

Abstract
Human immunodeficiency virus type 1 (HIV-1) subtype C, which is most predominant in sub-Saharan Africa as well as in Asia and India, is the most prevalent subtype worldwide. A large number of transcription factor families have been shown to be involved in regulating HIV-1 gene expression in T lymphocytes and cells of the monocyte-macrophage lineage. Among these, proteins of the CCAAT/enhancer binding protein (C/EBP) family are of particular importance in regulating HIV-1 gene expression within cells of the monocytic lineage during the course of hematologic development and cellular activation. Few studies have examined the role of C/EBPs in long terminal repeat (LTR)-directed viral gene expression of HIV-1 subtypes other than subtype B. Within subtype B viruses, two functional C/EBP sites located upstream of the TATA box are required for efficient viral replication in cells of the monocyte-macrophage lineage. We report the identification of three putative subtype C C/EBP sites, upstream site 1 and 2 (C-US1 and C-US2) and downstream site 1 (C-DS1). C-US1 and C-DS1 were shown to form specific DNA-protein complexes with members of the C/EBP family (C/EBPα, β, and δ). Functionally, within the U-937 monocytic cell line, subtype B and C LTRs were shown to be equally responsive to C/EBPβ-2, although the basal activity of subtype C LTRs appeared to be higher. Furthermore, the synergistic interaction between C/EBPβ-2 and Tat with the subtype C LTR was also observed in U-937 cells as previously demonstrated with the subtype B LTR.
Keywords: C/EBPβ, HIV-1, Subtype C, LTR, Transcription
1. Introduction
Human immunodeficiency virus type 1 (HIV-1) is divided into three groups, M (major), O (outlier), and N (non-M, non-O). Most HIV-1 isolates belong to the M group, which has been subdivided into at least nine subtypes: A–D, F–H, J–K, and several circulating recombinant forms (CRFs) [1–3]. Most experimental studies concerning HIV-1 center on subtype B, the predominant form in North America and Europe, which accounts for approximately 10% of HIV-1 infections worldwide. In contrast, subtype C, which causes more than 50% of HIV-1 infections and predominates in southern Africa, India, and other parts of Asia, has been under investigated [4,5].
Several studies have demonstrated that different HIV-1 subtypes have distinct long terminal repeat (LTR) structures that represent the promoter sequences that drive HIV-1 gene expression and have been shown to influence viral transcription and replication in a cell type–specific manner [6–8][9]. For example, several studies have demonstrated the presence of three nuclear factor (NF)-κB binding sites within the subtype C LTR, whereas most other subtypes contain two NF-κB binding sites. One NF-κB binding site has been identified in CRF01_AE [10,11]. The extra NF-κB site in the subtype C LTR has been shown to be related to the high level of basal transcription and increased tumor necrosis factor α–induced transcriptional activity [10,12,13]. Three CCAAT/enhancer binding protein (C/EBP) sites (B-US1, B-US2, and B-US3) located upstream of the transcriptional start site have been identified within the subtype B LTR [14]. Activation of one of the C/EBP binding sites (B-US1 or B-US2) through binding of C/EBP factors is required for HIV-1 subtype B transcription and replication in cells of the monocyte-macrophage lineage [15,16]. Studies have shown, however, that C/EBP B-US2, which is present in the −170 region of the HIV-1 subtype B LTR, is not present in the subtype C LTR [17]. No other information has been reported regarding the existence or function of C/EBP sites within the subtype C LTR.
The C/EBP family of proteins belong to the basic region leucine zipper transcription factor family, which includes at least six members: C/EBPα, β, γ, δ, ε, and ζ [18]. C/EBP family members, especially C/EBPβ, have been reported to regulate HIV-1 transcription within different cell types (monocyte-macrophages, T cells, glial and microglia cells, endothelial cells, and hepatocytes) through various mechanisms [15,16,19–25], including interacting with several other transcription factors that include NF-κB [26,27], Sp1 [20], activating transcription factor/cyclic adenosine monophosphate–responsive element binding protein (ATF/CREB) [28–30], viral proteins Vpr and Tat [31–33], and chromatin remodeling factors, CREB binding protein/p300 and SWI/SNF (switch/sucrose nonfermentable) [22,34,35]. There are three isoforms of human C/EBPβ, C/EBPβ-1, -2, and -3 [36]. C/EBPβ-1 and -2 isoforms act as transcriptional activators and C/EBPβ-3 functions as a transcriptional repressor [36].
CCR5 (R5) and CXCR4 (X4) are the predominant co-receptors for HIV-1 entry (for review see Clapham and McKnight [37]). CCR5 acts as the major co-receptor for macrophage-tropic HIV-1 strains [38], whereas T-cell line–adapted viruses use CXCR4 [39,40]. Interestingly, most subtype C viruses examined to date display a preferential usage of the CCR5 co-receptor [41–43]. Based on the role of the C/EBP transcription factor family in HIV-1 subtype B replication in cells of the monocyte-macrophage lineage and CCR5 usage appearing to be more frequent among subtype C viruses, we sought to determine whether C/EBP binding site(s) also exist in the subtype C LTR and to determine the functional properties of the C/EBP binding sites within the context of HIV-1 subtype C transcription.
The HIV-1 transactivator protein (Tat) is critical for HIV-1 transcription and regulation (reviewed in [44–47]) by interacting with the transactivation-responsive region, located at the 5′ end of all nascent HIV-1 transcripts [48,49]. Variation in viral genomic sequence has been reported, as well as the resultant amino acid changes and subtype-specific effects of Tat protein among the different HIV-1 subtypes [10,11,50–52]. Specifically, Tat proteins from subtype C and E have been shown to drive higher levels of viral gene expression than subtype B Tat regardless of the origin of the HIV-1 LTR [13,50]. The increased level of viral gene expression observed with subtype C and E is related in part to the longer half-life of Tat within infected cells [50].
The studies reported here have determined that one C/EBP binding site exists both upstream and downstream of the transcriptional start site within HIV-1 subtype C LTR. In general, the subtype C LTR exhibited a higher basal transcription level compared with the subtype B LTR under the experimental conditions examined. C/EBPβ-2 was able to activate HIV-1 subtype C LTR-driven transcription in the U-937 promonocytic cell line with an overall level of transcription close to that of the subtype B LTR. A further increase in subtype C LTR activity was observed in the presence of both C/EBPβ-2 and subtype C Tat.
2. Material and Methods
2.1. Cell culture and nuclear extract preparation
The U-937 human monocytic cell line (American Type Culture Collection, CRL-1593.2) was grown in RPMI-1640 (Roswell Memorial Park Institute medium) (Cellgro, Mediatech, Manassas, VA). Medium was supplemented with 10% heat-inactivated fetal bovine serum (HyClone, Logan, UT), antibiotics (penicillin, 100 U/ml, and streptomycin, 100 µg/ml; Cellgro), glucose (4.5 g/ml, Cellgro), sodium pyruvate (1 mM, Cellgro), and HEPES [4-(2-hydroxyethyl)-1-piperazine ethane sulfonic acid], 10 mM; Cellgro). The cells were maintained at 37°C with 5% CO2 in 90% relative humidity.
Nuclear extracts were prepared as described [53]. Low-passage, exponentially growing cells (1 × 107) were collected by centrifugation, washed with cold phosphate-buffered saline (Cellgro), and lysed in ice-cold lysis buffer. After centrifugation (1000 × g), the supernatant was discarded and the pelleted nuclei were resuspended in nuclear extract buffer, shaken vigorously for 30 min at 4°C, and subjected to centrifugation for 15 min (14,000 × g). The supernatant was collected, frozen in liquid nitrogen, and stored at −80°C. The protein concentration was determined by Bradford assay as described by the manufacturer (Bio-Rad, Hercules, CA).
2.2. Electrophoretic mobility shift (EMS) analyses
Complementary single-stranded DNA oligonucleotides were synthesized by Integrated DNA Technologies (Coralville, IA). Probes were annealed by heating at 100°C for 5 min and slowly cooling to room temperature. Probes were 5′ end-labeled using T4 DNA kinase (Promega, Madison, WI) and [γ-32P] adenosine triphosphate. Labeled probes (75,000 cpm/lane) were incubated with 2 µg of nonspecific DNA (poly [d(I-C)], Roche, Indianapolis, IN) and 20 µg of U-937 nuclear extract at 30 °C for 30 min. Competition studies were performed using unlabeled competitor oligonucleotides at concentrations up to 100-fold molar excess as compared with the probe concentration. Supershift analyses were initiated by incubation of the complexes for another 15 min with antibodies specific to C/EBPα (sc-9315x), C/EBPβ (sc-150x), C/EBPγ (sc-7659x), C/EBPε (sc-31929x), or C/EBPδ (sc-636x) (all from Santa Cruz Biotechnologies, Santa Cruz, CA). DNA-protein complexes were resolved on 5% nondenaturing polyacrylamide gels at 200 V at 4°C for approximately 2 h. Gels were dried at 80°C for 1 h and subjected to autoradiography.
2.3. Plasmids
The subtype C LTR-containing DNA fragment (approximately 630 bp) was derived from the HIV-1 subtype C molecular clone (pMJ4, National Institutes of Health AIDS Research and Reference Reagent Program, Rockville, MD). The HIV-1 subtype C LTR was PCR-amplified using the forward primer: 5′-GGGGTACCTGGAAGGGTTAATTTACTCT-3′ and reverse primer: 5′-CGACGCGTTAGAG ATTTTCCTCACTACC-3′ (Integrated DNA Technologies). The italicized nucleotides indicate the restriction endonuclease sites. The amplified product was digested with KpnI and MluI (Promega) and ligated into a modified pGL3-basic vector, which contained the firefly luciferase (Luc) gene (Promega), to construct the parental HIV-1 subtype C LTR-luciferase (C-LTR-Luc) expression construct. The HIV-1 LAI LTR was cloned into the pGL3-basic vector as previously described [54].
The C/EBPβ-2 expression construct was generated by PCR amplification from human C/EBPβ cDNA (Open Biosystems, Human Verified Full-length cDNA Clone, MHS 1011, Huntsville, AL) using forward primers for C/EBPβ-2, 5′-CACCATGGAAGTGGCCAACTTCTACTA-3′ and the reverse primer 5′-CTAGCAGTGGCCGGAGGAGGCGAG-3′ (Integrated DNA Technologies). The italicized nucleotides in the forward primer correspond to the sequence necessary for directional cloning into the pcDNA3.1 TOPO vector (Invitrogen, San Diego, CA), and the underlined portion corresponds to the respective start site of translation. The amplified C/EBPβ-2 PCR product was ligated into the pcDNA3.1 TOPO vector as described by the manufacturer (Invitrogen). The plasmids were verified by sequencing (Genewiz, South Plainfield, NJ). To confirm expression, 30 µg of the C/EBPβ-2 construct was transiently transfected into 3.0 × 107 293F cells using 293 fectin as described by the manufacturer (Invitrogen). Forty-eight hours after transfection, nuclear extracts were collected and western immunoblot analyses were performed on nuclear extracts using C/EBPβ antibody (C/EBPβ, C-19, Santa Cruz Biotechnologies) for detection of a 45-kDa protein, C/EBPβ-2. Electrophoretic mobility shift (EMS) supershift analyses were performed to determine the identity of proteins involved in DNA-protein complex formation [Liu and Wigdahl, unpublished observations].
2.4. Transient expression analyses
Exponentially growing U-937 cells (1 × 106) were seeded in 2 ml of growth medium in six-well plates. For each transfection, FuGENE 6 Transfection Reagent (6 µl) (Roche) was dispensed into serum-free medium (94 µl). After 5 min, the solution was added to the DNA dropwise, incubated for 15 min, and dispensed dropwise into the cell culture. Cells were transfected with 1000 ng firefly luciferase LTR expression constructs (C-LTR-Luc or B-LTR-Luc) in conjunction with the thymidine kinase promoter-Renilla luciferase reporter plasmid (pRL-TK) internal control vector (50 ng). Cells were harvested 24 h after the addition of the FuGENE 6 and DNA. Cell lysates were harvested and assayed using the Dual Luciferase Assay system as described by the manufacturer (Promega). Firefly luminescence was normalized to Renilla luminescence to control for variation in transfection efficiency. For the experiments that included C/EBPβ-2 expression vectors, single luciferase assays were performed because C/EBPβ-2 has been shown to activate the pRL-TK promoter [53]; Liu and Wigdahl, unpublished observations]. Cell lysates were first quantified by Bradford assays (Promega) and cell lysates (30 µg per sample) were used for performance of single luciferase assays. The results are presented as mean values with the indicated standard deviations. Each value shown represents the average of two or three independent experiments performed in triplicate.
2.5. Statistical analysis
The results were statistically analyzed by Student’s t-test. Differences between groups were considered significant if p values <0.05 were obtained.
3. Results
3.1. Identification of C/EBP binding sites within the HIV-1 subtype C LTR
To determine whether the HIV-1 subtype C LTR contained C/EBP binding site(s), 211 subtype C LTR sequences were collected from GenBank and Los Alamos HIV sequence databases. The consensus sequence of the subtype C LTR was determined by aligning the 211 LTR sequences using the MegAlign program (DNAStar, Madison, WI) with the Clustal W algorithm (European Bioinformatics Institute, Cambridge, UK). The LTR subtype C consensus sequence was analyzed using the Web-based program TRANSFAC (P-Match; Biobase Biological Databases, Wolfenbüttel, Germany), (0.75 core similarity, 0.85 matrix similarity threshold) for the presence of putative C/EBP binding sites. As a result of this analysis, two sites were located upstream and one site was located downstream of the transcriptional start site within the subtype C LTR. They were designated subtype C C/EBP upstream site 2 (C-US2), upstream site 1 (C-US1), and subtype C C/EBP downstream site 1 (C-DS1). C-US2 was located at nucleotide position −446 to −433; C-US1 was located at position −259 to −247 and C-DS1 was located at position +171 to +184, relative to the transcriptional start site (Fig. 1A). When compared with the three identified upstream C/EBP binding sites within subtype B LTR (B-US3, B-US2, and B-US1, Fig. 1A), the physical location and nucleotide sequences of subtype C were very different (Fig. 1A and B). In contrast, the physical location and nucleotide sequence of the C/EBP binding site located downstream of the transcriptional start site were similar between subtype B and C LTRs (Fig. 1A and B).
Fig. 1.
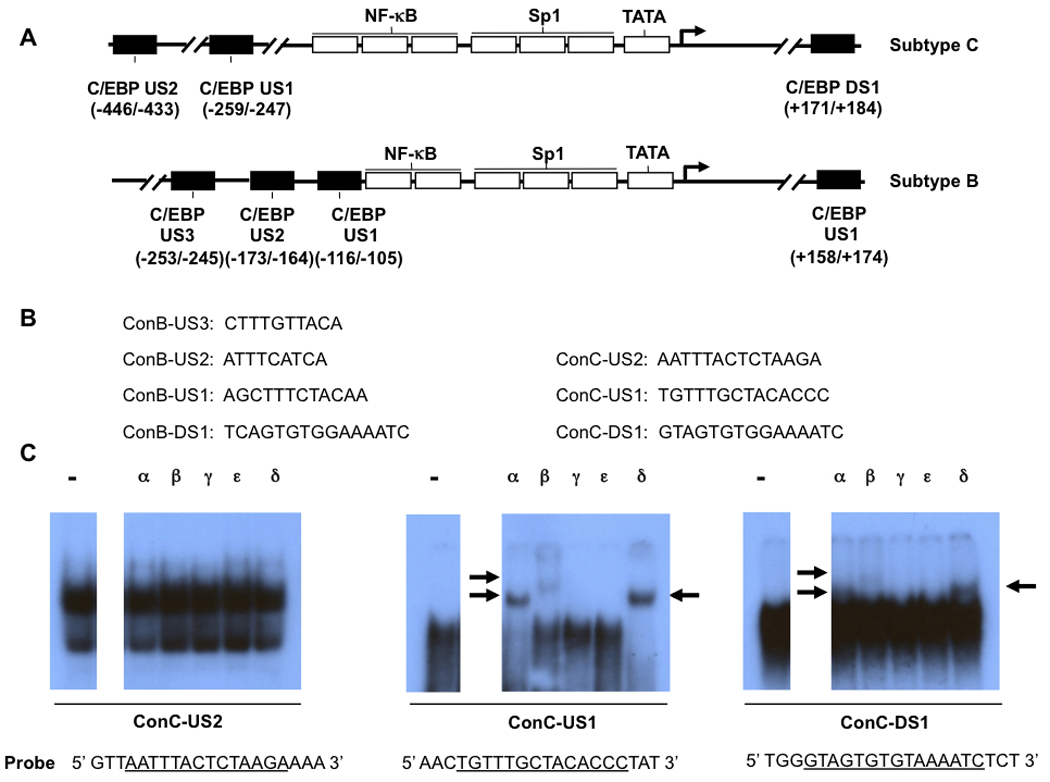
Members of the C/EBP transcription factor family bind to subtype C LTR. (A) The location of three putative C/EBP binding sites in subtype C LTR and four C/EBP binding sites within subtype B LTR are shown. (B) Sequence comparison of C/EBP sites between subtype B and C. (C) Consensus subtype C-US1 (ConC-US1) was able to form DNA-protein complexes containing C/EBPα, β, and δ. Supershift EMS analyses were performed as described in “Materials and Methods.” Radiolabeled ConC-US2, ConC-US1, and ConC-DS1 were used as probes and reacted with U-937 nuclear extract in the absence or presence of antibodies specific to C/EBPα, β, γ, ε, and δ. Arrows show the specific DNA-protein complexes.
To determine whether the three putative C/EBP binding sites were able to form DNA-protein complexes with C/EBP protein family members, supershift EMS analyses were performed. The oligonucleotides representing the three putative subtype C consensus C/EBP binding sites were radiolabeled and used as probes (Fig. 1C) and incubated with nuclear extracts prepared from U-937 monocytic cells. Antibodies specific to C/EBPα, β, γ, ε, or δ were used in EMS reactions. As shown in Fig. 1C, C-US2 was unable to form specific DNA-protein complexes and was therefore eliminated from further analysis. In contrast, the DNA-protein complexes formed by C-US1 or C-DS1 were shifted by antibodies specific to C/EBPα, β, and δ (Fig. 1C). These results indicated that the subtype C C/EBP binding sites C-US1 and C-DS1 had the ability to form specific DNA-protein complexes containing C/EBPα, β, and δ.
3.2. Comparison of C/EBP binding sites and other important transcription factor binding sites between subtype B and C LTRs
The interactions between subtype B C/EBP US1 (B-US1) and transcription factor binding sites located in proximity, including ATF/CREB, NF-κB, and Sp site III binding sites, have been reported [26,28,30]; Burdo and Wigdahl, unpublished observations]. To determine whether similar transcription factor binding sites were also located near subtype C-US1, the subtype C LTR consensus sequence was analyzed by TRANSFAC. Sequence analyses (0.75 core similarity, 0.85 matrix similarity threshold), however, failed to identify any transcription factor binding sites known to be important for HIV-1 transcription located in the 5′ or 3′ sequences immediately adjacent to C-US1. In contrast, the ATF/CREB site located 5′ of B-US1 also existed immediately 5′ of the promoter-distal NF-κB site in the subtype C LTR with the same physical location and only two nucleotide differences (Fig. 2). Also, three Sp binding sites (Sp sites I, II, and III) were located 3′ to the promoter-proximal NF-κB binding site in both subtype B and C LTRs with the same physical location and highly conserved nucleotide sequence (Fig. 2). Compared with consensus sequences of subtype B Sp binding sites, consensus sequences of subtype C Sp binding sites exhibited similar nucleotide sequences; Sp site I contained only one nucleotide variation (C-to-T change at position 10), and Sp site III contained three nucleotide variations (a deletion at position 1 and a C-to-T change at positions 5 and 10). With respect to the subtype C NF-κB binding sites, the nucleotide sequences of NF-κB site I and II were conserved between subtypes C and B (Fig. 2). The extra NF-κB binding site (NF-κB III), however, exhibited nucleotide sequences different from that of the subtype B or C consensus sequence.
Fig. 2.

Alignment and comparison of consensus sequences of indicated transcription factor binding sites within the LTR of HIV-1 subtypes B and C. Representative transcription factor binding sites within the HIV-1 subtype B and subtype C consensus sequences are shown. Arrow indicates the transcriptional start site.
The prevalences of the consensus sequence within the C/EBP US1 site, ATF/CREB site, NF-κB sites, and Sp sites I, II, and III were compared between the subtype B and C LTRs. For subtype B, the sequences from 348 peripheral blood–derived LTRs documented in three published studies [55–57] were used in the analyses of C/EBP US1; Sp sites I, II, III; and NF-κB site II. In addition, 267, 223, and 131 LTRs were used in C/EBP US2, NF-κB site I, and ATF/CREB site analyses, respectively. For subtype C LTR analysis, among the 211 collected LTRs, 197 LTRs have been used for three NF-κB sites, Sp sites I and II, and ATF/CREB binding sites with 205 and 185 LTRs used for C/EBP US1 site and Sp site III analyses, respectively. In the analyses in which a lower number of sequences were used, sequences for these regions were not available. Based on these analyses, C/EBP US1 exhibited similar conservation between subtype B (52.8%) and C (50.7%) (Fig. 3). Interestingly, among the subtype C transcription factor binding sites examined, the ATF/CREB site was highly conserved (93.9%); this was much higher than the subtype B ATF/CREB site (74.8%). The two classical NF-κB binding sites and Sp site II from subtype B and C LTRs also exhibited a high degree of conservation (subtype C NF-κB site II, I, Sp site II was 89.3%, 95.9% and 89.8%, respectively; subtype B NF-κB site II, I, Sp site II was 96%, 91%, and 83%, respectively), which was correlated with the important roles these transcription factor binding sites play in HIV-1 transcription, whereas Sp site I was significantly less conserved (36%) in the subtype C LTR than in subtype B LTR (86.2%). Currently, it is not clear why Sp site I is not conserved in subtype C. It has been suggested that this site may be critical for subtype B gene expression. The differences noted in relative conservation warrant further investigation. Importantly, the degree of conservation of Sp site III, which is known to be a critical regulator of HIV-1 replication in both T cells and cells of the monocyte-macrophage lineage, is similar for the both the subtype B and subtype C LTRs (32.5% and 43.2%, respectively). For the C/EBP binding site located downstream of the transcriptional start site within subtype B LTR, 77 of 100 LTRs (77%) exhibited the consensus sequence. With respect to the upstream C/EBP binding site located within subtype C LTR, 66 of 115 (57%) were found to represent the consensus sequence configuration [Liu and Wigdahl, unpublished observations].
Fig. 3.

Prevalence of the consensus configurations of indicated transcription binding sites between subtypes B and C. (A) Selected transcription binding sites within HIV-1 consensus subtype B (ConB) and C (ConC) LTRs. The arrow indicates the transcriptional start site. (B) HIV-1 subtype B LTRs from three published papers [55–57] and subtype C LTRs from GenBank and Los Alamos databases were compiled and aligned using the Clustal W method. The number shown indicates the percentage of each of the indicated transcription binding sites with the respective consensus configuration present in the total HIV-1 LTR sequences available for inspection.
3.3. Differing number and location of C/EBP binding sites within the subtype B and C LTRs, yet similar level of response to the transactivator C/EBPβ
Previous studies have demonstrated that representative subtype C LTRs exhibited higher basal transcription levels compared with several subtype B LTRs; however, most of these experiments were performed in T-cell lines and primary T lymphocytes [10,13,58,59]. Given that most isolated subtype C viruses preferentially use CCR5 as a co-receptor throughout the course of disease and therefore may rely more heavily on replication in cells of the monocyte-macrophage lineage as compared with a subtype B virus, which usually switches co-receptor usage from CCR5 to CXCR4 as HIV disease progresses, basal transcription levels of representative subtype B and C LTRs were compared in U-937 monocytic cells. Plasmids containing parental subtype B LTR (LAI) or a subtype C LTR (pMJ4) derived from a clinical isolate and obtained from the AIDS Reference and Reagent Program were transiently transfected into U-937 monocytic cells and LTR function was assessed based on the expression of the luciferase indicator gene. The representative subtype C LTR exhibited approximately 25% higher LTR basal activity than the representative subtype B LTR in U-937 monocytic cells (Fig. 4A). HIV-1 subtype C LTR transcription levels as a result of co-transfection with the C/EBPβ-2 expression vectors (Fig. 4B) were also examined in U-937 monocytic cells (Fig. 4C). C/EBPβ-2 (activator) was able to transactivate HIV-1 LTR transcription (Fig. 4B). As shown in Fig. 4C, in the presence of C/EBPβ-2, the transcription level of the subtype C LTR was enhanced up to 5.2-fold in U-937 cells. In parallel, the same assays were performed with the subtype B LTR. These results reveal that the level of transcription was increased up to 5.3-fold in U-937 cells. As shown, there was no significant difference with respect to C/EBPβ-2 activation of either the subtype B or C LTRs, even with the different C/EBP binding site number, nucleotide sequences, and/or physical location.
Fig. 4.

C/EBPβ-2 is able to activate the HIV-1 subtype C LTR. (A) and (C) U-937 monocytic cells were transiently transfected with luciferase reporter constructs under the control of subtype B or subtype C parental LTR in the absence (A) or presence of C/EBPβ-2 expression vector (C). Relative luciferase activity was quantitated 24 h after transfection. Results are shown as “fold” difference over the parental subtype C LTR. (B) C/EBPβ-2 isoform structure. Asterisk indicates the significant difference (p value <0.001). Error bars indicate the standard deviation for each data point.
3.4. Increased transactivation of subtype C LTR in the presence of Tat and C/EBPβ as compared with Tat alone
The transcription level of the subtype C LTR activated by subtype C Tat was determined in U-937 monocytic cells. As expected, in the presence of a subtype C Tat expression vector, the transcription level was enhanced up to 35.5-fold. Previous studies have shown the synergistic interaction between subtype B Tat and C/EBPβ-2 with respect to activating the subtype B LTR [27]. Similarly, the combined presence of subtype C Tat and C/EBPβ-2 resulted in a 73.7-fold increase in subtype C LTR transcriptional activity (Fig. 5).
Fig. 5.

The activities of HIV-1 subtype C LTR caused by Tat and C/EBPβ-2 in U-937 cells. U-937 cells were transiently transfected with parental subtype C LTR in the absence or presence of C/EBPβ-2, Tat alone, or in combination. Twenty-four hours after transfection, cell lysates were collected and followed by protein quantification and luciferase activity assays. Results are shown as “fold” difference over parental LTR. Error bars indicate the standard deviation for each data point.
4. Discussion and conclusion
Previous studies have demonstrated that (1) the corresponding subtype B C/EBP US2 does not exist in the subtype C LTR [17]; (2) subtype B US1 location has been replaced by an NF-κB site in the subtype C LTR; (3) C/EBP binding sites (US2 or US1) are required for HIV-1 subtype B transcription and replication in cells of the monocytic-macrophage lineage [15,16,25]; and (4) subtype C viruses prefer to use CCR5 as a co-receptor for virus entry throughout the course of HIV disease [41–43]. It has not been demonstrated, however, whether C/EBP binding sites exist within the HIV-1 subtype C LTR and, if they are present, whether they function within target cells representative of cells encountered by the subtype C virus during the course of disease. Our results demonstrate the presence of C/EBP binding sites within the subtype C LTR, located upstream and downstream of the transcriptional start site. Although the C/EBP binding site located upstream of the transcriptional start site within the subtype C LTR exists in a distinct location and also displays a unique spectrum of genotypic alternations as compared with the subtype B upstream functional C/EBP sites (B-US2 and -US1), both LTRs have been shown to be important for viral transcription and were shown to exhibit similar activity in the presence of C/EBPβ-2, suggesting that the function of C/EBPβ transactivation is conserved and important for HIV-1 transcription in different subtypes. Therefore, it would be interesting to investigate the roles of C/EBP factors and C/EBP sites in different HIV-1 subtypes. The location of C-US1 is close to that of B-US3 (Fig. 3A), which is not required for HIV-1 subtype B LTR transcription [16,19]. Thus, it is possible that the different genotype of C-US1 results in the functional C/EBP binding site or that C-DS1 plays an important role in C/EBPβ-2-mediated transactivation of the subtype C LTR. It remains possible, however, that the extra NF-κB binding site or some other transcription factor binding sites within the subtype C LTR impacts C/EBP-mediated function and plays a role in compensating for C/EBP binding site differences between subtype B and C LTRs. For example, the transcriptional activation of the HIV-1 LTR by p50-C/EBPβ heterodimeric complexes has been demonstrated within subtype B [26] and this functional interaction has been shown to be dependent on the DNA-binding ability of p50 and the transcription activation domain of C/EBPβ. With regard to the subtype C LTR, three NF-κB binding sites may result in enhanced binding of C/EBPβ–NF-κB heterodimeric complexes to the LTR and result in increased activation of the LTR. Whether the C/EBP sites are required for HIV-1 subtype C replication in mononcyte-macrophage cell lineages remains to be elucidated. The level of basal transcription driven by the representative subtype C LTR was higher than that of the representative subtype B LTR in U-937 monocytic cells (Fig. 4), which is consistent with previous studies that have been performed in Jurkat T-cell lines [13], suggesting that the role of the extra NF-κB site in subtype C LTR basal activity is independent of cell type.
Different HIV subtypes have been shown to exhibit different biologic properties, pathogenesis, disease progression, and antiretroviral drug resistance [60–64]. Specifically, differences in selected LTR regions may be associated with alterations in virus transcription, replication, and disease progression [6,10]. Interestingly, the subtype B Sp III-5T sequence variant (C-to-T change at position 5 compared with the consensus B Sp site III) that has been shown to be correlated with HIV-1 subtype B disease progression [65] appeared in the subtype C Sp site III consensus sequence. Whether this LTR sequence alteration is related to HIV-1 subtype C disease progression will require additional clinical information, viral genotyping, and molecular pathogenesis analyses to definitively delineate this potential structure/function correlation. The conservation of subtype C Sp site I was relatively low (36%) in the analysis, which is different from other reports showing that subtype C Sp site I [8] and Sp site II are the most conserved [66] among the three Sp sites. The number of LTRs examined in these studies was less than the number used in the present studies, however, and the function of Sp sites or the GC box array in subtype C gene expression needs additional investigation. Unexpectedly, subtype C virus exhibited a relatively high degree of conservation in the ATF/CREB site (percentage of consensus sequence: 93.9%, Fig. 3), which was comparable to that of the NF-κB sites. The function of the ATF/CREB site in subtype C gene expression is not known. Subtype B ATF/CREB site variants (TGACACAA, including C-to-A and G-to-A changes at position 7 and 8, respectively), however, were able to affect subtype B LTR basal activity and LTR function following interleukin-6 stimulation, a treatment that leads to increases in C/EBPβ activation [30]. This variant sequence is also similar to the subtype C ATF/CREB consensus sequence (TGACACAG). Therefore, it is possible that the subtype C ATF/CREB site is able to affect LTR basal transcription or transcription mediated by its corresponding transcription factors or the nearby NF-κB sites. Phenotypically, selected consensus sequences of subtype C (Sp site III 5T, ATF/CREB site 7C and 8C) have been found in subtype B as naturally occurring sequence variations and have been shown to exhibit low DNA binding ability. Others have reported that sequence variations in one transcriptional binding site affected DNA binding affinities of other neighboring transcriptional binding sites and viral proteins, and ultimately the promoter activation potential of subtype B LTRs [30,31]. Thus, it is possible that the replacement of C/EBP US1 and the low DNA binding affinity of the ATF/CREB and Sp site III binding sites within the subtype C LTR act together to decrease DNA binding competition with NF-κB, resulting in a better structural environment for NF-κB factors to bind the enhancer element rapidly and efficiently, which might be one of the factors contributing to subtype C replication and pathogenesis. Perhaps these specific LTR alterations facilitate viral replication in cells of the monocyte-macrophage lineage and enhance viral transmission capabilities.
As reported in other studies, subtype C LTR activities are enhanced in the presence of subtype C Tat in HEK293 cells and in Jurkat T cells [13,50,67]. The studies reported here have shown that subtype C LTR activities are also activated by subtype C Tat in U-937 monocytic cells (Fig. 5). The physical and functional interaction between C/EBPβ and subtype B Tat-regulating gene expression has been reported [27,32,33,68]. The cooperation of subtype B Tat and C/EBPβ has been shown to increase C/EBPβ DNA binding affinity for the HIV-1 LTR [27,33] and to regulate transcription of monocyte chemoattractant protein 1 in astrocytes [32]. Consequently, it has been proposed that subtype B Tat and C/EBPβ interaction contributes to HIV-1-associated brain dysfunction in patients with HIV-1 subtype B. Studies to date have shown, however, that HIV-1 subtype C is associated with a lower prevalence of HIV-1-associated dementia in India [51], although the exact role of subtype C in neurologic disease requires further investigation. Our results have demonstrated that subtype C Tat maintains transactivation capability and the ability to interact with other transcription factors, such as C/EBPβ, in U-937 cells. Thus, the interaction of subtype C Tat and C/EBP may not be associated with low occurrence of HIV-associated dementia. As reported previously [51,52,67], the relatively low occurrence of HIV-associated dementia in individuals infected with subtype C virus might be caused by other properties of subtype C Tat, including deficient chemoattractant properties, low binding affinity for chemokine receptor 2 (CCR2), or an inability to stimulate tumor necrosis factor-α production as a result of a serine-to-cysteine substitution at amino acid position 31.
In summary, two C/EBP binding sites have been identified within the HIV-1 subtype C LTR. Additional studies have suggested that transactivation ability of C/EBPβ may play an important functional role in the pathogenesis of different HIV-1 subtypes. Our results also suggest that viral LTR sequence variation in different subtypes as a result of viral genomic alterations and host selective pressures may be associated with differences in viral replication and disease progression. Further studies will determine how C/EBP US1 and DS1 affect subtype C gene expression in cells of monocyte-macrophage lineage and examine the roles of conserved ATF/CREB and Sp binding sites in HIV-1 subtype C gene expression.
Acknowledgements
These studies were funded in part by the Public Health Service, National Institutes of Health, through grants (B. Wigdahl, Principal Investigator) from the National Institute of Neurological Disorders and Stroke (NS32092 and NS46263) and the National Institute of Drug Abuse (DA19807).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Myers G, Korber B, Foley B, Jeang K-T, Mellers JW, Wain-Hubson S. Human Retroviruses and AIDS 1996: A compilation and Analysis of Nucleic Acid and Amino Acid Sequences. Los Alamos Nat Lab. 1996 [Google Scholar]
- 2.Korber B, Muldoon M, Theiler J, Gao F, Gupta R, Lapedes A, et al. Timing the ancestor of the HIV-1 pandemic strains. Science. 2000;288:1789–1796. doi: 10.1126/science.288.5472.1789. [DOI] [PubMed] [Google Scholar]
- 3.Zhang M, Foley B, Schultz AK, Macke JP, Bulla I, Stanke M, et al. The role of recombination in the emergence of a complex and dynamic HIV epidemic. Retrovirology. 2010;7:25. doi: 10.1186/1742-4690-7-25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Geretti AM. HIV-1 subtypes: epidemiology and significance for HIV management. Curr Opin Infect Dis. 2006;19:1–7. doi: 10.1097/01.qco.0000200293.45532.68. [DOI] [PubMed] [Google Scholar]
- 5.Arien KK, Vanham G, Arts EJ. Is HIV-1 evolving to a less virulent form in humans? Nat Rev Microbiol. 2007;5:141–151. doi: 10.1038/nrmicro1594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.van Opijnen T, Jeeninga RE, Boerlijst MC, Pollakis GP, Zetterberg V, Salminen M, et al. Human immunodeficiency virus type 1 subtypes have a distinct long terminal repeat that determines the replication rate in a host-cell-specific manner. J Virol. 2004;78:3675–3683. doi: 10.1128/JVI.78.7.3675-3683.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Centlivre M, Sommer P, Michel M, Ho Tsong Fang R, Gofflo S, Valladeau J, et al. HIV-1 clade promoters strongly influence spatial and temporal dynamics of viral replication in vivo. J Clin Invest. 2005;115:348–358. doi: 10.1172/JCI22873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.De Arellano ER, Soriano V, Holguin A. Genetic analysis of regulatory, promoter, and TAR regions of LTR sequences belonging to HIV type 1 Non-B subtypes. AIDS Res Hum Retroviruses. 2005;21:949–954. doi: 10.1089/aid.2005.21.949. [DOI] [PubMed] [Google Scholar]
- 9.Kilareski EM, Shah S, Nonnemacher MR, Wigdahl B. Regulation of HIV-1 transcription in cells of the monocyte-macrophage lineage. Retrovirology. 2009;6:118. doi: 10.1186/1742-4690-6-118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jeeninga RE, Hoogenkamp M, Armand-Ugon M, de Baar M, Verhoef K, Berkhout B. Functional differences between the long terminal repeat transcriptional promoters of human immunodeficiency virus type 1 subtypes A through G. J Virol. 2000;74:3740–3751. doi: 10.1128/jvi.74.8.3740-3751.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Montano MA, Novitsky VA, Blackard JT, Cho NL, Katzenstein DA, Essex M. Divergent transcriptional regulation among expanding human immunodeficiency virus type 1 subtypes. J Virol. 1997;71:8657–8665. doi: 10.1128/jvi.71.11.8657-8665.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Montano MA, Nixon CP, Ndung'u T, Bussmann H, Novitsky VA, Dickman D, et al. Elevated tumor necrosis factor-alpha activation of human immunodeficiency virus type 1 subtype C in Southern Africa is associated with an NF-kappaB enhancer gain-of-function. J Infect Dis. 2000;181:76–81. doi: 10.1086/315185. [DOI] [PubMed] [Google Scholar]
- 13.Roof P, Ricci M, Genin P, Montano MA, Essex M, Wainberg MA, et al. Differential regulation of HIV-1 clade-specific B, C, and E long terminal repeats by NF-kappaB and the Tat transactivator. Virology. 2002;296:77–83. doi: 10.1006/viro.2001.1397. [DOI] [PubMed] [Google Scholar]
- 14.Tesmer VM, Rajadhyaksha A, Babin J, Bina M. NF-IL6-mediated transcriptional activation of the long terminal repeat of the human immunodeficiency virus type 1. Proc Natl Acad Sci U S A. 1993;90:7298–7302. doi: 10.1073/pnas.90.15.7298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Henderson AJ, Calame KL. CCAAT/enhancer binding protein (C/EBP) sites are required for HIV-1 replication in primary macrophages but not CD4(+) T cells. Proc Natl Acad Sci U S A. 1997;94:8714–8719. doi: 10.1073/pnas.94.16.8714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Henderson AJ, Connor RI, Calame KL. C/EBP activators are required for HIV-1 replication and proviral induction in monocytic cell lines. Immunity. 1996;5:91–101. doi: 10.1016/s1074-7613(00)80313-1. [DOI] [PubMed] [Google Scholar]
- 17.Zacharova V, Becker ML, Zachar V, Ebbesen P, Goustin AS. DNA sequence analysis of the long terminal repeat of the C subtype of human immunodeficiency virus type 1 from Southern Africa reveals a dichotomy between B subtype and African subtypes on the basis of upstream NF-IL6 motif. AIDS Res Hum Retroviruses. 1997;13:719–724. doi: 10.1089/aid.1997.13.719. [DOI] [PubMed] [Google Scholar]
- 18.Ramji DP, Foka P. CCAAT/enhancer-binding proteins: structure, function and regulation. Biochem J. 2002;365:561–575. doi: 10.1042/BJ20020508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tesmer VM, Bina M. Regulation of HIV-1 gene expression by NF-IL6. J Mol Biol. 1996;262:327–335. doi: 10.1006/jmbi.1996.0516. [DOI] [PubMed] [Google Scholar]
- 20.Schwartz C, Catez P, Rohr O, Lecestre D, Aunis D, Schaeffer E. Functional interactions between C/EBP, Sp1, and COUP-TF regulate human immunodeficiency virus type 1 gene transcription in human brain cells. J Virol. 2000;74:65–73. doi: 10.1128/jvi.74.1.65-73.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yang Y, Tesmer VM, Bina M. Regulation of HIV-1 transcription in activated monocyte macrophages. Virology. 2002;299:256–265. doi: 10.1006/viro.2001.1530. [DOI] [PubMed] [Google Scholar]
- 22.Lee ES, Sarma D, Zhou H, Henderson AJ. CCAAT/enhancer binding proteins are not required for HIV-1 entry but regulate proviral transcription by recruiting coactivators to the long-terminal repeat in monocytic cells. Virology. 2002;299:20–31. doi: 10.1006/viro.2002.1500. [DOI] [PubMed] [Google Scholar]
- 23.Burdo TH, Gartner S, Mauger D, Wigdahl B. Region-specific distribution of human immunodeficiency virus type 1 long terminal repeats containing specific configurations of CCAAT/enhancer-binding protein site II in brains derived from demented and nondemented patients. J Neurovirol. 2004;10 Suppl 1:7–14. doi: 10.1080/753312746. [DOI] [PubMed] [Google Scholar]
- 24.Hogan TH, Krebs FC, Wigdahl B. Regulation of human immunodeficiency virus type 1 gene expression and pathogenesis by CCAAT/enhancer binding proteins in cells of the monocyte/macrophage lineage. J Neurovirol. 2002;8 Suppl 2:21–26. doi: 10.1080/13550280290167911. [DOI] [PubMed] [Google Scholar]
- 25.Henderson AJ, Zou X, Calame KL. C/EBP proteins activate transcription from the human immunodeficiency virus type 1 long terminal repeat in macrophages/monocytes. J Virol. 1995;69:5337–5344. doi: 10.1128/jvi.69.9.5337-5344.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ruocco MR, Chen X, Ambrosino C, Dragonetti E, Liu W, Mallardo M, et al. Regulation of HIV-1 long terminal repeats by interaction of C/EBP(NF-IL6) and NF-kappaB/Rel transcription factors. J Biol Chem. 1996;271:22479–22486. doi: 10.1074/jbc.271.37.22479. [DOI] [PubMed] [Google Scholar]
- 27.Mukerjee R, Sawaya BE, Khalili K, Amini S. Association of p65 and C/EBPbeta with HIV-1 LTR modulates transcription of the viral promoter. J Cell Biochem. 2007;100:1210–1216. doi: 10.1002/jcb.21109. [DOI] [PubMed] [Google Scholar]
- 28.Krebs FC, Goodenow MM, Wigdahl B. Neuroglial ATF/CREB factors interact with the human immunodeficiency virus type 1 long terminal repeat. J Neurovirol. 1997;3 Suppl 1:S28–S32. [PubMed] [Google Scholar]
- 29.Krebs FC, Mehrens D, Pomeroy S, Goodenow MM, Wigdahl B. Human immunodeficiency virus type 1 long terminal repeat quasispecies differ in basal transcription and nuclear factor recruitment in human glial cells and lymphocytes. J Biomed Sci. 1998;5:31–44. doi: 10.1007/BF02253354. [DOI] [PubMed] [Google Scholar]
- 30.Ross HL, Nonnemacher MR, Hogan TH, Quiterio SJ, Henderson A, McAllister JJ, et al. Interaction between CCAAT/enhancer binding protein and cyclic AMP response element binding protein 1 regulates human immunodeficiency virus type 1 transcription in cells of the monocyte/macrophage lineage. J Virol. 2001;75:1842–1856. doi: 10.1128/JVI.75.4.1842-1856.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hogan TH, Nonnemacher MR, Krebs FC, Henderson A, Wigdahl B. HIV-1 Vpr binding to HIV-1 LTR C/EBP cis-acting elements and adjacent regions is sequence-specific. Biomed Pharmacother. 2003;57:41–48. doi: 10.1016/s0753-3322(02)00333-5. [DOI] [PubMed] [Google Scholar]
- 32.Abraham S, Sweet T, Sawaya BE, Rappaport J, Khalili K, Amini S. Cooperative interaction of C/EBP beta and Tat modulates MCP-1 gene transcription in astrocytes. J Neuroimmunol. 2005;160:219–227. doi: 10.1016/j.jneuroim.2004.11.009. [DOI] [PubMed] [Google Scholar]
- 33.Ambrosino C, Ruocco MR, Chen X, Mallardo M, Baudi F, Trematerra S, et al. HIV-1 Tat induces the expression of the interleukin-6 (IL6) gene by binding to the IL6 leader RNA and by interacting with CAAT enhancer-binding protein beta (NF-IL6) transcription factors. J Biol Chem. 1997;272:14883–14892. doi: 10.1074/jbc.272.23.14883. [DOI] [PubMed] [Google Scholar]
- 34.Mink S, Haenig B, Klempnauer KH. Interaction and functional collaboration of p300 and C/EBPbeta. Mol Cell Biol. 1997;17:6609–6617. doi: 10.1128/mcb.17.11.6609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kowenz-Leutz E, Leutz A. A C/EBP beta isoform recruits the SWI/SNF complex to activate myeloid genes. Mol Cell. 1999;4:735–743. doi: 10.1016/s1097-2765(00)80384-6. [DOI] [PubMed] [Google Scholar]
- 36.Eaton EM, Hanlon M, Bundy L, Sealy L. Characterization of C/EBPbeta isoforms in normal versus neoplastic mammary epithelial cells. J Cell Physiol. 2001;189:91–105. doi: 10.1002/jcp.1139. [DOI] [PubMed] [Google Scholar]
- 37.Clapham PR, McKnight A. Cell surface receptors, virus entry and tropism of primate lentiviruses. J Gen Virol. 2002;83:1809–1829. doi: 10.1099/0022-1317-83-8-1809. [DOI] [PubMed] [Google Scholar]
- 38.Alkhatib G, Combadiere C, Broder CC, Feng Y, Kennedy PE, Murphy PM, et al. CC CKR5: a RANTES, MIP-1alpha, MIP-1beta receptor as a fusion cofactor for macrophage-tropic HIV-1. Science. 1996;272:1955–1958. doi: 10.1126/science.272.5270.1955. [DOI] [PubMed] [Google Scholar]
- 39.Feng Y, Broder CC, Kennedy PE, Berger EA. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science. 1996;272:872–877. doi: 10.1126/science.272.5263.872. [DOI] [PubMed] [Google Scholar]
- 40.Zhang L, Huang Y, He T, Cao Y, Ho DD. HIV-1 subtype and second-receptor use. Nature. 1996;383:768. doi: 10.1038/383768a0. [DOI] [PubMed] [Google Scholar]
- 41.Tscherning C, Alaeus A, Fredriksson R, Bjorndal A, Deng H, Littman DR, et al. Differences in chemokine coreceptor usage between genetic subtypes of HIV-1. Virology. 1998;241:181–188. doi: 10.1006/viro.1997.8980. [DOI] [PubMed] [Google Scholar]
- 42.Ping LH, Nelson JA, Hoffman IF, Schock J, Lamers SL, Goodman M, et al. Characterization of V3 sequence heterogeneity in subtype C human immunodeficiency virus type 1 isolates from Malawi: underrepresentation of X4 variants. J Virol. 1999;73:6271–6281. doi: 10.1128/jvi.73.8.6271-6281.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Bjorndal A, Sonnerborg A, Tscherning C, Albert J, Fenyo EM. Phenotypic characteristics of human immunodeficiency virus type 1 subtype C isolates of Ethiopian AIDS patients. AIDS Res Hum Retroviruses. 1999;15:647–653. doi: 10.1089/088922299310944. [DOI] [PubMed] [Google Scholar]
- 44.Brigati C, Giacca M, Noonan DM, Albini A. HIV Tat, its TARgets and the control of viral gene expression. FEMS Microbiol Lett. 2003;220:57–65. doi: 10.1016/S0378-1097(03)00067-3. [DOI] [PubMed] [Google Scholar]
- 45.Huigen MC, Kamp W, Nottet HS. Multiple effects of HIV-1 trans-activator protein on the pathogenesis of HIV-1 infection. Eur J Clin Invest. 2004;34:57–66. doi: 10.1111/j.1365-2362.2004.01282.x. [DOI] [PubMed] [Google Scholar]
- 46.Jeang KT, Xiao H, Rich EA. Multifaceted activities of the HIV-1 transactivator of transcription, Tat. J Biol Chem. 1999;274:28837–28840. doi: 10.1074/jbc.274.41.28837. [DOI] [PubMed] [Google Scholar]
- 47.Taube R, Fujinaga K, Wimmer J, Barboric M, Peterlin BM. Tat transactivation: a model for the regulation of eukaryotic transcriptional elongation. Virology. 1999;264:245–253. doi: 10.1006/viro.1999.9944. [DOI] [PubMed] [Google Scholar]
- 48.Berkhout B, Silverman RH, Jeang KT. Tat trans-activates the human immunodeficiency virus through a nascent RNA target. Cell. 1989;59:273–282. doi: 10.1016/0092-8674(89)90289-4. [DOI] [PubMed] [Google Scholar]
- 49.Churcher MJ, Lamont C, Hamy F, Dingwall C, Green SM, Lowe AD, et al. High affinity binding of TAR RNA by the human immunodeficiency virus type-1 tat protein requires base-pairs in the RNA stem and amino acid residues flanking the basic region. J Mol Biol. 1993;230:90–110. doi: 10.1006/jmbi.1993.1128. [DOI] [PubMed] [Google Scholar]
- 50.Desfosses Y, Solis M, Sun Q, Grandvaux N, Van Lint C, Burny A, et al. Regulation of human immunodeficiency virus type 1 gene expression by clade-specific Tat proteins. J Virol. 2005;79:9180–9191. doi: 10.1128/JVI.79.14.9180-9191.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ranga U, Shankarappa R, Siddappa NB, Ramakrishna L, Nagendran R, Mahalingam M, et al. Tat protein of human immunodeficiency virus type 1 subtype C strains is a defective chemokine. J Virol. 2004;78:2586–2590. doi: 10.1128/JVI.78.5.2586-2590.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Campbell GR, Watkins JD, Singh KK, Loret EP, Spector SA. Human immunodeficiency virus type 1 subtype C Tat fails to induce intracellular calcium flux and induces reduced tumor necrosis factor production from monocytes. J Virol. 2007;81:5919–5928. doi: 10.1128/JVI.01938-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Grant C, Nonnemacher M, Jain P, Pandya D, Irish B, Williams SC, et al. CCAAT/enhancer-binding proteins modulate human T cell leukemia virus type 1 long terminal repeat activation. Virology. 2006;348:354–369. doi: 10.1016/j.virol.2005.12.024. [DOI] [PubMed] [Google Scholar]
- 54.Alexaki A, Quiterio SJ, Liu Y, Irish B, Kilareski E, Nonnemacher MR, et al. PMA-induced differentiation of a bone marrow progenitor cell line activates HIV-1 LTR-driven transcription. DNA Cell Biol. 2007;26:387–394. doi: 10.1089/dna.2006.0542. [DOI] [PubMed] [Google Scholar]
- 55.Michael NL, D'Arcy L, Ehrenberg PK, Redfield RR. Naturally occurring genotypes of the human immunodeficiency virus type 1 long terminal repeat display a wide range of basal and Tat-induced transcriptional activities. J Virol. 1994;68:3163–3174. doi: 10.1128/jvi.68.5.3163-3174.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kirchhoff F, Greenough TC, Hamacher M, Sullivan JL, Desrosiers RC. Activity of human immunodeficiency virus type 1 promoter/TAR regions and tat1 genes derived from individuals with different rates of disease progression. Virology. 1997;232:319–331. doi: 10.1006/viro.1997.8586. [DOI] [PubMed] [Google Scholar]
- 57.Estable MC, Bell B, Merzouki A, Montaner JS, O'Shaughnessy MV, Sadowski IJ. Human immunodeficiency virus type 1 long terminal repeat variants from 42 patients representing all stages of infection display a wide range of sequence polymorphism and transcription activity. J Virol. 1996;70:4053–4062. doi: 10.1128/jvi.70.6.4053-4062.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Siddappa NB, Kashi VP, Venkatramanan M, Balasiddaiah A, Jayasuryan N, Mahadevan A, et al. Gene expression analysis from human immunodeficiency virus type 1 subtype C promoter and construction of bicistronic reporter vectors. AIDS Res Hum Retroviruses. 2007;23:1268–1278. doi: 10.1089/aid.2006.0305. [DOI] [PubMed] [Google Scholar]
- 59.Naghavi MH, Schwartz S, Sonnerborg A, Vahlne A. Long terminal repeat promoter/enhancer activity of different subtypes of HIV type 1. AIDS Res Hum Retroviruses. 1999;15:1293–1303. doi: 10.1089/088922299310197. [DOI] [PubMed] [Google Scholar]
- 60.Spira S, Wainberg MA, Loemba H, Turner D, Brenner BG. Impact of clade diversity on HIV-1 virulence, antiretroviral drug sensitivity and drug resistance. J Antimicrob Chemother. 2003;51:229–240. doi: 10.1093/jac/dkg079. [DOI] [PubMed] [Google Scholar]
- 61.Soares EA, Santos AF, Sousa TM, Sprinz E, Martinez AM, Silveira J, et al. Differential drug resistance acquisition in HIV-1 of subtypes B and C. PLoS ONE. 2007;2:e730. doi: 10.1371/journal.pone.0000730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kanki PJ, Hamel DJ, Sankale JL, Hsieh C, Thior I, Barin F, et al. Human immunodeficiency virus type 1 subtypes differ in disease progression. J Infect Dis. 1999;179:68–73. doi: 10.1086/314557. [DOI] [PubMed] [Google Scholar]
- 63.Ramirez de Arellano E, Martin C, Soriano V, Alcami J, Holguin A. Genetic analysis of the long terminal repeat (LTR) promoter region in HIV-1-infected individuals with different rates of disease progression. Virus Genes. 2007;34:111–116. doi: 10.1007/s11262-006-0054-z. [DOI] [PubMed] [Google Scholar]
- 64.Vasan A, Renjifo B, Hertzmark E, Chaplin B, Msamanga G, Essex M, et al. Different rates of disease progression of HIV type 1 infection in Tanzania based on infecting subtype. Clin Infect Dis. 2006;42:843–852. doi: 10.1086/499952. [DOI] [PubMed] [Google Scholar]
- 65.Nonnemacher MR, Irish BP, Liu Y, Mauger D, Wigdahl B. Specific sequence configurations of HIV-1 LTR G/C box array result in altered recruitment of Sp isoforms and correlate with disease progression. J Neuroimmunol. 2004;157:39–47. doi: 10.1016/j.jneuroim.2004.08.021. [DOI] [PubMed] [Google Scholar]
- 66.Scriba TJ, de Villiers T, Treurnicht FK, zur Megede J, Barnett SW, Engelbrecht S, et al. Characterization of the South African HIV type 1 subtype C complete 5' long terminal repeat, nef, and regulatory genes. AIDS Res Hum Retroviruses. 2002;18:149–159. doi: 10.1089/08892220252779692. [DOI] [PubMed] [Google Scholar]
- 67.Siddappa NB, Venkatramanan M, Venkatesh P, Janki MV, Jayasuryan N, Desai A, et al. Transactivation and signaling functions of Tat are not correlated: biological and immunological characterization of HIV-1 subtype-C Tat protein. Retrovirology. 2006;3:53. doi: 10.1186/1742-4690-3-53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Conant K, Garzino-Demo A, Nath A, McArthur JC, Halliday W, Power C, et al. Induction of monocyte chemoattractant protein-1 in HIV-1 Tat-stimulated astrocytes and elevation in AIDS dementia. Proc Natl Acad Sci U S A. 1998;95:3117–3121. doi: 10.1073/pnas.95.6.3117. [DOI] [PMC free article] [PubMed] [Google Scholar]