

Abstract
Both the humans as well as the quadrupedal spine have been shown to exhibit a pattern of pre-existent rotation that is similar in direction to what is found in the most common types of idiopathic scoliosis. It has been postulated that human bipedalism introduces forces to the spine that increase a tendency of the vertebrae to rotate. The objective of this study was to examine the effect of body position on vertebral rotation in vivo. Thirty asymptomatic volunteers underwent magnetic resonance imaging scanning of the spine (T2–L5) in three different body positions; upright, quadrupedal-like (on hands-and-knees) and supine. Vertebral rotation in the local transverse plane was measured according to a pre-established method and compared at different spinal levels between the three body positions. It was shown that in all three positions the mid- and lower thoracic vertebrae were predominantly rotated to the right. However, vertebral rotation was significantly less in the quadrupedal position than in both the standing upright and supine positions.
Keywords: Scoliosis, Etiology, Human bipedalism, Upright MRI, Vertebral rotation, Dorsally directed shear loads
Introduction
Idiopathic scoliosis (IS) is a complex three-dimensional deformity of the spine in which vertebral rotation plays an important role [1]. Spinal biomechanics are different in man than in other vertebrates and may play a role in the initiation and/or progression of this rotatory decompensation of the spine. The normal, non-scoliotic spine in man as well as in quadrupedal animals has been shown to also exhibit a rotational pattern that is similar in direction, but much less in magnitude to what is commonly found in IS [2]. We postulate that the human upright orientation of the spine, when compared with the quadrupedal orientation, plays a role in the expression of rotational patterns of certain spinal segments. In the present MRI study, we analyzed the effect of three different body positions and thus the orientation of the spine in space on the pre-existent rotation of the normal, non-scoliotic human spine in vivo. Vertebral rotation was measured according to a previously established method on MRI images of the spine in non-scoliotic human volunteers, acquired in the upright, “quadrupedal-like” (on hands-and-knees) and supine position.
Materials and methods
Study population and MRI technique
Thirty asymptomatic volunteers (15 males, 15 females) enrolled in this study. The women had a mean age of 23 years (range 19–27), mean height of 170 cm (range 160–180), mean weight of 65 kg (range 54–85) and a mean body mass index of 22.2 kg/m2 (range 19.0–27.4). The male volunteers had a mean age of 23 years (range 18–29), mean height of 180 cm (range 170–195), mean weight of 77 kg (range 67–100) and a mean body mass index of 23.3 kg/m2 (range 21.0–26.6). Exclusion criteria were a history of any spinal pathology, spinal or pelvic abnormalities or any contraindication for MRI. The study was approved by the Institutional Review Board and all participants provided written informed consent before participating in the study.
MRI scans of the spine of each volunteer were acquired in three different positions: (1) bipedal (standing fully upright), (2) supine, and (3) quadrupedal-like (on hands-and-knees) (Fig. 1). A 5° backward tilt, as is customary in this type of MRI, was kept in the standing upright position to improve standing stability, since volunteers had to maintain the same position for approximately 15 min. In the quadrupedal position, MRI compatible padding was provided under hand and knees so as to position the trunk horizontally (Fig. 1c).
Fig. 1.

Three positions in the upright open MRI scanner: a standing upright, b supine, c “quadrupedal” on hands-and-knees and d the quadrupedal position outside the scanner when seen from the side
All images were acquired on a 0.6-T upright open MRI unit (Upright Multi-Position MRI, Fonar Corporation, Melville, NY). First, multi-stack spin echo survey scans were acquired [repetition time (TR) 196 ms, echo time (TE) 15 ms, number of excitations (NEX) 1, slice thickness 9 mm, matrix 256 × 256, field of view (FOV) 480 × 480 mm2]. The survey images in the coronal and sagittal planes of the spine were then used for planning sixteen (L5–T2) axial T1-weighted fast-spin echo (FSE) multi-stack scans, on which the vertebral rotation measurements were performed. (TR 488 ms, TE 17 ms, NEX 2, slice thickness 10 mm, matrix 300 × 300, FOV 300 × 300 mm2).
Each stack consisted of a single slice of 10-mm thickness that was planned passing through the center of each vertebral body and pedicles. The scan planes could be planned ‘truly axially’ by orienting the slices perpendicular to the local axis of vertebral rotation both on the coronal and on the sagittal survey images, as is illustrated in Fig. 2. A relatively thick slice thickness of 10 mm was used to ensure an adequate signal-to-noise ratio. Planning of the slices always started caudal at L5 and proceeded in the cranial direction, when possible as far as T2. Quadrature receiver coils were used; a Quad TL thorax coil for the standing and supine position, and a Quad Planar coil for the quadrupedal position since the subject’s trunk was not on the scanner table. As time was needed for the acquisition of the survey scan plus slice planning, each position was sustained for at least 10 min prior to actual scanning. Thus, we assume that the body had sufficient time to adjust to each new position before the scans were made.
Fig. 2.

Planning of the scan planes on the coronal and sagittal survey scans. By orienting the slices perpendicular to the local axis of vertebral rotation both on the coronal and on the sagittal survey images, the scan planes could be planned ‘truly axially’
Measurements
We used the same definition for vertebral rotation as we did in our previous CT and MRI studies in humans and dogs [2–4]. Vertebral rotation was defined as the angle between the longitudinal axis of the vertebra and the mid-sagittal axis of the trunk. The longitudinal axis of the vertebra was defined as the line passing through the center of the spinal canal and the visually determined center of the anterior part of the vertebral body. The mid-sagittal axis of the trunk was defined as the line between the center of the spinal canal and the center of the sternum at level T8. Rotation of the anterior part of the vertebra to the right side of the patient was defined as a positive angle, to the left as a negative angle (Fig. 3). Three observers (M.J., T.B. and R.W., see “Acknowledgments”) independently measured the vertebral rotation of T2–L5 in all positions using an in-house developed interactive computer program. Circles, which were adjustable in size, assisted the observers in determining the centers of the spinal canal and sternum (Fig. 3). The delineations performed by the observer and the positional information about the orientation and location of the scan planes in the laboratory (scanner) frame (stored in the header of the DICOM files that contained the images) were used in calculating the rotation angles.
Fig. 3.

Vertebral rotation angle in a transverse MRI scan at level T8
One of the three observers (R.W.) performed all measurements two times on separate sittings. The centers of the spinal canal and sternum at level T8 (which determine the position the mid-sagittal axis of the trunk) as well as the vertebral rotation angle were tested for inter-observer and intra-observer variability. Furthermore, the centers of the spinal canal were used to calculate spinal shape in the coronal and sagittal plane in all three positions.
Statistical analysis
The statistical analysis was performed using SPSS 12.0 for Windows (SPSS, Inc., Chicago, IL). Repeated measures analysis was performed to determine whether vertebral rotation between the positions was statistically significant and to analyze differences in the coronal and sagittal shape of the spine in the three different positions. Inter-observer and intra-observer variability were calculated using intra-class correlation coefficients (ICCs) [5]. A P value below 0.017 was considered to be statistically significant because a post hoc Bonferroni’s correction was applied.
Results
The vertebral rotation angles of the vertebrae in each position are shown in Table 1 and Fig. 4. In all three positions, the mid- and lower thoracic vertebrae (T6–T11) were predominantly rotated to the right. Maximum rotation in the upright position and supine position was at level T7 (2.7° and 2.8°, respectively). Maximum rotation in the quadrupedal-like position was at level T8 of 1.4°. Vertebrae in the quadrupedal-like position were found to be significantly less rotated as compared to the supine (P = 0.001) and standing upright positions (P < 0.001). Rotation patterns in the supine and standing upright position were not significantly different (P = 0.785). No significant differences were observed in vertebral rotation in all three positions between the sexes.
Table 1.
Mean vertebral rotation angle of level T2–L5 in all three positions
| Vertebra | Standing upright | Supine | Quadrupedal | |||
|---|---|---|---|---|---|---|
| N | Rotation angle (°) (mean ± SEM) | N | Rotation angle (°) (mean ± SEM) | N | Rotation angle (°) (mean ± SEM) | |
| T2 | 11 | 0.2 ± 0.8 | 7 | −0.2 ± 1.4 | 9 | −0.1 ± 0.8 |
| T3 | 23 | −0.3 ± 0.6 | 24 | −0.9 ± 0.6 | 28 | −1.2 ± 0.5 |
| T4 | 27 | −0.6 ± 0.6 | 27 | 0.0 ± 0.6 | 29 | −0.9 ± 0.6 |
| T5 | 30 | 1.1 ± 0.5 | 30 | 1.1 ± 0.5 | 30 | 0.4 ± 0.5 |
| T6 | 30 | 2.4 ± 0.6 | 30 | 2.6 ± 0.5 | 30 | 1.3 ± 0.5 |
| T7 | 30 | 2.7 ± 0.6 | 30 | 2.8 ± 0.5 | 30 | 1.3 ± 0.5 |
| T8 | 30 | 2.7 ± 0.6 | 30 | 2.8 ± 0.5 | 30 | 1.4 ± 0.6 |
| T9 | 30 | 2.5 ± 0.6 | 30 | 2.3 ± 0.5 | 30 | 0.7 ± 0.6 |
| T10 | 30 | 2.3 ± 0.5 | 30 | 2.2 ± 0.5 | 30 | 0.5 ± 0.5 |
| T11 | 30 | 2.1 ± 0.6 | 30 | 2.2 ± 0.6 | 30 | 0.6 ± 0.5 |
| T12 | 30 | 1.1 ± 0.7 | 30 | 1.2 ± 0.7 | 30 | 0.3 ± 0.5 |
| L1 | 30 | 1.2 ± 0.9 | 30 | 0.9 ± 0.7 | 30 | 0.1 ± 0.6 |
| L2 | 30 | 0.4 ± 0.8 | 30 | 0.4 ± 0.7 | 30 | −0.3 ± 0.6 |
| L3 | 30 | 0.5 ± 0.6 | 30 | 0.2 ± 0.7 | 30 | −0.7 ± 0.6 |
| L4 | 30 | −0.2 ± 0.6 | 30 | −0.0 ± 0.6 | 30 | −0.5 ± 0.5 |
| L5 | 30 | −0.4 ± 0.5 | 30 | −0.1 ± 0.6 | 30 | −0.5 ± 0.5 |
Fig. 4.

Mean vertebral rotation angles (in degrees, with SEM) in the transverse plane at level T2–L5 in 30 young adults in different positions
Analysis of the spinal shape in the coronal plane did not show any difference between the three positions (Fig. 5). In the sagittal plane, the spinal curvature is significantly different in the upright position when compared with the quadrupedal and supine position (both P < 0.01). In the upright position, the spine is more kyphotic in the upper thoracic region, equally curved in the mid-thoracic and thoracolumbar region and more lordotic in the lower lumbar region. However, the sagittal spinal profile was not different between the quadrupedal and supine position (P = 0.761), even though rotation was significantly different in these positions.
Fig. 5.

Mean sagittal and coronal shape of (the centers of) the spinal canal at level T3–L5 of 30 young adults in different positions (in mm with SEM). T9 is set as origin
ICCs for inter-observer and intra-observer variability of the vertebral rotation angles were 0.82 and 0.87, respectively. The ICCs for the inter-observer and intra-observer variability of the centers of the spinal canal and sternum are shown in Table 2.
Table 2.
ICCs of the inter-observer and intra-observer variability of the x and y coordinates of the centers of the spinal canal and sternum at level T8 (see Fig. 3 for the directions of the x and y axes within the image)
| Inter-observer variability | Intra-observer variability | |||
|---|---|---|---|---|
| x | y | x | y | |
| Center of spinal canal | 0.97 | 0.99 | 0.99 | 0.99 |
| Center of sternum | 0.92 | 0.99 | 0.91 | 0.99 |
Discussion
Although certain kinds of scoliotic-like curvatures have been described in a number of experimental animal models, human bipedalism appears to be a prerequisite for the development of IS [6, 7]. Despite the fact that two-legged locomotion is a common trait in nature, a unique characteristic of human bipedalism is the way we ambulate in a fully erect position with extended knees and hips. Thus, in man the center of mass of the upper body is constantly positioned straight above the pelvis, that in itself has also adjusted its shape to upright locomotion [8]. All other vertebrates, also the bipedal ones, ambulate with flexed knees and hips, thus putting the center of mass of the upper body consistently in front of the pelvis (Fig. 6) [9, 10].
Fig. 6.
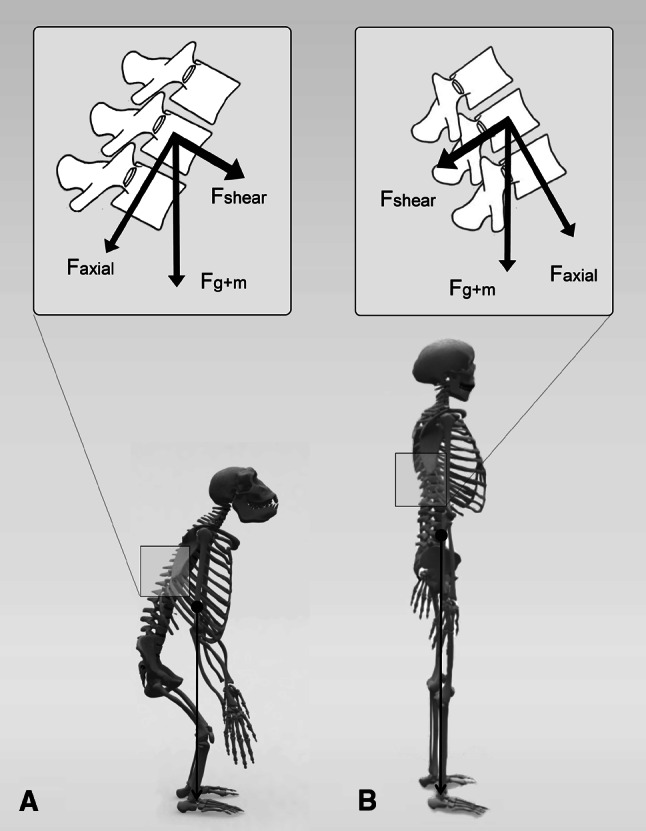
The sum of gravity and muscle force (F g+m) acting on a vertebra can be decomposed in an axial component (F axial) and an anterior-posterior shear component (F shear). Due to the horizontally orientated spine in quadrupeds, but also in occasional bipedal primates (a), this shear load will be ventrally directed. Only in the dorsally inclined thoracolumbar spinal segment of the human spine can shear loads be dorsally directed (b)
Forces acting on the human spine are obviously influenced in their magnitude and direction by this fully upright position [11]. In all vertebrates, the spine is predominantly loaded in an axial direction with superimposed ventrally directed shear loads (VDSL) due to gravity and muscle force. In the human spine, these VDSL are decreased due to a decrease in the anterior vector of gravity. Furthermore, dorsally directed shear loads (DDSL) are introduced to certain spinal areas (Fig. 6) [11]. In a previous biomechanical in vitro study, it was shown that these DDSL lead to a decrease in rotational stability of spinal segments [12].
The present in vivo study shows that body position, and thus the orientation of the spine in space, plays a role in the magnitude of vertebral rotation. Rotation was least in the so-called “quadruped-like” position, and increased in the human upright, but also in the supine position. Neither sagittal alignment, nor coronal alignment seems to be of influence for these findings. Both sagittal and coronal alignment of the spine were similar in the supine and quadrupedal position, although rotation differed significantly between these positions. The fully upright position showed a different sagittal profile, but rotation again was similar to the supine position.
We postulate that these findings can be explained by the fact that DDSL, that render the spine less stable in a rotatory sense, are actually non-existent in the quadrupedal position, when the spine and trunk are dependent in space. These DDSL increase, however, in both the upright and supine position. The finding that rotation increases in the supine position was initially surprising to us, but actually is very consistent with our other findings, and with the literature on infantile scoliosis. Obviously, in a supine position, the spine is very much subject to DDSL, making it less stable in rotation, as we showed in our in vitro experiment [12]. This finding is consistent with earlier literature showing that severe forms of early onset scoliosis occurred predominantly in infants that were put to sleep in a supine position. Infants that were put in their cots prone or in an alternate position did not develop this severe deformation [13]. Supposedly, being in a supine position for prolonged periods of time at a very young age with a more plastic skeleton can increase already present slight rotational patterns in the spine. This in turn could lead to morphologic changes in the developing spine under Hueter–Volkmann’s and Wolf’s laws, ultimately resulting in the vicious circle that ends in early onset scoliosis [14].
Similar to the pattern found in our previous CT study in the normal, non-scoliotic spine [2], the rotational pattern found in all three positions studied, corresponds to what is seen in the most prevalent types of IS, which show a primary thoracic curve rotated to the right with compensatory curves to the left above and below. An explanation for the predominance of right-rotating thoracic curves can possibly be found in this pre-existent rotation. We assume that once the spine starts to deteriorate into progressive deformity, it will most likely follow this built-in rotational pattern, rather than revert to the opposite direction.
It must be noted that in some tall subjects (mainly males) the field of view of the MRI scanner was not large enough to include the most cranial vertebrae. In addition, some upper thoracic vertebrae were excluded due to motion artefacts or a low signal-to-noise ratio. The present study underscores the importance of sagittal spinal alignment. The balance between the axially, ventrally and dorsally directed mechanical loads is influenced by differences in sagittal spinal alignment, and determines the rotational stability of the spine.
Which vertebrae are less stable in rotation depends on their orientation in space: the more backwardly inclined the spinal segment is, or the greater the number of superimposed vertebrae that are backwardly inclined, the more likely the segment will develop rotation (Fig. 6). Whether this rotation will progress may depend on several factors, one of which being the overall balance and magnitude of axial, ventral and dorsal forces that act on the spine during critical periods of rapid spinal growth, but also on the individual’s various compensating mechanisms, such as proprioceptive ability, ligament stiffness, trunk musculature and many others that have been suggested to play a role in the aetiopathogenesis of IS.
It is well known that sagittal spinal alignment changes with growth. Unfortunately, this is not well quantified at the individual vertebral level, neither for different periods of growth nor for differences between the sexes. We recently showed in young asymptomatic adults that the female spine is more backwardly inclined than the male spine [15]. If this is also the case during the period of peak adolescent growth velocity, it means that at this crucial moment the female spine may be less rotationally stable than the male spine. This might be one of the reasons why IS has a higher incidence in girls.
Conclusion
Pre-existent vertebral rotation in a supine, upright and quadrupedal position in the normal spine was analyzed in vivo in a human model. In all three positions, the mid- and lower thoracic vertebrae were predominantly rotated to the right. However, rotation was less in the quadrupedal position than in both the standing upright and supine positions. Neither coronal, nor sagittal alignment in the different positions appears to play a role in this difference in rotation. The erect as well as the supine spine show a higher tendency to rotate than the quadrupedal spine with its dependent thorax and abdomen. In depth analysis of the balance between the forces that act on the spine during critical periods of rapid growth, may shed more light on the still intangible aetiopathogenesis of IS.
Acknowledgments
This study was funded by Biomet and the Anna Foundation. The authors would like to thank radiology technologists S. Kleuskens, H. Klein Gebbink and A. van Aalten of the Sint Maartenskliniek for their invaluable help in the acquisition of the MRI scans and the independent observers R. de Wilde and T. van den Bosch (both medical students). In addition, the authors would like to thank all the volunteers who participated in the study. No benefits in any form have been or will be received from a commercial party related directly or indirectly to the subject of this manuscript.
Conflict of interest statement
This study was funded by Biomet and the Anna Foundation.
References
- 1.Xiong B, Sevastik J, Hedlund R, Sevastik B. Segmental vertebral rotation in early scoliosis. Eur Spine J. 1993;2:37–41. doi: 10.1007/BF00301053. [DOI] [PubMed] [Google Scholar]
- 2.Kouwenhoven JW, Vincken KL, Bartels LW, Castelein RM. Analysis of preexistent vertebral rotation in the normal spine. Spine. 2006;31:1467–1472. doi: 10.1097/01.brs.0000219938.14686.b3. [DOI] [PubMed] [Google Scholar]
- 3.Kouwenhoven JW, Vincken KL, Bartels LW, Meij BP, Oner FC, Castelein RM. Analysis of preexistent vertebral rotation in the normal quadruped spine. Spine. 2006;31:E754–E758. doi: 10.1097/01.brs.0000240209.85498.01. [DOI] [PubMed] [Google Scholar]
- 4.Kouwenhoven JW, Bartels LW, Vincken KL, Viergever MA, Verbout AJ, Delhaas T, Castelein RM. The relation between organ anatomy and pre-existent vertebral rotation in the normal spine: magnetic resonance imaging study in humans with situs inversus totalis. Spine. 2007;32:1123–1128. doi: 10.1097/01.brs.0000261563.75469.b0. [DOI] [PubMed] [Google Scholar]
- 5.Shrout PE, Fleiss JL. Intraclass correlations: uses in assessing rater reliability. Psychol Bull. 1979;86(2):420–428. doi: 10.1037/0033-2909.86.2.420. [DOI] [PubMed] [Google Scholar]
- 6.Naique SB, Porter R, Cunningham AA, Hughes SP, Sanghera B, Amis AA. Scoliosis in an Orangutan. Spine. 2003;28:E143–E145. doi: 10.1097/00007632-200304010-00026. [DOI] [PubMed] [Google Scholar]
- 7.Arkin AM. The mechanism of the structural changes in scoliosis. J Bone Joint Surg. 1949;31A(3):519–528. [PubMed] [Google Scholar]
- 8.Hogervorst T, Bouma HW, De Vos J. Evolution of the hip and pelvis. Acta Orthop. 2009;(Suppl 336)80:1–39. doi: 10.1080/17453690610046620. [DOI] [PubMed] [Google Scholar]
- 9.Alexander RM. Bipedal animals, and their differences from humans. J Anat. 2004;204:321–330. doi: 10.1111/j.0021-8782.2004.00289.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.D’Août K, Aerts P, De Clercq D, De Meester K, Van Elsacker L. Segment and joint angles of hind limb during bipedal and quadrupedal walking of the bonobo (Pan paniscus) Am J Phys Anthropol. 2002;119:37–51. doi: 10.1002/ajpa.10112. [DOI] [PubMed] [Google Scholar]
- 11.Castelein RM, van Dieen JH, Smit TH. The role of dorsal shear forces in the pathogenesis of adolescent idiopathic scoliosis—a hypothesis. Med Hypotheses. 2005;65:501–508. doi: 10.1016/j.mehy.2005.03.025. [DOI] [PubMed] [Google Scholar]
- 12.Kouwenhoven JW, Smit TH, van der Veen AJ, Kingma I, van Dieën JH, Castelein RM. Effects of dorsal versus ventral shear loads on the rotational stability of the thoracic spine: a biomechanical porcine and human cadaveric study. Spine. 2007;32:2545–2550. doi: 10.1097/BRS.0b013e318158cd86. [DOI] [PubMed] [Google Scholar]
- 13.Mau H. Etiology of idiopathic infantile scoliosis. Reconstr Surg Traumatol. 1972;13:184–190. [PubMed] [Google Scholar]
- 14.Stokes IA. Analysis and simulation of progressive adolescent scoliosis by biomechanical growth modulation. Eur Spine J. 2007;16(10):1621–1628. doi: 10.1007/s00586-007-0442-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Janssen MM, Drevelle X, Humbert L, Skalli W, Castelein RM. Differences in male and female spino-pelvic alignment in asymptomatic young adults: a three-dimensional analysis using upright low-dose digital biplanar X-rays. Spine. 2009;34(23):E826–E832. doi: 10.1097/BRS.0b013e3181a9fd85. [DOI] [PubMed] [Google Scholar]