

Abstract
BACKGROUND AND PURPOSE
Cannabinoid CB1 receptor antagonists/inverse agonists, potentiate toxin-induced nausea and vomiting in animal models. Here, we sought to determine if this potentiated nausea was mediated by inverse agonism or neutral antagonism of the CB1 receptor, and if the potentiated nausea would be produced by intracerebroventricular (icv) administration of an inverse agonist.
EXPERIMENTAL APPROACH
The conditioned gaping model of nausea in rats was used to compare the CB1 receptor antagonist/inverse agonist, AM251, and the CB1 receptor neutral antagonists, AM6527 (centrally and peripherally active) and AM6545 (peripherally active), in potentiating conditioned gaping produced by lithium chloride (LiCl) solution. The effect of icv (lateral ventricle and 4th ventricle) administration of AM251 on LiCl-induced gaping in this model was also evaluated.
KEY RESULTS
At a dose that did not produce conditioned gaping on its own, systemically administered AM251 (1.25 mg·kg−1) potentiated LiCl-induced conditioned gaping and reduced sucrose palatability; however, even doses as high as 8 mg·kg−1 of AM6545 and AM6527 neither potentiated LiCl-induced conditioned gaping nor reduced sucrose palatability. Infusions of AM251 into the lateral ventricles (1.25, 12.5 and 125 µg) or the 4th ventricle (2.5, 12.5 and 125 µg) did not potentiate LiCl-induced conditioned gaping reactions, but all doses attenuated saccharin palatability during the subsequent test.
CONCLUSIONS AND IMPLICATIONS
Inverse agonism, but not neutral antagonism, of CB1 receptors potentiated toxin-induced nausea. This effect may be peripherally mediated or may be mediated centrally by action on CB1 receptors, located distal to the cerebral ventricles.
Keywords: CB1 receptor, inverse agonist, neutral antagonist, conditioned gaping, nausea, AM251, AM6545, AM6527, intracerebroventricular, palatability, rat
Introduction
The marijuana plant has been used for several centuries for control of nausea and vomiting (Iversen, 2000). The anti-emetic effects of cannabinoids appear to be mediated by actions at the cannabinoid CB1 receptor (Darmani, 2001; Van Sickle et al., 2001; receptor nomenclature follows Alexander et al., 2009). CB1 receptors are found in the gastrointestinal tract and its enteric nervous system (Pertwee, 2001) as well as within the emetic system of the brain (Van Sickle et al., 2001; Van Sickle et al., 2003) in the dorsal vagal complex, consisting of the area postrema (AP), nucleus of the solitary tract (NTS) and the dorsal motor nucleus of the vagus (DMNX) in the brainstem of rats, ferrets and the least shrew (Simoneau et al., 2001; Van Sickle et al., 2001; Darmani and Johnson, 2004). CB1 receptors in the NTS are activated by centrally administered Δ9-THC and this activation is blocked by the selective CB1 receptor antagonists/inverse agonists, SR-141716 (Darmani and Johnson, 2004) and AM251 (Van Sickle et al., 2003). Indeed, c-Fos expression induced by cisplatin in the DMNX, NTS and AP is significantly reduced by Δ9-THC (Van Sickle et al., 2001; 2003). However, recently it has been also been reported that activation of peripheral CB1 receptors by Δ9-THC may also play a role in the suppression of vomiting (Darmani and Johnson, 2004) and in the regulation of feeding (Gomez et al., 2002; Sink et al., 2009b).
Activation of CB1 receptors clearly attenuates vomiting produced by toxins; however, the effect of activation of these receptors on nausea is less well understood. Although vomiting is well controlled by current anti-emetic therapies, the control of nausea in chemotherapy patients continues to be problematical (Andrews and Horn, 2006). Understanding the neuropharmacology of nausea so that it can be treated as effectively as vomiting would improve clinical treatment. However, the subjective nature of nausea has limited its investigation to human self-report measures. Traditionally, the potential of drugs to produce nausea has been assessed by the conditioned taste avoidance measure (see Garcia et al., 1974). However, this measure has been shown to be non-selective to the nauseating effects of drugs, because drugs that do not produce nausea (and in fact even those shown to be rewarding) produce taste avoidance, and anti-emetic drugs generally do not interfere with conditioned taste avoidance (Grant, 1987; see Parker et al., 2008).
Considerable recent evidence, however, suggests that conditioned gaping reactions (rapid, large amplitude opening of the mandible with retraction of the corners of the mouth) in rats represents a preclinical test for the evaluation of nausea-inducing drug effects (Parker et al., 2008; Rock et al., 2009; Tuerke et al., 2009). Although rats are not capable of vomiting, they display characteristic conditioned gaping reactions when exposed to a flavor previously paired with a nausea-inducing treatment, such as lithium chloride (LiCl) solution (see Parker et al., 2008). Unlike conditioned taste avoidance, only emetic treatments produce conditioned gaping reactions and anti-emetic drugs attenuate conditioned gaping reactions (Parker, 2003).
Cannabinoid compounds modify conditioned gaping in rats in a manner that suggests they reduce nausea as well as vomiting. Cannabinoid agonists, including Δ9-tetrahydrocannabinol (Δ9-THC), HU210 and URB597, interfere with the establishment of LiCl-induced conditioned gaping reactions in rats, presumably by interfering with nausea produced by LiCl (Parker and Mechoulam, 2003; Parker et al., 2003; Cross-Mellor et al., 2007). The interference with nausea by cannabinoid agonists is reversed by pretreatment with SR141716 (rimonabant; Parker et al., 2003; Cross-Mellor et al., 2007), suggesting a mechanism of action, mediated via CB1 receptors.
Rimonabant and AM251 are not receptor neutral antagonists, instead they act as CB1 receptor inverse agonists/antagonists (Pertwee, 2005). When administered on their own for inhibition of food intake and body weight gain, these agents produce nausea as a side effect in humans (Janero and Makriyannis, 2009). As well, they produce nausea in rats (McLaughlin et al., 2005) and vomiting in the least shrew (Darmani, 2001). More recently CB1 receptor neutral antagonists, such as AM4113, have been developed which bind to the receptor without eliciting changes in constitutive or intrinsic cellular endocannbinoid signalling. CB1 receptor neutral antagonists (AM4113, AM6545 and AM6527) have also been shown to reduce feeding and weight gain in rats at doses comparable with Rimonabant and AM251, but without the side effect of nausea in rats or vomiting in ferrets (Chambers et al., 2007; Salamone et al., 2007; Sink et al., 2008; Cluny et al., 2010; Tam et al., 2010). AM6545 also completely reversed the effects of WIN55, 212-2 in an assay of colonic motility, but had no effect on its own (Cluny et al., 2010). These effects suggest that the nausea-inducing effects of compounds acting like rimonabant are mediated by their inverse agonist effects at the CB1 receptor.
When administered at a dose that does not produce nausea or vomiting on its own, AM251 also potentiated LiCl-induced conditioned gaping reactions in rats (Parker et al., 2003) and morphine 6-glucuronide (M6G) induced vomiting in ferrets (Van Sickle et al., 2001; Chambers et al., 2007). That is, the emetic effects of known emetic agents are potentiated in rats and ferrets pretreated with a subthreshold dose of a CB1 inverse agonist/antagonist. Interestingly, Chambers et al. (2007) recently reported that, unlike AM251, the CB1 receptor neutral antagonist, AM4113, did not potentiate vomiting produced by M6G in the ferret, suggesting that the potentiation of toxin-induced vomiting was also mediated by inverse agonism of the CB1 receptor. Here we present evidence that inverse agonism, but not neutral antagonism, of the CB1 receptor also potentiated LiCl-induced nausea (at non-nauseating doses) in the conditioned gaping model in rats. The following experiments evaluated the potential of a low dose of AM251 (1.25 mg·kg−1) and a range of doses of the CB1 receptor neutral antagonists, AM6545 [with limited penetration across the blood–brain barrier (BBB); see Makriyannis et al., 2009; Cluny et al., 2010; Tam et al., 2010] and AM6527 (which crosses the BBB; see Sink et al., 2009a), to potentiate LiCl-induced nausea in rats.
As there is evidence that both central and peripheral CB1 receptors may be involved in the control of vomiting responses in emetic species (van Sickle et al., 2001; Darmani and Johnson, 2004), it is unclear whether the potentiation of LiCl-induced nausea by systemically administered CB1 receptor inverse agonists/antagonists in rats is centrally or peripherally mediated. Therefore, additional experiments evaluated the effect of intracerebroventricular (icv) injections of AM251 on LiCl-induced nausea in rats. In separate experiments, AM251 was infused into the lateral ventricles (primarily targeting forebrain structures) and to the 4th ventricles (primarily targeting brainstem structures) and the effect on LiCl-induced conditioned gaping was measured.
Methods
Animals
All animal care and experimental procedures were approved by the Animal Care Committees of Wilfrid Laurier University and the University of Guelph and adhered to the guidelines of the Canadian Council of Animal Care. Animals were maintained on ad libitum food (Highland Rat Chow [8640]) and water throughout the experiments. We used male Sprague Dawley rats (Charles River Lab, St Constant, Quebec) weighing from 300 to 380 g on the day of conditioning. The rats were individually housed in Plexiglas cages (48 × 26 × 20 cm) in the colony room at an ambient temperature of 21°C with a 12/12 reverse light/dark schedule (lights off at 7 am). All experimental manipulations were conducted during the dark phase of the cycle. Rats were provided with two clean paper towels (replenished during weekly cage changes) and a soft plastic container that was 14 cm long and 12 cm in diameter that remained in the home cage.
Intraoral cannulation surgery
All rats were implanted with intraoral cannula for delivery of the 0.1% saccharin solution directly to the oral cavity. On the day of surgery, the rats were injected with an antibiotic (Derapin: 100 mg·kg−1, sc; Ayerst) 30 min prior to being anaesthetized with isoflurane (4–5% induction, 1.5% maintenance in O2). Once anaesthetized a 2 cm2 section of skin was shaved at the back of the neck at the level of the scapula. The skin was prepared by cleaning with soap (Bactistat; Ecolab, St. Paul, MN, USA) and wiping with 70% isopropyl alcohol followed by 7% Betadine solution (Purdue Products L.P., Stamford, CT, USA). The rat was then administered a 5 mg·kg−1 injection (ip) of the anti-inflammatory/analgesic drug carprofen (Rimadyl; Pfizer Canada Inc., Kirkland, QC, Canada). A thin-walled 15 G stainless steel needle was inserted at the shaved area on the neck, directed subcutaneously around the ear and brought out behind the first molar inside the mouth. A 10 cm length of Intra Medic PE90 tubing (Clay Adams Brand; Becton Dickinson and Co., Sparks, MD, USA) with an inner diameter (I.D.) of 0.86 mm and an outer diameter (O.D.) of 1.27 mm was then inserted through the needle after which the needle was removed. Betadine (10%) was applied to the puncture site and two elastic discs (2 cm2) were placed over the exposed end of the tubing and drawn to the skin at the back of the neck for the purpose of stabilizing the cannula. The cannula was held secure in the oral cavity by a 6 mm disc of polypropylene mesh (297 micron; Small Parts Inc., Miramar, FL, USA) secured behind the heat flanged intraoral opening. Each cannula was constructed prior to surgery and kept in a cold sterilent solution (Germiphene Corporation, Brantford, ON, Canada). Rats were returned to their home cage immediately following surgery and food pellets were loosely provided in the cage. For 3 days following surgery indices of recovery were assessed (body weight, facial swelling, activity, etc.) and the cannula was flushed daily with an oral cleansing solution (Nolvadent; Ayerst, Fort Dodge, Iowa, USA).
Stereotaxic guide cannula surgery
For the studies of central effects of AM251, rats had a unilateral indwelling guide cannula surgically implanted in the lateral ventricle (Experiment 3a) or the 4th ventricle (Experiment 3b). They were anaesthetized with isoflurane gas and prepared for surgery by shaving a strip of skin between the ears (2.5–3 cm long), cleaning the skin and injecting carprofen (5 mg·kg−1; ip) and a local anaesthetic (0.1 mL; sc; Marcaine; Hospira, Montreal, QC, Canada) on either side of the skull. They were then stabilized in the flat skull position (according to Paxinos and Watson, 1998) in the stereotaxic frame and the skull was exposed. In Experiment 3a, the stainless steel guide cannula (22G, 8 mm below pedestal) was lowered into either the left or right lateral ventricle (counterbalanced among groups) using the following coordinates from Bregma: −1.0 mm AP, ±1.4 mm LM, –3.6 mm DV (Erb et al., 2003). In Experiment 3b, the guide cannula (22 G, 11 mm below pedestal) was lowered into the 4th ventricle in the right or left hemisphere (counterbalanced among groups) using the following coordinates from interaural zero: AP −3.5; LM ± 1.4; DV −6.2 (Blevins et al., 2004). The guide cannula was stabilized by four screws secured in the skull and dental cement. Once the dental cement had hardened, a stainless steel obdurator was inserted in the guide cannula to maintain patency. Immediately following removal from the stereotaxic frame, the rats were surgically implanted with an intraoral cannula then placed in a heated recovery area and monitored until they were ambulatory, at which time they were returned to the colony room. The rats were allowed to recover for 5 days following surgery and were monitored daily as previously described.
Histology
Verification of lateral ventricle cannula placement was conducted 3 days following surgery by injecting 2 µL angiotensin II (50 ng/2 µL) at a rate of 2 µL·min−1. If the rat drank from a water bottle within 1 min of the infusion and sustained drinking for 2–3 min the cannula was considered to be accurately placed (Erb et al., 2003), otherwise it was removed from further testing.
Verification of cannula placement into the 4th ventricle was evaluated by histological evaluation of tissue. Rats were deeply anaesthetized using an 85 mg·kg−1 injection of Euthansol (Intervet Canada Corp., Kirkland, QC, Canada) followed by transcardial perfusion with phosphate buffered saline (0.1 M) and 4% formalin. The brains were removed and stored at 4°C in 4% formalin solution for 24–48 h after which they were placed in a 20% sucrose solution overnight at room temperature. The brains were then sliced in 60 µm sections using a CM1850 Leica cryostat and relevant sections were mounted on glass microscope slides. The tissue was stained with cresyl violet 24 h later and examined for accurate cannula placement using a Leica MZ6 Stereomicroscope with a Leica DFC420 Digital Camera and Leica Application Suite software. Data from any rat with improper cannula placement were excluded from the behavioural analyses.
Apparatus
Conditioning and testing was conducted in a square Plexiglas ‘Taste Reactivity’ (TR) chamber (26.5 × 26.5 × 12 cm) placed on a glass-topped table with a mirror set at 45° below the glass top to facilitate videotaping the ventral surface of the rat while in the chamber. The rat's cannula was attached to the infusion pump (KDS100; KD Scientific Inc, Holliston, MA, USA) using a length of PE160 tubing fitted over the intraoral cannula that ran through a hole in the top of the chamber and connected to the pump. Orofacial responses were captured directly to the computer using the Roxio Videowave Premiere Suite 8 video capture program and a Sony DCR-HC28 Handycam camera.
For central administration of solutions, microinfusions were conducted using a 28 gauge injector that extended 1 mm beyond the guide cannula tip and was attached to a microinfusion pump (KDS101; KD Scientific Inc, Holliston, MA, USA) using Tygon tubing with a 0.1905 mm ID (Cole Parmer, Vernon Hills, IL, USA).
Procedures
Experiment 1: Potential of systemic AM251 to potentiate LiCl-induced conditioned gaping
The TR procedure was similar in all experiments except for the drugs delivered. Three days after recovering from the intraoral cannulation surgery, the rats received an adaptation trial; they were individually placed in the TR chamber with their cannula attached to an infusion pump for fluid delivery. Water was infused into their intraoral cannula over a period of 2 min at the rate of 1 mL·min−1 after which they were returned to their home cage. The rats received a conditioning trial 24 h after the adaptation trial. Prior to the conditioning trial, the rats were randomly assigned to groups on the basis of treatment drug (AM251/vehicle) and conditioning drug (LiCl/saline) as follows: AM251/LiCl (n = 7), AM251/Saline (n = 7), Vehicle/LiCl (n = 8) and Vehicle/Saline (n = 8). Thirty min prior to the conditioning trial, the rats were injected ip with either 1.25 mg·kg−1 AM251 or vehicle according to group assignment. They were then placed in the TR chamber and were intraorally infused with 0.1% saccharin for 2 min at a rate of 1 mL·min−1. Immediately following the saccharin infusion, the rats were injected with either 20 mL·kg−1 of LiCl or saline according to group assignment and returned to their home cage. The animals were given a second adaptation trial with a 2 min intraoral infusion of water, 48 h following the conditioning trial and 24 h prior to a drug-free TR test trial. The rats were placed in the TR chamber and infused with 0.1% saccharin solution over a period of 2 min (1 mL·min−1) while their orofacial reactions were recorded from the mirror set at a 45° angle beneath the glass floor of the chamber.
Experiment 2: Potential of CB1 receptor neutral antagonists to potentiate LiCl-induced conditioned gaping
The rats were treated as in Experiment 1 except as indicated. In Experiment 2a, the rats were assigned to groups on the basis of treatment drug (AM6545/Vehicle) and conditioning drug (LiCl/Saline) as such: Vehicle/LiCl (n = 8), Vehicle/Saline (n = 8), 1.25 mg·kg−1 AM6545/LiCl (n = 8), 1.25 mg·kg−1 AM6545/Saline (n = 8), 2.5 mg·kg−1 AM6545/LiCl (n = 8), 2.5 mg·kg−1 AM6545/Saline (n = 9), 8 mg·kg−1 AM6545/LiCl (n = 8), 8 mg·kg−1 AM6545/Saline (n = 8). In Experiment 2b, the rats were assigned to groups on the basis of treatment drug (AM6527/Vehicle) and conditioning drug (LiCl/Saline) as such: Vehicle/LiCl (n = 8), Vehicle/Saline (n = 6), 8 mg·kg−1 AM6527/LiCl (n = 8), 8 mg·kg−1 AM6527/Saline (n = 8). Twenty-four hours after the adaptation trial, on the conditioning trial, the rats were injected with the treatment drug 30 min prior to receiving a 2 min intraoral infusion of 0.1% saccharin solution (1 mL·min−1) and were immediately injected with conditioning drug. Seventy-two hours later (24 h after the second adaptation trial), the rats received a drug-free 2 min TR test with 0.1% saccharin solution.
Experiment 3: Potential of icv AM251 given into the lateral ventricle or the 4th ventricle to potentiate LiCl-induced conditioned gaping
Experiment 3 was conducted as Experiment 1, except AM251 or vehicle was delivered icv to either the lateral or the 4th ventricle. All rats had a unilateral indwelling cannula surgically implanted in the lateral ventricle (Experiment 3a) or the 4th ventricle (Experiment 3b). The TR adaptation trial occurred on the sixth day following surgery and was conducted as in Experiment 1. The conditioning trial occurred 24 h after the adaptation trial. In Experiment 3a (lateral ventricle placement), the rats were randomly assigned to groups on the basis of treatment drug (Vehicle, 1.25 µg AM251, 12.5 µg AM251 or 125 µg AM251) and conditioning drug (LiCl or saline) as follows: Vehicle/LiCl (n = 7), Vehicle/Saline (n = 7), 1.25 µg AM251/LiCl (n = 7), 1.25 µg AM251/Saline (n = 7), 12.5 µg AM251/LiCl (n = 7), 12.5 µg AM251/Saline (n = 8), 125 µg AM251/LiCl (n = 6), 125 µg AM251/Saline (n = 6). In Experiment 3b (4th ventricle placement), the rats were assigned to the following groups: Vehicle/LiCl (n = 8), Vehicle Saline (n = 6), 2.5 µg AM251/LiCl (n = 8), 2.5 µg AM251/Saline (n = 7), 12.5 µg AM251/LiCl (n = 6) and 12.5 µg AM251/Saline (n = 5). In Experiment 3b, an additional group was also administered 125 µg AM251/LiCl (n = 4) and was compared with group Vehicle/LiCl. All n-values refer to rats with correct cannula placements.
On the conditioning trial of both experiments, the obdurator was removed from the guide cannula and while the rat was restrained (wrapped in a towel) a 28 G injector was inserted and 2 uL of the treatment drug was microinfused at a rate of 2 µL·min−1. Following the infusion, the injector was left in place for 1 min before removal and replacement of the obdurator. Thirty min later, the rats were placed in the TR chamber and intraorally infused with 2 mL of 0.1% saccharin solution at the rate of 1 mL·min−1 and their orofacial reactions were videotaped. Forty-eight hours later, the rats received an adaptation trial followed 24 h later by the drug-free test trial during which they were intraorally infused with 0.1% saccharin solution and their orofacial reactions videorecorded.
Behavioural measures
The video recordings were scored using the Observer (Noldus Information Technology, Sterling, VA) event recording program for the following behaviours: The number of gaping reactions (rapid, large amplitude opening of the mandible with retraction of the corners of the mouth) was counted during the 2 min sessions. The number of seconds that the rats displayed hedonic reactions of mouth movements (movement of the lower mandible without opening the mouth) and tongue protrusions (extensions of the tongue out of the mouth) was also measured and the total of the two measures was used as the hedonic score.
Data analysis
In each experiment, the rats did not display gaping reactions during the conditioning trial. Therefore, the number of gaping reactions during the test trials only were entered into a between factors analysis of variance (anova), with the factors of treatment drug and conditioning drug. Subsequent Bonferroni post hoc comparison or planned comparison tests were used as appropriate. The hedonic reaction scores were entered into mixed factors anovas, with treatment drug, conditioning drug and trial (conditioning trial/ test trial) as factors. When appropriate, separate, two factor, between groups, anovas were then conducted for the conditioning trial and the test trial.
Materials
Lithium chloride and angiotensin II were obtained from Sigma-Aldrich Canada, Toronto, ON, Canada; Isoflurane from Benson Medical Industries Inc. Toronto, ON Canada and 2-hydroxypropyl-β-cyclodextrin (2-HPBCD) from ONBIO Inc, Richmond Hill, ON, Canada. The AM compounds (AM251, AM6545 and AM6527) were synthesized at the Center for Drug Discovery, Northwestern University. The stainless steel guide cannula was from Plastics One Inc., Roanoke, VA, United States.
Lithium chloride was prepared in a 0.15 M solution with sterile water and was administered i.p. in a volume of 20 mL·kg−1 (127.2 mg·kg−1). AM251, AM6545 and AM6527 were prepared (1 mg·mL−1) in a vehicle of 45% 2-HPBCD in sterile water.
Results
Experiment 1: Effects of systemic AM251 on LiCl-induced conditioned gaping
When administered ip, a low dose of AM251 potentiated LiCl-induced conditioned gaping, as has been reported with rimonbant (Parker et al., 2003). The upper section in Figure 1 presents the mean number of gaping reactions displayed by the various groups in Experiment 1 during the conditioning trial and during the test trial. As is apparent, the rats did not display gaping reactions during the conditioning trial. The mean number of gaping reactions during the test trial was entered into a 2 × 2 anova which revealed significant effects of treatment drug, F (1, 26) = 7.8, P < 0.01, conditioning drug, F (1, 26) = 56.7; P < 0.001 and a significant (treatment drug × conditioning drug) interaction, F (1, 26) = 5.0; P < 0.05. Subsequent planned comparison tests revealed that the groups Veh-LiCl and AM251-LiCl displayed more gaping reactions than the groups Veh-Saline or AM251-Saline (P < 0.01); however, the group AM251-LiCl also displayed significantly more gaping reactions than the group Veh-LiCl (P < 0.01).
Figure 1.
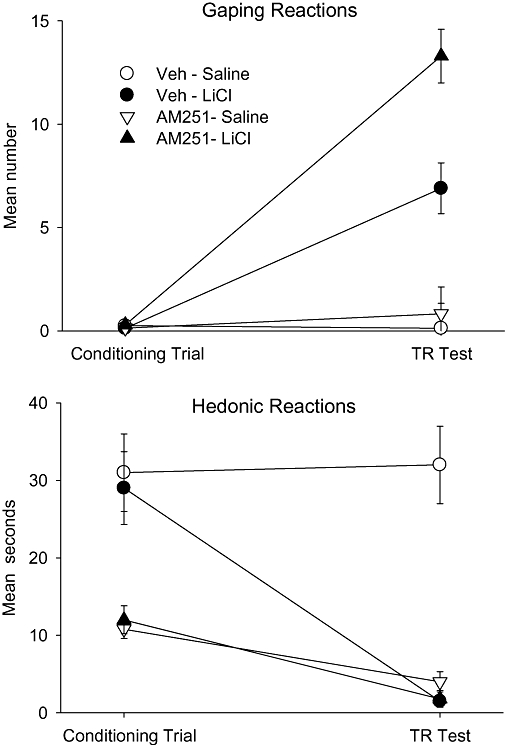
Mean (±SEM) number of gaping reactions (upper section) and mean (±SEM) seconds spent displaying hedonic reactions (tongue protrusions + mouth movements) during the conditioning trial and the drug-free TR test trial by rats injected (ip) with vehicle or 1.25 mg·kg−1 AM251 30 min prior to a saccharin–LiCl or saccharin–saline pairing. LiCl, lithium chloride; TR, Taste Reactivity.
Systemic administration of AM251 also modified hedonic reactions elicited by saccharin solution during both conditioning and testing. The bottom section of Figure 1 presents the mean number of seconds that the rats displayed hedonic reactions during the conditioning trial and the test trial of Experiment 1. The mixed factors anova revealed significant effects of treatment drug, F (1, 26) = 16.5; P < 0.001, conditioning drug, F (1, 26) = 4.5; P < 0.05, and a (treatment drug × conditioning drug) interaction, F (1, 26) = 5.3; P < 0.05, as well as trial, F (1, 26) = 24.5; P < 0.001, (conditioning drug × trial), F (1, 26) = 13.8; P < 0.001 and a (treatment drug × conditioning drug × trial) interaction, F (1, 26) = 7.0; P < 0.025. On the conditioning trial, a 2 × 2 anova revealed only a significant effect of treatment drug, F (1, 26) = 19.8; P < 0.001, with rats treated with AM251 displaying fewer hedonic reactions than rats treated with vehicle (replicating Jarrett et al., 2007). On the test trial, a 2 × 2 anova revealed a significant effect of treatment drug, F (1, 26) = 7.5; P < 0.01, conditioning drug, F (1, 26) = 10.6; P < 0.01 and an (treatment drug × conditioning drug) interaction, F (1, 26) = 9.2; P < 0.01. Planned comparison tests of the data for the test trial revealed that the group Veh-Saline displayed more hedonic reactions than any other group (Ps < 0.01) and the remaining groups did not differ from one another.
Experiment 2: Effects of CB1 receptor neutral antagonists on LiCl-induced conditioned gaping
Systemic administration of neither the CB1 receptor neutral antagonist, AM6545 (peripherally restricted) nor AM6527 (crosses BBB), potentiated LiCl-induced conditioned gaping. These two compounds did not suppress hedonic reactions – during conditioning or during the subsequent drug-free test.
Experiment 2a
Even at a dose as high as 8 mg·kg−1, which suppresses feeding (Cluny et al., 2010; Tam et al., 2010), the peripherally restricted CB1 neutral antagonist, AM6545, did not potentiate LiCl-induced conditioned gaping in rats. The upper section of Figure 2 presents the mean number of gaping reactions displayed by the rats in the various groups on the conditioning trial and on the test trial. On the test trial, the 4 × 2 anova revealed only a significant effect of conditioning drug, F (1, 57) = 23.6; P < 0.001; LiCl produced conditioned gaping, but the strength of the response did not significantly differ among the treatment drug groups.
Figure 2.
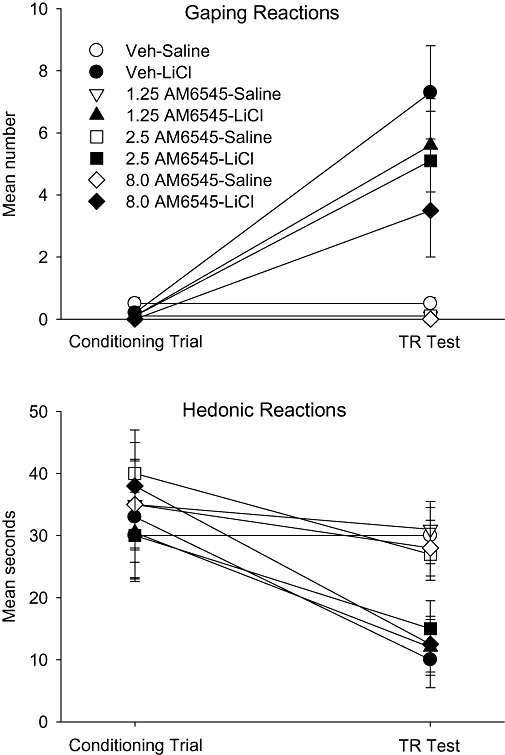
Mean (±SEM) number of gaping reactions (upper section) and mean (±SEM) seconds spent displaying hedonic reactions (lower section) during the conditioning trial and the drug-free TR test trial by rats injected (ip) with vehicle, 1.25, 2.5 or 8 mg·kg−1 AM6545 30 min prior to a saccharin–LiCl or saccharin–saline pairing. LiCl, lithium chloride; TR, Taste Reactivity.
Also, unlike AM251, AM6545 neither unconditionally nor conditionally suppressed hedonic reactions elicited by saccharin solution. The lower section of Figure 2 presents the mean seconds of hedonic reactions displayed by the various groups on the conditioning trial and on the TR test trial. The mixed factors anova revealed only a significant effect of trial, F (1, 57) = 22.5; P < 0.001 and an (conditioning drug × trial) interaction, F (1, 57) = 7.2; P = 0.01. Separate 4 × 2 between groups anovas for each trial revealed only a significant effect of conditioning drug on the test trial, F (1, 57) = 28.6; P < 0.001. Rats showed suppressed hedonic reactions to LiCl-paired saccharin on the test trial, but the strength of the suppression was not affected by treatment with AM6545.
Experiment 2b
The centrally active CB1 receptor neutral antagonist, AM6527 (8 mg·kg−1), neither potentiated LiCl-induced conditioned gaping nor suppressed hedonic reactions elicited by saccharin solution during the conditioning or test trials. The upper section of Figure 3 presents the mean number of gaping reactions displayed by the rats in the various groups on the conditioning trial and on the TR test trial. A 2 × 2 between groups anova for the test trial revealed only a significant effect of conditioning drug, F (1, 26) = 14.5; P < 0.001; that is, LiCl produced conditioned gaping that did not significantly differ among treatment groups. The lower section of Figure 3 presents the mean seconds of hedonic reactions displayed by the various groups on the conditioning trial and TR test trial. The mixed factors anova revealed only a significant (conditioning drug × trial) interaction, F (1, 26) = 6.6; P = 0.016. Planned comparison tests revealed that regardless of treatment condition, the LiCl-conditioned groups displayed fewer hedonic reactions on the test trial than on the conditioning trial (P < 0.01), while the saline-conditioned groups did not differ in hedonic reactions displayed during conditioning or testing.
Figure 3.
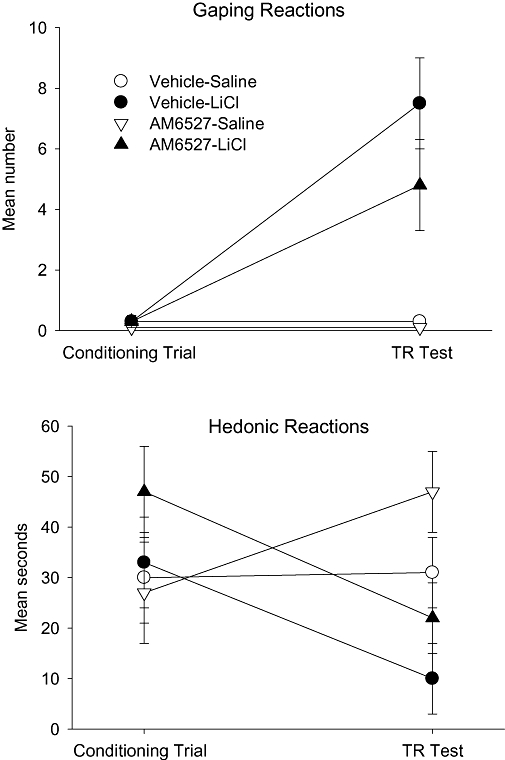
Mean (±SEM) number of gaping reactions (upper section) and mean (±SEM) seconds spent displaying hedonic reactions (lower section) during the conditioning trial and the drug-free TR test trial by rats injected (ip) with vehicle or 8 mg·kg−1 AM6527 30 min prior to a saccharin–LiCl or saccharin–saline pairing. LiCl, lithium chloride; TR, Taste Reactivity.
Experiment 3: Effects of AM251, given icv, to the lateral or the 4th ventricle on LiCl-induced conditioned gaping
Whether administered to the lateral ventricle or to the 4th ventricle, AM251 did not potentiate LiCl-induced conditioned gaping, even though it did modify saccharin palatability. The upper sections of Figures 4 and 5 present the mean number of gaping reactions displayed by the various groups on the conditioning trial and the test trial of Experiments 3a and 3b when AM251 or vehicle was delivered to the lateral ventricle or the 4th ventricle, respectively, prior to conditioning. In Experiment 3a (lateral ventricle) on the test trial, the 4 × 2 between groups anova of gaping reactions revealed only a significant conditioning drug effect, F (1, 47) = 25.6; P < 0.001. Likewise, the 3 × 2 between groups anova of gaping reactions on the test trial in Experiment 3b (4th ventricle) revealed a significant effect of conditioning drug, F (1, 34) = 27.7; P < 0.001. Finally in Experiment 3b, the mean number of gaping reactions displayed by the additional four rats given 125 µg AM251/LiCl (10.2 ± 4.3) did not significantly differ from the group Vehicle/LiCl (12.8 ± 4.0). In both Experiments 3a and 3b, LiCl did induce conditioned gaping in rats, but the treatment drug did not modify the strength of the gaping reactions.
Figure 4.
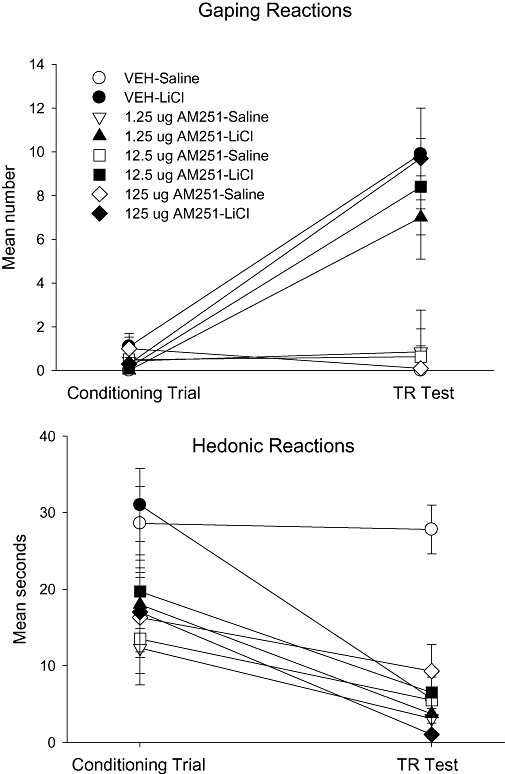
Mean (±SEM) number of gaping reactions (upper section) and mean (±SEM) seconds spent displaying hedonic reactions (lower section) during the conditioning trial and the drug-free TR test trial by rats injected (icv) with vehicle, 1.25, 12.5 or 125 µg AM251 into the lateral ventricles 30 min prior to a saccharin–LiCl or saccharin–saline pairing. LiCl, lithium chloride; TR, Taste Reactivity.
Figure 5.
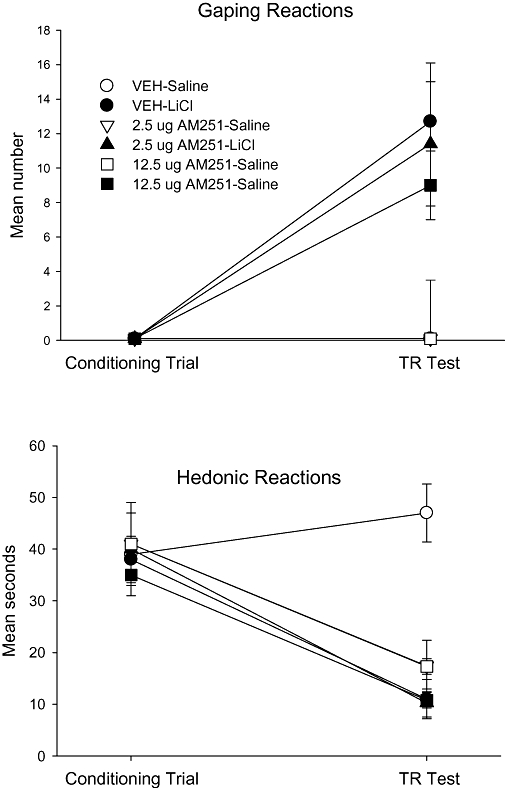
Mean (±SEM) number of gaping reactions (upper section) and mean (±SEM) seconds spent displaying hedonic reactions (lower section) during the conditioning trial and the drug-free TR test trial by rats injected (icv) with vehicle, 2.5 or 12.5 µg AM251 into the fourth ventricle 30 min prior to a saccharin–LiCl or saccharin–saline pairing. LiCl, lithium chloride; TR, Taste Reactivity.
Central administration of AM251 to either the lateral or 4th ventricle did, however, conditionally attenuate the palatability of saccharin reflected in reduced hedonic reactions during the test trial, as can be seen in the lower sections of Figures 4 and 5 respectively. Furthermore, when delivered to the lateral ventricle, but not to the 4th ventricle, AM251 also suppressed hedonic reactions unconditionally during the conditioning trial. In Experiment 3a (lateral ventricle), the mixed factors anova of hedonic reactions revealed significant effects of treatment drug, F (3, 47) = 6.9; P < 0.001, trial, F (1, 47) = 56.7; P < 0.001 (conditioning drug × trial), F (1, 47) = 12.5; P < 0.001 and an (treatment drug × conditioning drug × trial) interaction, F (3, 47) = 2.8; P < 0.05. A separate 4 × 2 anova for the conditioning trial in Experiment 3a (lateral ventricles) revealed a significant effect of pretreatment condition, F (3, 47) = 4.1; P < 0.025; Bonferroni post hoc pairwise comparison tests revealed only that rats pretreated with vehicle displayed significantly more hedonic reactions than did the rats pretreated with any dose of AM251 (P < 0.05). The 4 by 2 anova for the test trial revealed significant effects of treatment drug, F (3, 47) = 7.0; P < 0.001, conditioning drug, F (1, 47) = 9.7; P < 0.01 and an (treatment drug × conditioning drug) interaction, F (3, 47) = 5.7; P < 0.01. Bonferroni post hoc pairwise comparison tests revealed that the group Vehicle-Saline displayed significantly more (P < 0.01) hedonic reactions than any other group, which did not differ from each other. In Experiment 3b (4th ventricle), the mixed factors anova of hedonic reactions revealed significant effects of conditioning drug, F (1, 34) = 5.5; P < 0.025, trial, F (1, 34) = 39.7; P < 0.001, (conditioning drug × trial), F (1, 34) = 4.8; P < 0.05 and (treatment drug × conditioning drug × trial), F (2, 34) = 3.4; P < 0.05. Separate 3 by 2 anovas of the hedonic reactions for the conditioning and test trials, revealed only significant effects on the TR test trial of treatment drug, F (2, 34) = 7.5; P < 0.01, conditioning drug, F (1, 34) = 19.4; P < 0.01 and an (treatment drug × conditioning drug) interaction, F (2, 34) = 7.0; P < 0.01. Bonnferoni post hoc pairwise comparison tests revealed that on the test trial the group Vehicle-Saline displayed significantly more hedonic reactions than any other group (P < 0.01), but the other groups did not significantly differ from one another.
Discussion
Systemic administration of a low dose (1.25 mg·kg−1) of the CB1 antagonist/inverse agonist, AM251, that does not produce gaping on its own (McLaughlin et al., 2005), potentiated conditioned gaping reactions elicited by a LiCl-paired saccharin solution in rats, as has been reported with another inverse agonist, rimonabant (Parker et al., 2003). On the other hand, systemic injections of much higher doses (up to 8 mg·kg−1) of the CB1 antagonists with no inverse agonist action, AM6545 (peripherally restricted; see Makriyannis et al., 2009; Cluny et al., 2010; Tam et al., 2010) and AM6527 (peripherally and centrally active; Sink et al., 2009a), did not potentiate LiCl- induced conditioned gaping in rats. At the same range of doses both AM6545 (Makriyannis et al., 2009; Cluny et al., 2010) and AM6527 (Sink et al., 2009a) have been shown to suppress feeding and reduce body weight in rats.
Our results suggest that in the presence of a toxin, inverse agonism of the CB1 receptor, enhances the nausea produced by that toxin. This effect on LiCl-induced nausea may be peripherally mediated, because icv administration of AM251 (even at a dose 1/10th that of the peripheral dose) either to the lateral ventricle or the 4th ventricle did not produce a similar potentiation of conditioned gaping reactions. Recent evidence suggests that differential effects on feeding occur with administration of a melanocortin antagonist (Fan et al., 2004) or an oxytocin antagonist (Blevins et al., 2004) into the 3rd or the 4th ventricles. As brainstem sites have been shown to be important for the emetic reaction in ferrets (e.g.,Van Sickle et al., 2001) and LiCl-induced conditioned gaping (Eckel and Ossenkopp, 1996) and taste avoidance (Ritter et al., 1980) in rats, the 4th ventricle injections were expected to produce a greater effect on LiCl-induced potentiated nausea than the lateral ventricle injections. However, neither infusions of AM251, at doses as high as 1/10th the peripheral dose, to the lateral or the 4th ventricle produced enhancement of LiCl-induced conditioned gaping reactions. Interestingly, when LiCl is administered to the lateral ventricles in rats, it is also ineffective in producing conditioned taste avoidance (Smith, 1980).
Given that 5-HT3 receptor antagonists, such as ondansetron, are the most commonly employed anti-emetic agents to treat the side effects of emesis and nausea in cancer chemotherapy, and that ondansetron blocks LiCl-induced conditioned gaping in rats (Limebeer and Parker, 2000), it is conceivable that the interaction of peripheral CB1 receptor inverse agonism and 5-HT3 receptor agonism mediated the potentiated aversion. Cannabinoid agonists have been reported to inhibit the activity of 5-HT3 receptors on the rat nodose ganglion (Fan, 1995), an effect that may be mediated by allosteric modulation of the 5-HT3 receptor by the cannabinoid agonist (Barann et al., 2002). CB1 receptors are expressed in the nodose ganglion and largely transported to peripheral terminals of the vagus (Partosoedarso et al., 2003), providing another potential site of interaction between 5-HT3 and CB1 receptors in the regulation of nausea and vomiting. It has also been suggested that cannabinoids may act on the interneurons synapsing with the vagal motor neurons (Hornby and Prouty, 2004). The vagal afferents project to the NTS, clearly a site of anti-emetic action of CB1 receptor agonists (Van Sickle et al., 2003), and from here information is disseminated to higher regions of the brain including the parabrachial nucleus, hypothalamus, amygdala and insular cortex (Andrews and Sanger, 2002). The generation of the sensation of nausea is most likely cortically driven (insular cortex; Penfield and Faulk, 1955), but the present data suggest that the initial site of action of the CB1 receptor inverse agonist in enhancing LiCl-induced nausea may be in the periphery. This effect would be expected to be blocked by subdiaphramatic vagotomy. Future studies aimed at evaluating the effect of systemic pretreatment with AM251 on LiCl-induced potentiated conditioned gaping in vagotomized rats would be valuable in testing this hypothesis.
On the other hand, it is also conceivable that the action of AM251 on central CB1 receptors located distal from the cerebral ventricles is responsible for the potentiation of LiCl-induced nausea. One candidate region is the insular cortex and there is considerable evidence that the insular cortex regulates the sensation of nausea. Electrical stimulation of the insular cortex produces vomiting in cats (Kaada, 1951) and humans (Catenoix et al., 2008), as well as a sensation of nausea in humans (Fiol et al., 1988; Penfield and Faulk, 1955). Contreras et al. (2007) recently reported that inactivation of the interoceptive insula attenuated LiCl-induced malaise in rats as measured by flattened ‘lying on belly’ postures (Parker et al., 1984), which was also attenuated by AP lesions (Bernstein et al., 1992) and ondansetron pretreatment (unpublished observations). Finally, ablation of the insular cortex (Kiefer and Orr, 1992), but not the amygdala (Rana and Parker, 2008), prevents LiCl-induced conditioned gaping reactions in rats (Kiefer and Orr, 1992). Indeed, recent human evidence indicates that the insular cortex, but not the amygdala, is activated by scenes of disgusting food (Calder et al., 2007).
Depletion of forebrain 5-HT by 5,7-dihydroxytryptamine (5,7-DHT) lesions of the dorsal and median raphe nuclei completely blocks LiCl-induced conditioned gaping reactions in rats (Limebeer et al., 2004). Therefore, it is likely that the action of 5-HT in forebrain regions, most likely the insular cortex, is necessary for the sensation of nausea. CB1 receptors are found within this region (Tsou et al., 1998) and they are co-expressed with 5-HT3 receptors in the insular cortex (Morales et al., 2004). As well, CB1 receptors have been shown to be expressed on 5-hydroxytryptaminergic neurons (Lau and Schloss, 2008) which suggests that one potential mechanism of the regulation of nausea by cannabinoid agonists is in the suppression of the release of 5-HT (Nakazi et al., 2000). Indeed, 5-HT has been shown to evoke endocannabinoid release, which retrogradely suppresses excitatory synapses (Best and Regeher, 2008). Kwiatkowska et al. (2004) reported that combined sub-threshold low doses of Δ9- THC and ondansetron (that individually were ineffective) completely suppressed cisplatin-induced vomiting in shrews, suggesting a synergistic effect. The potentiation of LiCl-induced nausea by AM251 could therefore be mediated by inverse agonism of the presynaptic CB1 receptors on the terminals of 5-hydroxytryptaminergic neurons, thereby elevating the release of 5-HT in the insular cortex. Future studies that deliver AM251 directly to the insular cortex are needed to test this hypothesis.
Although central AM251 did not potentiate LiCl-induced conditioned gaping reactions in rats, it did unconditionally suppress saccharin palatability when delivered to the lateral ventricle but not to the 4th ventricle, as was also seen with systemic AM251 in Experiment 1 (replicating Jarrett et al., 2007). As icv administration to the 4th ventricle would be less likely to activate forebrain mechanisms than icv administration to the lateral ventricle (Blevins et al., 2004; Fan et al., 2004; Sink et al., 2009b), the palatability shift produced by AM251 during conditioning, is most likely to be mediated by its action on forebrain mechanisms. Indeed, recent evidence suggests that direct administration of anandamide to the medial shell of the nucleus of the accumbens potently doubled the number of positive ‘liking’ facial reactions that sucrose taste elicits from rats (Mahler et al., 2007). The unconditional attenuation of hedonic reactions to intraoral delivery of saccharin solution that was produced by AM251 when administered to the lateral ventricle or systemically may also reflect anhedonia, a depression-like effect in rats (Hill and Gorzalka, 2005). This anhedonia was not produced by systemic administration of the neutral CB1 receptor antagonists, AM6545 or AM6527, even at doses as high as 8 mg·kg−1 (6 times higher than the effective systemic dose of AM251, 1.25 mg·kg−1). As the debilitating side effect of depression prevented the therapeutic use of the inverse agonist, rimonabant, in the treatment of obesity, the finding that the neutral CB1 receptor antagonists did not attenuate hedonic reactions elicited by saccharin solution at doses that suppress feeding and weight gain (Sink et al., 2008; Cluny et al., 2010; Tam et al., 2010), suggest that these compounds may have therapeutic potential.
When administered systemically or centrally to either the lateral ventricle or the 4th ventricle, AM251 also conditionally suppressed the palatability of saccharin solution on the subsequent drug-free test trial. This conditioned suppression of hedonic reactions, however, was not accompanied by a conditioned enhancement of gaping reactions. Such a conditional shift in hedonic reactions, does not appear to be a specific marker of nausea in rats, because even non-emetic treatments, such as amphetamine and cocaine, produce such a shift, as well as taste avoidance (Parker, 1995). As suppressed ingestive behaviour in the TR test follows a pattern to that of taste avoidance in a consumption test, it would be interesting to determine if central administration of AM251 would produce conditioned taste avoidance in rats, which may reflect conditioned fear rather than conditioned nausea in this non-emetic species (Parker et al., 2008).
One of the primary therapeutic targets of CB1 receptor antagonists is the suppression of feeding and food-motivated behaviours; however, the locus of action of these agents is controversial. CB1 receptor agonists injected into the hypothalamic nuclei or into the nucleus accumbens have been shown to produce hyperphagia (Williams and Kirkham, 1999; Jamshidi and Talyor, 2001; Verty and Mallet, 2005; Soria-Gomez et al., 2007) and these effects were blocked by CB1 receptor inverse agonists. Although some investigators have reported that forebrain injections of CB1 receptor inverse agonists produce anorexia (Werner and Koch, 2003; Verty et al., 2004a,b), others have reported that feeding-related actions of these compounds may be peripherally mediated (Gomez et al., 2002; Sink et al., 2009b). Most recently, Sink et al. (2009b) reported no effect on feeding-related behaviours, of icv administration of AM251 to the lateral ventricles at doses up to 1/10 the peripherally effective dose. As CB1 receptor inverse agonists produce nausea and depression in humans (see Janero and Makriyannis, 2009), there is considerable interest in the development of CB1 receptor antagonists without central inverse agonist properties. Indeed, the CB1 receptor specific neutral antagonist, AM6545, with limited CNS penetration dose-dependently reduced food intake and induced sustained reduction in body weight in mice and rats, and did not produce conditioned gaping at effective doses (Cluny et al., 2010). The effect on food intake was maintained in rats with a complete subdiaphragmatic vagotomy. The results reported here, suggest that neutral CB1 receptor antagonists, AM6545 and AM6527, neither enhanced the nauseating effects of LiCl, nor produced anhedonia in the TR test as did the inverse agonist AM251. The potentiation of LiCl-induced nausea by inverse agonism of the CB1 receptor by AM251 may be peripherally mediated or centrally mediated by its action on CB1 receptors distal to the cerebral ventricles.
Acknowledgments
This research was supported by an NSERC Post-doctoral Fellowship to C. Limebeer, NSERC Operating grants to L. Parker (NSERC 92057) and to K.P Ossenkopp and DA7215, DA3801, DA009158 NIH Awards to A. Makriyannis.
Glossary
Abbreviations
- AP
area postrema
- BBB
blood–brain barrier
- DMNX
dorsal motor nucleus of the vagus
- icv
intracerebroventricular
- LiCl
lithium chloride
- M6G
morphine 6-glucuronide
- NTS
nucleus of the solitary tract
- TR
Taste Reactivity
Conflicts of interest
There are no conflicts of interest among any authors.
Supplemental material
Supporting Information: Teaching Materials; Figs 1–5 as PowerPoint slide.
References
- Alexander SPH, Mathie A, Peters JA. Guide to Receptors and Channels (GRAC), 4th edn. Br J Pharmacol. 2009;158(Suppl. 1):S1–S254. doi: 10.1111/j.1476-5381.2009.00499.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrews PLR, Horn CC. Signals for nausea and emesis: implications for models of upper gastrointestinal diseases. Auton Neurosci. 2006;125:100–115. doi: 10.1016/j.autneu.2006.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrews PLR, Sanger GJ. Abdominal vagal afferent neurones: an important target for the treatment of gastrointestinal dysfunction. Cur Opin Pharmacol. 2002;2:650–656. doi: 10.1016/s1471-4892(02)00227-8. [DOI] [PubMed] [Google Scholar]
- Barann M, Molderings G, Brüss M, Bönisch H, Urban BW, Göthert M. Direct inhibition by cannabinoids of human 5-HT3A receptors: probable involvement of an allosteric modulatory site. Br J Pharmacol. 2002;137:589–596. doi: 10.1038/sj.bjp.0704829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernstein IL, Chavez M, Allen D, Taylor EM. Area postrema mediation of physiological and behavioral effects of lithium chloride in the rat. Br Res. 1992;575:132–137. doi: 10.1016/0006-8993(92)90432-9. [DOI] [PubMed] [Google Scholar]
- Best AR, Regeher WG. Serotonin evokes endocannabinoid release and retrogradely suppresses excitatory synapses. J Neurosci. 2008;28:6508–6515. doi: 10.1523/JNEUROSCI.0678-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blevins JE, Schwartz MW, Baskin DG. Evidence that paraventricular nucleus oxytocin neurons link hypothalamic leptin action to caudal brain stem nuclei controlling meal size. Am J Physiol Regul Integr Comp Physiol. 2004;287:R87–R96. doi: 10.1152/ajpregu.00604.2003. [DOI] [PubMed] [Google Scholar]
- Calder AK, Beaver JD, Davis MH, van Ditzhuijzen J, Keane J, Lawrence AD. Disgust sensitivity predicts the inusla and pallidal response to pictures of disgusting foods. Eur J Neurosci. 2007;25:3422–3428. doi: 10.1111/j.1460-9568.2007.05604.x. [DOI] [PubMed] [Google Scholar]
- Catenoix H, Isnard J, Guénot M, Petit J, Remy C, Mauguiére F. The role of the anterior insular cortex in ictal vomiting: a stereotactic electroencephalography study. Epilepsy Behav. 2008;13:560–563. doi: 10.1016/j.yebeh.2008.06.019. [DOI] [PubMed] [Google Scholar]
- Chambers AP, Vemuri VK, Peng Y, Wood JT, Olszewska T, Pittman QJ, et al. A neutral CB1 receptor antagonist reduces weight gain in rats. Am J Physiol Regul Integr Comp Physiol. 2007;293:R2185–R2193. doi: 10.1152/ajpregu.00663.2007. [DOI] [PubMed] [Google Scholar]
- Cluny NL, Vemuri VK, Chambers AP, Limebeer CL, Bedard H, Wood JT, et al. A novel peripherally restricted, cannabinoid 1 (CB1) receptor antagonist, AM6545, reduces food intake and body weight, but does not cause malaise, in rodents. Br J Pharmacol. 2010 doi: 10.1111/j.1476-5381.2010.00908.x. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Contreras M, Ceric F, Torrealba F. Inactivation of the interoceptive insula disrupts drug craving and malaise induced by lithium. Science. 2007;318:655–658. doi: 10.1126/science.1145590. [DOI] [PubMed] [Google Scholar]
- Cross-Mellor SK, Ossenkopp KP, Piomelli D, Parker LA. Effects of the FAAH inhibitor, URB597, and anandamide on lithium-induced taste reactivity responses: a measure of nausea in rats. Psychopharmacology. 2007;190:135–143. doi: 10.1007/s00213-006-0589-7. [DOI] [PubMed] [Google Scholar]
- Darmani NA. The cannabinoid CB1 receptor antagonist SR 141716A reverses the antiemetic and motor depressant actions of WIN 55, 212-2. Eur J Pharm. 2001;430:49–58. doi: 10.1016/s0014-2999(01)01355-3. [DOI] [PubMed] [Google Scholar]
- Darmani NS, Johnson CJ. Central and peripheral mechanisms contribute to the antiemetic actions of delta-9-tetrahydrocannabinol against 5-hydroxytryptophan-induced emesis. Eur J Pharmacol. 2004;488:201–212. doi: 10.1016/j.ejphar.2004.02.018. [DOI] [PubMed] [Google Scholar]
- Eckel LA, Ossenkopp KP. Area postrema mediates the formation of rapid conditioned palatability shifts in area postrema lesioned rats. Behav Neurosci. 1996;110:202–212. [PubMed] [Google Scholar]
- Erb S, Funk D, Lê AD. Prior, repeated exposure to cocaine potentiates locomotor responsivity to central injections of corticotrophin-releasing factor (CRF) in rats. Psychopharmacology. 2003;170:383–389. doi: 10.1007/s00213-003-1556-1. [DOI] [PubMed] [Google Scholar]
- Fan P. Cannabinoid agonists inhibit the activation of 5-HT3 receptors in rat nodose ganglion neurons. J Neurophysiol. 1995;73:907–910. doi: 10.1152/jn.1995.73.2.907. [DOI] [PubMed] [Google Scholar]
- Fan W, Ellacott KLJ, Halatchev IG, Takahashi K, Yu P, Cone RD. Cholecystokinin-mediated suppression of feeding involves the brainstem melanocortin system. Nat Neurosci. 2004;7:335–336. doi: 10.1038/nn1214. [DOI] [PubMed] [Google Scholar]
- Fiol M, Leppik IE, Mireless R, Maxwell R. Ictus emeticus and the insular cortex. Epilepsy Res. 1988;2:127–131. doi: 10.1016/0920-1211(88)90030-7. [DOI] [PubMed] [Google Scholar]
- Garcia J, Hankins WG, Rusiniak KW. Behavioral regulation of the milieu interne in men and rat. Science. 1974;185:824–831. doi: 10.1126/science.185.4154.824. [DOI] [PubMed] [Google Scholar]
- Gomez R, Navarro M, Ferrer B, Trigo JM, Bilbao A, Del Arco I, et al. A peripheral mechanism for CB1 cannabinoid receptor-dependent modulation of feeding. J Neurosci. 2002;22:9612–9617. doi: 10.1523/JNEUROSCI.22-21-09612.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grant VL. Do conditioned taste aversions result from activation of emetic mechanisms? Psychopharmacology. 1987;93:405–416. doi: 10.1007/BF00207227. [DOI] [PubMed] [Google Scholar]
- Hill MN, Gorzalka BB. Is there role for the endocannabinoid system in the etiology and treatment of melancholic depression? Behav Pharmacol. 2005;16:333–352. doi: 10.1097/00008877-200509000-00006. [DOI] [PubMed] [Google Scholar]
- Hornby PJ, Prouty SM. Involvement of cannabinoid receptors in gut motility and visceral perception. Br J Pharmacol. 2004;141:1335–1345. doi: 10.1038/sj.bjp.0705783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iversen LL. The Science of Marijuana. New York: Oxford University Press; 2000. [Google Scholar]
- Jamshidi N, Talyor DA. Anandamide administration itno the ventromeidal hypothalamus stimulates appetite in rats. Br J Pharmacol. 2001;134:1151–1154. doi: 10.1038/sj.bjp.0704379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janero DR, Makriyannis A. Cannabinoid receptor antagonists: pharmacological opportunities, clinical experience, and translational prognosis. Exp Opin Emerg Drugs. 2009;14:1–23. doi: 10.1517/14728210902736568. [DOI] [PubMed] [Google Scholar]
- Jarrett MM, Scantlebury J, Parker LA. The effect of CB1 antagonist, AM251, on sucrose and quinine palatability: direct measurement through the taste reactivity test. Physiol Behav. 2007;90:425–430. doi: 10.1016/j.physbeh.2006.10.003. [DOI] [PubMed] [Google Scholar]
- Kaada BR. Somato-motor, autonomic and electrocorticographic responses to electrical stimulation of rhinencephalic and other structures in primates, cat, and dog; a study of responses from the limbic, subcallosal, orbito-insular, piriform and temporal cortex, hippocampus-fornix and amygdala. Acta Physiol Scand Suppl. 1951;24:1–262. [PubMed] [Google Scholar]
- Kiefer SW, Orr MS. Taste avoidance, but not aversion, learning in rats lacking gustatory cortex. Behav Neurosci. 1992;106:140–146. doi: 10.1037//0735-7044.106.1.140. [DOI] [PubMed] [Google Scholar]
- Kwiatkowska M, Parker LA, Burton P, Mechoulam R. A comparative analysis of the potential of cannabinoids and ondansetron to suppress cisplatin-induced emesis in the Suncus murinus (house musk shrew) Psychopharmacology. 2004;174:254–259. doi: 10.1007/s00213-003-1739-9. [DOI] [PubMed] [Google Scholar]
- Lau T, Schloss P. The cannabinoid CB1 receptor is expressed on serotonergic and dopaminergic neurons. Eur J Pharm. 2008;578:137–141. doi: 10.1016/j.ejphar.2007.09.022. [DOI] [PubMed] [Google Scholar]
- Limebeer CL, Parker LA. The anti-emetic drug, ondansetron, interferes with lithium-induced conditioned rejection reactions, but not lithium-induced conditioned taste avoidance. J Exp Psychol Anim Behav Process. 2000;26:371–384. doi: 10.1037//0097-7403.26.4.371. [DOI] [PubMed] [Google Scholar]
- Limebeer CL, Parker LA, Fletcher P. 5,7-Dihydroxytryptamine lesions of the dorsal and median raphe nuclei interfere with lithium-induced conditioned gaping, but not conditioned taste avoidance, in rats. Behav Neurosci. 2004;118:1391–1399. doi: 10.1037/0735-7044.118.6.1391. [DOI] [PubMed] [Google Scholar]
- Mahler SV, Smith KS, Berridge KC. Endocannabinoid hedonic hotspot for sensory pleasure: anandamide in the nucleus accumbens shell enhances ‘liking’ of a sweet reward. Neuropsychopharmacology. 2007;32:2267–2278. doi: 10.1038/sj.npp.1301376. [DOI] [PubMed] [Google Scholar]
- Makriyannis A, Vemuri VK, Thotapally R, Olszewska T. 2009. International Patent Application No. PCT//US09//01054.
- McLaughlin PJ, Winston KM, Limebeer CL, Parker LA, Makriyannis A, Salamone JD. The cannabinoid antagonist AM251 produces food avoidance and behaviors associated with nausea but does not impair feeding efficiency in rats. Psychopharmacology. 2005;180:286–293. doi: 10.1007/s00213-005-2171-0. [DOI] [PubMed] [Google Scholar]
- Morales M, Wang S, Diaz-ruiz O, Jho DH. Cannabinoid CB1 receptor and serotonin 3 receptor subunit A (5-HT3A) are co-expressed in GABA neurons in the telencephalon. J Comp Neurol. 2004;468:205–216. doi: 10.1002/cne.10968. [DOI] [PubMed] [Google Scholar]
- Nakazi M, Bauer U, Nickel T, Kathmann M, Schlicker E. Inhibition of serotonin release in the mouse brain via presynaptic cannabinoid CB1 receptors. Naunyn Schmiedebergs Arch Pharmacol. 2000;361:19–24. doi: 10.1007/s002109900147. [DOI] [PubMed] [Google Scholar]
- Parker LA. Rewarding drugs produce taste avoidance, but not taste aversion. Neurosci Biobehav Rev. 1995;19:143–151. doi: 10.1016/0149-7634(94)00028-y. [DOI] [PubMed] [Google Scholar]
- Parker LA. Taste avoidance and taste aversion: Evidence for two different processes. Learn Behav. 2003;31:165–172. doi: 10.3758/bf03195979. [DOI] [PubMed] [Google Scholar]
- Parker LA, Mechoulam R. Cannabinoid agonists and an antagonist modulate lithium-induced conditioned gaping in rats. Integr Physiol Behav Sci. 2003;38:134–146. doi: 10.1007/BF02688831. [DOI] [PubMed] [Google Scholar]
- Parker LA, Hills K, Jensen K. Behavioral conditioned responses elicited by lithium- and amphetamine-paired contextual cues. Anim Learn Behav. 1984;12:307–315. [Google Scholar]
- Parker LA, Mechoulam R, Schlievert C, Abbott LA, Fudge ML, Burton P. Cannabinoid agonists attenuate and a cannabinoid antagonist potentiates lithium-induced conditioned rejection reactions in a rat model of nausea. Psychopharmacology. 2003;166:156–162. doi: 10.1007/s00213-002-1329-2. [DOI] [PubMed] [Google Scholar]
- Parker LA, Rana SA, Limebeer CL. Conditioned nausea in rats: Assessment by conditioned disgust reactions, rather than conditioned taste avoidance. Can J Exp Psychol. 2008;62:198–209. doi: 10.1037/a0012531. [DOI] [PubMed] [Google Scholar]
- Partosoedarso ER, Abrahams TP, Scullion RT, Moerschabaecher JM, Hornby PJ. Cannabinoid 1 receptor in the dorsal vagal complex modulates lower oesophageal sphincter relaxation in ferrets. J Physiol. 2003;550:149–158. doi: 10.1113/jphysiol.2003.042242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paxinos G, Watson C. The Rat Brain. 4th edn. New York: Academic Press; 1998. [Google Scholar]
- Penfield W, Faulk ME. The insula: further observations on its function. Brain. 1955;78:445–470. doi: 10.1093/brain/78.4.445. [DOI] [PubMed] [Google Scholar]
- Pertwee RG. Cannabinoids and the gastrointestinal tract. Gut. 2001;48:859–867. doi: 10.1136/gut.48.6.859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pertwee RC. Inverse agonism and neutral antagonism at cannabinoid CB1 receptors. Life Sci. 2005;76:1307–1324. doi: 10.1016/j.lfs.2004.10.025. [DOI] [PubMed] [Google Scholar]
- Rana SA, Parker LA. Differential effects of neurotoxic lesions of the basolateral amygdale and the central nucleus of the amygdale on lithium-induced conditioned disgust reactions and conditioned taste avoidance. Behav Brain Res. 2008;189:284–297. doi: 10.1016/j.bbr.2008.01.005. [DOI] [PubMed] [Google Scholar]
- Ritter S, McGlone JJ, Kelley KW. Absence of lithium-induced taste aversion after area postrema lesion. Brain Res. 1980;201:501–506. doi: 10.1016/0006-8993(80)91061-6. [DOI] [PubMed] [Google Scholar]
- Rock EM, Benzaquen J, Limebeer CL, Parker LA. Potential of the rat model of conditional gaping to detect nausea produced by rolipram, a phosphodiesterase-4 (PDE4) inhibitor. Pharmacol Biochem Behav. 2009;91:537–541. doi: 10.1016/j.pbb.2008.09.005. [DOI] [PubMed] [Google Scholar]
- Salamone JD, McLaughlin PJ, Sink K, Makriyannis A, Parker LA. Cannabinoid CB1 receptor inverse agonists and neutral antagonists: Effects on food intake, food-reinforced behavior and food aversions. Physiol Behav. 2007;91:383–388. doi: 10.1016/j.physbeh.2007.04.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simoneau II, Hamza MS, Mata HP, Siegel EM, Vanderah TW, Porreca F, et al. The cannabinoid agonist WIN55, 212-2 suppresses opioid-induced emesis in ferrets. Anesthesiology. 2001;94:882–887. doi: 10.1097/00000542-200105000-00029. [DOI] [PubMed] [Google Scholar]
- Sink KS, McLaughlin PJ, Wood JA, Brown C, Fan P, Vemuri VK, et al. The novel cannabinoid CB1 receptor neutral antagonist, AM4113, suppresses food intake and food-reinforced behaviour but does not induce signs of nausea in rats. Neuropsychopharmacology. 2008;33:946–955. doi: 10.1038/sj.npp.1301476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sink KS, Vemuri VK, Wood J, Makriyannis A, Salamone JD. Oral bioavailability of the novel cannabinoid CB1 antagonist AM6527: effects on food-reinforced behaviour and comparisons with AM4113. Pharmacol Biochem Behav. 2009a;91:303–306. doi: 10.1016/j.pbb.2008.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sink KS, Segovia KN, Nunes EJ, Collins LE, Vemuri VK, Thakur G, et al. Intracerebroventricular administration of cannabinoid CB1 receptor antagonists AM251 and AM4113 fails to alter food-reinforced behaviour in rats. Psychopharmacology. 2009b;206:223–232. doi: 10.1007/s00213-009-1602-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith DF. Central and peripheral effects of lithium on conditioned taste aversions in rats. Psychopharmacology. 1980;68:315–317. doi: 10.1007/BF00428123. [DOI] [PubMed] [Google Scholar]
- Soria-Gomez E, Matias I, Rueda-Orozco PE, Cisneros M, Petrosino S, Navarro L, et al. Pharmacological enhancement of the endocannabinoid system in the nucleus accumbens shell stimulates food intake and increases c-fos expression in the hypothalamus. Br J Pharmacol. 2007;151:1109–1116. doi: 10.1038/sj.bjp.0707313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tam J, Vemuri VK, Liu J, Bátkai S, Mukhopadhyay B, Gadlewski G, et al. Peripheral CB, cannabinoid receptor blockade improves cardiometabolic risk in mouse models of obesity. J Clin Investigation. 2010 doi: 10.1172/JCI42551. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsou K, Brown S, Sanudo-Pena MC, Mackie K, Walker JM. Immunohistocmehical distribution of cannabinoid CB1 receptors in the rat central nervous system. Neuroscience. 1998;83:393–411. doi: 10.1016/s0306-4522(97)00436-3. [DOI] [PubMed] [Google Scholar]
- Tuerke KJ, Leri F, Parker LA. Antidepressant-like effects of paroxetine are produced by lower doses than those which produce nausea. Pharmacol Biochem Behav. 2009;93:190–195. doi: 10.1016/j.pbb.2009.05.011. [DOI] [PubMed] [Google Scholar]
- Van Sickle MD, Oland LD, Ho W, Hillard CJ, Mackie K, Davison JJ, et al. Cannabinoids inhibit emesis through CB1 receptors in the brainstem of the ferret. Gastroenterology. 2001;121:767–767. doi: 10.1053/gast.2001.28466. [DOI] [PubMed] [Google Scholar]
- Van Sickle MD, Oland LD, Mackie K, Davison JS, Sharkey KA. Δ9-Tetrahydrocannabinol selectively acts on CB1 receptors in specific regions of dorsal vagal complex to inhibit emesis in ferrets. Am J Physiol Gastrointest Liver Physiol. 2003;285:G566–G576. doi: 10.1152/ajpgi.00113.2003. [DOI] [PubMed] [Google Scholar]
- Verty AN, Mallet PE. Paraventricular hypothalamic CB1 cannabinoid receptors are involved in the feeding stimulatory effects of delta9-tetrahydrocannabinol. Neuropharmacology. 2005;49:1101–1109. doi: 10.1016/j.neuropharm.2005.03.025. [DOI] [PubMed] [Google Scholar]
- Verty AN, McFarlane JR, McGregor IS, Mallet PE. Evidence for an interaction between CB1 Cannabinoid and oxytocin receptors in food and water intake. Neuropharmacology. 2004a;47:593–603. doi: 10.1016/j.neuropharm.2004.06.002. [DOI] [PubMed] [Google Scholar]
- Verty AN, McFarlane JR, McGregor IS, Mallet PE. Evidence for an interaction between CB1 cannabinoid and melanocortin MCR-4 receptors in regulating food intake. Endocrinology. 2004b;145:3224–3231. doi: 10.1210/en.2004-0059. [DOI] [PubMed] [Google Scholar]
- Werner NA, Koch JE. Effects of the cannabinoid antagonists AM281 and AM630 on deprivation-induced intake in Lewis rats. Brain Res. 2003;967:290–292. doi: 10.1016/s0006-8993(02)04274-9. [DOI] [PubMed] [Google Scholar]
- Williams C, Kirkham TC. Anandamide induces overeating: mediation by central cannabinoid (CB1) receptors. Psychopharmacology. 1999;143:315–317. doi: 10.1007/s002130050953. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.