was analysed for heart rate variability using fast fourier transformations (and displayed as log transformed data (ln)). In comparison with the first hypoxic exercise exposure, the second exposure of the CW group resulted in an increased ln high frequency (ln HF) power (P < 0.001) and reduced adrenaline (P < 0.001) and noradrenaline concentrations (P < 0.001). Adrenaline and noradrenaline concentrations were lower in the CW group during the second hypoxic exercise compared to the TW group (P = 0.042 and P = 0.003), but ln HF was not. When separated into hypoxic sensitive and hypoxic insensitive subgroups, ln HF was higher in the hypoxic sensitive CW group during the second hypoxic exercise than in any of the other subgroups. Cold habituation reduced the sympathetic response (indicated by the reduced catecholamine concentrations) and elevated the parasympathetic activity (increased ln HF power) to hypoxic exercise. These data suggest a generic autonomic cross-adaptive effect between cold habituation and exposure to acute hypoxia in humans.
0.12) rest and exercise (100 W on a recumbent cycle ergometer). These were repeated after a 96 h interval, during which participants completed six, 5 min immersions in either cold (12°C, CW) or thermoneutral water (35°C, TW). Venous blood samples were taken immediately after each bout, for determination of catecholamine concentrations. A three-lead ECG was recorded throughout and the final 5 min of each bout was analysed for heart rate variability using fast fourier transformations (and displayed as log transformed data (ln)). In comparison with the first hypoxic exercise exposure, the second exposure of the CW group resulted in an increased ln high frequency (ln HF) power (P < 0.001) and reduced adrenaline (P < 0.001) and noradrenaline concentrations (P < 0.001). Adrenaline and noradrenaline concentrations were lower in the CW group during the second hypoxic exercise compared to the TW group (P = 0.042 and P = 0.003), but ln HF was not. When separated into hypoxic sensitive and hypoxic insensitive subgroups, ln HF was higher in the hypoxic sensitive CW group during the second hypoxic exercise than in any of the other subgroups. Cold habituation reduced the sympathetic response (indicated by the reduced catecholamine concentrations) and elevated the parasympathetic activity (increased ln HF power) to hypoxic exercise. These data suggest a generic autonomic cross-adaptive effect between cold habituation and exposure to acute hypoxia in humans.
Introduction
Physiological adaptation to environmental stressors is often studied in isolation, but these stressors are frequently combined outside of laboratory settings, for example cold and hypoxia at altitude. There is limited information about the effect that adaptation to one environment has on exposure to another. Animal experiments suggest that a generic autonomic adaptation provided by short repeated cold-air exposures can result in a reduction in the sympathetic response to novel environmental stimuli, such as acute hypoxia (forming a ‘cross-adaptive’ effect; LeBlanc, 1969). Groups of rats that were not habituated to short cold exposures or lived in consistently cool environments did not demonstrate an improved response to hypoxic conditions. Others have also reported that prolonged moderately cold conditions had no effect (Fregly, 1954) or a detrimental effect on responses to hypoxia (Fregly, 1953; Altland et al. 1972, 1973).
In humans it is known that repeated short cold-water immersions attenuate the initial cardio-respiratory responses to cold immersion (Keatinge & Evans, 1961; Golden & Tipton, 1988) as well as decrease circulating adrenaline and noradrenaline concentrations (Huttunen et al. 2001). However, others have observed a decrease in plasma noradrenaline concentrations, but not adrenaline (Jansky et al. 1996; Leppaluoto et al. 2001). This suggests that repeated immersions in cold-water result in a habituation which involves the sympathetic nervous system pathways. At present, it is not known if this also provides a generic autonomic cross-adaptation to another environmental stressor, such as exposure to acute hypoxia.
Autonomic activity has been assessed non-invasively using heart rate variability (HRV), with high frequency power representing the level of parasympathetic activity (Akselrod et al. 1981; Pomeranz et al. 1985; Pagani et al. 1986) and low frequency power having both sympathetic and parasympathetic components (Akselrod et al. 1981; Pomeranz et al. 1985). The HRV responses to hypoxia have previously been investigated, and several studies suggest that compared to normoxia, an acute hypoxic/altitude exposure reduces the high frequency component at rest (Zuzewicz et al. 1999; Bernardi et al. 2001; Kanai et al. 2001; Buchheit et al. 2004). This effect was also established during exercise in hypoxic and normoxic environments (Zupet et al. 2009), but has not been reported by all (Buchheit et al. 2004). In addition, habituation to repeated short cold-air exposures has been reported to increase high frequency activity (parasympathetic activity) (Westerlund et al. 2006). However, there are no experiments which have investigated the response of HRV to head-out, whole body immersions in cold-water, or used this technique to establish if a cross-adaptive effect occurs between repeated cold water immersions and hypoxia. The variability of the measurement technique (Sandercock, 2007) and a concern about how representative of sympathetic activity the low frequency power is (Berntson et al. 1997) raise questions about the sole use of HRV as an accurate and reliable indicator of autonomic activity. Therefore, a second measurement technique, such as circulating catecholamine concentrations may be used.
Accordingly, the present study tested the hypothesis that a cross-adaptation occurs between the habituation response to repeated short cold-water immersions and exposure to hypoxia in humans.
Methods
Ethical approval
Thirty-two healthy male volunteers gave their written informed consent to participate, and the study received ethical approval from the University of Portsmouth BioSciences research Ethics Committee. None of the subjects had been to significant altitude (>3000 m) or exposed to cold environments in the preceding year. A 12-lead ECG was performed and venous blood drawn for a full blood count screening prior to an assessment by an Independent Medical Officer for the subjects’ fitness to participate in the study. The physical characteristics of the participants are presented in Table 1.
Table 1.
Mean (s.d.) personal characteristics of the cold water (CW) and thermoneutral water (TW) groups separated into hypoxic sensitive (HS) and hypoxic insensitive (HI) subgroups
| Age (years) | Height (m) | Mass (kg) | ∑ 8 skinfolds (mm)* |
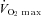 (L min−1) (L min−1) |
FEV1 (L) | FVC (L) | |
|---|---|---|---|---|---|---|---|
| CW (n = 16) | 27 (7) | 1.79 (0.08) | 77.3 (8.4) | 82 (24) | 4.3 (0.7) | 4.4 (0.4) | 5.2 (0.5) |
| CWHS (n = 8) | 27 (7) | 1.80 (0.1) | 75.9 (10.3) | 85 (24) | 4.3 (0.8) | 4.6 (0.5) | 5.3 (0.6) |
| CWHI (n = 8) | 26 (7) | 1.78 (0.06) | 78.5 (5.8) | 79 (22) | 4.3 (0.8) | 4.4 (0.3) | 5.3 (0.4) |
| TW (n = 16) | 22 (3) | 1.80 (0.07) | 76.7 (10.3) | 82 (26) | 4.3 (0.6) | 4.5 (0.7) | 5.5 (0.7) |
| TWHS (n = 8) | 24 (3) | 1.76 (0.06) | 77.5 (10.8) | 80 (11) | 4.1 (0.7) | 4.2 (0.5) | 5.2 (0.5) |
| TWHI (n = 8) | 22 (4) | 1.84 (0.05) | 75.8 (10.5) | 82 (11) | 4.5 (0.6) | 4.9 (0.7) | 5.6 (0.7) |
| All (n = 32) | 25 (6) | 1.79 (0.07) | 76.9 (9.6) | 88 (26) | 4.3 (0.7) | 4.5 (0.6) | 5.3 (0.6) |
Familiarisation
In the fortnight prior to the study, participants attended the laboratory on one occasion and were familiarised with the equipment and procedures. They performed a 10 min period of recumbent cycling (at 100 W) breathing normoxic air, and then breathing an oxygen fraction of 12% (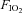 0.12). The cardiac frequency (fC) and minute ventilation (
0.12). The cardiac frequency (fC) and minute ventilation (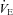 ) results obtained were used to allocate participants to two groups: ‘hypoxic sensitive’ and ‘hypoxic insensitive’. The hypoxic sensitive group had the largest increase in fC and
) results obtained were used to allocate participants to two groups: ‘hypoxic sensitive’ and ‘hypoxic insensitive’. The hypoxic sensitive group had the largest increase in fC and 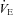 during hypoxic exercise compared to normoxic exercise and the hypoxic insensitive group had smaller fC and
during hypoxic exercise compared to normoxic exercise and the hypoxic insensitive group had smaller fC and 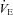 responses. The volunteers from these two groups were then randomly assigned to either the cold-water (CW) or thermoneutral-water control (TW) groups.
responses. The volunteers from these two groups were then randomly assigned to either the cold-water (CW) or thermoneutral-water control (TW) groups.
Experimental outline
All participants performed two sets of normoxic and hypoxic exposures at rest and during exercise; each bout was 10 min in length. A recovery break (breathing normoxic air for 10–20 min) was given between each bout for the participants to recover their fC, 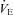 and blood oxygen saturation (
and blood oxygen saturation (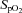 ; estimated from pulse oximetry) to baseline values. All cycling exercise was performed at a cadence of 60 rev min−1.The first and second set of normoxic and hypoxic exposures were separated by 96 h during which participants were immersed into CW or TW six times for a duration of 5 min on each occasion. Two immersions were performed each day, one each morning and afternoon, with a minimum of 4 h between the immersions.
; estimated from pulse oximetry) to baseline values. All cycling exercise was performed at a cadence of 60 rev min−1.The first and second set of normoxic and hypoxic exposures were separated by 96 h during which participants were immersed into CW or TW six times for a duration of 5 min on each occasion. Two immersions were performed each day, one each morning and afternoon, with a minimum of 4 h between the immersions.
Normoxic and hypoxic exposures
Participants had a light meal approximately 3 h prior to the start of the test session. The exposure involved volunteers resting and exercising at 100 W on a recumbent cycle ergometer (Angio, Lode, Groningen, The Netherlands) following a Latin square design, whilst breathing normoxic (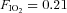 ) and hypoxic (
) and hypoxic (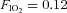 ) gas mixtures in a hypoxic chamber (Sporting Edge, Sherfield on Loddon, UK). Volunteers were instrumented with a three-lead ECG (HME Lifepulse, HME Ltd, Potters Bar, UK), a pulse oximeter finger probe (Nonin 7500, Nonin Medical Inc., Plymouth, MN, USA), respiratory mask and a cannula (18 gauge BD Venflon, Helsingborg, Sweden) was inserted into a prominent vein in the antecubital fossa. Inspired O2 and CO2 values were measured using a portable gas analyser (IBRID MX6, Industrial Scientific Corp., Oakdale, PA, USA), and mixed expired air measurements were collected via a non-rebreathing valve (Hans Rudolph Inc., Shawnee, KS, USA) which was linked to an online O2 and CO2 gas analysis system (GIR 250, Hi-tech Instruments Ltd, Great Marlings, UK) for the measurement of
) gas mixtures in a hypoxic chamber (Sporting Edge, Sherfield on Loddon, UK). Volunteers were instrumented with a three-lead ECG (HME Lifepulse, HME Ltd, Potters Bar, UK), a pulse oximeter finger probe (Nonin 7500, Nonin Medical Inc., Plymouth, MN, USA), respiratory mask and a cannula (18 gauge BD Venflon, Helsingborg, Sweden) was inserted into a prominent vein in the antecubital fossa. Inspired O2 and CO2 values were measured using a portable gas analyser (IBRID MX6, Industrial Scientific Corp., Oakdale, PA, USA), and mixed expired air measurements were collected via a non-rebreathing valve (Hans Rudolph Inc., Shawnee, KS, USA) which was linked to an online O2 and CO2 gas analysis system (GIR 250, Hi-tech Instruments Ltd, Great Marlings, UK) for the measurement of 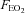 and
and 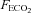 . The volume of each expired breath (VT), was measured using a flow turbine. Expired air temperatures were recorded continuously during all experiments using thermistors placed in the expiratory loop (ADInstruments, Sydney, Australia). Data were recorded using a Powerlab data acquisition system (ADInstruments), and time sequenced using Chart Pro (ADInstruments).
. The volume of each expired breath (VT), was measured using a flow turbine. Expired air temperatures were recorded continuously during all experiments using thermistors placed in the expiratory loop (ADInstruments, Sydney, Australia). Data were recorded using a Powerlab data acquisition system (ADInstruments), and time sequenced using Chart Pro (ADInstruments).
Following instrumentation, participants remained in a seated position for 20 min to allow recovery. At the end of this period, a baseline blood sample was taken via the cannula. Further blood samples were taken at the end of each period of rest and exercise for the determination of haematocrit (Hct), haemoglobin concentration ([Hb]) and catecholamine concentrations. Participants performed the test at the same time of day on both occasions.
Barometric pressure was recorded from a Fortins Mercury Barometer (Russell Scientific Instruments, Dereham, UK) immediately before each experimental session. The mean ±s.d. ambient air temperature in the laboratory used during this part of the study was 20.4 ± 0.7°C with 53.6 ± 2.8% relative humidity.
Upon completion of each hypoxic test, the participants answered a hypoxia symptoms questionnaire similar to that described by Evetts et al. (2005).
Water immersions
The first water immersions took place on the second experimental day, after the first normoxic and hypoxic exposures. On arrival at the laboratory, participants changed into swimming shorts and were instrumented with a three-lead ECG and mouthpiece, connected to a flow turbine (KL Engineering, CA, USA) via a respiratory hose for the measurement of inspiratory volume (VI). Each participant undertook six 5 min head-out seated immersions in stirred water. The TW group were immersed in mean ±s.d. water temperature of 34.7 ± 0.4°C and the CW group in 12.1 ± 0.1°C. At the start of the water immersion the participant was seated in a chair and suspended above the surface of the water for 30 s before being lowered into the water at 8 m min−1 to the level of the axilla. The mean ±s.d. ambient temperature of the immersion laboratory was 25.1 ± 1.1°C with 72.6 ± 6.8% relative humidity.
Blood analyses
Venous blood samples were collected into heparinised vacutainer tubes (Vacutainer Vacuette, Greiner Bio-One GmbH, Kremsmuenster, Austria). Plasma volume changes were calculated from measurements of Hct and [Hb] following the method of Strauss et al. (1951) and Dill & Costill (1974). [Hb] was assessed using a HemoCue (HemoCue AB, Helsingborg, Denmark). Duplicate aliquots of blood were expressed into heparinised micro-centrifuge tubes and spun at 15,115×g for 5 min in a microcentrifuge (Heraeus Pico 17, Thermo Electron Corp., Karlsruhe, Germany) before visual examination of the Hct was made using a micro-Hct reader (Hawsley, Lancing, UK).
The heparin tubes were placed in a refrigerated centrifuge (Heraeus multifuge 3 S-R, Thermo Electron Corp.) at 4°C and spun for 10 min at 812×g. The resultant plasma was removed and frozen at −80°C (snap frozen within 20 min of blood sampling).
Blood plasma catecholamine concentrations were measured from four participants in each group by high performance liquid chromatography with electrochemical detection (HPLC-ECD) at King's College Pathology Laboratory, London. The intra-assay c.v. was 7.2% and inter-assay c.v. was 9.1%.
HRV data analysis
The R-R intervals were calculated from the recorded ECG traces over the final 5 min period of each normoxic and hypoxic exposure. The R-R intervals were recorded to an accuracy of 1 ms. All R-R intervals were visually inspected to exclude premature beats and noise.
Once signal processing was completed, time and frequency domain parameters were calculated using KubiosHRV analysis Software (Biomedical Signal Analysis Group, University of Kuopio, Finland). HRV was calculated in accordance with the recommendations of The Task Force of the European Society of Cardiology and the North American Society of Pacing Electrophysiology (1996). The frequency domain HRV measurements were calculated using fast Fourier transformations and Welch's periodogram method with 256 point window width and a 50% overlap. The time domain measurements include mean fC, mean R-R interval and the standard deviation of the R-R intervals (SDNN). Frequency domain measurements of total power, low frequency power (LF) (range 0.04–0.15 Hz), high frequency power (HF) (range 0.15–0.4 Hz) and low:high frequency ratio (LF:HF ratio) were calculated from absolute values (ms2) and presented in log transformed terms using the natural logarithm (ln).
Statistical analyses
A mixed model ANOVA was conducted to assess the impact of the repeated CW intervention on participants’ responses to hypoxia at rest and during moderate intensity recumbent cycling. This was used for all variables except the catecholamine concentrations, which were analysed using a non-parametric Mann–Whitney U test and Bonferroni's adjustment applied for multiple comparisons, due to the sample size (n = 4 in each group) on which the analysis was performed. Catecholamine analysis was performed on two randomly selected hypoxic sensitive and two hypoxic insensitive participants in each of the TW and CW groups. Spearman's rank order correlation coefficient was also used to assess the relationship in the CW group between the change in fC from the first to the final immersion, and the change in fC from the first to final hypoxic exercise exposure.
Results
All volunteers underwent six water immersions, between the two normoxic and hypoxic exposures; however one participant from the TW and CW groups was removed early from one immersion due to an excessive number of ectopic heart beats.
There were no differences in any variables when normoxic and hypoxic rest periods were compared before or after the CW or TW immersions.
Response to hypoxia and normoxia during exercise
During the first hypoxic exercise condition compared to the normoxic exercise (Table 2), there was a significant reduction in R-R intervals (P < 0.001), ln Total power (P < 0.001), ln HF power (P < 0.001), ln LF power (P < 0.001), ln LF:HF power ratio (P = 0.011) and 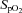 (P < 0.001), and an increase in fC (P < 0.001), and adrenaline (P < 0.001) and noradrenaline concentrations (P < 0.001). The following were significantly increased: fR (P = 0.019), VT (P < 0.001),
(P < 0.001), and an increase in fC (P < 0.001), and adrenaline (P < 0.001) and noradrenaline concentrations (P < 0.001). The following were significantly increased: fR (P = 0.019), VT (P < 0.001), 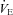 (P < 0.001),
(P < 0.001), 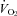 (P < 0.001),
(P < 0.001), 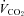 (P = 0.003) and RER (P < 0.001). No differences in the SDNN were found in either group when normoxic and hypoxic exercise was compared.
(P = 0.003) and RER (P < 0.001). No differences in the SDNN were found in either group when normoxic and hypoxic exercise was compared.
Table 2.
Mean (s.d.) HRV indices and ventilatory variables during cycling exercise (100 W) whilst breathing hypoxic gas (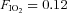 ) before (first exposure) and after (second exposure) the water immersions in thermoneutral (TW) or cold-water (CW)
) before (first exposure) and after (second exposure) the water immersions in thermoneutral (TW) or cold-water (CW)
| CW group | TW group | |||
|---|---|---|---|---|
| First exposure | Second exposure | First exposure | Second exposure | |
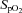 (%) (%) |
74.4 (4.3) | 72.1 (4.6) | 75.1 (3.7) | 73.9 (4.9) |
| fC (beats min−1) | 142 (13) | 134 (13)*† | 141 (12) | 144 (9) |
| R-R intervals (ms) | 426 (40) | 447 (47)*† | 428 (38) | 417 (27) |
| SDNN (ms) | 12 (11) | 9 (5) | 16 (20) | 14 (11) |
| ln total power | 4.0 (1.4) | 4.0 (1.3) | 4.1 (1.3) | 3.5 (0.8) |
| ln HF | 1.3 (1.1) | 2.4 (1.2)* | 1.5 (1.4) | 1.6 (1.3) |
| ln LF | 2.3 (1.5) | 1.5 (0.9) | 2.4 (1.4) | 2.1 (1.4) |
| ln LF:HF | 1.4 (0.6) | 0.7 (0.3)*† | 1.4 (0.7) | 1.2 (0.5) |
| VT (L) | 2.3 (0.2) | 2.6 (0.3)*† | 2.3 (0.2) | 2.4 (0.3) |
| fR (breaths min−1) | 27 (6) | 22 (5)*† | 27 (4) | 26 (4) |
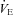 (L·min−1) (L·min−1) |
50.9 (7.1) | 47.2 (3.9)*† | 50.1 (6.2) | 50.5 (6.2) |
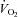 (L·min−1) (L·min−1) |
1.8 (0.3) | 1.7 (0.2)† | 1.9 (0.4) | 2.0 (0.3) |
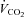 (L·min−1) (L·min−1) |
2.3 (0.2) | 1.8 (0.3)*† | 2.2 (0.3) | 2.3 (0.3) |
| RER | 1.2 (0.1) | 1.1 (0.1)*† | 1.3 (0.1) | 1.2 (0.1) |
Different from the first exposure P < 0.05.
Different from the TW group P < 0.05.
There were no differences in any variables during normoxic exercise before and after the immersions in either the CW or TW group. In the TW group, no differences were found in any of the measured variables between the two hypoxic exercise exposures.
In the CW group the second hypoxic exercise exposure resulted in a significant increase in R-R interval (P = 0.004) and ln HF power (P < 0.001), and reductions in fC (P = 0.002), ln LF:HF power ratio (P = 0.003), adrenaline (P = 0.042, Fig. 1) and noradrenaline concentrations (P = 0.003, Fig. 2), 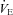 (P = 0.016), and
(P = 0.016), and 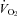 (P = 0.001, Table 2). No differences were observed in ln LF power, ln total power, SDNN and
(P = 0.001, Table 2). No differences were observed in ln LF power, ln total power, SDNN and 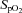 between the hypoxic exercise exposures in the CW group before and after the cold habituation.
between the hypoxic exercise exposures in the CW group before and after the cold habituation.
Figure 1. Adrenaline concentrations during 100 W of hypoxic cycling.

Mean (dotted line) and individual data (continuous lines) for plasma volume corrected adrenaline concentrations during 100 W of hypoxic cycling before and after the repeated cold water (n = 4) or thermoneutral water immersions (n = 4). Mean (s.d.) for the normoxic rest (filled diamonds), exercise (filled squares) and hypoxic rest (filled triangles) conditions are also shown. *Difference between hypoxic exposure 1 and 2 (P < 0.05); †difference between groups (P < 0.05).
Figure 2. Noradrenaline concentrations during 100 W of hypoxic cycling.
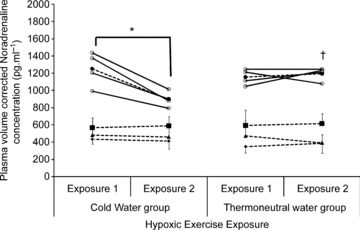
Mean (dotted line) and individual data (continuous lines) for plasma volume corrected noradrenaline concentrations during 100 W of hypoxic cycling before and after the repeated cold water (n = 4) or thermoneutral water immersions (n = 4). See Fig. 1 for further details.
During the second hypoxic exercise exposure, the ln HF power was not significantly different between the CW and TW groups, but R-R intervals were higher (P = 0.041) and fC, ln LF:HF power ratio (Table 2), adrenaline and noradrenaline concentrations (Figs 1 and 2), and  (Table 2) were significantly lower in the CW compared to the TW groups (P = 0.045, 0.002, 0.004, 0.002 and 0.003, respectively).
(Table 2) were significantly lower in the CW compared to the TW groups (P = 0.045, 0.002, 0.004, 0.002 and 0.003, respectively).
Sensitivity to hypoxia
During the hypoxic exposures, there were no differences in any variables in the TW hypoxic sensitive and insensitive subgroups, before or after the immersion in thermoneutral water. After exposure to the repeated cold-water immersions, the CW hypoxic sensitive subgroup had a significant increase in the ln HF power compared to the CW hypoxic insensitive (P = 0.045), TW hypoxic sensitive (P = 0.003) and TW hypoxic insensitive (P = 0.001) subgroups (Fig. 3).
Figure 3. High frequency power between the first and second hypoxic exercise exposures.
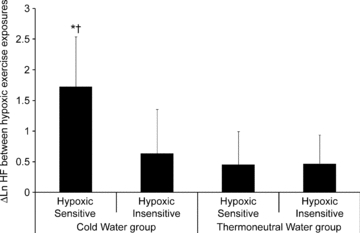
Mean (s.d.) change in log transformed high frequency power between the first and second hypoxic exercise exposures when the cold water and thermoneutral water groups have been separated into the hypoxic sensitive and hypoxic insensitive subgroups. *Different from other subgroups (P < 0.05).
Those in the hypoxic sensitive CW group had the greatest reduction in peak fC between the first and final cold-water immersions and the largest reduction in peak fc between the two hypoxic exercise exposures (r = 0.59, P = 0.017, Fig. 4).
Figure 4.
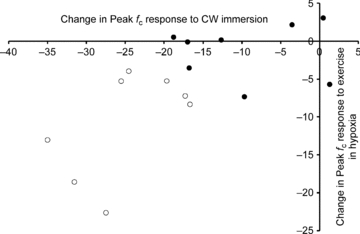
Change in peak cardiac frequency (beats·min−1) from the first to the final cold-water immersions against the change in peak cardiac frequency to hypoxic exercise separated by those in the hypoxic sensitive group (open circles) and hypoxia insensitive group (filled circles).
Subjective responses to hypoxia
The number of hypoxic symptoms and symptom severity did not differ between the CW and TW groups during the first hypoxic exposure. During the second hypoxic exposure, after cold habituation the CW group had a significant reduction in the number of hypoxic symptoms (P = 0.035) and symptom severity (P = 0.016) compared to the first exposure, and in comparison to the second exposure in the TW group (P = 0.042, P = 0.013, number and severity of symptoms respectively, Figs 5 and 6). No additional differences were found when the groups were separated into the hypoxic sensitive and hypoxic insensitive subgroups.
Figure 5. Number of symptoms during the hypoxic exposures.

Mean (s.d.) number of symptoms during the hypoxic exposures for participants in the thermoneutral water immersion group (black bars) and cold water immersion (white bars). †Different from first hypoxic exposure (P < 0.05); *difference between CW and TW (P < 0.05).
Figure 6. Symptom scores during the hypoxic exposures.
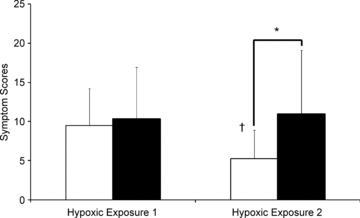
Mean (s.d.) symptom scores during the hypoxic exposures for participants in the thermoneutral water immersion group (black bars) and cold water immersion (white bars). †Different from first hypoxic exposure (P < 0.05); *difference between CW and TW (P < 0.05).
Discussion
The present study found that habituation to cold-water immersion reduced the circulating catecholamine concentrations and increased the HF power of HRV indices during acute hypoxia when exercising. These findings suggest that cold habituation reduced sympathetic stimulation and increased parasympathetic activity when exercising during subsequent hypoxic exposures. The environmental cross-adaptation resulted in reduced physiological strain (an alteration in autonomic balance and reduced cardio-respiratory responses) during exercise in hypoxic conditions. The hypothesis that cold habituation has a cross-adaptive effect on the response to acute hypoxia when exercising is accepted.
The results of the present study support the work of LeBlanc (1969) with rodents, and extend it to human participants. However, these experimental results are in contrast to the findings of Fregly (1953, 1954) and Altland et al. (1973) who exposed rodents to prolonged moderately cold air exposures before the addition of a hypoxic environment. A similar cross-sensitisation response to restraint stress was also described in rats that were exposed to intermittent hypoxia (Ma et al. 2008). The effects reported during hypoxic exposures by Fregly (1953, 1954) and Altland et al. (1973) may be the result of a specific metabolic habituation to the cold, which increases oxygen consumption to defend internal body temperature. In an oxygen depleted environment, the increased metabolic rate results in the rats’ reduced tolerance. In contrast, in the present study, cardio-respiratory and autonomic responses to hypoxia were reduced following the cold-water immersions. The type of cold exposure used in the present study and that of LeBlanc (1969) resulted in a habituation to short repeated cold-water immersions. This is not a metabolic response to prolonged cold exposure, but an attenuation of the sympathetic autonomic response to a short severe repeated stressor (short water immersions). This suggests that the cross adaptation between cold habituation and hypoxic conditions observed in the present study may be mediated by a generic habituation to a stressor rather than a specific response to long-term cold exposure. However, the duration of the cross-adaptation effect has not been established, or whether it will hinder more specific physiological changes which result in an adaptive response to altitude and hypoxia. In addition, it has not been established if reverse cross-adaptation with repeated hypoxic exposures would affect the response to cold; this may require a hypoxic stimulus which would elicit a response of the magnitude observed during initial immersion in cold water.
Sensitivity to hypoxia
The elevated ln HF power in the CW hypoxic sensitive group and the reduction in catecholamine concentrations during the second hypoxic exercise suggests that autonomic balance may have shifted away from sympathetic predominance to include more parasympathetic activity. This suggests that those with the greatest change in cardio-respiratory variables and autonomic balance when cold habituated also have the larger reductions during exposure to acute hypoxic conditions. However, identification of these hypoxic-sensitive individuals is not straightforward as there were no discernable differences in their physical characteristics including 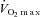 , in comparison with the other subgroups (Table 1).
, in comparison with the other subgroups (Table 1).
The catecholamine results suggest that reductions in circulating volumes occurred in the CW group during the second hypoxic exercise exposure, even though two of the participants were in the hypoxic insensitive group. This suggests that a reduction in sympathetic activity occurred in both the hypoxic sensitive and hypoxic insensitive participants in the CW group, but HRV may not have been sensitive enough to detect the change. This adds further weight to the argument that the large variability in the HRV data may have prevented a significant increase in ln HF power from being observed in the CW group as a whole, compared to the TW group, during the second hypoxic exposure. In addition, the reduction in ventilatory drive during the second hypoxic exposure in the CW group may account for some of the autonomic changes. Elevated inspiratory activity inhibits the cardiac vagal tone (Brown et al. 1993). Consequently, the reduction in fR of the CW group during the second hypoxic exercise exposure may result in an increase in HF power. As a result, changes in HRV indices may not reflect alterations in autonomic balance. The combination of the two measurement techniques (HRV and plasma catecholamines concentrations) was justified, to indicate and corroborate the balance of autonomic activity which occurred during the experiment.
It is concluded that repeated cold-water immersions resulted in a reduction in the sympathetic autonomic and cardio-respiratory responses during moderate exercise in hypoxic conditions. This response may provide a beneficial generic autonomic cross-adaptive effect for some individuals who are exposed acutely to hypoxia. The duration of this adaptation when exposed to hypoxia is not known, but warrants further investigation.
Acknowledgments
The catecholamine samples were analysed by the Clinical Biochemistry Department of King's College Hospital NHS Foundation Trust and technical assistance was provided by Mr Geoff Long and Miss Nicola Ferguson. The Independent Medical Officer for this study was Dr Daniel Roiz de Sa. The work was funded by an Institute of Biomedical and Biomolecular Sciences student bursary.
Glossary
Abbreviations
- CW
cold water
- HRV
heart rate variability
- TW
thermoneutral water
Author contributions
All authors contributed to conception and design of experiments, collection analysis and interpretation of data, and drafting the article or revising it critically. All authors have approved the final version for publication.
References
- Akselrod S, Gordon D, Ubel F, Shannon D, Berger A, Cohen R. Power spectrum analysis of heart rate fluctuations: a quantitative probe of beat-to-beat cardiovascular control. Science. 1981;213:220–222. doi: 10.1126/science.6166045. [DOI] [PubMed] [Google Scholar]
- Altland P, Highman B, Dieter M. Reduced hypoxic tolerance of cold-acclimated rats: serum enzyme and tissue changes. Am J Physiol. 1972;222:1441–1445. doi: 10.1152/ajplegacy.1972.222.6.1441. [DOI] [PubMed] [Google Scholar]
- Altland PD, Highman B, Sellner RG. Tolerance of cold – acclimated and unacclimated rats to hypoxia at 1.7°C. Int J Biometerol. 1973;17:59–71. doi: 10.1007/BF01553646. [DOI] [PubMed] [Google Scholar]
- Bernardi L, Passino C, Wilmerding V, Dallam G, Parker D, Robergs R, Appenzeller O. Breathing patterns and cardiovascular autonomic modulation during hypoxia induced by simulated altitude. J Hypertens. 2001;19:947–958. doi: 10.1097/00004872-200105000-00016. [DOI] [PubMed] [Google Scholar]
- Berntson G, Bigger T, Eckberg DL, Grossman P, Kaufmann P, Malik M, Nagaraja H, Porges S, Saul P, Stone P, Van Der Molen M. Heart rate variability: origins, methods and interpretive caveats. Psychophysiology. 1997;34:623–648. doi: 10.1111/j.1469-8986.1997.tb02140.x. [DOI] [PubMed] [Google Scholar]
- Brown T, Beightol L, Koh J, Eckberg DL. Important influence of respiration on human R-R interval power spectra is largely ignored. J Appl Physiol. 1993;75:2310–2317. doi: 10.1152/jappl.1993.75.5.2310. [DOI] [PubMed] [Google Scholar]
- Buchheit M, Richard R, Doutreleau S, Lonsdorfer-Wolf E, Brandenberger G, Simon C. Effect of acute hypoxia on heart rate variability at rest and during exercise. Int J Sports Med. 2004;25:264–269. doi: 10.1055/s-2004-819938. [DOI] [PubMed] [Google Scholar]
- Dill DB, Costill DL. Calculation of percentage changes in volumes of blood, plasma, and red cells in dehydration. J Appl Physiol. 1974;37:247–248. doi: 10.1152/jappl.1974.37.2.247. [DOI] [PubMed] [Google Scholar]
- Evetts G, Hartley A, Keane S, Keegan J, Simpson A, Taylor A, Hurley A, Ernsting J. SAFE (Europe) Henlow and Cardiff, UK: Kings College Aerospace Medicine Group, London and Royal Air Force Centre of Aviation Medicine; 2005. A comparison of acute hypoxia induced by low concentrations of oxygen at ground level, 10,000 feet and by air at 25,000 feet. Implications for military aircrew training. [Google Scholar]
- Fregly M. Minimal exposures needed to acclimatize rats to cold. Am J Physiol. 1953;173:393–402. doi: 10.1152/ajplegacy.1953.173.3.393. [DOI] [PubMed] [Google Scholar]
- Fregly M. Cross-acclimation between cold and altitude in rats. Am J Physiol. 1954;176:267–273. doi: 10.1152/ajplegacy.1954.176.2.267. [DOI] [PubMed] [Google Scholar]
- Golden F, Tipton M. Human adaptation to repeated cold immersions. J Physiol. 1988;396:349–363. doi: 10.1113/jphysiol.1988.sp016965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huttunen P, Rintamaki H, Hirvonen J. Effect of regular winter swimming on the activity of the sympathoadrenal system before and after a single cold water immersion. Int J Circumpolar Health. 2001;60:400–406. [PubMed] [Google Scholar]
- Jansky L, Sramek P, Savlikova J, Ulicny B, Janakova H, Horky K. Change in sympathetic activity, cardiovascular functions and plasma hormone concentrations due to cold water immersion in men. Eur J Appl Physiol Occup Physiol. 1996;74:148–152. doi: 10.1007/BF00376507. [DOI] [PubMed] [Google Scholar]
- Kanai M, Nishihara F, Shiga T, Shimada H, Saito S. Alterations in autonomic nervous control of heart rate amoung tourists at 2700 and 3700 m above sea level. Wilderness Environ Med. 2001;12:8–12. doi: 10.1580/1080-6032(2001)012[0008:aianco]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- Keatinge WR, Evans M. The respiratory and cardiovascular response to immersion in cold and warm water. Q J Exp Physiol. 1961;46:83–94. doi: 10.1113/expphysiol.1961.sp001519. [DOI] [PubMed] [Google Scholar]
- LeBlanc J. Stress and interstress adaptation. Fed Proc. 1969;28:996–1000. [PubMed] [Google Scholar]
- Leppaluoto J, Korhonen I, Hassi J. Habituation of thermal sensations, skin temperatures and norepinephrine in men exposed to cold air. J Appl Physiol. 2001;90:1211–1218. doi: 10.1152/jappl.2001.90.4.1211. [DOI] [PubMed] [Google Scholar]
- Ma S, Mifflin SW, Cunningham JT, Morilak DA. Chronic intermittent hypoxia sensitizes acute hypothalamic-pituitary-adrenal stress reactivity and Fos induction in the rat locus coeruleus in response to subsequent immobilization stress. Neuroscience. 2008;154:1639–1647. doi: 10.1016/j.neuroscience.2008.04.068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norton K, Olds T. UNSW Press. Australia: 1996. Anthropometrica: a textbook of body measurements for sports and health courses. [Google Scholar]
- Pagani M, Lombardi F, Guzzetti S, Rimoldi O, Furlan R, Pizzinelli P, Sandrone G, Malfatto G, Dell’Orto S, Piccaluga E. Power spectral analysis of heart rate and arterial pressure variabilities as a marker of sympatho-vagal interaction in man and conscious dog. Circ Res. 1986;59:178–193. doi: 10.1161/01.res.59.2.178. [DOI] [PubMed] [Google Scholar]
- Pomeranz B, Macaulay R, Caudill M, Kutz I, Adam D, Gordon D, Kilborn K, Barger A, Shannon D, Cohen R, Benson H. Assessment of autonomic function in humans by heart rate spectral analysis. Am J Physiol Heart Circ Physiol. 1985;248:H151–H153. doi: 10.1152/ajpheart.1985.248.1.H151. [DOI] [PubMed] [Google Scholar]
- Sandercock G. Normative values, reliability and sample size estimates in heart rate variability. Clin Sci. 2007;113:129–130. doi: 10.1042/CS20070137. [DOI] [PubMed] [Google Scholar]
- Strauss M, Davis R, Resenbaum J, Rossmeisl E. “Water diuresis” produced during recumbency by the intravenous infusion of isotonic saline solution. J Clin Invest. 1951;30:862–868. doi: 10.1172/JCI102501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- The Task Force of the European Society of Cardiology and the North American Society of Pacing Electrophysiology. Heart rate variability: standards of measurement, physiological interpretation and clinical use. Circulation. 1996;93:1043–1065. [PubMed] [Google Scholar]
- Westerlund T, Uusitalo A, Smolander J, Mikkelsson M. Heart rate variability in women exposed to very cold air (−110°C) during whole-body cryotherapy. J Thermal Biol. 2006;31:342–346. [Google Scholar]
- Zupet P, Princi T, Finderle Z. Effect of hypobaric hypoxia on heart rate variability during exercise: a pilot field study. Eur J Appl Physiol. 2009;107:345–350. doi: 10.1007/s00421-009-1123-5. [DOI] [PubMed] [Google Scholar]
- Zuzewicz K, Biernat B, Kempa G, Kwarecki K. Heart rate variability in exposure to high altitude hypoxia of short duration. Int J Occup Saf Ergon. 1999;5:337–346. doi: 10.1080/10803548.1999.11076424. [DOI] [PubMed] [Google Scholar]