

Abstract
The right temporoparietal junction (R TPJ) is involved in stimulus-driven attentional control in response to the appearance of an unexpected target or a distractor that shares features with a task-relevant target. An unresolved question is whether these responses in R TPJ are due simply to the presence of a stimulus that is a potential target, or instead responds to any task-relevant information. Here, we addressed this issue by testing the sensitivity of R TPJ to a perceptually salient, non-target stimulus -- a contextual cue. Although known to be a non-target, the contextual cue carried probabilistic information regarding the presence of a target in the opposite visual field. The contextual cue was therefore always of potential behavioral relevance, but only sometimes paired with a target. The appearance of the contextual cue alone increased activation in R TPJ, but more so when it appeared with a target. There was also greater connectivity between R TPJ and a network of attentional control and decision areas when the contextual cue was present. These results demonstrate that R TPJ is involved in the stimulus-driven representation of task-relevant information that can be used to engage an appropriate behavioral response.
Introduction
In order to behave efficiently in a complex environment, it is necessary to not only direct top-down attention to task-relevant information, but also to integrate sensory events of contextual relevance. Contextual information may not always provide sufficient information to immediately execute goal-relevant actions, but may nevertheless predict the occurrence of events and behaviors related to the current goal. Successful use of such contextual information requires the rapid integration of previously unattended stimuli with ongoing top-down selection processes. In this study, we explored the role of TPJ in representing contextual information that could be used to bias mechanisms of top-down information selection.
The right temporo-parietal junction (R TPJ) has consistently been found to be activated by unexpected stimulus events of behavioral relevance. Behavioral relevance has typically been defined by task-relevant target properties. For example, in variations of the Posner cueing paradigm, (Posner 1980), R TPJ and the right inferior frontal gyrus (IFG) were activated specifically by “invalid” targets that appeared in an unexpected location, (e.g. Corbetta et al 2000; Corbetta et al 2008; Indovina & Macaluso 2007; Kincade et al 2005; Natale et al 2009; Shulman et al 2007; Vossel et al 2009). Similarly, R TPJ is sensitive to non-targets that share the target color (Hu et al 2009; Natale et al 2009; Serences et al 2005)or category (Hampshire et al 2007), and task cues that indicate a new target location (Shulman et al 2009). Unexpected stimuli in these paradigms produced shifts in spatial attention, but shifts of attention are not a necessary condition for R TPJ activation: R TPJ also responds to oddball targets within an attended stream (Marois et al 2000), cues indicating low frequency target locations (Shulman et al 2009) and sudden changes in continuous sensory events that violate expectations (Downar et al 2000; 2002). While some of these studies also report left TPJ activation under similar conditions, R TPJ activation is generally more robust and reliable. R TPJ activation is stimulus-driven, but only in response to unexpected stimuli of potential behavioral relevance. This has led to the hypothesis that R TPJ and the ventral attentional network acts as a filter that selectively gates stimulus information based on task goals (Corbetta et al 2008; Shulman et al 2007)
In contrast, stimulus-driven attentional capture by perceptually salient stimuli that are completely task-irrelevant do not activate R TPJ. Instead, salient stimuli of this kind tend to interfere with task processing and activate the intraparietal sulcus (IPS) and frontal eye fields (FEF) (e.g. Corbetta et al 2008; Indovina & Macaluso 2007; Kincade et al 2005). IPS and FEF form a dorsal attentional network that is thought to reflect the current distribution of spatial attention across the visual field, (e.g., Bisley & Goldberg 2003; Goldberg et al 2006; Gottlieb 2007), and to control the voluntary orientation of attention (e.g. Corbetta 1998; Corbetta et al 2000; Geng et al 2006; Serences & Yantis 2007; Yantis et al 2002). Sensory stimuli that are completely irrelevant to current goals but yet capture attention are represented within the dorsal attentional priority map, and do not engage R TPJ and the ventral attentional network (e.g., Geng & Mangun 2009).
These previous studies suggest that the mechanisms of stimulus-driven attention depend on the task-relevance of the stimulus. TPJ and the ventral attentional network are selectively activated by unexpected task-relevant stimuli, but task-relevance has so far been defined primarily by information that is directly related to target processing (e.g., targets, non-targets with target features, or target-related cues). Thus, while it has been hypothesized that TPJ is sensitive generally to any behaviorally relevant stimulus that leads to a change in the focus of attention, current studies have not addressed whether or not TPJ is sensitive to non-target contextual information that may facilitate the current behavioral goal.
In this study, we define task-relevance flexibly by manipulating the statistical relationship between a perceptually salient contextual cue and the appearance of a target stimulus. The contextual cue and target appeared in opposite visual fields within bilateral streams of “standard” stimuli (see Figure 1). The contextual cue was only partially predictive of the appearance of the target, which allowed us to dissociate responses to the target alone, the salient contextual cue alone, and the coincidence of the two. The salient stimulus was therefore always of potential behavioral relevance, but only sometimes paired with a target. This allowed us to dissociate activation related to the detection of the potentially informative stimulus and its actual task-relevance.
Figure 1.

Illustration of experimental conditions. Standard stimuli were presented in rapid bilateral visual streams. The central fixation indicated which stimulus stream targets would appear in and whether the target discrimination would be easy or hard (e.g, the right-sided blue rectangle at fixation in the illustration above instructed subjects to attend to the right-sided stream for easy targets). There were three event types of interest within each block: 1) target alone trials in which a target appeared in the attended stream and a standard in the unattended stream; 2) target + contextual cue trials in which a target and a contextual cue appeared simultaneously; and 3) contextual cue alone trials in which the stimulus in the unattended stream increased in luminance and a standard stimulus appeared in the attended location.
Materials and Methods
Subjects
Imaging and behavioral data were collected from 16 subjects (8 females, 1 left handed). Three were excluded based on technical problems with data acquisition or excessive head motion; data from 13 subjects were entered into our analyses. All were screened for MRI compatibility and gave written informed consent in accord with the local ethics clearance as approved by NIH. All had normal or corrected-to-normal vision and none had a history of neurological or psychiatric disorder.
Experimental Design
A factorial design was used such that attention was blocked to either the left or right stimulus stream, the target was present or absent, target discrimination was hard or easy and the contextual cue was either present (i.e., a high luminance stimulus in the unattended stream) or absent (i.e., a standard stimulus). Targets were identical to standard stimuli except narrower (see Figure 1); easy targets were more narrow and therefore the change from a standard was easier to detect. Target location and difficulty were blocked and indicated by the location and color of the rectangle within the fixation square prior to the start of each block (e.g., a blue rectangle on the left indicated that an easy target would appear in the left visual stream) (Figure 1). The cue remained visible throughout the block. Blocks lasted between 12 and 18 seconds and were separated by a blank fixation period lasting 2 or 12 seconds. Each subject experienced 10 scan sessions composed of 12 blocks each.
Standard stimuli appeared simultaneously with the onset of the spatial cue and were presented bilaterally in homologous locations. The corner nearest fixation was 3.5° of horizontal and .21° of vertical visual angle from central fixation. A standard stimulus subtended .2° of horizontal visual angle and .44° of vertical visual angle, at central fixation. Standards appeared every 500 ms, lasted 100 ms and were continuously presented throughout the block in the bilateral rapid visual presentation streams. Targets appeared in the stream indicated by the location cue with 100% validity and were identical to the standard except narrower. Difficult targets subtended .12°, and easy targets, .05° of horizontal visual angle, but were identical to the standard stimulus in height and luminance (6 cd/m2). Subjects were asked to press a single button with their index finger whenever a target appeared (in the attended stream). Subjects were instructed to only respond if they were “50% or more sure that they detected a target”. Non-target contextual cues only appeared in the unattended stimulus stream and were identical to a standard except for its luminance (25 cd/m2). These high luminance contextual cues were perceptually salient to ensure its primacy in bottom-up processing compared to the less salient target. This permitted measurement of its contextual relevance (i.e., whether it facilitated or interfered with target processing).
Two targets and two contextual cues appeared in each block and they coincided once, resulting in the following three trial types of primary interest: target + contextual cue, target alone (with standard in the unattended stream), and contextual cue alone (with standard in the attended stream) (see Figure 1). Half of all targets appeared with a salient contextual cue and half of all contextual cues appeared with a target. The remaining targets and contextual cues appeared with a standard stimulus in the opposite stimulus stream. Standard stimuli appeared bilaterally every 500 ms. The inter-trial interval between trials of interest was 2 to 10 seconds, jittered in intervals of 2 seconds to allow for uniform sampling of event-related BOLD responses across the whole TR and to maintain behavioral uncertainty regarding the onset of the next target stimulus.
The most critical aspect of the design involved the predictive value of the contextual cue for target discrimination. Although the salient contextual cue only appeared with the target 50% of the time, it still provided predictive power compared to standards, only 3% of which coincided with a target (recall that standard stimuli occurred every 500 ms throughout a block of trials). Thus, the appearance of the contextual cue signaled a relative increase in likelihood of a target being present. This design allowed us to evaluate the behavioral and brain responses to a contextual cue that carries task relevant information both when it occurs with a target and when it does not.
Behavioral training was conducted outside the scanner until participants understood the task and demonstrated the ability to correctly detect targets in every block within an experimental session. Stimuli were presented via a video projector and front projection screen. The screen was viewed via a mirror system attached to the head coil. All subjects were instructed to maintain visual fixation. Eye-tracking was performed at 60 Hz using Applied Science Laboratories (ASL, Inc. Bedford, MA) model 504 with long-range remote optics.
Image and data processing
MRI data were acquired from a 3T Siemens Trio scanner (Siemens, Erlangen, Germany) equipped with an 8 channel phased array head coil. A T2*-weighted echo planar imaging (EPI) sequence was used to acquire volumes of 34 slices of 3 mm thickness (3.4 × 3.4 mm in-plane resolution) with a distance factor of 10%, every 2000 ms. Slices were axially oriented and covered the whole brain. 222 volumes were collected in each session for 10 sessions. Image data were analyzed using SPM5 (Wellcome Department of Imaging Neuroscience, London, UK; Friston et al., 1995). Prior to statistical analysis, the first four volumes were discarded to allow for T1 equilibrium effects. Images were realigned and unwarped to correct for interactions between movement and field inhomogeneities (Andersson et al., 2001), normalized to the MNI EPI template available in SPM 5, and resampled to a resolution of 2 × 2 × 2 mm. Data used in group image analyses were additionally smoothed with a three-dimensional 9 mm full-width half maximum (FWHM) Gaussian kernel.
High resolution T1-weighted structural images were acquired using a MPRAGE sequence, coregistered with each subject’s EPI images, and normalized to the MNI template brain. These were used for identifying individual anatomical landmarks for ROI selection (see below). An average structural image was created from the normalized T1-weighted images for the purpose of displaying functional results from the group.
Hemodynamic responses to stimuli in the 12 experimental conditions were modeled for each subject by stick functions convolved with a canonical hemodynamic response function. The 12 conditions were given by crossing the attended side, target difficulty, and the three combinations of target and contextual cue presence. The fourth combination where the target and contextual were absent (effectively two standard stimuli) was not separately modeled, but served as part of the implicit baseline against which responses to targets and contextual cues were estimated (e.g., Jack et al 2007; Marois et al 2000; Serences et al 2005). Misses and false alarms were modeled separately as two additional conditions. Linear contrasts of parameter estimates were estimated for each subject and combined for the group level in random-effects GLM. Behavioral data and image data extracted from individual regions of interest (ROIs) were analyzed using R software (www.r-project.org).
Eye position data
Eye position data for each subject and run were normalized to the global mean signal across the entire run to account for drift. Values exceeding a z score of 3 or greater, which could be caused by artifacts unrelated to the experimental manipulation such as blinks, or loss of data, were removed and replaced with a mean of the neighboring values. The data were then smoothed with a five-point moving average to remove remaining noise artifacts.
Region of interest (ROI) selection
Regions of interest were selected in each individual based on a combination of functional activation and anatomy; the peak activation closest to the group functional results from the contrast of target + salient contextual cue minus target alone was used in combination with the anatomical region defined by the intersection between the superior temporal sulcus the angular gyrus to identify the ROI center (see Supplemental Table 1 for individual ROI coordinates). Each ROI was a spherical volume with a radius of 2 mm. TPJ ROIs could not be reliably identified in one subject and that subject was excluded from all ROI analyses. Mean estimates were extracted for each individual corresponding to all conditions of the experiment. To address the primary experimental question, extracted parameter estimates corresponding to the contextual cue alone were compared to those for the two target present conditions using paired t-tests. Additional ANOVAS were performed on all extracted parameter estimate values to test for effects related to target side and difficulty.
Psychophysiological interactions analysis
We applied the “psychophysiological interactions” (PPI) approach (Friston 2002; Friston et al 1997) that tests for condition dependent covariation in activity between a ‘seed’ region and any other brain area, after the effects of the experimental factors in the model have been accounted for. The PPI reflects changes in the degree to which activation in one region covaries with activation in another region based on an experimental manipulation. The PPI analyses were free of any assumptions about which areas in the brain should show functional connectivity with the seed regions.
For each subject, mean-adjusted data (first eigenvariates of the timeseries) were extracted from all voxels within each left and right TPJ ROI (see above). The PPI procedure in SPM5 was then used to create regressors representing the timecourse of activation in each TPJ seed region and their interaction with the presence or absence of the contextual cue, (see, Gitelman et al 2003). These regressors were added to the existing subject-specific models, and two new random-effects models were calculated to identify any regions (on a voxel-by-voxel basis) showing increased coupling with left or right TPJ for a target appearing with a salient contextual cue compared to a target alone.
Results
Eyetracking
Eye data were acquired from 9 out of the 13 subjects whose fMRI data were included for analysis. Data quality from 4 out of 13 was inadequate for analysis. Data from each subject were divided into the twelve experimental conditions given by crossing the attended side (left, right), target difficulty (easy, hard), and the three target plus contextual cue conditions of interest (target + contextual cue, target alone and contextual cue alone). The distribution of fixation points in each condition was normal and an average of 93% of the data from each condition were within 1°of the fixation point. There were no significant differences between corresponding left- and right-sided conditions in the proportion of central fixations (all t < 1.7); these data demonstrate that subjects were able to maintain central fixation throughout the experiment (Figure 2).
Figure 2.

Eye-position data from all 12 conditions plotted as the proportion of trials in each condition at each distance in degrees of visual angle from central fixation.
Behavioral performance on the target discrimination task
Target discrimination was moderately difficult and subjects correctly detected an average of 82% of targets. We first report reaction time (RT) data for correct target discriminations and then the signal detection measures of sensitivity (d′) and decision criterion (c). In order to calculate signal detection measures despite relatively high accuracy, subjects with 100% accurate performance in any condition were assigned a hit rate of 99.9% and false alarm rate of .1% (Macmillan & Creelman 1991) (Two subjects had 100% accuracy in 1 condition and 2 had 100% accuracy in 2 conditions. All conditions with 100% accuracy included the contextual cue.) Sensitivity, d′ = (z(hits) - z(false alarms)), and decision criterion, c = (z(hits) + z(false alarms))/2), were calculated for each subject in each condition (Green & Swets 1966; Levine & Parkinson 1994).
Analysis of RT data only included trials where a target was correctly detected. A repeated measures ANOVA of the six conditions given by the attended side (left, right), difficulty (easy, hard) and contextual cue (present, absent). First, RTs were shorter when the contextual cue was present, (F(1,12) = 56.7, p < .00005), (means: contextual cue present = 593 ms, absent= 639 ms; Figure 3A), suggesting that the contextual cue facilitated performance despite being in the unattended visual stream opposite to the target. Second, as expected, RTs were shorter when the target was easy compared to hard, (F(1,12) = 23.2, p < .0005), (means: easy = 600 ms; hard = 633 ms). There were no differences in performance between left and right sided targets (RT: (F(1,12) = 2.0, p > .1), nor any significant interactions. An additional analysis was also conducted on only target + contextual cue trials as a function of whether this trial preceded or followed the contextual cue alone trial within each block; there was no significant difference, (t(11) = 0.1).
Figure 3.

Behavioral data from trials with correct target detection showing interactions between contextual cue presence and target discrimination difficulty. ‘*’ signifies statistically significant differences. A) RTs were shorter when the contextual cue was present and the difference was bigger when detection was hard; B) sensitivity (d′) was greater for easy targets overall, but amongst hard targets, sensitivity was greater when the contextual cue was present; and C) the decision criterion (c) was more liberal in the presence of a contextual cue, particularly when target discrimination was hard. Error bars are standard error of the mean.
Sensitivity (d′) and decision criterion (c) data were entered into the same repeated measures ANOVA as the RT data and again the contextual cue and difficulty were found to impact both measures of performance. There was a significant main effect of difficulty, (F(1,12) = 19.7, p < .001), and an interaction between difficulty and contextual cue on d′, (F(1,12) = 10.3, p < .01). d′ was significantly higher when target discrimination was easy overall, but when detection was hard, d′ was higher when the contextual cue was present (Figure 3B). Importantly, despite differences between conditions, d′ was high in all conditions (easy lo = 4.0, easy high = 3.8. hard lo = 2.8, hard high = 3.2), demonstrating that the salient contextual cue was not used as a proxy for true target identification (if it were, there would be an equal number of hits and false alarms when the contextual cue appeared resulting in a d′ of zero). No other effects were significant.
The contextual cue had a bigger effect on subjects’ decision criterion (c): c was lower overall when the contextual cue was present, (F(1,12) = 27.6, p < .0005), but more so when target discrimination was hard, (F(1,12) = 5.1, p < .05) (Figure 3C). The more liberal decision criterion corresponded to an increase in false alarm rates, (easy target, contextual cue absent = 2.3%; easy, cue present = 5.1%; hard, cue absent = 3.8%; hard, cue present = 8.9%), but all decision criterion values were positive (i.e., values were to the right of the intersection between the noise and signal-plus-noise distributions). This indicates that subjects were relatively conservative in their decisions that a target had appeared (Green & Swets 1966; Levine & Parkinson 1994). Subjects were likely conservative due to the relative infrequency and difficulty of target discriminations and instructions to only respond if they felt “more than 50% sure” that they saw a target. There were no other significant effects.
fMRI data
The main goal of the study was to understand the attentional mechanisms underlying the use of a contextual cue that operated as a predictor of target presence. The behavioral results demonstrated that the presence of the contextual cue facilitated target processing primarily by changing the decision criterion that a target was present. We hypothesized that R TPJ and the ventral attentional network would be involved in processing the contextual cue. To functionally localize TPJ and to identify all regions that responded more when a contextual cue appeared with a target, we first conducted a whole-brain group analysis of the contrast of target + contextual cue minus target alone (i.e., with a standard stimulus). The contrast threshold was set at voxel-wise significance of p < .001 and cluster-level significance of p < .001, corrected for family-wise error, (Poline et al 1997). There were three significant regions: two clusters were in the left and right TPJ (−40 −50 24, peak-level z = 5.8; 46 −58 16, peak-level z = 4.4) and one was in the precuneus, spanning both hemispheres (−6 −56 46, peak-level z = 4.5) (see Figure 4A). These were the only areas within the whole brain analysis that showed greater activation to targets with a contextual cue compared to a target alonei.
Figure 4.

A) TPJ and precuneus regions activated by contrast of target + contextual cue minus target alone. B) Parameter estimates for left and right TPJ ROIs in each of the three conditions of interest.
Based on the group results, we next extracted ROIs for both left and right TPJ (although L TPJ was not part of our original hypothesis) to better understand their functional properties in response to the predictive contextual cue (see Methods). The main goal of the ROI analyses was to distinguish between interpretations of the previous group results by examining TPJ activation on “correct rejection” trials in which the contextual cue appeared alone and a response was correctly withheld. If the greater response in TPJ from the whole-brain analysis for targets + contextual cue compared to targets alone was due solely to stimulus-driven reorienting of attention, or non-specific effects of arousal, then we would expect equivalent activation in response to the contextual cue alone and to a target + contextual cue. Alternatively, if TPJ acts as a “coincidence detector” that responds only to target processing facilitated by contextual information, then activation to the contextual cue alone should be similar to that of a target alone. Finally, if the response in TPJ was due to a combination of stimulus-driven reorienting and the initiation of task-related decisions associated with the stimulus, then we would expect an intermediate response to the contextual cue alone. These contrasts involving the contextual cue alone against the two target present conditions were independent of the contrast with which the ROIs were initially identified.
The pattern of activation was different in left and right TPJ. R TPJ activation in response to the contextual cue alone was intermediate to the two target conditions. This suggests that R TPJ was sensitive to the stimulus-driven aspects of the contextual cue, but more so when it triggered a decision that a target was detected: activation was greater in response to a target + contextual cue than a contextual cue alone (t(11) = 4.2, p < .005); and a contextual cue alone produced greater activation than a target alone, (t(11) = 4.2, p < .005), (Figure 4B). Furthermore, consistent with previous findings showing that R TPJ is deactivated under conditions of top-down attentional focus (Shulman et al 2007), R TPJ activation was negative in response to a target alone, even though the target was relatively infrequent within in the attended stimulus stream, (t(11) = 3.8, p < .005).
In contrast to R TPJ, the response in L TPJ did not differ between targets and contextual cues alone, (t(11) = .44) (Figure 4B). Instead, L TPJ was only activated by the coincidence of a target and contextual cue: targets + contextual cues produced significantly greater activation than contextual cues alone, (t(11) = 2.6, p < .05), or targets alone, (t(11) = 10.2, p < .001). The precuneus cluster from the whole brain analysis produced a similar result (see Supplemental Figure 1 and supporting text). This suggests that L TPJ and the precuneus were not sensitive to the stimulus-driven aspects of the contextual cue, but rather to target discrimination prompted by the informative cue.
In addition to the main effects of the target and contextual cue, we tested for interactions with the manipulations of the target side (left, right) and difficulty (easy, hard) in the two TPJ ROIs. In both left and right TPJ, there were significant interactions with the target side, (L TPJ: F(2,22) = 8.4, p < .005; R TPJ: F(2,22) = 12.7) p < .0005). The interactions were due to the pattern of differences (described above; see Figure 4) being larger when then target was in the ipsilateral visual field (i.e., the contextual cue was contralateral). These results were surprising and suggest that while left and right TPJ produced similar patterns of activation in response to left and right target conditions, they both demonstrated some spatial sensitivity. There were no significant interactions with target difficulty, (both F(2,22) < 2.1, p > .14), although the trend was towards larger effect sizes when the target was hard compared to easy.
Connectivity analysis
To further understand the role of left and right TPJ in stimulus-driven decision processes, functional connectivity analyses were conducted. We conducted “psychophysiological interactions” (PPI; see Methods) (Friston et al 1997), to test for regions whose activity covaried more with activity in left or right TPJ on target + contextual cue trials compared to target alone trials. An inclusive significance threshold of p < .001, uncorrected, was used in conjunction with a cluster size criterion of 10 or more voxels.
There was greater connectivity between R TPJ and a number of attentional and decision areas when a salient contextual cue accompanied the target, including the right inferior frontal gyrus, right IPS, medial frontal cortex, the right pulvinar, bilateral cuneus, and the right parahippocampal gyrus (see Table 1; Figure 5). Lowering the statistical threshold to, p < .005, uncorrected with a cluster size of 10 or more voxels revealed additional activations in bilateral FEF and the left central sulcus (motor cortex) and are reported here for completeness (marked with an “*” in Table 1).
Table 1.
Significant regions from right TPJ PPI connectivity analysis
| Cluster size | T- value | Z-value | MNI XYZ coord | Anatomical region |
|---|---|---|---|---|
| R TPJ seed | ||||
| 27 | 5.74 | 3.83 | 66 6 20 | R inferior frontal gyrus |
| 143 | 5.71 | 3.81 | −42 −34 46 | Left postcentral gyrus |
| 295 | 5.54 | 3.75 | 14 −20 10 | R pulvinar |
| 361 | 5.01 | 3.54 | 14 −100 10 | R cuneus extending to MOG and fusiform gyrusb |
| 128 | 5.33 | 3.67 | 2 22 54 | R medial superior frontal gyrus |
| 85 | 5.50 | 3.74 | −16 −100 12 | L cuneusb |
| 94 | 5.31 | 3.66 | −30 −94 12 | L MOG |
| 300 | 4.94 | 3.51 | 36 −48 44 | R IPSb |
| 26 | 4.82 | 3.46 | −46 −52 2 | L middle temporal lobe |
| 24 | 4.56 | 3.35 | 48 42 −2 | R middle frontal gyrus |
| 256 | 4.36 | 3.25 | −48 −18 56 | L central sulcus (motor cortex)a |
| 23 | 4.05 | 3.11 | 40 −34 26 | R insula a |
| 23 | 4.01 | 3.08 | 34 6 56 | R suprior frontal sulcus (FEF) a |
| 21 | 3.71 | 2.93 | −54 6 −14 | R suprior temporal gyrus a |
| 115 | 6.16 | 3.97 | −44 8 50 | L superior frontal gyrus |
| 42 | 4.13 | 3.14 | −28 2 58 | L sup frontal sulcus (FEF) a |
| 64 | 5.89 | 3.88 | 28 −52 −2 | R parahippocampal gyrus |
| L TPJ seed | ||||
| 10 | 4.72 | 3.42 | 58 −56 38 | R TPJ |
regions significant at p < .005, cluster size 10+
regions also significantly activated by contrast between left and right target hit trials (see Table 2)
Figure 5.
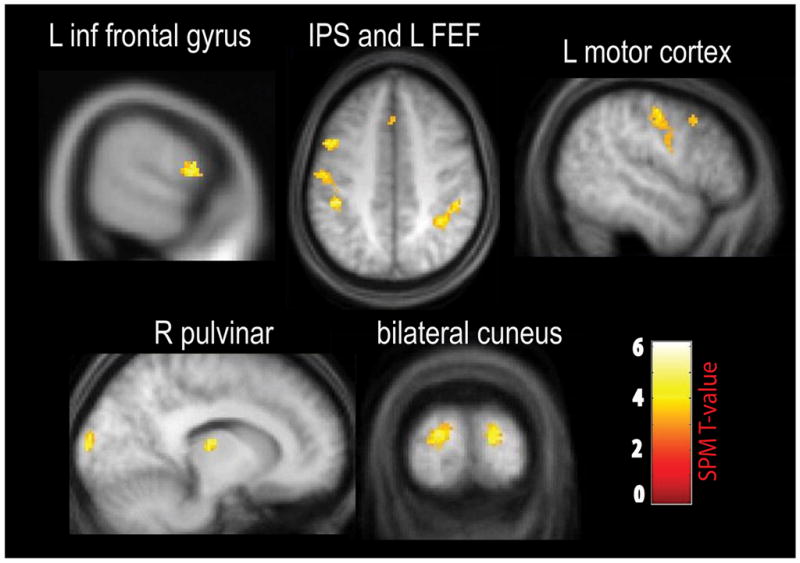
PPI results from whole brain using R TPJ as the seed region. There was greater connectivity between R TPJ and these attentional control and decision areas when the target appeared with a contextual cue compared to a standard in the unattended visual stream.
To test whether the right IPS and bilateral cuneus regions from the PPI were also related to top-down attentional selectivity, the PPI results were masked with the independent contrasts of contralateral attention minus ipsilateral attention (i.e. left targets minus right targets and the reverse; Table 2). All three activations were still present, verifying that the same IPS and extrastriate areas that showed greater connectivity with R TPJ when the contextual cue was present were also involved in attentional selection and task-relevant sensory processing. Thus, detection of a contextual cue that predicted the presence of a target resulted in greater R TPJ connectivity with dorsal attentional control areas, task-relevant visual cortex, and regions involved with task decisions. While it is not possible to infer causal directionality of the connectivity results, they suggest that R TPJ participates in a network that translates a sensory cue into a task-relevant behavioral action.
Table 2.
Target-contralateral activations
| Cluster size | T-value | Z-value | MNI XYZ coord | Anatomical region |
|---|---|---|---|---|
| Attend right minus Left | ||||
| 2644 | 8.83 | 4.83 | −24 −74 28 | L cuneus, extending into L fusiform gyrus |
| 392 | 5.76 | 3.92 | −26 −60 56 | L IPS |
| Attend reft minus Right | ||||
| 2371 | 7.39 | 4.45 | 42 −70 −2 | R middle occipital gyrus fusiform |
| 89 | 6.78 | 4.27 | −24 −94 −14 | L inferior occipital gyurs |
| 89 | 6.45 | 4.16 | 30 −54 50 | R IPS |
| 18 | 5.05 | 3.63 | −8 −52 48 | L precuneus |
In contrast to the results from R TPJ, the only significant cluster from the L TPJ PPI was in the vicinity of the R TPJ ROI (see Table 1). Even using a lower threshold of p < .01, uncorrected with the R TPJ results as an inclusive mask did not reveal any significant clusters of activation. Differences from the connectivity analyses between left and right TPJ are consistent with idea that stimulus-driven attentional control is right lateralized (e.g. Corbetta et al 2008; Friedrich et al 1998; Kincade et al 2005; Kinsbourne 1977; Mort et al 2003; Shulman et al 2009; Vossel et al 2009). The current results go further and indicate that R TPJ is sensitive to stimulus-driven contextual information that is used for generating task-relevant responses.
Discussion
The temporoparietal region is thought to be a critical hub within the ventral attentional control network that supports the detection of behaviorally relevant stimuli. This is in contrast to the dorsal attentional network (including IPS and FEF), which contain attentional priority maps that reflect the current distribution of attention amongst all stimuli, regardless of task-relevance (e.g., Balan & Gottlieb 2006; Bisley & Goldberg 2003; Donner et al 2002; Geng & Mangun 2009; Gottlieb 2007; Hu et al 2009). TPJ (particularly in the right hemisphere) is selectively activated in fMRI studies by the unexpected occurrence of information that is potentially task-relevant. “Task-relevance” has most frequently been defined by a target object or features of the target template (e.g., the target’s color or location) (e.g. Corbetta et al 2000; Corbetta et al 2008; Downar et al 2000; Hampshire et al 2009; Hu et al 2009; Indovina & Macaluso 2007; Natale et al 2009; Serences et al 2005; Shulman et al 2007; Vossel et al 2009). These studies have demonstrated that TPJ is actively involved in reorienting attention towards a target, or target-like stimulus, but have left open the question of whether TPJ is sensitive to non-target information that nevertheless could predict task-relevant behaviors.
In this study we defined task-relevance with a contextual cue (in an unattended visual field) that was probabilistically related to the appearance of the target. Although in the unattended stream, the contextual cue was perceptually salient and produced stimulus-driven attentional capture. This paradigm differs intentionally from those in which salient non-targets were distractors that interfered with processing (e.g., Balan & Gottlieb 2006; Bichot & Schall 2002; Bisley & Goldberg 2006; Donner et al 2002; Geng & Mangun 2009; Gottlieb 2007; Hu et al 2009; Serences et al 2005). Using a probabilistic contextual cue allowed us to test TPJ sensitivity to a non-target stimulus that carried task-relevant information both when it was only of potential relevance (contextual cue only condition) compared to when it was of practical relevance (target + contextual cue).
Using this paradigm, we found that R TPJ increased activation in response to the stimulus-driven contextual cue that likely captured attention, but more so when that contextual cue was accompanied by a target. Activation was the lowest (and negative) when a target appeared alone, suggesting that R TPJ activity was specifically related to processes triggered by the salient contextual stimulus. We hypothesize that detection of the contextual cue by R TPJ produced a stimulus-driven increase in the expectation that an infrequent target would be present. Consistent with this, there was greater connectivity between R TPJ and several attentional control and decision areas (including the right IFG) when the contextual cue appeared with a target than a target alone. The increased expectation facilitated responses by decreasing target discrimination RTs and making the decision criterion more liberal, which occasionally led to a false alarm. This suggests that R TPJ is part of a network that translates stimulus-driven contextual information into task-relevant processing and responses. This is consistent with previous results showing that R TPJ and IFG form a ventral attentional control network (Fox et al 2006; Shulman et al 2009; Umarova et al 2010) and further suggests that TPJ is more sensitive to the sensory event and R IFG to the decision that a target was detected.
Interestingly, activation in R TPJ did not appear to be specifically related to the frequency of the target or the contextual cue since R TPJ did not respond to a target alone, which appeared with the same frequency as a target with the contextual cue. Instead it seems that R TPJ responded to the stimulus-driven shift of attention towards the salient contextual cue, which then increased activity within attention and decision regions in expectation of a target. This is in contrast to the results from L TPJ, which responded only to the coincidence of a contextual cue with a target, but not to either alone. L TPJ also did not show any changes in connectivity specific to the presence of the contextual cue. This suggests that L TPJ is involved in the process of discriminating a target that coincides with a contextual cue, but may not be involved in detecting the contextual cue nor the network relating the cue with task decisions and responses. Importantly, activation in both ROIs in response to the contextual cue alone differed from that of the contextual cue with a target, suggesting that TPJ was not responding solely to task-independent changes in arousal or alertness due to the onset of a salient stimulus.
The characterization of R TPJ as being involved in stimulus-driven changes in behavior also fits generally with the role of R TPJ in theory-of-mind tasks where complex expectations and inferences are integrated for generating an appropriate behavioral response(Hein & Knight 2008; Saxe & Kanwisher 2003; Saxe et al 2004). Thus, a more concrete understanding of the role of TPJ in stimulus-to-behavior transformations should help clarify the specific roles of left and right TPJ in the different tasks where these brain regions have been found to be active.
In conclusion, the current study demonstrated that a salient contextual cue that was probabilistically related to a target stimulus resulted in shorter RTs and more liberal decision criteria. R TPJ was activated by the contextual cue alone and more so when it appeared with a target. R TPJ also showed greater connectivity with other attentional control and decision areas when the contextual cue accompanied the target, suggesting that R TPJ was important for executing stimulus-driven changes in behavior. Our data suggest that R TPJ and L TPJ are both sensitive to the association of a contextual stimulus and task responses, but that only R TPJ is part of a network that detects and translates sensory information into task-relevant decisions.
Research Highlights.
Salient contextual cue increases TPJ connectivity with attention and decision areas
TPJ is sensitive to probabilistic contextual cues that facilitate behavior
Supplementary Material
Acknowledgments
This work was supported by grants from NIMH (MH055714) and NEI (EY015387). We would like to thank Pia Rotshtein and two anonymous reviewers for many helpful comments on the manuscript.
Footnotes
The higher luminance contextual cue was expected to activate visual cortex in this contrast. The following areas were significant at the less stringent voxel level significance threshold of p < .001, uncorrected: the right cuneus (14 −98 14; 18 −98 12), left cuneus (−14 −86 0) and left calcarine sulcus (−6 −94 2). Activation in these regions is consistent with the location of stimuli in the lower visual quadrants.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Balan PF, Gottlieb J. Integration of exogenous input into a dynamic salience map revealed by perturbing attention. J Neurosci. 2006;26:9239–49. doi: 10.1523/JNEUROSCI.1898-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bichot NP, Schall JD. Priming in macaque frontal cortex during popout visual search: feature-based facilitation and location-based inhibition of return. J Neurosci. 2002;22:4675–85. doi: 10.1523/JNEUROSCI.22-11-04675.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bisley JW, Goldberg ME. Neuronal activity in the lateral intraparietal area and spatial attention. Science. 2003;299:81–6. doi: 10.1126/science.1077395. [DOI] [PubMed] [Google Scholar]
- Bisley JW, Goldberg ME. Neural correlates of attention and distractibility in the lateral intraparietal area. Journal of neurophysiology. 2006;95:1696–717. doi: 10.1152/jn.00848.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corbetta M. Frontoparietal cortical networks for directing attention and the eye to visual locations: identical, independent, or overlapping neural systems? Proceedings of the National Academy of Sciences of the United States of America. 1998;95:831–8. doi: 10.1073/pnas.95.3.831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corbetta M, Kincade JM, Ollinger JM, McAvoy MP, Shulman GL. Voluntary orienting is dissociated from target detection in human posterior parietal cortex. Nature neuroscience. 2000;3:292–7. doi: 10.1038/73009. [DOI] [PubMed] [Google Scholar]
- Corbetta M, Patel G, Shulman GL. The reorienting system of the human brain: from environment to theory of mind. Neuron. 2008;58:306–24. doi: 10.1016/j.neuron.2008.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donner TH, Kettermann A, Diesch E, Ostendorf F, Villringer A, Brandt SA. Visual feature and conjunction searches of equal difficulty engage only partially overlapping frontoparietal networks. Neuroimage. 2002;15:16–25. doi: 10.1006/nimg.2001.0951. [DOI] [PubMed] [Google Scholar]
- Downar J, Crawley AP, Mikulis DJ, Davis KD. A multimodal cortical network for the detection of changes in the sensory environment. Nature neuroscience. 2000;3:277–83. doi: 10.1038/72991. [DOI] [PubMed] [Google Scholar]
- Downar J, Crawley AP, Mikulis DJ, Davis KD. A cortical network sensitive to stimulus salience in a neutral behavioral context across multiple sensory modalities. Journal of neurophysiology. 2002;87:615–20. doi: 10.1152/jn.00636.2001. [DOI] [PubMed] [Google Scholar]
- Fox MD, Corbetta M, Snyder AZ, Vincent JL, Raichle ME. Spontaneous neuronal activity distinguishes human dorsal and ventral attention systems. Proceedings of the National Academy of Sciences of the United States of America. 2006;103:10046–51. doi: 10.1073/pnas.0604187103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedrich FJ, Egly R, Rafal RD, Beck D. Spatial attention deficits in humans: a comparison of superior parietal and temporal-parietal junction lesions. Neuropsychology. 1998;12:193–207. doi: 10.1037//0894-4105.12.2.193. [DOI] [PubMed] [Google Scholar]
- Friston K. Beyond phrenology: what can neuroimaging tell us about distributed circuitry? Annual review of neuroscience. 2002;25:221–50. doi: 10.1146/annurev.neuro.25.112701.142846. [DOI] [PubMed] [Google Scholar]
- Friston KJ, Buechel C, Fink GR, Morris J, Rolls E, Dolan RJ. Psychophysiological and modulatory interactions in neuroimaging. Neuroimage. 1997;6:218–29. doi: 10.1006/nimg.1997.0291. [DOI] [PubMed] [Google Scholar]
- Geng JJ, Eger E, Ruff CC, Kristjansson A, Rotshtein P, Driver J. On-line attentional selection from competing stimuli in opposite visual fields: Effects on human visual cortex and control processes. Journal of neurophysiology. 2006 doi: 10.1152/jn.01245.2005. [DOI] [PubMed] [Google Scholar]
- Geng JJ, Mangun GR. Anterior Intraparietal Sulcus is Sensitive to Bottom-Up Attention Driven by Stimulus Salience. Journal of cognitive neuroscience. 2009;21:1584–601. doi: 10.1162/jocn.2009.21103. [DOI] [PubMed] [Google Scholar]
- Gitelman DR, Penny WD, Ashburner J, Friston KJ. Modeling regional and psychophysiologic interactions in fMRI: the importance of hemodynamic deconvolution. NeuroImage. 2003;19:200–7. doi: 10.1016/s1053-8119(03)00058-2. [DOI] [PubMed] [Google Scholar]
- Goldberg ME, Bisley JW, Powell KD, Gottlieb J. Saccades, salience and attention: the role of the lateral intraparietal area in visual behavior. Progress in brain research. 2006;155:157–75. doi: 10.1016/S0079-6123(06)55010-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gottlieb J. From thought to action: the parietal cortex as a bridge between perception, action, and cognition. Neuron. 2007;53:9–16. doi: 10.1016/j.neuron.2006.12.009. [DOI] [PubMed] [Google Scholar]
- Green DM, Swets JA. Signal Detection Theory and Psychophysics. New York: Wiley; 1966. [Google Scholar]
- Hampshire A, Duncan J, Owen AM. Selective tuning of the blood oxygenation level-dependent response during simple target detection dissociates human frontoparietal subregions. J Neurosci. 2007;27:6219–23. doi: 10.1523/JNEUROSCI.0851-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hampshire A, Thompson R, Duncan J, Owen AM. Selective tuning of the right inferior frontal gyrus during target detection. Cognitive, affective & behavioral neuroscience. 2009;9:103–12. doi: 10.3758/CABN.9.1.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hein G, Knight RT. Superior temporal sulcus—It’s my area: or is it? Journal of cognitive neuroscience. 2008;20:2125–36. doi: 10.1162/jocn.2008.20148. [DOI] [PubMed] [Google Scholar]
- Hu S, Bu Y, Song Y, Zhen Z, Liu J. Dissociation of attention and intention in human posterior parietal cortex: an fMRI study. The European journal of neuroscience. 2009;29:2083–91. doi: 10.1111/j.1460-9568.2009.06757.x. [DOI] [PubMed] [Google Scholar]
- Indovina I, Macaluso E. Dissociation of stimulus relevance and saliency factors during shifts of visuospatial attention. Cereb Cortex. 2007;17:1701–11. doi: 10.1093/cercor/bhl081. [DOI] [PubMed] [Google Scholar]
- Jack AI, Patel GH, Astafiev SV, Snyder AZ, Akbudak E, et al. Changing human visual field organization from early visual to extra-occipital cortex. PLoS ONE. 2007;2:e452. doi: 10.1371/journal.pone.0000452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kincade JM, Abrams RA, Astafiev SV, Shulman GL, Corbetta M. An event-related functional magnetic resonance imaging study of voluntary and stimulus-driven orienting of attention. J Neurosci. 2005;25:4593–604. doi: 10.1523/JNEUROSCI.0236-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kinsbourne M. Hemi-neglect and hemisphere rivalry. Adv Neurol. 1977;18:41–9. [PubMed] [Google Scholar]
- Levine G, Parkinson S. Experimental Methods in Psychology. Hillsdale, New Jersey: Lawrence Erlbaum Associates; 1994. [Google Scholar]
- Macmillan NA, Creelman CD. Detection Theory: A User’s Guide. New York: Cambridge University Press; 1991. [Google Scholar]
- Marois R, Leung HC, Gore JC. A stimulus-driven approach to object identity and location processing in the human brain. Neuron. 2000;25:717–28. doi: 10.1016/s0896-6273(00)81073-9. [DOI] [PubMed] [Google Scholar]
- Mort DJ, Malhotra P, Mannan SK, Rorden C, Pambakian A, et al. The anatomy of visual neglect. Brain. 2003;126:1986–97. doi: 10.1093/brain/awg200. [DOI] [PubMed] [Google Scholar]
- Natale E, Marzi CA, Macaluso E. Right temporal-parietal junction engagement during spatial reorienting does not depend on strategic attention control. Neuropsychologia. 2009 doi: 10.1016/j.neuropsychologia.2009.11.012. [DOI] [PubMed] [Google Scholar]
- Poline JB, Worsley KJ, Evans AC, Friston KJ. Combining spatial extent and peak intensity to test for activations in functional imaging. NeuroImage. 1997;5:83–96. doi: 10.1006/nimg.1996.0248. [DOI] [PubMed] [Google Scholar]
- Posner MI. Orienting of attention. Q J Exp Psychol. 1980;32:3–25. doi: 10.1080/00335558008248231. [DOI] [PubMed] [Google Scholar]
- Saxe R, Kanwisher N. People thinking about thinking people. The role of the temporo-parietal junction in “theory of mind”. NeuroImage. 2003;19:1835–42. doi: 10.1016/s1053-8119(03)00230-1. [DOI] [PubMed] [Google Scholar]
- Saxe R, Xiao DK, Kovacs G, Perrett DI, Kanwisher N. A region of right posterior superior temporal sulcus responds to observed intentional actions. Neuropsychologia. 2004;42:1435–46. doi: 10.1016/j.neuropsychologia.2004.04.015. [DOI] [PubMed] [Google Scholar]
- Serences JT, Shomstein S, Leber AB, Golay X, Egeth HE, Yantis S. Coordination of voluntary and stimulus-driven attentional control in human cortex. Psychol Sci. 2005;16:114–22. doi: 10.1111/j.0956-7976.2005.00791.x. [DOI] [PubMed] [Google Scholar]
- Serences JT, Yantis S. Spatially selective representations of voluntary and stimulus-driven attentional priority in human occipital, parietal, and frontal cortex. Cereb Cortex. 2007;17:284–93. doi: 10.1093/cercor/bhj146. [DOI] [PubMed] [Google Scholar]
- Shulman GL, Astafiev SV, Franke D, Pope DL, Snyder AZ, et al. Interaction of stimulus-driven reorienting and expectation in ventral and dorsal frontoparietal and basal ganglia-cortical networks. J Neurosci. 2009;29:4392–407. doi: 10.1523/JNEUROSCI.5609-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shulman GL, Astafiev SV, McAvoy MP, d′Avossa G, Corbetta M. Right TPJ deactivation during visual search: functional significance and support for a filter hypothesis. Cereb Cortex. 2007;17:2625–33. doi: 10.1093/cercor/bhl170. [DOI] [PubMed] [Google Scholar]
- Umarova RM, Saur D, Schnell S, Kaller CP, Vry MS, et al. Structural connectivity for visuospatial attention: significance of ventral pathways. Cereb Cortex. 2010;20:121–9. doi: 10.1093/cercor/bhp086. [DOI] [PubMed] [Google Scholar]
- Vossel S, Weidner R, Thiel CM, Fink GR. What is “odd” in Posner’s location-cueing paradigm? Neural responses to unexpected location and feature changes compared. Journal of cognitive neuroscience. 2009;21:30–41. doi: 10.1162/jocn.2009.21003. [DOI] [PubMed] [Google Scholar]
- Yantis S, Schwarzbach J, Serences JT, Carlson RL, Steinmetz MA, et al. Transient neural activity in human parietal cortex during spatial attention shifts. Nature neuroscience. 2002;5:995–1002. doi: 10.1038/nn921. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.