

Abstract
To be the fittest is central to proliferation in evolutionary games. Individuals
thus adopt the strategies of better performing players in the hope of successful
reproduction. In structured populations the array of those that are eligible to
act as strategy sources is bounded to the immediate neighbors of each
individual. But which one of these strategy sources should potentially be
copied? Previous research dealt with this question either by selecting the
fittest or by selecting one player uniformly at random. Here we introduce a
parameter  that interpolates between these two extreme options.
Setting
that interpolates between these two extreme options.
Setting  equal to zero returns the random selection of the
opponent, while positive
equal to zero returns the random selection of the
opponent, while positive  favor the fitter
players. In addition, we divide the population into two groups. Players from
group
favor the fitter
players. In addition, we divide the population into two groups. Players from
group 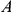 select their opponents as dictated by the parameter
select their opponents as dictated by the parameter
 , while players from group
, while players from group
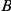 do so randomly irrespective of
do so randomly irrespective of
 . We denote the fraction of players contained in groups
. We denote the fraction of players contained in groups
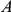 and
and 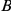 by
by
 and
and  , respectively. The
two parameters
, respectively. The
two parameters  and
and
 allow us to analyze in detail how aspirations in the
context of the prisoner's dilemma game influence the evolution of
cooperation. We find that for sufficiently positive values of
allow us to analyze in detail how aspirations in the
context of the prisoner's dilemma game influence the evolution of
cooperation. We find that for sufficiently positive values of
 there exist a robust intermediate
there exist a robust intermediate
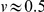 for which cooperation thrives best. The robustness of
this observation is tested against different levels of uncertainty in the
strategy adoption process
for which cooperation thrives best. The robustness of
this observation is tested against different levels of uncertainty in the
strategy adoption process 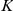 and for different
interaction networks. We also provide complete phase diagrams depicting the
dependence of the impact of
and for different
interaction networks. We also provide complete phase diagrams depicting the
dependence of the impact of  and
and
 for different values of
for different values of 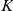 , and contrast the
validity of our conclusions by means of an alternative model where individual
aspiration levels are subject to evolution as well. Our study indicates that
heterogeneity in aspirations may be key for the sustainability of cooperation in
structured populations.
, and contrast the
validity of our conclusions by means of an alternative model where individual
aspiration levels are subject to evolution as well. Our study indicates that
heterogeneity in aspirations may be key for the sustainability of cooperation in
structured populations.
Introduction
Understanding the evolution of cooperation among selfish individuals in human and animal societies remains a grand challenge across disciplines. Evolutionary games are employed frequently as the theoretical framework of choice in order to interpret the emergence and survival of cooperative behavior [1]–[6]. The prisoner's dilemma game, in particular, has attracted considerable interest [7]–[11] as the essential yet minimalist example of a social dilemma. In the original two-person one-shot game the two players have two strategies to choose from (cooperation and defection), and their payoffs depend on the simultaneous decision of both. If they choose to cooperate they will receive the highest collective payoff, which will be shared equally among them. Mutual defection, on the other hand, yields the lowest collective payoff. Yet to defect is tempting because it yields a higher individual payoff regardless of the opponent's decision. It is thus frequently so that both players choose not to cooperate, thus procreating the inevitable social dilemma. In reality, however, interactions may be repeated and the reputation of players compromised [12], [13]. Additionally, individuals may alter with whom they interact [14], and different behaviors may be expressed when participants in a social interaction occupy different roles [15]–[18]. Such and similar considerations have been very successful in elucidating why the unadorned scenario of total defection is often at odds with reality [19], where it is clear that both humans and animals cooperate to achieve what would be impossible by means of isolated efforts. Mechanisms supporting cooperation identified thus far include kin selection [20] as well as many others [6], [21]–[23], and there is progress in place aimed at unifying some of these approaches [24], [25].
Probably the most vibrant of all in recent years have been advances building on the
seminal paper by Nowak and May [26], who showed that spatial structure may sustain
cooperation without the aid of additional mechanisms or strategic complexity.
Although in part anticipated by Hamilton's comments on viscous populations
[20], it was
fascinating to discover that structured populations, including complex and social
networks [27]–[29], provide an optimal
playground for the pursuit of cooperation. Notably, a simple rule for the evolution
of cooperation on graphs and social networks is that natural selection favors
cooperation if 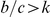 , where
, where 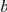 is the benefit of the
altruistic act,
is the benefit of the
altruistic act,  is its cost, while
is its cost, while  is the average number
of neighbors [30]. This is similar to Hamilton's rule stating that
is the average number
of neighbors [30]. This is similar to Hamilton's rule stating that
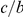 should be larger than the coefficient of genetic relatedness
between individuals [20]. In fact, on graphs and social networks the evolution of
altruism can thus be fully explained by the inclusive fitness theory since the
population is structured such that interactions are between genetic relatives on
average [25],
[31], [32].
should be larger than the coefficient of genetic relatedness
between individuals [20]. In fact, on graphs and social networks the evolution of
altruism can thus be fully explained by the inclusive fitness theory since the
population is structured such that interactions are between genetic relatives on
average [25],
[31], [32].
According to the “best takes all” rule [33], [34] players are allowed to adopt the
strategy of one of their neighbors, provided its payoff is higher than that from the
other neighbors as well as from the player aspiring to improve by changing its
strategy. Based on this relatively simple setup, it was shown that on a square
lattice cooperators form compact clusters and so protect themselves against being
exploited by defectors. The “best takes all” strategy adoption rule is,
however, just one of the many possible alternatives that were considered in the
past. Other examples include the birth-death and imitation rule [35], the
proportional imitation rule [36], the reinforcement learning adoption rule [37], or the
Fermi-function based strategy adoption rule [38]. The latter received substantial
attention, particularly in the physics community, for its compatibility with the
Monte Carlo simulation procedure and the straightforward adjustment of the level of
uncertainty governing the strategy adoptions 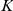 . However, with this
rule the potential donor of the new strategy is selected uniformly at random from
all the neighbors. This is somewhat untrue to what can be observed in reality, where
in fact individuals typically aspire to their most successful neighbors rather than
just somebody random. In this sense the “best takes all” rule seems more
appropriate, although it fails to account for errors in judgment, uncertainty,
external factors, and other disturbances that may vitally affect how we evaluate and
see our co-players. Here we therefore propose a simple tunable function that
interpolates between the “best takes all” and the random selection of a
neighbor in a smooth fashion by means of a single parameter
. However, with this
rule the potential donor of the new strategy is selected uniformly at random from
all the neighbors. This is somewhat untrue to what can be observed in reality, where
in fact individuals typically aspire to their most successful neighbors rather than
just somebody random. In this sense the “best takes all” rule seems more
appropriate, although it fails to account for errors in judgment, uncertainty,
external factors, and other disturbances that may vitally affect how we evaluate and
see our co-players. Here we therefore propose a simple tunable function that
interpolates between the “best takes all” and the random selection of a
neighbor in a smooth fashion by means of a single parameter
 . In this sense the parameter
. In this sense the parameter  acts as an aspiration
parameter, determining to what degree neighbors with a higher payoff will be
considered more likely as potential strategy sources than other (randomly selected)
neighbors.
acts as an aspiration
parameter, determining to what degree neighbors with a higher payoff will be
considered more likely as potential strategy sources than other (randomly selected)
neighbors.
Aiming to further disentangle the role of aspirations, we also consider two types of
players, denoted by type 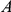 and
and
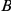 , respectively. While players of type
, respectively. While players of type
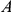 conform to the aspirations imposed by the value of the
aspiration parameter
conform to the aspirations imposed by the value of the
aspiration parameter  , type
, type
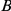 players choose whom to potentially copy uniformly at random
irrespective of
players choose whom to potentially copy uniformly at random
irrespective of  . We denote the fraction of type
. We denote the fraction of type
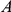 and
and 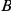 players by
players by
 and
and  , respectively. This
additional division of players into two groups is motivated by the overwhelming
evidence indicating that heterogeneity, almost irrespective of its origin, promotes
cooperative actions. Most notably associated with this statement are complex
networks, including small-world networks [39]–[41], random regular graphs [5], [42], scale-free
networks [43]–[48], as well as adaptive and growing networks [49]–[56]. Furthermore, we
follow the work by McNamara et al. on the coevolution of choosiness
and cooperation [57], in particular by omitting the separation of the
population on two types of players and introducing the heterogeneity by means of
normally distributed individual aspiration levels that are then also subject to
evolution.
, respectively. This
additional division of players into two groups is motivated by the overwhelming
evidence indicating that heterogeneity, almost irrespective of its origin, promotes
cooperative actions. Most notably associated with this statement are complex
networks, including small-world networks [39]–[41], random regular graphs [5], [42], scale-free
networks [43]–[48], as well as adaptive and growing networks [49]–[56]. Furthermore, we
follow the work by McNamara et al. on the coevolution of choosiness
and cooperation [57], in particular by omitting the separation of the
population on two types of players and introducing the heterogeneity by means of
normally distributed individual aspiration levels that are then also subject to
evolution.
At present, we thus investigate how aspirations on an individual level affect the
evolution of cooperation. Having something to aspire to is crucial for progress and
betterment. But how high should we set our goals? Should our role models be only
overachievers and sports heroes, or is it perhaps better to aspire to achieving
somewhat more modest goals? Here we address these questions in the context of the
evolutionary prisoner's dilemma game and determine just how strong and how
widespread aspirations should be for cooperation to thrive best. As we will show, a
strong drive to excellence in the majority of the population may in fact act
detrimental on the evolution of cooperation, while on the other hand, properly
spread and heterogeneous aspirations may be just the key to fully eliminating the
defectors. We will show that this holds irrespective of the structure of the
underlying interaction network, as well as irrespective of the level of uncertainty
by strategy adoptions 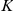 . In addition, the
presented results will be contrasted with the output of a simple coevolutionary
model, where individual aspirations will also be subject to evolution by means of
natural selection. We will conclude that appropriately tuned aspirations may be seen
as a universally applicable promoter of cooperation, which will hopefully inspire
new studies along this line of research.
. In addition, the
presented results will be contrasted with the output of a simple coevolutionary
model, where individual aspirations will also be subject to evolution by means of
natural selection. We will conclude that appropriately tuned aspirations may be seen
as a universally applicable promoter of cooperation, which will hopefully inspire
new studies along this line of research.
Results
Depending on the interaction network, the strategy adoption rule and other simulation
details (see e.g.
[4], [6], [23], [58]), there always
exists a critical cost-to-benefit ratio 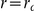 at which cooperators
in the prisoner's dilemma die out. This is directly related to Hamilton's
rule stating that natural selection favors cooperation if
at which cooperators
in the prisoner's dilemma die out. This is directly related to Hamilton's
rule stating that natural selection favors cooperation if
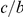 is larger than the coefficient of genetic relatedness
between individuals [20]. If the aspiration parameter
is larger than the coefficient of genetic relatedness
between individuals [20]. If the aspiration parameter
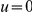 (note that then the division of players to those of type
(note that then the division of players to those of type
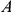 and those of type
and those of type 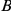 is irrelevant),
is irrelevant),
 , and the interaction network is a square lattice, then, in
our case,
, and the interaction network is a square lattice, then, in
our case, 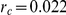 . In what follows, we will typically set
. In what follows, we will typically set
 slightly below this threshold to
slightly below this threshold to
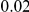 and examine how different values of
and examine how different values of
 ,
,  ,
,
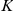 , as well as different interaction networks influence the
outcome of the prisoner's dilemma game.
, as well as different interaction networks influence the
outcome of the prisoner's dilemma game.
It is instructive to first examine characteristic snapshots of the square lattice for
different values of  and
and
 . Results presented in Fig. 1 hint to the conclusion that heterogenous
aspiration to the fittest promotes cooperation, although the details of this claim
depend somewhat on the value of the aspiration parameter
. Results presented in Fig. 1 hint to the conclusion that heterogenous
aspiration to the fittest promotes cooperation, although the details of this claim
depend somewhat on the value of the aspiration parameter
 . For small values of
. For small values of  it is best if all the
players, i.e.
it is best if all the
players, i.e.
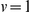 , aspire to their slightly (note that
, aspire to their slightly (note that
 is small) fitter neighbors and thus none actually choose the
potential strategy sources uniformly at random. This can be deduced from the top
three panels of Fig. 1 if
compared from left to right. For large
is small) fitter neighbors and thus none actually choose the
potential strategy sources uniformly at random. This can be deduced from the top
three panels of Fig. 1 if
compared from left to right. For large  , however, it is best
if only half of the players, i.e.
, however, it is best
if only half of the players, i.e.
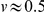 , aspire to their most fittest neighbors, while the other
half chooses their role models randomly. This can be observed if one compares the
bottom three panels of Fig. 1
with one another, although the difference in the overall density of cooperators
(depicted green and blue) between the middle and the right panel is fairly small.
Finally, the role of the aspiration parameter is more clear cut since larger
, aspire to their most fittest neighbors, while the other
half chooses their role models randomly. This can be observed if one compares the
bottom three panels of Fig. 1
with one another, although the difference in the overall density of cooperators
(depicted green and blue) between the middle and the right panel is fairly small.
Finally, the role of the aspiration parameter is more clear cut since larger
 clearly favor the cooperative strategy if compared to small
clearly favor the cooperative strategy if compared to small
 . This can be observed if comparing the snapshots presented
in Fig. 1 vertically.
. This can be observed if comparing the snapshots presented
in Fig. 1 vertically.
Figure 1. Characteristic snapshots of strategy distributions on the square lattice.

Top row depicts results for the aspiration parameter
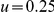 while the bottom row features results for
while the bottom row features results for
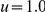 . In both rows the fraction of type
. In both rows the fraction of type
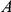 players
players  is
is
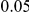 ,
, 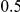 and
and
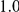 from left to right. Cooperators of type
from left to right. Cooperators of type
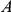 and
and 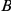 are colored
green and blue, respectively. Defectors of type
are colored
green and blue, respectively. Defectors of type
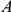 and
and 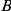 , on the other
hand, are colored red and yellow. If comparing the snapshots vertically, it
can be observed that larger values of
, on the other
hand, are colored red and yellow. If comparing the snapshots vertically, it
can be observed that larger values of  (top
(top
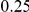 , bottom
, bottom 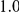 ) clearly
promote the evolution of cooperation. The scenario from left to right via
increasing the fraction of type
) clearly
promote the evolution of cooperation. The scenario from left to right via
increasing the fraction of type 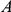 players is not
so clear cut. For
players is not
so clear cut. For 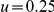 (top row) we
can conclude that larger
(top row) we
can conclude that larger  favor
cooperative behavior, as clearly the cooperators flourish more and more from
the left toward the right panel. For
favor
cooperative behavior, as clearly the cooperators flourish more and more from
the left toward the right panel. For 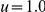 (bottom row),
however, it seems that for
(bottom row),
however, it seems that for 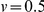 (bottom
middle) cooperators actually fare better then for both
(bottom
middle) cooperators actually fare better then for both
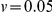 (bottom left) and
(bottom left) and 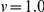 (bottom
right). Hence, the conclusion imposes that for higher
(bottom
right). Hence, the conclusion imposes that for higher
 values an intermediate (rather than the maximal, as
is the case for lower
values an intermediate (rather than the maximal, as
is the case for lower  ) fraction of
type
) fraction of
type 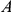 players (those that aspire to their most fittest
neighbors only) is optimal for the evolution of cooperation. Results in all
panels were obtained for
players (those that aspire to their most fittest
neighbors only) is optimal for the evolution of cooperation. Results in all
panels were obtained for 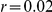 and
and
 .
.
Since the snapshots presented in Fig.
1 can be used primarily for an initial qualitative assessment of the
impact of heterogeneous aspirations, we present in Fig. 2 the fraction of cooperators
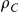 (left) and the critical cost-to-benefit ratio
(left) and the critical cost-to-benefit ratio
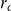 (right) in dependence on
(right) in dependence on  for different values
of
for different values
of  . It can be observed that the promotion of cooperation for
the optimal combination of the two parameters, being
. It can be observed that the promotion of cooperation for
the optimal combination of the two parameters, being 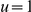 and
and
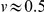 , is really remarkable. The fraction of cooperators rises
from
, is really remarkable. The fraction of cooperators rises
from 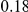 to
to 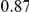 , while the critical
cost-to-benefit ratio rises a full order of magnitude from
, while the critical
cost-to-benefit ratio rises a full order of magnitude from
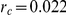 to
to 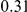 . As tentatively
deduced from the lower three snapshots in Fig. 1, it can also be observed that for high
values of
. As tentatively
deduced from the lower three snapshots in Fig. 1, it can also be observed that for high
values of  an intermediate fraction of type
an intermediate fraction of type
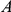 players is optimal for the evolution of cooperation.
Conversely, for low
players is optimal for the evolution of cooperation.
Conversely, for low  the fraction of
cooperators
the fraction of
cooperators 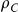 and the critical cost-to-benefit ratio
and the critical cost-to-benefit ratio
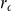 both increase monotonously with increasing
both increase monotonously with increasing
 . If, however, selecting a particular value of
. If, however, selecting a particular value of
 , then the impact of the aspiration parameter
, then the impact of the aspiration parameter
 is always such that cooperation is the more promoted the
larger the value of
is always such that cooperation is the more promoted the
larger the value of  . This can be observed
clearly from both panels, and indeed seems like the main driving force behind the
elevated levels of cooperation. Fine-tuning the fraction of players making use of
the aspiration to the fittest (from
. This can be observed
clearly from both panels, and indeed seems like the main driving force behind the
elevated levels of cooperation. Fine-tuning the fraction of players making use of
the aspiration to the fittest (from 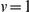 downwards since the
downwards since the
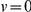 limit trivially returns the random selection of potential
strategy sources) at high
limit trivially returns the random selection of potential
strategy sources) at high  can rise the
cooperation level further, but more in the sense of minor adjustments, similarly as
was observed in the past for the impact of uncertainty by strategy adoptions [42] or the impact of
noise [59].
can rise the
cooperation level further, but more in the sense of minor adjustments, similarly as
was observed in the past for the impact of uncertainty by strategy adoptions [42] or the impact of
noise [59].
Figure 2. Promotion of cooperation due to heterogenous aspirations on the square lattice.

Left panel depicts the density of cooperators 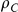 in dependence
on the fraction of type
in dependence
on the fraction of type 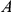 players
players
 for different values of the aspiration parameter
for different values of the aspiration parameter
 . Right panel depicts the critical cost-to-benefit
ratio
. Right panel depicts the critical cost-to-benefit
ratio 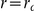 at which cooperators die out, i.e.
at which cooperators die out, i.e.
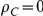 , in dependence on
, in dependence on  for different
values of
for different
values of  . Results in both panels convey the message that low
values of
. Results in both panels convey the message that low
values of  require a high fraction of type
require a high fraction of type
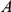 players for cooperation to flourish. Conversely,
higher values of
players for cooperation to flourish. Conversely,
higher values of  sustain
cooperation optimally if only half (
sustain
cooperation optimally if only half (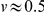 ) of the
players aspires to their most fittest neighbors while the rest chooses whom
to potentially imitate uniformly at random. Optimal conditions for the
evolution of cooperation thus require
) of the
players aspires to their most fittest neighbors while the rest chooses whom
to potentially imitate uniformly at random. Optimal conditions for the
evolution of cooperation thus require  and
and
 to be fine-tuned jointly. Depicted results in both
panels were obtained for
to be fine-tuned jointly. Depicted results in both
panels were obtained for 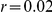 and
and
 .
.
Aiming to generalize the validity of our results, we present in Fig. 3 the fraction of cooperators
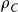 in dependence on
in dependence on  for different values
of
for different values
of  as obtained on the random regular graph (left) and the
small-world network (right). The goal is to test to what extend above conclusions
hold also on interaction networks other than the square lattice, in particular such
that are more complex and spatially heterogenous. If comparing the obtained results
with those presented in the left panel of Fig. 2, it seems save to conclude that they are
to a very large degree qualitatively identical. Some differences nevertheless can be
observed. The first is that what constitutes a high
as obtained on the random regular graph (left) and the
small-world network (right). The goal is to test to what extend above conclusions
hold also on interaction networks other than the square lattice, in particular such
that are more complex and spatially heterogenous. If comparing the obtained results
with those presented in the left panel of Fig. 2, it seems save to conclude that they are
to a very large degree qualitatively identical. Some differences nevertheless can be
observed. The first is that what constitutes a high  limit is a bit higher
on complex networks than on the regular lattice. Note that for
limit is a bit higher
on complex networks than on the regular lattice. Note that for
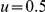 the optimal fraction of type
the optimal fraction of type 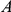 players is practically
still
players is practically
still 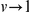 . Even for
. Even for 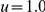 the bell-shaped
dependence on
the bell-shaped
dependence on  is far less pronounced than on the square lattice, and the
optimal
is far less pronounced than on the square lattice, and the
optimal  (the peak of
(the peak of 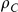 ) is closer to
) is closer to
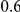 than
than 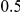 . The second difference
is, looking relatively to the starting point at
. The second difference
is, looking relatively to the starting point at 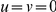 , that the promotion of
cooperation due to positive
, that the promotion of
cooperation due to positive  and
and
 is somewhat less prolific. This is, however, not that
surprising since complex networks in general promote cooperation already on their
own [6], and thus
secondary promotive mechanisms may therefore become less expressed. Aside from these
fairly mild differences though, we can conclude that heterogenous aspirations do
promote cooperation irrespective of the underlying interaction network, and that the
details of the promotive effect are largely universal and predictable.
is somewhat less prolific. This is, however, not that
surprising since complex networks in general promote cooperation already on their
own [6], and thus
secondary promotive mechanisms may therefore become less expressed. Aside from these
fairly mild differences though, we can conclude that heterogenous aspirations do
promote cooperation irrespective of the underlying interaction network, and that the
details of the promotive effect are largely universal and predictable.
Figure 3. Promotion of cooperation due to heterogenous aspirations on the random regular graph (RRG) and the small-world (SW) network.

Left panel depicts the density of cooperators 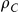 in dependence
on the fraction of type
in dependence
on the fraction of type 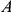 players
players
 for different values of the aspiration parameter
for different values of the aspiration parameter
 for the RRG. Right panel depicts
for the RRG. Right panel depicts
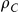 in dependence on
in dependence on  for different
values of
for different
values of  for the Watts-Strogatz SW network with the fraction
of rewired links equalling
for the Watts-Strogatz SW network with the fraction
of rewired links equalling 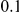 . These results
are in agreement with those presented in Fig. 2, supporting the conclusion that
the impact of heterogenous aspirations on the evolution of cooperation is
robust against alterations of the interaction network. As on the square
lattice, low, but also intermediate, values of
. These results
are in agreement with those presented in Fig. 2, supporting the conclusion that
the impact of heterogenous aspirations on the evolution of cooperation is
robust against alterations of the interaction network. As on the square
lattice, low, but also intermediate, values of
 require
require 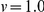 for
cooperation to thrive, while higher values of
for
cooperation to thrive, while higher values of
 sustain cooperation optimally only if
sustain cooperation optimally only if
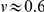 . Depicted results in both panels were obtained for
. Depicted results in both panels were obtained for
 and
and  .
.
Next, we proceed with examining how positive values of
 and
and  fare under different
levels of uncertainty by strategy adoptions. The latter can be tuned via
fare under different
levels of uncertainty by strategy adoptions. The latter can be tuned via
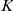 [see Eq. (3)], which acts as a temperature
parameter in the employed Fermi strategy adoption function [38]. Accordingly, when
[see Eq. (3)], which acts as a temperature
parameter in the employed Fermi strategy adoption function [38]. Accordingly, when
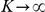 all information is lost and the strategies are adopted by
means of a coin toss. Note that this aspect has thus far not received any attention
here as
all information is lost and the strategies are adopted by
means of a coin toss. Note that this aspect has thus far not received any attention
here as  was fixed. The matter is not trivial to address because
uncertainty and noise can have a rather profound impact on the evolution of
cooperation [40],
[42], [59]–[62], and thus
care needs to be exercised. The safest and most accurate way to approach the problem
is by means of phase diagrams. Since we have two additional parameters
(
was fixed. The matter is not trivial to address because
uncertainty and noise can have a rather profound impact on the evolution of
cooperation [40],
[42], [59]–[62], and thus
care needs to be exercised. The safest and most accurate way to approach the problem
is by means of phase diagrams. Since we have two additional parameters
( and
and  ) against which we want
to test the impact of
) against which we want
to test the impact of 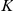 , we determined full
, we determined full
 phase diagrams for six characteristic combinations of
phase diagrams for six characteristic combinations of
 and
and  on the square lattice.
Obtained results are presented in Fig.
4. Notably, the phase diagram presented in the top left panel of Fig. 4 is well-known, implying the
existence of an optimal level of uncertainty for the evolution of cooperation, as
was previously reported in [42], [59]. In particular, note that the
on the square lattice.
Obtained results are presented in Fig.
4. Notably, the phase diagram presented in the top left panel of Fig. 4 is well-known, implying the
existence of an optimal level of uncertainty for the evolution of cooperation, as
was previously reported in [42], [59]. In particular, note that the
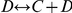 transition line is bell shaped, indicating that
transition line is bell shaped, indicating that
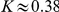 is the optimal temperature at which cooperators are able to
survive at the highest value of
is the optimal temperature at which cooperators are able to
survive at the highest value of  . Importantly though,
this phenomenon can only be observed on interaction topologies lacking overlapping
triangles [63],
[64].
Interestingly, increasing
. Importantly though,
this phenomenon can only be observed on interaction topologies lacking overlapping
triangles [63],
[64].
Interestingly, increasing  from
from
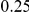 (top row) to
(top row) to 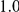 (bottom row)
completely eradicates (as do interaction networks incorporating overlapping
triangles) the existence of an optimal
(bottom row)
completely eradicates (as do interaction networks incorporating overlapping
triangles) the existence of an optimal 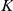 , and in fact
qualitatively reverses the dependence. The
, and in fact
qualitatively reverses the dependence. The 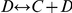 transition line has an
inverted bell-shaped outlay, indicating the existence of the worst rather than an
optimal temperature
transition line has an
inverted bell-shaped outlay, indicating the existence of the worst rather than an
optimal temperature 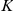 for the evolution of
cooperation. The qualitative changes are less profound if
for the evolution of
cooperation. The qualitative changes are less profound if
 is kept constant at
is kept constant at 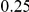 (top row) and
(top row) and
 increases (from left to right). Still, however, the
bell-shaped outlay of the
increases (from left to right). Still, however, the
bell-shaped outlay of the 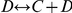 transition gives way
to a monotonically increasing curve, saturating only for high
transition gives way
to a monotonically increasing curve, saturating only for high
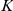 . These qualitative changes in the phase diagrams imply that
increasing the aspiration parameter
. These qualitative changes in the phase diagrams imply that
increasing the aspiration parameter  or the fraction of
players abiding to it (type
or the fraction of
players abiding to it (type 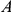 ) effectively alters
the interaction network. While the square lattice obviously lacks overlapping
triangles and thus enables the observation of an optimal
) effectively alters
the interaction network. While the square lattice obviously lacks overlapping
triangles and thus enables the observation of an optimal
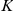 for small enough values of
for small enough values of  and
and
 (or a combination thereof, as is the case in the top left
panels), trimming the likelihood of who will act as a strategy source and how many
players will actually aspire to their fittest neighbors seems to effectively enhance
linkage among essentially disconnected triplets and thus precludes the same
observation. It is instructive to note that a similar phenomenon was observed
recently in public goods games, where the joint membership in large groups was also
found to alter the effective interaction network and thus the impact of uncertainly
on the evolution of cooperation [64].
(or a combination thereof, as is the case in the top left
panels), trimming the likelihood of who will act as a strategy source and how many
players will actually aspire to their fittest neighbors seems to effectively enhance
linkage among essentially disconnected triplets and thus precludes the same
observation. It is instructive to note that a similar phenomenon was observed
recently in public goods games, where the joint membership in large groups was also
found to alter the effective interaction network and thus the impact of uncertainly
on the evolution of cooperation [64].
Figure 4. Full 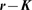 phase diagrams for the square lattice.
phase diagrams for the square lattice.

Top row depicts results for the aspiration parameter
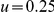 while the bottom row features results for
while the bottom row features results for
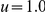 . In both rows the fraction of type
. In both rows the fraction of type
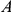 players
players  is
is
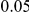 ,
, 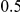 and
and
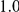 from left to right. The outline of panels thus
corresponds to the snapshots presented in Fig. 1. Thin blue and thick red lines
mark the border between stationary pure
from left to right. The outline of panels thus
corresponds to the snapshots presented in Fig. 1. Thin blue and thick red lines
mark the border between stationary pure 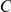 and
and
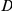 phases and the mixed
phases and the mixed 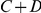 phase,
respectively. In agreement with previous works [42], [63], it can be observed that
for
phase,
respectively. In agreement with previous works [42], [63], it can be observed that
for 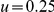 and
and 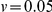 (top left)
there exists an intermediate uncertainty in the strategy adoption process
(an intermediate value of
(top left)
there exists an intermediate uncertainty in the strategy adoption process
(an intermediate value of 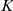 ) for which the
survivability of cooperators is optimal, i.e.
) for which the
survivability of cooperators is optimal, i.e.
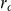 is maximal. Conversely, while the borderline
separating the pure
is maximal. Conversely, while the borderline
separating the pure 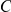 and the mixed
and the mixed
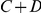 phase for all the other combinations of
phase for all the other combinations of
 and
and  exhibits a
qualitatively identical outlay as for the
exhibits a
qualitatively identical outlay as for the 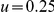 and
and
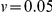 case, the
case, the 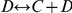 transition is
qualitatively different and very much dependent on the particularities
players' aspirations. Note that in all the bottom panels there exist an
intermediate value of
transition is
qualitatively different and very much dependent on the particularities
players' aspirations. Note that in all the bottom panels there exist an
intermediate value of 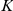 for which
for which
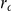 is minimal rather than maximal, while towards the
large
is minimal rather than maximal, while towards the
large 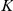 limit
limit 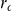 increases,
saturating only for
increases,
saturating only for 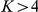 (not shown).
In the top middle and right panel, on the other hand, the bell-shaped outlay
of the
(not shown).
In the top middle and right panel, on the other hand, the bell-shaped outlay
of the 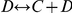 transition gives way to a monotonically increasing
curve, again saturating only for
transition gives way to a monotonically increasing
curve, again saturating only for 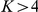 . It can thus
be concluded that, while the aspiration based promotion of cooperation is
largely independent of
. It can thus
be concluded that, while the aspiration based promotion of cooperation is
largely independent of 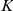 , the details
of phase transition are very much affected, which can be attributed to an
effective alterations of the interaction network due to preferred strategy
sources (see also main text for details).
, the details
of phase transition are very much affected, which can be attributed to an
effective alterations of the interaction network due to preferred strategy
sources (see also main text for details).
In terms of the facilitation of cooperation, however, it can be concluded that the
promotive impact of positive values of  and
and
 prevails irrespective of
prevails irrespective of 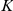 . By comparing the
extend of pure
. By comparing the
extend of pure 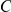 and mixed
and mixed 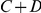 regions for different
pairs of the two parameters, we can observe that for small values of
regions for different
pairs of the two parameters, we can observe that for small values of
 (top panels in Fig. 4) it is best if all the players, i.e.
(top panels in Fig. 4) it is best if all the players, i.e.
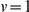 , aspire to their slightly (note that
, aspire to their slightly (note that
 is small) fitter neighbors, while for large
is small) fitter neighbors, while for large
 (bottom panels in Fig. 4) it is best if only approximately half of
the players, i.e.
(bottom panels in Fig. 4) it is best if only approximately half of
the players, i.e.
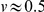 , aspire to their most fittest neighbors. The same
conclusions were stated already upon the inspection of results presented in Figs. 2 and 3, and with this we now affirm that not only is
the promotion of cooperation via heterogeneous aspirations robust against
differences in the interaction networks, but also against variations in the
uncertainty by strategy adoptions.
, aspire to their most fittest neighbors. The same
conclusions were stated already upon the inspection of results presented in Figs. 2 and 3, and with this we now affirm that not only is
the promotion of cooperation via heterogeneous aspirations robust against
differences in the interaction networks, but also against variations in the
uncertainty by strategy adoptions.
It remains of interest to elucidate why then cooperative behavior is in fact promoted
by positive values of  and
and
 . To provide answers, we show in Fig. 5 time courses of
. To provide answers, we show in Fig. 5 time courses of
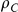 for different characteristic combination of the two main
parameters that we have used throughout this work. What should attract the attention
is the fact that in the most early stages of the evolutionary process (note that
values of
for different characteristic combination of the two main
parameters that we have used throughout this work. What should attract the attention
is the fact that in the most early stages of the evolutionary process (note that
values of 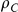 were recorded also in-between full iteration steps) it
appears as if defectors would actually fare better than cooperators. This is
actually what one would expect, given that defectors are, as individuals, more
successful than cooperators and will thus be chosen more likely as potential
strategy donors if
were recorded also in-between full iteration steps) it
appears as if defectors would actually fare better than cooperators. This is
actually what one would expect, given that defectors are, as individuals, more
successful than cooperators and will thus be chosen more likely as potential
strategy donors if  is positive. This
should in turn amplify their chances of spreading and ultimately result in the
decimation of cooperators (indeed, only between 20–30% survive). Quite
surprisingly though, the tide changes fairly fast, and as one can observe from the
presented time courses, frequently the more so the deeper the initial downfall of
cooperators. We argue that for positive values of
is positive. This
should in turn amplify their chances of spreading and ultimately result in the
decimation of cooperators (indeed, only between 20–30% survive). Quite
surprisingly though, the tide changes fairly fast, and as one can observe from the
presented time courses, frequently the more so the deeper the initial downfall of
cooperators. We argue that for positive values of  and
and
 a negative feedback effect occurs, which halts and
eventually reverts what appears to be a march of defectors toward dominance. Namely,
in the very early stages of the game defectors are able to plunder very efficiently,
which quickly results in a state where there are hardly any cooperators left to
exploit. Consequently, the few remaining clusters of cooperators start recovering
lost ground against weakened defectors. Crucial thereby is the fact that the
clusters formed by cooperators are impervious to defector attacks even at high
values of
a negative feedback effect occurs, which halts and
eventually reverts what appears to be a march of defectors toward dominance. Namely,
in the very early stages of the game defectors are able to plunder very efficiently,
which quickly results in a state where there are hardly any cooperators left to
exploit. Consequently, the few remaining clusters of cooperators start recovering
lost ground against weakened defectors. Crucial thereby is the fact that the
clusters formed by cooperators are impervious to defector attacks even at high
values of  because of the positive selection towards the fittest
neighbors acting as strategy sources (occurring for
because of the positive selection towards the fittest
neighbors acting as strategy sources (occurring for 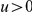 ). In a sea of
cooperators this is practically always another cooperator rather than a defector
trying to penetrate into the cluster. This newly identified mechanism ultimately
results in fairly widespread cooperation that goes beyond what can be warranted by
the spatial reciprocity alone (see e.g.
[6]), and this
irrespective of the underlying interaction network and uncertainty by strategy
adoptions.
). In a sea of
cooperators this is practically always another cooperator rather than a defector
trying to penetrate into the cluster. This newly identified mechanism ultimately
results in fairly widespread cooperation that goes beyond what can be warranted by
the spatial reciprocity alone (see e.g.
[6]), and this
irrespective of the underlying interaction network and uncertainty by strategy
adoptions.
Figure 5. Time courses of the density of cooperators on the square lattice.

Results are presented for the aspiration parameters
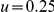 (solid lines) and
(solid lines) and 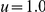 (dashed
lines), each for three different fractions of type
(dashed
lines), each for three different fractions of type
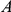 players
players  , as depicted
on the figure. The crucial feature of all time courses is the initial
temporary downfall of cooperators, which sets in for all depicted
combinations of
, as depicted
on the figure. The crucial feature of all time courses is the initial
temporary downfall of cooperators, which sets in for all depicted
combinations of  and
and
 . Quite remarkably, what appears to become an ever
faster extinction eventually becomes a rise to, at least in some cases,
near-dominance. Note that the horizontal axis is logarithmic and that values
of
. Quite remarkably, what appears to become an ever
faster extinction eventually becomes a rise to, at least in some cases,
near-dominance. Note that the horizontal axis is logarithmic and that values
of 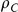 were recorded also in-between full iteration steps
to ensure a proper resolution. Depicted results were obtained for
were recorded also in-between full iteration steps
to ensure a proper resolution. Depicted results were obtained for
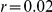 and
and  .
.
Finally, it is instructive to examine whether an optimal intermediate value of
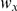 , determining the aspiration level of player
, determining the aspiration level of player
 , can emerge spontaneously from an initial array of normally
distributed values. This would imply that natural selection indeed favors
individuals with a specific aspiration level, which would in turn extend the
credibility of thus far presented results that were obtained primarily in a top-down
manner [by optimizing a population-level property (cooperation) by means of an
appropriate selection of parameters determining the aspiration level of
individuals]. For this purpose we omit the division of the population on
players of type
, can emerge spontaneously from an initial array of normally
distributed values. This would imply that natural selection indeed favors
individuals with a specific aspiration level, which would in turn extend the
credibility of thus far presented results that were obtained primarily in a top-down
manner [by optimizing a population-level property (cooperation) by means of an
appropriate selection of parameters determining the aspiration level of
individuals]. For this purpose we omit the division of the population on
players of type 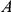 and
and 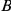 , and initially assign
to every player a value
, and initially assign
to every player a value 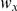 that is drawn randomly
from a Gaussian distribution with a given mean
that is drawn randomly
from a Gaussian distribution with a given mean  and standard deviation
and standard deviation
 . Then if player
. Then if player  adopts the strategy
from player
adopts the strategy
from player  also
also 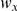 becomes equal to
becomes equal to
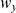 (see Methods for
details). Results obtained with this alternative coevolutionary model are presented
in Fig. 6. It can be observed
that the initial Gaussian distribution sharpens fast around an intermediate value of
(see Methods for
details). Results obtained with this alternative coevolutionary model are presented
in Fig. 6. It can be observed
that the initial Gaussian distribution sharpens fast around an intermediate value of
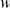 , which then gradually becomes more and more frequent in the
population as the natural selection spontaneously eliminates the less favorable
values that warrant a lower individual fitness. The final state is a population
where virtually all players have an identical aspiration level
, which then gradually becomes more and more frequent in the
population as the natural selection spontaneously eliminates the less favorable
values that warrant a lower individual fitness. The final state is a population
where virtually all players have an identical aspiration level
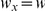 , and accordingly, the outcome in terms of the stationary
density of cooperators is equal to that obtained with the original model having
, and accordingly, the outcome in terms of the stationary
density of cooperators is equal to that obtained with the original model having
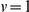 and
and  . In this sense the
preceding results are validated and their generality extended by means of a
bottom-up approach entailing a spontaneous coevolution towards an intermediate
individually optimal aspiration level. We note, however, that with this simple
coevolutionary model the result that heterogeneous aspirations promote cooperation
is not exactly reproduced. Further studies on more sophisticated models
incorporating coevolving aspirations are required to arrive spontaneously at a
heterogeneous distribution of individual aspiration levels. Inspirations for this
can be found in the recent review on coevolutionary games [23], and we are looking forward to
further developments in this direction.
. In this sense the
preceding results are validated and their generality extended by means of a
bottom-up approach entailing a spontaneous coevolution towards an intermediate
individually optimal aspiration level. We note, however, that with this simple
coevolutionary model the result that heterogeneous aspirations promote cooperation
is not exactly reproduced. Further studies on more sophisticated models
incorporating coevolving aspirations are required to arrive spontaneously at a
heterogeneous distribution of individual aspiration levels. Inspirations for this
can be found in the recent review on coevolutionary games [23], and we are looking forward to
further developments in this direction.
Figure 6. Spontaneous fixation towards an intermediate aspiration level by means of natural selection.

Presented results were obtained with the alternative model where players are
not divided into two groups and initially every player is assigned a random
aspiration level 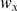 drawn from a
Gaussian distribution with the mean
drawn from a
Gaussian distribution with the mean 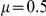 and standard
deviation
and standard
deviation 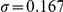 . The main panel depicts the distributions
. The main panel depicts the distributions
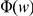 of individual aspiration levels as recorded at
of individual aspiration levels as recorded at
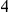 (solid black line),
(solid black line), 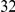 (dashed red
line) and
(dashed red
line) and 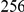 (dotted green line) full iteration steps. The
fixation towards a dominant average value
(dotted green line) full iteration steps. The
fixation towards a dominant average value 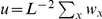 due to natural
selection is evident since the interval of
due to natural
selection is evident since the interval of 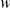 values still
present in the population becomes more and more narrow as time progresses.
The inset shows the convergence of
values still
present in the population becomes more and more narrow as time progresses.
The inset shows the convergence of  (solid gray
line) and
(solid gray
line) and 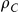 (dotted blue line). The initial temporary downfall
of cooperators, followed by the rise to near-dominance, is well-expressed
also in the coevolutionary setup, and the stationary density agrees well
with the results obtained by means of the original model with
(dotted blue line). The initial temporary downfall
of cooperators, followed by the rise to near-dominance, is well-expressed
also in the coevolutionary setup, and the stationary density agrees well
with the results obtained by means of the original model with
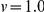 and
and 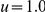 (compare with
the dashed cyan line in Fig.
5). Note that in the inset the horizontal axis is logarithmic and
that values of
(compare with
the dashed cyan line in Fig.
5). Note that in the inset the horizontal axis is logarithmic and
that values of  and
and
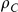 were recorded also in-between full iteration steps
to ensure a proper resolution. Depicted results were obtained for
were recorded also in-between full iteration steps
to ensure a proper resolution. Depicted results were obtained for
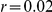 and
and  .
.
Discussion
We have shown that heterogenous aspiration to the fittest, i.e. the
propensity of designating the most successful neighbor as being the role model, may
be seen as a universally applicable promoter of cooperation that works on different
interaction networks and under different levels of stochasticity. For low and
moderate values of the aspiration parameter  cooperation thrives
best if the total population abides to aspiring to the fittest. For large values of
cooperation thrives
best if the total population abides to aspiring to the fittest. For large values of
 , however, it is best if only approximately half of the
players persuasively attempt to copy their most successful neighbors while the rest
chooses their opponents uniformly at random. The optimal evolution of cooperation
thus requires fine-tuning of both, the density of players that are prone to aspiring
to the fittest, as well as the aspiration parameter that determines how fit a
neighbor actually must be in order to be considered as the potential source of the
new strategy. In addition, by studying an alternative model where individual
aspiration levels were also subject to evolution, we have shown that an intermediate
value of the aspiration level emerges spontaneously through natural selection, thus
supplementing the main results by means of a coevolutionary approach.
, however, it is best if only approximately half of the
players persuasively attempt to copy their most successful neighbors while the rest
chooses their opponents uniformly at random. The optimal evolution of cooperation
thus requires fine-tuning of both, the density of players that are prone to aspiring
to the fittest, as well as the aspiration parameter that determines how fit a
neighbor actually must be in order to be considered as the potential source of the
new strategy. In addition, by studying an alternative model where individual
aspiration levels were also subject to evolution, we have shown that an intermediate
value of the aspiration level emerges spontaneously through natural selection, thus
supplementing the main results by means of a coevolutionary approach.
Notably, the extensions of the prisoner's dilemma game we have considered here
seem very reasonable and are in fact easily justifiable with realistic examples. For
example, it is a fact that people will, in general, much more likely follow a
successful individual than somebody who is just struggling to get by. Under certain
adverse circumstances, like in a state of rebelion or in revolutionary times,
however, it is also possible that individuals will be inspired to copy their less
successful partners or those that seem to do more harm than good. In many ways it
seems that the ones who are satisfied with just picking somebody randomly to aspire
to are the ones that are most difficult to come by. In this sense the rather
frequently adopted random selection of a neighbor, retrieved in our case if
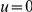 (or equivalently
(or equivalently 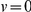 ), seems in many ways
like the least probable alternative. In this sense it is interesting to note that
our aspiring to the fittest becomes identical to the frequently adopted, especially
in the early seminal works on games on grids [26], [33], [34], “best takes all”
adoption rule if
), seems in many ways
like the least probable alternative. In this sense it is interesting to note that
our aspiring to the fittest becomes identical to the frequently adopted, especially
in the early seminal works on games on grids [26], [33], [34], “best takes all”
adoption rule if 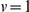 ,
, 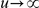 in Eq. (2), and
in Eq. (2), and
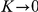 in Eq. (3). Although in our simulations we never quite reach
the “best takes all” limit, and thus a direct comparison with the
seminal works is somewhat circumstantial, we find here that the intermediate regions
of heterogenous aspirations offer fascinating new insights into the evolution of
cooperation, and we hope that this work will inspire future studies, especially in
terms of understanding the emergence of successful leaders in societies via a
coevolutionary process [23].
in Eq. (3). Although in our simulations we never quite reach
the “best takes all” limit, and thus a direct comparison with the
seminal works is somewhat circumstantial, we find here that the intermediate regions
of heterogenous aspirations offer fascinating new insights into the evolution of
cooperation, and we hope that this work will inspire future studies, especially in
terms of understanding the emergence of successful leaders in societies via a
coevolutionary process [23].
Methods
An evolutionary prisoner's dilemma game with the temptation to defect
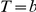 (the highest payoff received by a defector if playing
against a cooperator), reward for mutual cooperation
(the highest payoff received by a defector if playing
against a cooperator), reward for mutual cooperation 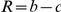 , the punishment for
mutual defection
, the punishment for
mutual defection 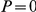 , and the sucker's payoff
, and the sucker's payoff
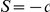 (the lowest payoff received by a cooperator if playing
against a defector) is used as the basis for our simulations. Without loss of
generality the payoffs can be rescaled as
(the lowest payoff received by a cooperator if playing
against a defector) is used as the basis for our simulations. Without loss of
generality the payoffs can be rescaled as 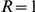 ,
,
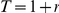 ,
, 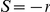 and
and
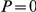 , where
, where 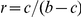 is the cost-to-benefit
ratio [5]. For
positive
is the cost-to-benefit
ratio [5]. For
positive  we have
we have 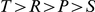 , thus strictly
satisfying the prisoner's dilemma payoff ranking.
, thus strictly
satisfying the prisoner's dilemma payoff ranking.
As the interaction network, we use either a regular 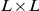 square lattice, the
random regular graph (RRG) constructed as described in [65], or the small-world (SW) topology
with an average degree of four generated via the Watts-Strogatz algorithm [66]. Each vertex
square lattice, the
random regular graph (RRG) constructed as described in [65], or the small-world (SW) topology
with an average degree of four generated via the Watts-Strogatz algorithm [66]. Each vertex
 is initially designated as hosting either players of type
is initially designated as hosting either players of type
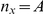 or
or 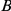 with the probability
with the probability
 and
and  , respectively. This
division of players is performed uniformly at random irrespective of their initial
strategies and remains unchanged during the simulations. According to established
procedures, each player is initially also designated either as a cooperator
(
, respectively. This
division of players is performed uniformly at random irrespective of their initial
strategies and remains unchanged during the simulations. According to established
procedures, each player is initially also designated either as a cooperator
(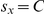 ) or defector (
) or defector (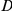 ) with equal
probability. The game is iterated forward in accordance with the sequential
simulation procedure comprising the following elementary steps. First, player
) with equal
probability. The game is iterated forward in accordance with the sequential
simulation procedure comprising the following elementary steps. First, player
 acquires its payoff
acquires its payoff 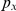 by playing the game
with all its neighbors. Next, we evaluate in the same way the payoffs of all the
neighbors of player
by playing the game
with all its neighbors. Next, we evaluate in the same way the payoffs of all the
neighbors of player  and subsequently
select one neighbor
and subsequently
select one neighbor  via the
probability
via the
probability
| (1) |
where the sum runs over all the neighbors of player  . Importantly,
. Importantly,
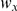 is the so-called selection or aspiration parameter that
depends on the type of player
is the so-called selection or aspiration parameter that
depends on the type of player  according
to
according
to
| (2) |
Evidently, if the aspiration parameter 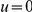 then irrespective of
then irrespective of
 (density of type
(density of type 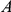 players) the most
frequently adopted situation is recovered where player
players) the most
frequently adopted situation is recovered where player
 is chosen uniformly at random from all the neighbors of
player
is chosen uniformly at random from all the neighbors of
player  . For
. For 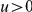 and
and
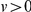 , however, Eqs. (1) and (2) introduce a preference in all
players of type
, however, Eqs. (1) and (2) introduce a preference in all
players of type 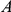 (but not in players of type
(but not in players of type 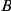 ) to copy the strategy
of those neighbors who have a high fitness, or equivalently, a high payoff
) to copy the strategy
of those neighbors who have a high fitness, or equivalently, a high payoff
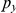 . Lastly then, after the neighbor
. Lastly then, after the neighbor
 that is aspired to by player
that is aspired to by player  is chosen, player
is chosen, player
 adopts the strategy
adopts the strategy 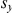 from the selected
player
from the selected
player  with the probability
with the probability
| (3) |
where 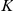 denotes the amplitude of noise or its inverse
(
denotes the amplitude of noise or its inverse
(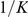 ) the so-called intensity of selection [38]. Irrespective of the values of
) the so-called intensity of selection [38]. Irrespective of the values of
 and
and  one full iteration
step involves all players
one full iteration
step involves all players 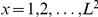 having a chance to
adopt a strategy from one of their neighbors once.
having a chance to
adopt a strategy from one of their neighbors once.
An alternative model, allowing for individual 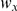 values to be subject
to evolution as well, entails omitting the division of the population on two types
of players and assigning to every individual an initial
values to be subject
to evolution as well, entails omitting the division of the population on two types
of players and assigning to every individual an initial
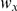 value that is drown randomly from a Gaussian distribution
having mean
value that is drown randomly from a Gaussian distribution
having mean  and standard deviation
and standard deviation  , as was done recently
in [57], for
example. Subsequently, if player
, as was done recently
in [57], for
example. Subsequently, if player  adopts the strategy
from player
adopts the strategy
from player  following the identical procedure as described above for the
original model, then the value of
following the identical procedure as described above for the
original model, then the value of 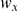 changes to that of
changes to that of
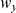 as well. The key question that we aim to answer with this
model is whether a specific aspiration level is indeed optimal for an individual to
prosper, and if yes, does the selection pressure favor it spontaneously.
Essentially, we are interested in the distribution of
as well. The key question that we aim to answer with this
model is whether a specific aspiration level is indeed optimal for an individual to
prosper, and if yes, does the selection pressure favor it spontaneously.
Essentially, we are interested in the distribution of
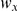 values after the stationary fraction of strategies in the
population is reached. A link with the original model can be established by
considering in this case
values after the stationary fraction of strategies in the
population is reached. A link with the original model can be established by
considering in this case  to equal one and
to equal one and
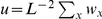 .
.
Results of computer simulations were obtained on populations comprising
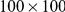 to
to 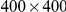 individuals, whereby
the fraction of cooperators
individuals, whereby
the fraction of cooperators 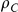 was determined within
was determined within
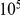 full iteration steps after sufficiently long transients were
discarded. Moreover, since the heterogeneous preferential selection of neighbors may
introduce additional disturbances, final results were averaged over up to
full iteration steps after sufficiently long transients were
discarded. Moreover, since the heterogeneous preferential selection of neighbors may
introduce additional disturbances, final results were averaged over up to
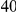 independent runs for each set of parameter values in order
to assure suitable accuracy.
independent runs for each set of parameter values in order
to assure suitable accuracy.
Acknowledgments
We have benefited substantially from the insightful comments of the Editor James Arthur Robert Marshall and an anonymous referee, which were instrumental for elevating the quality of this work. Helpful discussions with Professor Lianzhong Zhang are gratefully acknowledged as well.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: MP acknowledges support from the Slovenian Research Agency (Grant No. Z1-2032). ZW acknowledges support from the Center for Asia Studies of Nankai University (Grant No. 2010-5) and from the National Natural Science Foundation of China (Grant No. 10672081). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Hofbauer J, Sigmund K. Cambridge, UK: Cambridge University Press; 1998. Evolutionary Games and Population Dynamics. [Google Scholar]
- 2.Nowak MA. Cambridge, MA: Harvard University Press; 2006. Evolutionary Dynamics. [Google Scholar]
- 3.Maynard Smith J. Cambridge, UK: Cambridge University Press; 1982. Evolution and the Theory of Games. [Google Scholar]
- 4.Roca CP, Cuesta JA, Sánchez A. Evolutionary game theory: Temporal and spatial effects beyond replicator dynamics. Phys Life Rev. 2009;6:208–249. doi: 10.1016/j.plrev.2009.08.001. [DOI] [PubMed] [Google Scholar]
- 5.Hauert C, Szabó G. Game theory and physics. Am J Phys. 2005;73:405–414. [Google Scholar]
- 6.Szabó G, Fáth G. Evolutionary games on graphs. Phys Rep. 2007;446:97–216. [Google Scholar]
- 7.Nowak M, Sigmund K. Game-dynamical aspects of the prisoner's dilemma. Appl Math Comp. 1989;30:191–213. [Google Scholar]
- 8.Moyano LG, Sánchez A. Evolving learning rules and emergence of cooperation in spatial prisoner's dilemma. J Theor Biol. 2009;259:84–95. doi: 10.1016/j.jtbi.2009.03.002. [DOI] [PubMed] [Google Scholar]
- 9.Doebeli M, Hauert C. Models of cooperation based on prisoner's dilemma and snowdrift game. Ecol Lett. 2005;8:748–766. [Google Scholar]
- 10.Szabó G, Szolnoki A, Sznaider GA. Segregation process and phase transition in cyclic predator-prey models with even number of species. Phys Rev E. 2007;76:051921. doi: 10.1103/PhysRevE.76.051921. [DOI] [PubMed] [Google Scholar]
- 11.Szolnoki A, Perc M, Szabó G, Stark HU. Impact of aging on the evolution of cooperation in the spatial prisoner's dilemma game. Phys Rev E. 2009;80:021901. doi: 10.1103/PhysRevE.80.021901. [DOI] [PubMed] [Google Scholar]
- 12.Milinski M, Semmann D, Krambeck HJ. Reputation helps to solve the ‘tragedy of the commons’. Nature. 2002;415:424–426. doi: 10.1038/415424a. [DOI] [PubMed] [Google Scholar]
- 13.Fehr E. Don't lose your reputation. Nature. 2004;432:449–450. doi: 10.1038/432449a. [DOI] [PubMed] [Google Scholar]
- 14.Van Segbroeck S, Santos FC, Lenaerts T, Pacheco JM. Reacting differently to adverse ties promotes cooperation in social networks. Phys Rev Lett. 2009;102:058105. doi: 10.1103/PhysRevLett.102.058105. [DOI] [PubMed] [Google Scholar]
- 15.Maynard Smith J, Parker GA. The logic of asymmetric contests. Anim Behav. 1976;24:159–175. [Google Scholar]
- 16.Selten R. A note on evolutionary stable strategies in asymmetric animal conflicts. J Theor Biol. 1980;84:93–101. doi: 10.1016/s0022-5193(80)81038-1. [DOI] [PubMed] [Google Scholar]
- 17.Hammerstein P. The role of asymmetries in animal contests. Anim Behav. 1981;29:193–205. [Google Scholar]
- 18.Marshall JAR. The donation game with roles played between relatives. J Theor Biol. 2009;260:386–391. doi: 10.1016/j.jtbi.2009.07.008. [DOI] [PubMed] [Google Scholar]
- 19.Milinski M. TIT FOR TAT in sticklebacks and the evolution of cooperation. Nature. 1987;325:533–535. doi: 10.1038/325433a0. [DOI] [PubMed] [Google Scholar]
- 20.Hamilton WD. Genetical evolution of social behavior I & II. J Theor Biol. 1964;7:1–52. doi: 10.1016/0022-5193(64)90038-4. [DOI] [PubMed] [Google Scholar]
- 21.Nowak MA. Five rules for the evolution of cooperation. Science. 2006;314:1560–1563. doi: 10.1126/science.1133755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Schuster S, Kreft JU, Schroeter A, Pfeiffer T. Use of game-theoretical methods in biochemistry and biophysics. J Biol Phys. 2008;34:1–17. doi: 10.1007/s10867-008-9101-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Perc M, Szolnoki A. Coevolutionary games – a mini review. BioSystems. 2010;99:109–125. doi: 10.1016/j.biosystems.2009.10.003. [DOI] [PubMed] [Google Scholar]
- 24.Lehmann L, Keller L. The evolution of cooperation and altruism - a general framework and a classification of models. J Evol Biol. 2006;19:1365–1376. doi: 10.1111/j.1420-9101.2006.01119.x. [DOI] [PubMed] [Google Scholar]
- 25.Marshall JAR. Ultimate causes and the evolution of altruism. Behav Ecol Sociobiol: in press. 2010.
- 26.Nowak MA, May RM. Evolutionary games and spatial chaos. Nature. 1992;359:826–829. [Google Scholar]
- 27.Albert R, Barabási AL. Statistical mechanics of complex networks. Rev Mod Phys. 2002;74:47–97. [Google Scholar]
- 28.Newman MEJ. The structure and function of complex networks. SIAM Review. 2003;45:167–256. [Google Scholar]
- 29.Boccaletti S, Latora V, Moreno Y, Chavez M, Hwang D. Complex networks: Structure and dynamics. Phys Rep. 2006;424:175–308. [Google Scholar]
- 30.Ohtsuki H, Hauert C, Lieberman E, Nowak MA. A simple rule for the evolution of cooperation on graphs and social networks. Nature. 2006;441:502–505. doi: 10.1038/nature04605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lehmann L, Keller L, Sumpter D. The evolution of helping and harming on graphs: the return of the inclusive fitness effect. J Evol Biol. 2007;20:2284–2295. doi: 10.1111/j.1420-9101.2007.01414.x. [DOI] [PubMed] [Google Scholar]
- 32.Grafen A. An inclusive fitness analysis of altruism on a cyclical network. J Evol Biol. 2007;20:2278–2283. doi: 10.1111/j.1420-9101.2007.01413.x. [DOI] [PubMed] [Google Scholar]
- 33.Nowak MA, May RM. The spatial dilemmas of evolution. Int J Bifurcat Chaos. 1993;3:35–78. [Google Scholar]
- 34.Nowak MA, Bonhoeffer S, May RM. More spatial games. Int J Bifurcat Chaos. 1994;4:33–56. [Google Scholar]
- 35.Ohtsuki H, Nowak MA. Evolutionary games on cycles. Proc R Soc Lond B. 2006;273:2249–2256. doi: 10.1098/rspb.2006.3576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Schlag KH. Why imitate, and if so, how? a bounded rational approach to multi-armed bandits. J Econ Theory. 1998;78:130–156. [Google Scholar]
- 37.Wang S, Szalay MS, Zhang C, Csermely P. Learning and innovative elements of strategy adoption rules expand cooperative network topologies. PLoS ONE. 2008;3:e1917. doi: 10.1371/journal.pone.0001917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Szabó G, Tőke C. Evolutionary prisoner's dilemma game on a square lattice. Phys Rev E. 1998;58:69–73. [Google Scholar]
- 39.Abramson G, Kuperman M. Social games in a social network. Phys Rev E. 2001;63:030901(R). doi: 10.1103/PhysRevE.63.030901. [DOI] [PubMed] [Google Scholar]
- 40.Ren J, Wang WX, Qi F. Randomness enhances cooperation: coherence resonance in evolutionary game. Phys Rev E. 2007;75:045101(R). doi: 10.1103/PhysRevE.75.045101. [DOI] [PubMed] [Google Scholar]
- 41.Chen XJ, Wang L. Promotion of cooperation induced by appropriate payoff aspirations in a small-world networked game. Phys Rev E. 2008;77:017103. doi: 10.1103/PhysRevE.77.017103. [DOI] [PubMed] [Google Scholar]
- 42.Vukov J, Szabó G, Szolnoki A. Cooperation in the noisy case: Prisoner's dilemma game on two types of regular random graphs. Phys Rev E. 2006;73:067103. doi: 10.1103/PhysRevE.73.067103. [DOI] [PubMed] [Google Scholar]
- 43.Santos FC, Pacheco JM. Scale-free networks provide a unifying framework for the emergence of cooperation. Phys Rev Lett. 2005;95:098104. doi: 10.1103/PhysRevLett.95.098104. [DOI] [PubMed] [Google Scholar]
- 44.Santos FC, Rodrigues JF, Pacheco JM. Graph topology plays a determinant role in the evolution of cooperation. Proc R Soc Lond B. 2006;273:51–55. doi: 10.1098/rspb.2005.3272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Poncela J, Gómez-Gardeñes J, Floría LM, Moreno Y. Robustness of cooperation in the evolutionary prisoner's dilemma on complex systems. New J Phys. 2007;9:184. [Google Scholar]
- 46.Assenza S, Gómez-Gardeñes J, Latora V. Enhancement of cooperation in highly clustered scale-free networks. Phys Rev E. 2008;78:017101. doi: 10.1103/PhysRevE.78.017101. [DOI] [PubMed] [Google Scholar]
- 47.Szolnoki A, Perc M, Danku Z. Towards effective payoffs in the prisoner's dilemma game on scale-free networks. Physica A. 2008;387:2075–2082. [Google Scholar]
- 48.Perc M. Evolution of cooperation on scale-free networks subject to error and attack. New J Phys. 2009;11:033027. [Google Scholar]
- 49.Zimmermann MG, Eguíluz V, Miguel MS. Coevolution of dynamical states and interactions in dynamic networks. Phys Rev E. 2004;69:065102(R). doi: 10.1103/PhysRevE.69.065102. [DOI] [PubMed] [Google Scholar]
- 50.Pacheco JM, Traulsen A, Nowak MA. Coevolution of strategy and structure in complex networks with dynamical linking. Phys Rev Lett. 2006;97:258103. doi: 10.1103/PhysRevLett.97.258103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Szolnoki A, Perc M, Danku Z. Making new connections towards cooperation in the prisoner's dilemma game. EPL. 2008;84:50007. [Google Scholar]
- 52.Poncela J, Gómez-Gardeñes J, Floría LM, Sánchez A, Moreno Y. Complex cooperative networks from evolutionary preferential attachment. PLoS ONE. 2008;3:e2449. doi: 10.1371/journal.pone.0002449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Szolnoki A, Perc M. Resolving social dilemmas on evolving random networks. EPL. 2009;86:30007. [Google Scholar]
- 54.Poncela J, Gómez-Gardeñes J, Traulsen A, Moreno Y. Evolutionary game dynamics in a growing structured population. New J Phys. 2009;11:083031. [Google Scholar]
- 55.Szolnoki A, Perc M. Emergence of multilevel selection in the prisoner's dilemma game on coevolving random networks. New J Phys. 2009;11:093033. [Google Scholar]
- 56.Wu B, Zhou D, Fu F, Luo Q, Wang L, et al. Evolution of cooperation on stochastic dynamical networks. PloS ONE. 2010;5:1–7. doi: 10.1371/journal.pone.0011187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.McNamara JM, Barta Z, Fromhage L, Houston AI. The coevolution of choosiness and cooperation. Nature. 2008;451:189–192. doi: 10.1038/nature06455. [DOI] [PubMed] [Google Scholar]
- 58.Hauert C. Effects of space in 2×2 games. Int J Bifurc Chaos. 2002;12:1531–1548. [Google Scholar]
- 59.Perc M. Coherence resonance in spatial prisoner's dilemma game. New J Phys. 2006;8:22. [Google Scholar]
- 60.Perc M, Marhl M. Evolutionary and dynamical coherence resonance in the pair approximated prisoner's dilemma game. New J Phys. 2006;8:142. [Google Scholar]
- 61.Tanimoto J. Promotion of cooperation by payoff noise in a 2×2 game. Phys Rev E. 2007;76:041130. doi: 10.1103/PhysRevE.76.041130. [DOI] [PubMed] [Google Scholar]
- 62.Traulsen A, Nowak MA, Pacheco JM. Stochastic payoff evaluation increases the temperature of selection. J Theor Biol. 2007;244:349–356. doi: 10.1016/j.jtbi.2006.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Szabó G, Vukov J, Szolnoki A. Phase diagrams for an evolutionary prisoner's dilemma game on two-dimensional lattices. Phys Rev E. 2005;72:047107. doi: 10.1103/PhysRevE.72.047107. [DOI] [PubMed] [Google Scholar]
- 64.Szolnoki A, Perc M, Szabó G. Topology-independent impact of noise on cooperation in spatial public goods games. Phys Rev E. 2009;80:056109. doi: 10.1103/PhysRevE.80.056109. [DOI] [PubMed] [Google Scholar]
- 65.Szabó G, Szolnoki A, Izsák R. Rock-scissors-paper game on regular small-world networks. J Phys A: Math Gen. 2004;37:2599–2609. [Google Scholar]
- 66.Watts DJ, Strogatz SH. Collective dynamics of ‘small world’ networks. Nature. 1998;393:440–442. doi: 10.1038/30918. [DOI] [PubMed] [Google Scholar]